Introduction
Distribution of white adipose tissues in humans is
associated with metabolic disorders. Individuals who are
peripherally obese (fat accumulation predominantly in the
gluteofemoral region) are at little or no risk of developing
metabolic disease, whereas individuals who are centrally obese (fat
distribution predominantly in visceral depots) are prone to
developing metabolic complications (1). However, the underlying mechanisms that
regulate fat distribution and link excess visceral fat to metabolic
complications are yet to be elucidated.
Environmental obesogens are chemical compounds that
promote or exacerbate the development of obesity and its associated
health outcomes (2) by disrupting or
interfering with critical pathways associated with energy balance,
adipogenesis and lipid metabolism (3). Phthalates are a class of candidate
obesogens that are ingested in food. Phthalate metabolites have
been detected in >80% of the population and fetal exposure
levels are readily detectable (4–6). In a
previous cross-sectional study, urinary phthalate metabolite
concentrations were demonstrated to be associated with an increased
waist circumference and insulin resistance in adult males in the
USA (7,8). This correlation indicates that
increased phthalate exposure may be associated with increased
abdominal obesity and fat distribution. In addition, there are age
and sex differences in the association between phthalate exposure
and obesity (9,10). For example, male children and
adolescents are at high risk for obesity associated with urinary
low molecular weight phthalate metabolites, whereas adults are at
high risk for obesity associated with high molecular weight
phthalate metabolites (9). Moreover,
associations were positive for low molecular weight phthalate
metabolites with BMI z-score in boys >10 years of age, but no
association was detected in girls <10 years of age (10).
Di-(2-ethylhexyl) phthalate (DEHP), which is a
phthalate ester, is predominately used as an industrial plasticizer
and is found in cosmetics, industrial paints and solvents (11). The presence of DEHP metabolites in
urine is associated with adiposity and insulin resistance in
children (12). Previous studies
have reported that perinatal exposure to DEHP may induce obesity
and metabolic disorders in mice (13,14);
however, the mechanisms underlying these associations are yet to be
investigated.
T-box 15 (Tbx15) and glypican 4 (Gpc4)
are developmental genes that have roles in the origins of obesity
and body fat distribution in mice and humans (15). In humans, Tbx15 expression is
negatively correlated with waist/hip ratio (16). Low levels of Tbx15 mRNA and
high levels of Gpc4 mRNA in visceral adipose tissue, and low
levels of Gpc4 mRNA and high levels of Tbx15 mRNA in
subcutaneous adipose tissue appear to be associated with a high
waist/hip ratio (15) and,
therefore, may be associated with obesity and fat distribution.
These genes may be correlated with an increased risk of developing
metabolic and cardiovascular complications (15).
In utero exposure to endocrine-disrupting
chemicals may alter adipose tissue development by affecting the
number, size, and distribution of adipocytes formed, as well as
larger regulatory systems associated with body weight homeostasis
(17). Therefore, it was
hypothesized in the present study that in utero exposure to
DEHP may, in part, affect obesity and fat distribution by altering
the expression of Tbx15 and Gpc4 in murine offspring.
The effects of exposure to DEHP in pregnant dams on their
9-week-old offspring was investigated by measuring: i) Serum
leptin, insulin, lipid, and glucose concentrations; ii) body weight
and adipose tissue deposition; and iii) mRNA expression levels of
Tbx15 and Gpc4 in subcutaneous and visceral adipose
tissues.
Materials and methods
Reagents
DEHP, serum triglyceride determination kit (TR0100)
and cholesterol quantitation kit (MAK043) were purchased from
Sigma-Aldrich (Merck Millipore, Darmstadt, Germany). Leptin ELISA
kit (EK0438) was obtained from Boster Biological Technology (Wuhan,
China), and insulin ELISA kit (EZRMI-13K) was purchased from Merck
Millipore (Shanghai, China). RNAiso Plus, PrimeScript RT Perfect
Real Time reagent kit and SYBR Premix Ex Taq II kit (Tli
RNaseH Plus) were purchased from Takara Biotechnology Co., Ltd.
(Dalian, China). Primers were synthesized by Sangon Biotech Co.,
Ltd. (Shanghai, China).
Animals
Male (n=25) and female (n=50) C57BL/6J mice (age, 8
weeks; weight: male, 21.9±1.0 g; female, 19.6±0.9 g) were purchased
from the Animal Center of China Medical University (Shenyang,
China). Mice were kept at a controlled temperature of 20±2°C and a
relative humidity of 50±10% with a 12-h light/dark cycle. Mice were
provided water and a standard chow diet containing 10% kcal from
fat ad libitum. Animal procedures were conducted according
to an animal protocol approved by the Institutional Animal Care and
Use Committee of China Medical University.
Fetal exposure to DEHP
A total of 25 male and 50 female mice (gender ratio,
1:2) were mated overnight. Females were examined for a copulation
plug the following day, which was designated as gestational day
(GD) 1. Pregnant females were randomly divided into three groups
(F0 control, F0 DEHP0.05 and F0 DEHP500) and exposed to various
concentrations of DEHP (0, 0.05 and 500 mg/kg/day). DEHP (diluted
in olive oil) or vehicle (olive oil) was administered via gavage
every 24 h from GD1 to GD19. Food intake of F0 dams was recorded
weekly. Spontaneous abortion was determined in dams that did not
deliver. Pups (F1 control, n=29 (female/male ratio, 15:14) and F1
DEHP0.05, n=30 (female/male ratio, 14:16) were maintained with
their dams and weaned at three weeks of age. Pups were subsequently
fed a standard diet until nine weeks of age.
Serum measurements
Blood samples of F1 offspring were harvested from
the abdominal aorta at nine weeks of age. Samples were centrifuged
at 1,000 × g for 15 min at room temperature and stored at
−20°C prior to analysis. Levels of serum leptin and insulin were
determined using ELISA kits. Serum triglyceride and total
cholesterol levels were determined using commercial kits. Serum
glucose concentration was measured via the glucose oxidase method
(18). All experimental protocols
were performed according to the manufacturers' protocols. All data
were obtained from three independent experiments.
Adipose tissue collection
F1 offspring were anesthetized with ether (Tianjin
Guangfu Fine Chemical Research Institute, Tianjin, China) and
sacrificed at nine weeks of age via cervical dislocation. Gonadal
and inguinal fat pads were immediately removed, weighed, frozen in
liquid nitrogen and stored at −80°C prior to RT-qPCR analysis.
Gonadal and inguinal fat pads were representative of visceral and
subcutaneous fat, respectively.
RT-qPCR
Total RNA of gonadal and inguinal fat pads was
extracted using RNAiso Plus, according to the manufacturer's
protocol. RT-qPCR reactions were performed using the ABI Prism 7500
system (Thermo Fisher Scientific, Inc., Waltham, MA, USA) as
described previously (19). Briefly,
RT was performed with a reaction mixture containing 2 µl 5X
PrimeScript buffer, 0.5 µl PrimeScript RT Enzyme Mix, 0.5 µl Oligo
dT Primer (50 µM) and/or 0.5 µl Random hexamers (100 µM) and 500 ng
mRNA. The mixture was maintained for 5 min at 37°C followed by 5
sec at 85°C and final hold at 4°C. Subsequently, each qPCR reaction
mixture contained 10 µl 2X SYBR Premix Ex Taq II, 0.8 µl
forward and reverse primers (10 µmol/µl), 0.4 ml Rox Reference Dye
II (50X) and 2 µl cDNA. Thermal cycling was performed to amplify
the respective targets in a total volume of 20 µl: Initial
denaturation at 95°C for 30 sec for 1 cycle, followed by
denaturation at 95°C for 5 sec and annealing and extension 60°C for
34 sec for 40 cycles. β-actin was used as the endogenous control
gene. RT-qPCR data were analyzed using the 2−ΔΔCq method
(20). All PCR reactions were
performed in triplicate. Primers were designed as follows:
β-actin, forward 5′-CATCCGTAAAGACCTCTATGCCAAC-3′ and reverse
5′-ATGGAGCCACCGATCCACA-3′; Gpc4, forward
5′-AGAGCAACGCCCAACCAC-3′ and reverse 5′-GCCATTCCAGCAGTCATC-3′; and
Tbx15, forward 5′-AGCTTCTGGAGACACCTGGATGA-3′ and reverse
5′-CGTGGACTCGAGGCTGGTATTTA-3′.
Statistical analysis
Data are expressed as mean ± standard deviation.
Statistical analyses were performed using the two-tailed unpaired
Student's t-test with unequal variance or one-way analysis of
variance and Spearman's correlation analysis using SPSS 13.0
software (SPSS Inc., Chicago, IL, USA). P<0.05 was considered to
indicate a statistically significant difference.
Results
Effects of exposure to DEHP on
reproductive outcomes of F0 dams
As indicated in Table
I, the food intake of pregnant mice was not significantly
altered by DEHP exposure. However, reproductive outcome was
impaired in dams exposed to a high dose (500 mg/kg) of DEHP. A 100%
abortion rate was exhibited by the F0 DEHP500 group, whereas no
adverse reproductive effect was demonstrated by the F0 DEHP0.05
group. Therefore, the offspring of the F0 DEHP0.05 group (F1
DEHP0.05) and of the control mice (F1 control) were used for
subsequent stages of the present study.
 | Table I.Reproductive outcome of F0 dams
treated with DEHP from gestational day 1 to 19. |
Table I.
Reproductive outcome of F0 dams
treated with DEHP from gestational day 1 to 19.
|
| DEHP (mg) |
|---|
|
|
|
|---|
| Criteria | 0 | 0.05 | 500 |
|---|
| Pregnant dams
(n) | 6 | 6 | 7 |
| Food intake
(g/day) | 4.9±0.5 | 5.0±0.8 | 5.1±0.7 |
| Delivery (n) | 6 | 6 | 0 |
| Abortion (%) | 0 | 0 | 100 |
| F1 offspring
(n) | 29 | 30 | 0 |
| F1 offspring
size | 4.83±1.94 | 4.29±1.80 | – |
| F1 females (n) | 15 | 14 | – |
| F1 males (n) | 14 | 16 | – |
Effects of in utero exposure to DEHP
on serum leptin and insulin concentrations in F1 offspring
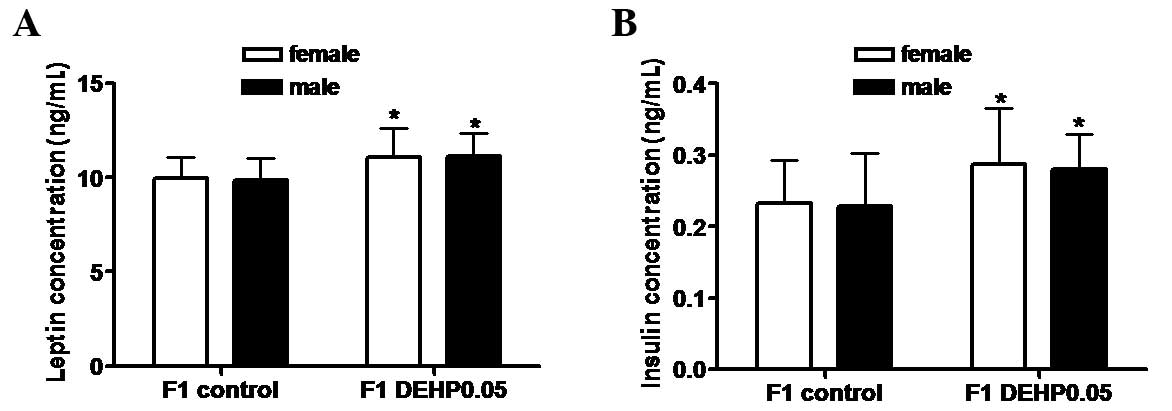
As shown in Fig. 1,
serum leptin (female, 11.09±1.53 ng/ml; male, 11.11±1.23 ng/ml) and
insulin (female, 0.29±0.08 ng/ml; male, 0.28±0.05 ng/ml)
concentrations in the F1 DEHP0.05 group were significantly higher
than those in the F1 control group (leptin: female, 9.95±1.14
ng/ml; male, 9.86±1.19 ng/ml; and insulin: female, 0.23±0.06 ng/ml;
male, 0.23±0.07 ng/ml; P<0.05). No significant differences were
observed between female and male F1 offspring.
Effect of in utero exposure to DEHP on
body weight, fat distribution, serum lipid levels, and glucose
concentrations in F1 offspring
In utero exposure to DEHP had no significant
effect on the body weights (Fig. 2A)
or subcutaneous fat weights (Fig.
2B) of F1 offspring in the DEHP0.05 group compared with the
control group. However, as shown in Fig.
2C, visceral fat weights in the F1 DEHP0.05 group (female,
0.36±0.10 g; male, 0.41±0.10 g) were significantly higher than
those in the F1 control group (female, 0.29±0.05 g; male, 0.30±0.05
g; P<0.05). Consistent with this increase in visceral fat mass,
serum total triglycerides, total cholesterol, and fasting glucose
levels were significantly increased by ~8, 13, and 16%,
respectively, compared with the controls (Fig. 2D-F; P<0.05). No significant
difference was observed between female and male F1 offspring.
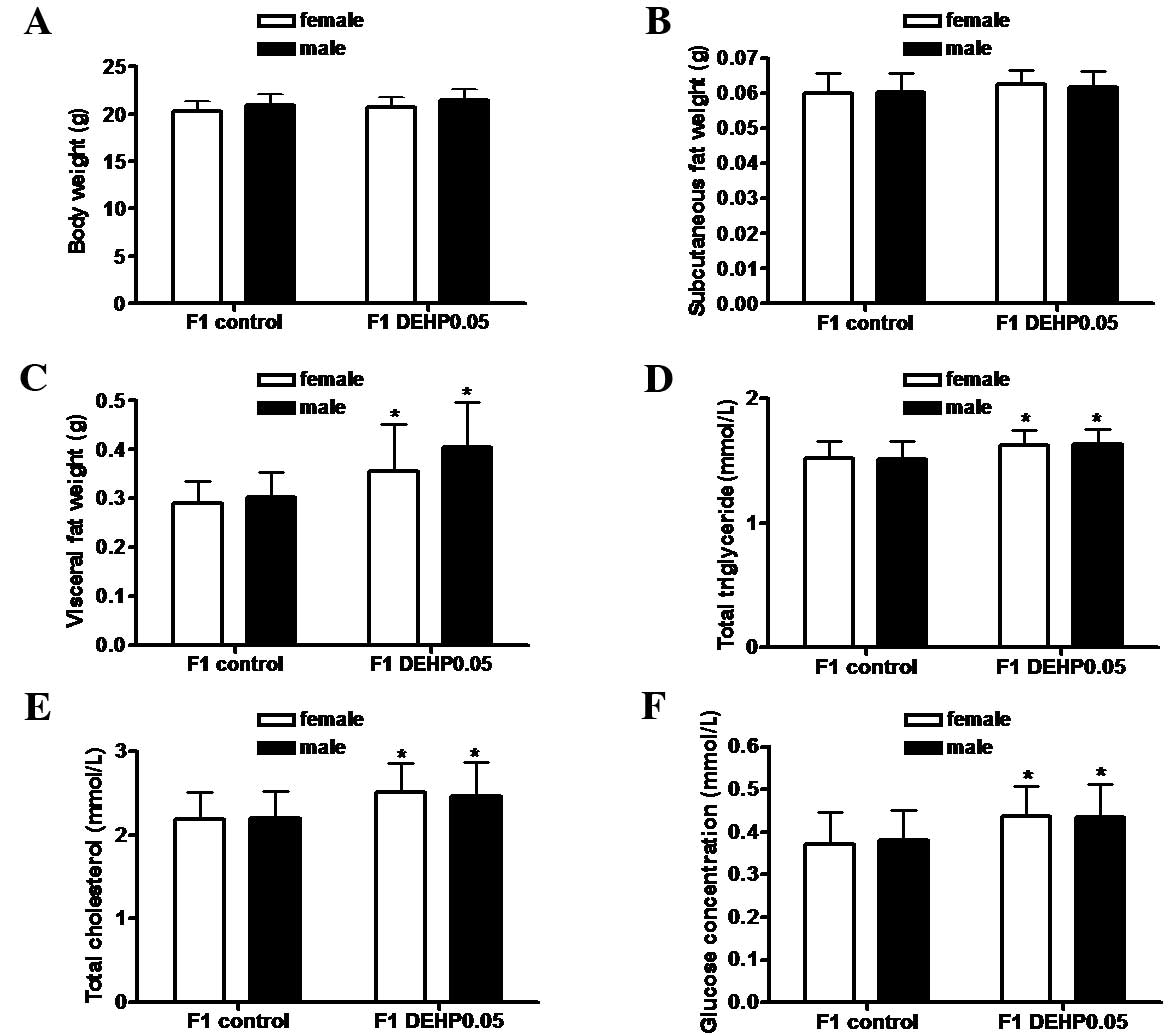 | Figure 2.Effects of in utero exposure
to DEHP on body weight, fat distribution, and serum lipid and
glucose concentration in F1 offspring. (A) Body weight, (B) weight
of subcutaneous fat, (C) weight of visceral fat, (D) total
triglyceride levels, (E) total cholesterol levels, and (F) serum
fasting glucose levels. Results are shown as the mean ± standard
deviation (F1 control group: female, n=15 and male, n=14; F1
DEHP0.05: female, n=14 and male, n=16). DEHP, di-(2-ethylhexyl)
phthalate. *P<0.05 vs. the F1 control group. |
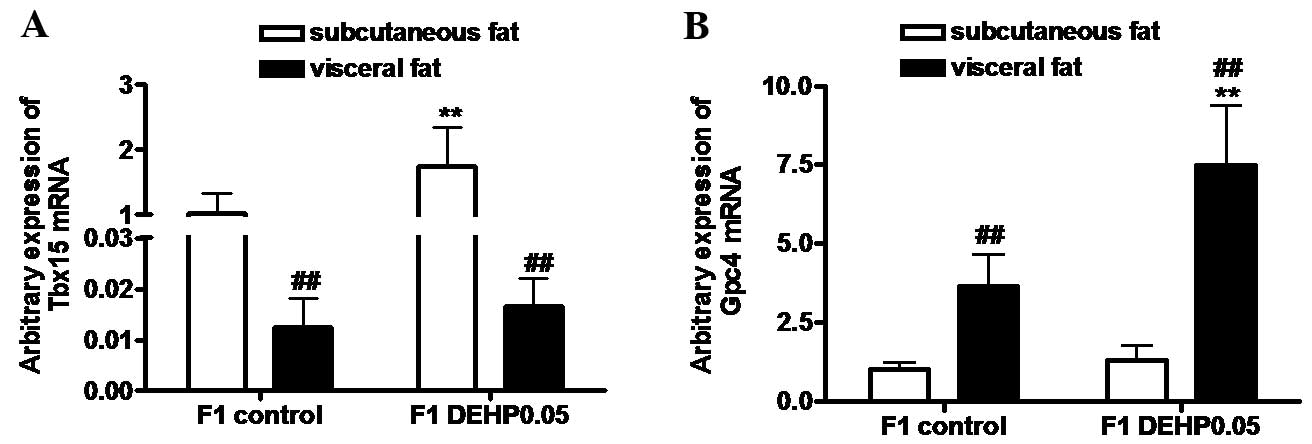
Effects of in utero exposure to DEHP
on the mRNA expression levels of Tbx15 and Gpc4 in subcutaneous and
visceral fat in F1 offspring
As shown in Fig. 3A,
Tbx15 mRNA expression levels in subcutaneous fat in the F1
control and F1 DEHP0.05 groups were significantly higher than those
in visceral fat (P<0.01). The expression of Tbx15 mRNA
expression levels in subcutaneous fat were significantly increased
in the F1 DEHP0.05 group compared with the F1 control group
(P<0.01), whereas no significant increase was observed in
visceral fat. Expression levels of Gpc4 mRNA in subcutaneous fat in
the control and DEHP-treated groups were significantly lower than
those in visceral fat (P<0.01; Fig.
3B). Compared with the F1 control group, the expression of
Gpc4 mRNA in visceral fat was significantly upregulated in
the F1 DEHP0.05 group (P<0.01), but no significant increase was
detected in the subcutaneous fat.
Association between leptin, insulin,
serum lipid and glucose concentration, and mRNA expression levels
of Tbx15 and Gpc4 in subcutaneous and visceral fat of F1
offspring
As shown in Table
II, Spearman's correlation analysis demonstrated that serum
leptin concentration was positively correlated with Tbx15
and Gpc4 mRNA in visceral fat. Serum insulin and glucose
concentrations were positively correlated with Tbx15 mRNA in
subcutaneous fat, as well as Gpc4 mRNA in visceral fat.
Serum total triglyceride was positively correlated with
Tbx15 mRNA in subcutaneous fat, whereas total cholesterol
was positively correlated with Gpc4 mRNA in visceral
fat.
| Table II.Spearman correlation coefficient
analysis between serum measurements and mRNA expression levels in
the fat tissues of F1 offspring. |
Table II.
Spearman correlation coefficient
analysis between serum measurements and mRNA expression levels in
the fat tissues of F1 offspring.
|
| Subcutaneous
fat | Visceral fat |
|---|
|
|
|
|
|---|
| Measurement | Tbx15 | Gpc4 | Tbx15 | Gpc4 |
|---|
| Leptin (ng/ml) | 0.164 | 0.120 | 0.317a | 0.290a |
| Insulin
(ng/ml) | 0.387b | 0.027 | 0.027 | 0.423b |
| Total triglyceride
(mmol/l) | 0.462b | 0.260 | 0.178 | 0.160 |
| Total cholesterol
(mmol/l) | 0.245 | 0.140 | 0.074 | 0.386b |
| Glucose
(mmol/l) | 0.303a | 0.208 | 0.113 | 0.305a |
Discussion
In the present study, DEHP was administered to
pregnant C57BL/6J mice by gavage. DEHP was absorbed through the
intestines, a route that mimics one of the most prominent exposure
routes in humans (11). DEHP was
delivered at 0.05 and 500 mg/kg/day. The lower dose (0.05
mg/kg/day) is considered to be a ‘safe dose’ for humans and is
within the tolerable daily intake (TDI) (21). In contrast, the higher dose (500
mg/kg/day) has caused adverse reproductive and developmental
effects in previous animal studies (22). The principal results of the present
study were as follows. Firstly, exposure to 0.05 mg/kg DEHP did not
significantly affect the food intake or reproductive capacity of F0
dams. Secondly, the weight of visceral fat and serum leptin,
insulin, serum lipid, and glucose concentrations were significantly
elevated in F1 offspring following in utero exposure to 0.05
mg/kg DEHP. Thirdly, expression levels of Tbx15 mRNA in
subcutaneous fat and Gpc4 mRNA in visceral fat were
significantly upregulated in F1 offspring exposed to 0.05 mg/kg
DEHP in utero.
Previous studies have reported that DEHP is able to
induce a dose-dependent decrease in mouse fertility (23), and affect reproductive outcomes in
female mice (24). In the present
study, no significant decline in fertility was exhibited by F0 dams
treated with 0.05 mg/kg DEHP; however, the abortion rate was 100%
in the 500 mg/kg DEHP dose group compared with 0% in the control
and 0.05 mg/kg DEHP groups. These findings indicate that exposure
to DEHP at the TDI level did not affect the reproductive outcomes
of mice; however, a high dose of DEHP may damage the reproductive
capacity. In addition, 0.05 mg/kg DEHP exposure did not
significantly affect the food intake or reproductive outcome of
dams, although it did induce metabolic disorders in the offspring.
Previous studies have reported a correlation between in
utero exposure to endocrine-disrupting chemicals and the
development of metabolic disorders in adulthood (2,14,25,26).
In the present study, serum leptin, insulin, serum lipid, and
glucose concentrations were significantly elevated in male and
female offspring at postnatal week nine, indicating that in
utero exposure to DEHP may influence metabolic function in
adulthood.
Leptin has an important role in the developmental
programming of obesity and insulin resistance (27). In the present study, in utero
DEHP exposure correlated with a significant elevation in leptin
concentration in F1 offspring, which was consistent with the
increased weight of visceral fat. This suggested that in
utero DEHP exposure may cause dysregulation of the central
effect of leptin in F1 offspring. Similarly, a significant increase
in insulin concentration was also demonstrated, consistent with the
increase in glucose levels. This implied the potential development
of insulin resistance. Elevated leptin and insulin levels in adult
offspring may result from hypoleptinemia and hypoinsulinemia during
fetal development due to in utero exposure to DEHP (28). Fetal hypoinsulinemia may lead to
reduced fetal glucose uptake and subsequent insulin resistance
(28). In the present study, leptin
and insulin levels were not detected during the fetal period.
However, a previous study found that fetal rats exposed to
diisobutyl phthalate, another phthalate, exhibited altered insulin
and leptin levels, which implies that perinatal phthalate exposure
may increase the risk of insulin resistance (27). Rajesh and Balasubramanian (29) also demonstrated that gestational
exposure to DEHP may lead to β-cell dysfunction and whole body
glucometabolic abnormalities in F1 offspring.
Fat distribution in F1 offspring in the present
study was also affected by in utero exposure to DEHP. The
body and subcutaneous fat weights of DEHP-exposed F1 offspring were
not significantly increased in male and female mice, whereas the
visceral fat weights were significantly elevated. These results
indicate that excessive visceral fat storage due to in utero
exposure to DEHP may be associated with a high risk of developing
metabolic disorders. The present results agreed in part with two
previous studies, which demonstrated that direct exposure to DEHP,
through placenta and milk, increased body weight and adipose
storage in offspring (13,14). However, in contrast to the effects of
DEHP on body weight and visceral fat tissue in the F1 offspring
observed in the present study, previous studies have detected no
effect (3) or reductions (30) in body weight and white adipose
tissue, suggesting that developmental exposure to DEHP is unlikely
to cause metabolic disorders in adulthood (3). These discrepancies may be due to the
use of different exposure dosages, exposure periods (in
utero/lactation), or strain differences due to gene mutations
or enzyme inactivation accounting for differences in DEHP
metabolism (11). Therefore, more
extensive studies are required to determine whether there are any
adverse effects of DEHP on F1 offspring.
Exposure to certain environmental chemicals, such as
diethylstilbestrol may alter the expression of genes involved in
fat distribution (31).
Overexpression of Tbx15 in 3T3-L1 preadipocytes has been
demonstrated to impair adipocyte differentiation and decrease
triglyceride content (32).
Therefore, differential expression of Tbx15 between fat
depots has a principal role in the interdepot differences in
adipocyte differentiation, triglyceride accumulation and
mitochondrial function, which may contribute to the risk of
developing diabetes and metabolic disease (32). Gpc4 is thought to have a
critical role in the control of cell growth, differentiation, and
morphogenesis (33). This gene is
developmentally regulated in 3T3-F442A adipocytes, and its
expression level may contribute to the regulation of preadipocyte
differentiation (34). Therefore,
the expression of Tbx15 and Gpc4 mRNA in subcutaneous
and visceral fat tissues was detected in the present study to
investigate whether in utero exposure to DEHP may modify
their expression levels in F1 offspring. It was demonstrated that
Tbx15 and Gpc4 mRNA expression levels were
significantly increased in subcutaneous fat and visceral fat
tissues, respectively, of filial mice exposed to DEHP in
utero compared with the control group. DEHP-mediated alteration
of developmental gene levels in subcutaneous and visceral fat
tissues may be one explanation for the excessive visceral fat
weight in the DEHP group.
The transcriptional activity and mRNA expression of
Tbx15 may be regulated by methylation status in the distal
promoter region. Hypomethylation of Tbx15 has previously
been correlated with low birth weight, which is a risk factor for
the development of obesity in adults (35). In the present study, DNA methylation
of Tbx15 was not detected; however, a recent study has
reported that gestational exposure to DEHP at 40 mg/kg may affect
the DNA methylation of imprinting genes in fetal germ cells and
growing oocytes, and in the offspring's oocytes (36). Therefore, it was inferred in the
present study that the differential expression levels of
Tbx15 mRNA in subcutaneous and visceral fat tissues between
the control and DEHP groups may be due to an alteration in
Tbx15 methylation status following in utero DEHP
exposure.
Another developmental gene, Gpc4, may be
regulated by the ratio of specificity protein 3 to specificity
protein 1 (Sp3/Sp1) (34,37). Moreover, there is a physical
interaction between Sp1 and peroxisome proliferator activated
receptor-γ (PPAR-γ) (38,39). The authors of the present study have
previously demonstrated that the Sp3/Sp1 ratio is correlated with
Gpc4 expression levels in the subcutaneous and visceral fat
tissues of mice treated with the PPAR-γ agonist, rosigilitazone,
indicating that the Sp3/Sp1 ratio may regulate Gpc4
expression during the PPAR-γ activation process (37). In the present study, DEHP exposure
was associated with a significant increase in Gpc4 mRNA
expression levels in visceral fat in F1 offspring, which was
consistent with the increase of visceral fat. This suggested that
DEHP exposure may affect fat distribution by regulating Gpc4
mRNA expression. In animals and humans, intestinal lipases convert
DEHP to its monoester equivalent, monoethyl-hexyl-phthalate (MEHP),
which is a selective PPAR-γ modulator (40). MEHP has previously been demonstrated
to induce adipogenesis by modulating PPAR-γ activity (41). In utero exposure to a low dose
of MEHP significantly increased the body and fat pad weights, and
the serum cholesterol, triacylglycerol and glucose levels of male
offspring at postnatal day 60 (42).
These observations suggest that in utero exposure to DEHP
may directly or indirectly regulate the expression levels of Sp1
and Sp3 by activating PPAR-γ, thereby altering the expression of
Gpc4 in visceral and subcutaneous fat tissues.
In conclusion, prenatal exposure to safe,
environmentally relevant concentrations of DEHP caused an increase
in serum leptin and insulin levels in male and female offspring.
This finding indicates that in utero exposure to DEHP may
influence metabolic function later in life. The effects of DEHP
were analyzed in terms of the increased visceral fat weight and
increased expression levels of two developmental genes:
Tbx15 in subcutaneous fat and Gpc4 in visceral fat.
These data suggest that prenatal DEHP exposure may alter fat
distribution by upregulating Tbx15 and Gpc4 in
subcutaneous and visceral adipose tissues, although the exact
mechanisms are yet to be elucidated. The present study stresses the
importance of further investigation into the mechanisms by which
prenatal and postnatal DEHP exposure may affect fat distribution
and lead to metabolic disorders.
Acknowledgements
This work was supported by the Liaoning Educational
Foundation (grant no. L2011138), the Liaoning Natural Scientific
Foundation (grant no. 201102285), and the Key Platform Foundation
of Science and Technology for the Universities in Liaoning
Province, China (grant no. 16010).
References
|
1
|
Kissebah AH and Krakower GR: Regional
adiposity and morbidity. Physiol Rev. 74:761–811. 1994.PubMed/NCBI
|
|
2
|
Grün F and Blumberg B: Perturbed nuclear
receptor signaling by environmental obesogens as emerging factors
in the obesity crisis. Rev Endocr Metab Disord. 8:161–171. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Casals-Casas C, Feige JN and Desvergne B:
Interference of pollutants with PPARs: Endocrine disruption meets
metabolism. Int J Obes (Lond). 32:(Suppl 6). S53–S61. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Wittassek M, Wiesmüller GA, Koch HM,
Eckard R, Dobler L, Müller J, Angerer J and Schlüter C: Internal
phthalate exposure over the last two decades-a retrospective human
biomonitoring study. Int J Hyg Environ Health. 210:319–333. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Wittassek M, Angerer J, Kolossa-Gehring M,
Schäfer SD, Klockenbusch W, Dobler L, Günsel AK, Müller A and
Wiesmüller GA: Fetal exposure to phthalates-a pilot study. Int J
Hyg Environ Health. 212:492–498. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Serrano SE, Braun J, Trasande L, Dills R
and Sathyanarayana S: Phthalates and diet: A review of the food
monitoring and epidemiology data. Environ Health. 13:432014.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Stahlhut RW, van Wijngaarden E, Dye TD,
Cook S and Swan SH: Concentrations of urinary phthalate metabolites
are associated with increased waist circumference and insulin
resistance in adult US males. Environ Health Perspect. 115:876–882.
2007. View
Article : Google Scholar : PubMed/NCBI
|
|
8
|
Hatch EE, Nelson JW, Qureshi MM, Weinberg
J, Moore LL, Singer M and Webster TF: Association of urinary
phthalate metabolite concentrations with body mass index and waist
circumference: A cross-sectional study of NHANES data, 1999–2002.
Environ Health. 7:272008. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Buser MC, Murray HE and Scinicariello F:
Age and sex differences in childhood and adulthood obesity
association with phthalates: Analyses of NHANES 2007–2010. Int J
Hyg Environ Health. 217:687–694. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Zhang Y, Meng X, Chen L, Li D, Zhao L,
Zhao Y, Li L and Shi H: Age and sex-specific relationships between
phthalate exposures and obesity in Chinese children at puberty.
PLoS One. 9:e1048522014. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Feige JN, Gerber A, Casals-Casas C, Yang
Q, Winkler C, Bedu E, Bueno M, Gelman L, Auwerx J, Gonzalez FJ and
Desvergne B: The pollutant diethylhexyl phthalate regulates hepatic
energy metabolism via species-specific PPARalpha-dependent
mechanisms. Environ Health Perspect. 118:234–241. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Smerieri A, Testa C, Lazzeroni P, Nuti F,
Grossi E, Cesari S, Montanini L, Latini G, Bernasconi S, Papini AM
and Street ME: Di-(2-ethylhexyl) phthalate metabolites in urine
show age-related changes and associations with adiposity and
parameters of insulin sensitivity in childhood. PLoS One.
10:e01178312015. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Hao C, Cheng X, Guo J, Xia H and Ma X:
Perinatal exposure to diethyl-hexyl-phthalate induces obesity in
mice. Front Biosci (Elite Ed). 5:725–733. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Schmidt JS, Schaedlich K, Fiandanese N,
Pocar P and Fischer B: Effects of di(2-ethylhexyl) phthalate (DEHP)
on female fertility and adipogenesis in C3H/N mice. Environ Health
Perspect. 120:1123–1129. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Gesta S, Blüher M, Yamamoto Y, Norris AW,
Berndt J, Kralisch S, Boucher J, Lewis C and Kahn CR: Evidence for
a role of developmental genes in the origin of obesity and body fat
distribution. Proc Natl Acad Sci USA. 103:6676–6681. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Yamamoto Y, Gesta S, Lee KY, Tran TT,
Saadatirad P and Kahn CR: Adipose depots possess unique
developmental gene signatures. Obesity (Silver Spring). 18:872–878.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Hatch EE, Nelson JW, Stahlhut RW and
Webster TF: Association of endocrine disruptors and obesity:
Perspectives from epidemiological studies. Int J Androl.
33:324–332. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Lott JA and Turner K: Evaluation of
Trinder's glucose oxidase method for measuring glucose in serum and
urine. Clin Chem. 21:1754–1760. 1975.PubMed/NCBI
|
|
19
|
Liu L, Gu H, Yang J, Ma S, Yu F, Ren Y and
An L: Adipogenic differentiation is not influenced by
lentivirus-mediated shRNA targeting the SOCS3 gene in
adipose-derived stromal cells. Mol Biol Rep. 37:2455–2462. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta Delta C(T)) Method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Naville D, Pinteur C, Vega N, Menade Y,
Vigier M, Le Bourdais A, Labaronne E, Debard C, Luquain-Costaz C,
Bégeot M, et al: Low-dose food contaminants trigger sex-specific,
hepatic metabolic changes in the progeny of obese mice. FASEB J.
27:3860–3870. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Moore RW, Rudy TA, Lin TM, Ko K and
Peterson RE: Abnormalities of sexual development in male rats with
in utero and lactational exposure to the antiandrogenic plasticizer
Di(2-ethylhexyl) phthalate. Environ Health Perspect. 109:229–237.
2001. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Lamb JC IV, Chapin RE, Teague J, Lawton AD
and Reel JR: Reproductive effects of four phthalic acid esters in
the mouse. Toxicol Appl Pharmacol. 88:255–269. 1987. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Niermann S, Rattan S, Brehm E and Flaws
JA: Prenatal exposure to di-(2-ethylhexyl) phthalate (DEHP) affects
reproductive outcomes in female mice. Reprod Toxicol. 53:23–32.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Alonso-Magdalena P, Quesada I and Nadal Á:
Prenatal Exposure to BPA and offspring outcomes: The diabesogenic
behavior of BPA. Dose Response. 13:1559325815590395. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Vaiserman A: Early-life exposure to
endocrine disrupting chemicals and later-life health outcomes: An
epigenetic bridge? Aging Dis. 5:419–429. 2014.PubMed/NCBI
|
|
27
|
Boberg J, Metzdorff S, Wortziger R,
Axelstad M, Brokken L, Vinggaard AM, Dalgaard M and Nellemann C:
Impact of diisobutyl phthalate and other PPAR agonists on
steroidogenesis and plasma insulin and leptin levels in fetal rats.
Toxicology. 250:75–81. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Holemans K, Aerts L and Van Assche FA:
Lifetime consequences of abnormal fetal pancreatic development. J
Physiol. 547:11–20. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Rajesh P and Balasubramanian K:
Gestational exposure to di(2-ethylhexyl) phthalate (DEHP) impairs
pancreatic β-cell function in F1 rat offspring. Toxicol Lett.
232:46–57. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Martinelli MI, Mocchiutti NO and Bernal
CA: Effect of di(2-ethylhexyl) phthalate (DEHP) on lipolysis and
lipoprotein lipase activities in adipose tissue of rats. Hum Exp
Toxicol. 29:739–745. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Newbold RR: Impact of environmental
endocrine disrupting chemicals on the development of obesity.
Hormones (Athens). 9:206–217. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Gesta S, Bezy O, Mori MA, Macotela Y, Lee
KY and Kahn CR: Mesodermal developmental gene Tbx15 impairs
adipocyte differentiation and mitochondrial respiration. Proc Natl
Acad Sci USA. 108:2771–2776. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Tumova S, Woods A and Couchman JR: Heparan
sulfate proteoglycans on the cell surface: Versatile coordinators
of cellular functions. Int J Biochem Cell Biol. 32:269–288. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Li H, Melford K, Judson A and Bensadoun A:
Murine glypican-4 gene structure and expression; Sp1 and Sp3 play a
major role in glypican-4 expression in 3T3-F442A cells. Biochim
Biophys Acta. 1679:141–155. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Chelbi ST, Doridot L, Mondon F, Dussour C,
Rebourcet R, Busato F, Gascoin-Lachambre G, Barbaux S, Rigourd V,
Mignot TM, et al: Combination of promoter hypomethylation and PDX1
overexpression leads to TBX15 decrease in vascular IUGR placentas.
Epigenetics. 6:247–255. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Li L, Zhang T, Qin XS, Ge W, Ma HG, Sun
LL, Hou ZM, Chen H, Chen P, Qin GQ, et al: Exposure to diethylhexyl
phthalate (DEHP) results in a heritable modification of imprint
genes DNA methylation in mouse oocytes. Mol Biol Rep. 41:1227–1235.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Liu L, Gu H, Zhao Y, An L and Yang J:
Glypican 4 may be involved in the adipose tissue redistribution in
high-fat feeding C57BL/6J mice with peroxisome
proliferators-activated receptor γ agonist rosiglitazone treatment.
Exp Ther Med. 8:1813–1818. 2014.PubMed/NCBI
|
|
38
|
Sassa Y, Hata Y, Aiello LP, Taniguchi Y,
Kohno K and Ishibashi T: Bifunctional properties of peroxisome
proliferator-activated receptor gamma1 in KDR gene regulation
mediated via interaction with both Sp1 and Sp3. Diabetes.
53:1222–1229. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Sugawara A, Uruno A, Kudo M, Ikeda Y, Sato
K, Taniyama Y, Ito S and Takeuchi K: Transcription suppression of
thromboxane receptor gene by peroxisome proliferator-activated
receptor-gamma via an interaction with Sp1 in vascular smooth
muscle cells. J Biol Chem. 277:9676–9683. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Huber WW, Grasl-Kraupp B and
Schulte-Hermann R: Hepatocarcinogenic potential of di(2-ethylhexyl)
phthalate in rodents and its implications on human risk. Crit Rev
Toxicol. 26:365–481. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Feige JN, Gelman L, Rossi D, Zoete V,
Métivier R, Tudor C, Anghel SI, Grosdidier A, Lathion C,
Engelborghs Y, et al: The endocrine disruptor
monoethyl-hexyl-phthalate is a selective peroxisome
proliferator-activated receptor gamma modulator that promotes
adipogenesis. J Biol Chem. 282:19152–19166. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Hao C, Cheng X, Xia H and Ma X: The
endocrine disruptor mono-(2-ethylhexyl) phthalate promotes
adipocyte differentiation and induces obesity in mice. Biosci Rep.
32:619–629. 2012. View Article : Google Scholar : PubMed/NCBI
|