Introduction
Sevoflurane is a commonly used general anesthetic,
which has been widely used in the anesthesia of infants and young
children (1). Long-term exposure to
sevoflurane anesthesia can cause neurological dysfunction,
especially in infants, which can lead to developmental
neurodegeneration (1). It may also
cause the central nervous system degradation of infants, affecting
memory and cognitive function (2).
Recent studies have shown that the exposure to clinically relevant
doses of narcotic drugs, such as isoflurane and sevoflurane, leads
to neurological disorders in rats and mice (3,4). All
these results indicate that anesthetic exposure in the early
postpartum period may cause permanent damage to the developing
brain (3).
Hypoxia inducible factor-1 (HIF-1) is an
oxygen-sensitive transcriptional regulator that regulates more than
70 downstream target genes, including vascular endothelial growth
factor, erythropoietin, glucose transporter and so on (5). Upregulation of HIF-1 expression may
play a neuroprotective role in animal models of focal cerebral
ischemia (6).
MAPKs are short for the mitogen-activated protein
kinase family. The family members are a group of evolutionarily
conserved enzymes in mammalian cells, including four subclasses:
ERK1/2, JNK, p38 and ERK5 (7). In
recent years, the MAPK family has been found to be an important
signal-regulating enzyme between cell surface receptors and
determinants of gene expression (8).
Therefore, the MAPK family governs almost all the physiological
functions and processes such as cell adaptation, proliferation,
differentiation, survival, and programmed cell death. And it is the
hub between the cytoplasm and the nucleus (8).
VEGF is purified from bovine pituitary follicular
cells in 1959 and is a vascular endothelial cell-specific mitogen
that promotes the growth of vascular endothelial cells. It lifts
the mRNA levels of plasminogen activator (PA) and plasminogen
activator inhibitor (PAI) to play a role of activation and
regulation on PA and PAI-1, thus enhancing the activity of
plasminogen activator and promoting the formation of new
capillaries. In addition, the addition of VEGF to the culture
medium can increase the survival rate of neurons in the cerebral
cortex as well as the number and length of ganglion cell processes,
to promote nerve growth.
Vitexin is mainly found in the Crataeguspinnatifida,
as one of the main active components of the total flavonoids
extracted from the dried fruits (9).
In addition, they are also widely found in other plants, such as
Trollius, Cajan leaves, willow leaves, short bamboo leaves and
Humulus (9). Vitexin has anti-tumor,
anti-oxidation, anti-virus, anti-inflammatory, anti-bacterial,
antihypertensive and antispasmodic and other biological activities
(10). In recent years, it has been
reported that vitexin has anti-inflammatory effect (10). The present study was designed to
investigate whether the protective effect of vitexin protects
against sevoflurane-induced neuronal apoptosis and the mechanisms
of this protective effect.
Materials and methods
Cell culture and experimental
protocols
Human neuroglioma cells H4 cell was purchased from
Shanghai Cell Bank and cultured in Dulbecco's modified Eagle's
medium (DMEM) (Gibco, Grand Island, NY, USA) containing 10%
heat-inactivated fetal bovine serum (Gibco) at 37°C with 5%
CO2 in a humidified incubator. Cells were gassed with 0%
(Control group), 4.1% sevoflurane (Model) and 4.1% sevoflurane +
100 µM vitexin (Vitexin). In Model group, H4 cell was gassed with
4.1% sevoflurane for 15 min. Then, H4 cell was gassed with 4.1%
sevoflurane for 15 min, and then treated with 100 µM vitexin for 24
h.
Methyl-thiazolyl-tetrazolium (MTT)
assay
After vitexin treatment, H4 cell was added with MTT
(10 µl, 5 mg/ml) for 4 h at 37°C. Medium was removed and DMSO sassy
was added for 20 min at 37°C. The absorbance was measured by a
microplate reader at 490 nm using ELISA reader (Epoch™; BioTek
Instruments, Inc., Winooski, VT, USA).
Sevoflurane exposure
Male SD rats weighing (20 g, 1–2 weeks) and SD rat
were maintained at 22±2°C, 55–5% humidity, at 12 h light/dark
cycle, and ad libitum access to food and water. Thirty rats were
randomly and equally assigned into three groups: control group,
Sevoflurane model group, Sevoflurane model + Vitexin group.
Sevoflurane-induced rat was placed into a chamber and exposed to
2.1% sevoflurane for 2 h. Then, rats of Sevoflurane model + Vitexin
group was treated with 50 mg/kg of Vitexin for 4 weeks.
Assessment of oxidative stress and
caspase-3 markers
Total protein was extracted from cell or tissue
samples using RIPA sassy (Sigma, St. Louis, MO, USA) and total
protein concentration was analyzed with a BCA kit (Sigma). Equal
amounts of the proteins (5 µg) was incubated with malondialdehyde
(MDA) level, and increased superoxide dismutase (SOD) and
glutathione peroxidase (GSH-px) activity ELISA Kits or caspase-3
activity ELISA kit. The absorbance was measured by a microplate
reader at 450 or 405 nm using ELISA reader (Epoch™; BioTek
Instruments, Inc.).
Western blot analysis
Total protein was extracted from cell or hippocampus
tissue samples using RIPA sassy (Sigma) and total protein
concentration was analyzed with a BCA kit (Sigma). Equal amounts of
the proteins (50 µg) were subjected to 6–12% SDS-PAGE and then
transferred to a 0.2 µm polyvinylidene difluoride membrane (EMD
Millipore, Billerica, MA, USA). The membrane was blocked with 5%
non-fat milk in TBST, and immunobloted overnight using primary
antibodies against Bax (Cell Signaling Technology, Inc., Danvers,
MA, USA), HIF-1α (Cell Signaling Technology, Inc.), VEGF (Cell
Signaling Technology, Inc.), phosphorylation-p38 MAPK (p-p38 MAPK,
Cell Signaling Technology, Inc.) and GAPDH (Cell Signaling
Technology, Inc.) overnight at 4°C. Subsequently, the membranes
were washed thrice with TBST and incubated with HRP-linked
anti-mouse IgG (Cell Signaling Technology, Inc.) for 2 h at room
temperature and observed using an enhanced chemiluminescence kit
(GE Healthcare Life Sciences, Chalfont, UK). Image Lab 3.0 software
(Bio-Rad Laboratories, Hercules, CA, USA) was measured Band
intensities.
Statistical analysis
Data from each group were expressed as mean ±
standard error. Data were analyzed using one-way analysis of
variance (ANOVA). P<0.05 was considered to indicate a
statistically significant difference.
Results
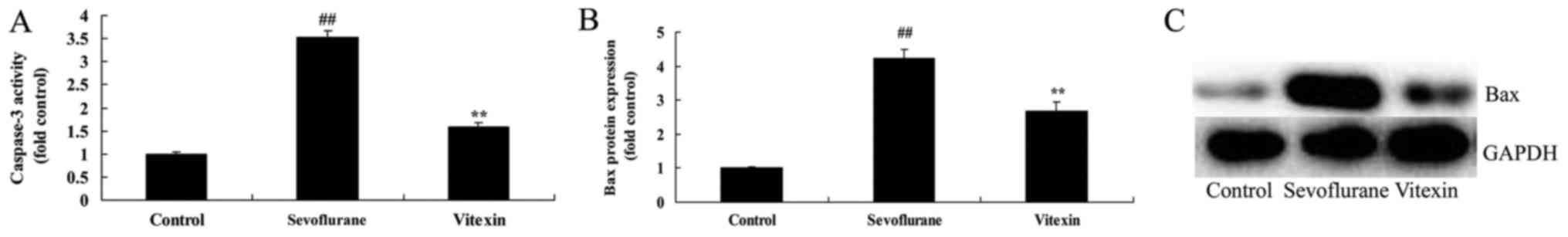
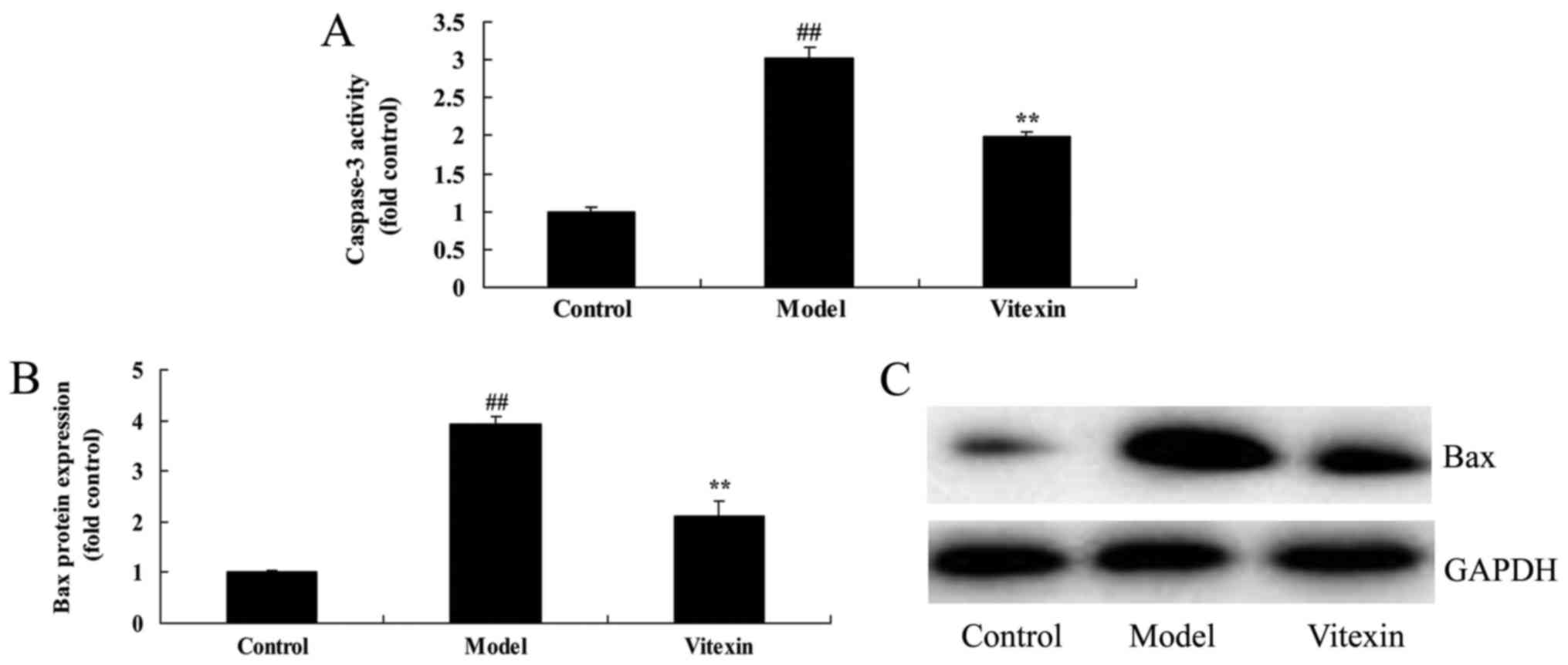
The effects of vitexin on caspase-3
activity and Bax protein expression of sevoflurane-induced newborn
rat
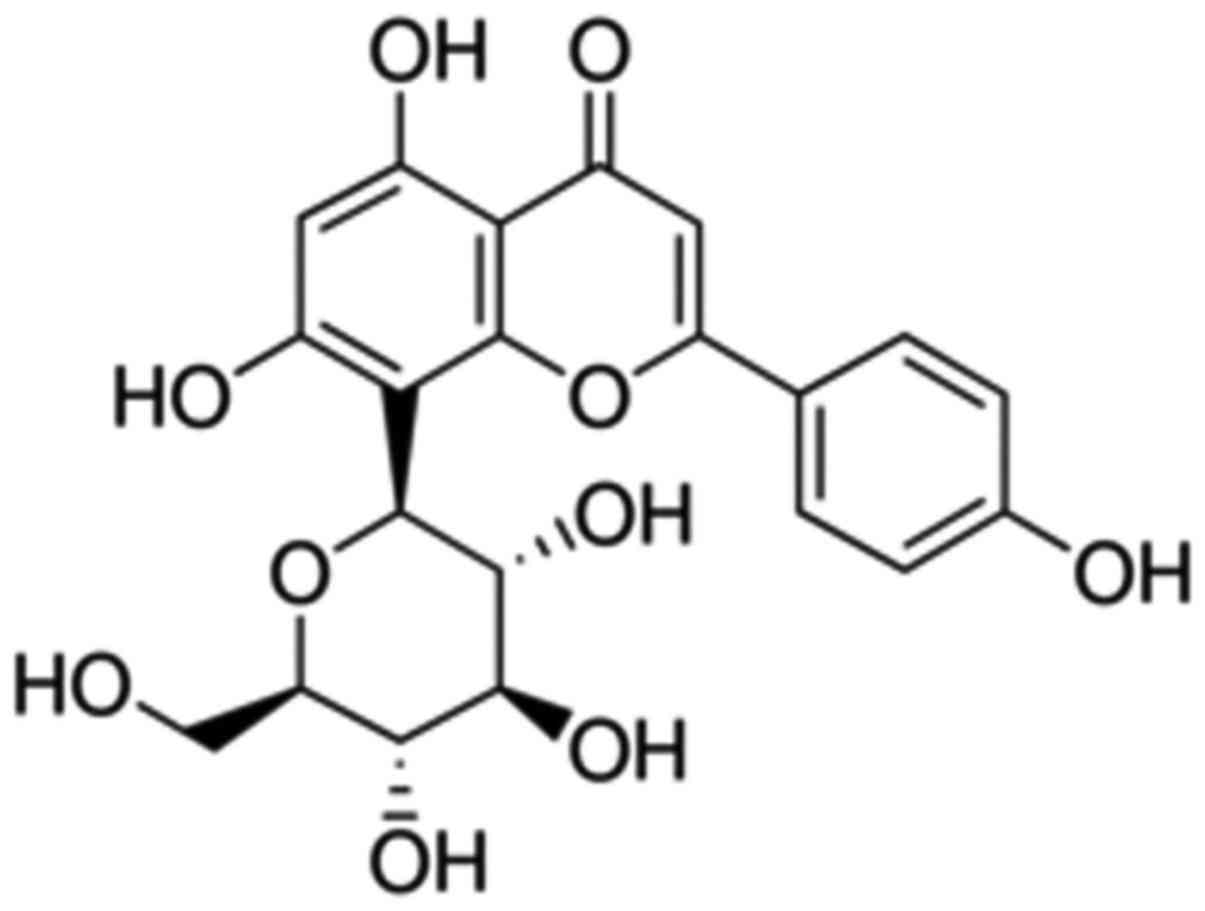
The chemical structure of Vitexin was showed at
Fig. 1. We firstly used
sevoflurane-induced newborn rat treated by vitexin to evaluate the
effect of vitexin on nerve apoptosis of sevoflurane-induced newborn
rat. we analyzed caspase-3 activity and Bax protein expression of
sevoflurane-induced newborn rat. As shown in Fig. 2, there were significant increases of
caspase-3 activity and Bax protein expression in
sevoflurane-induced newborn rat, compared with control group.
Treatment with Vitexin significantly inhibited sevoflurane-induced
caspase-3 activity and Bax protein expression in
sevoflurane-induced newborn rat (Fig.
2).
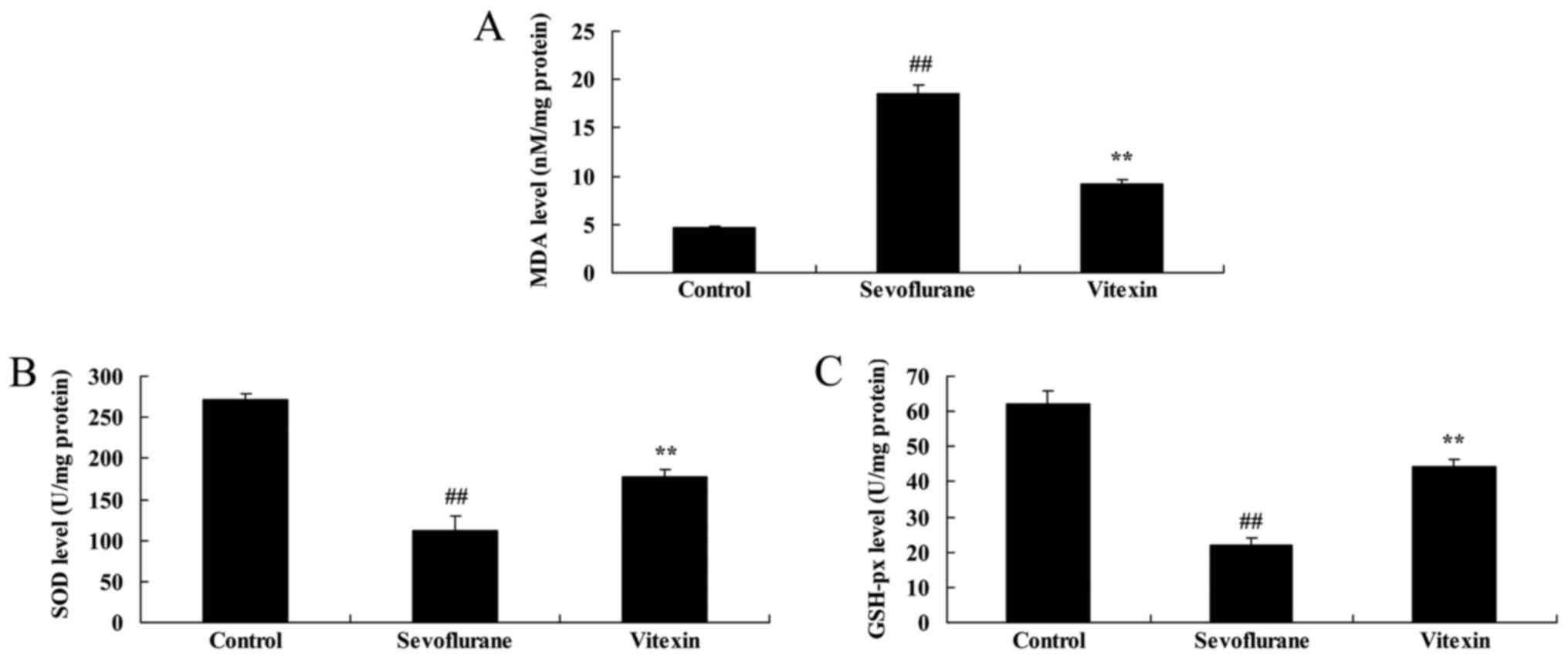
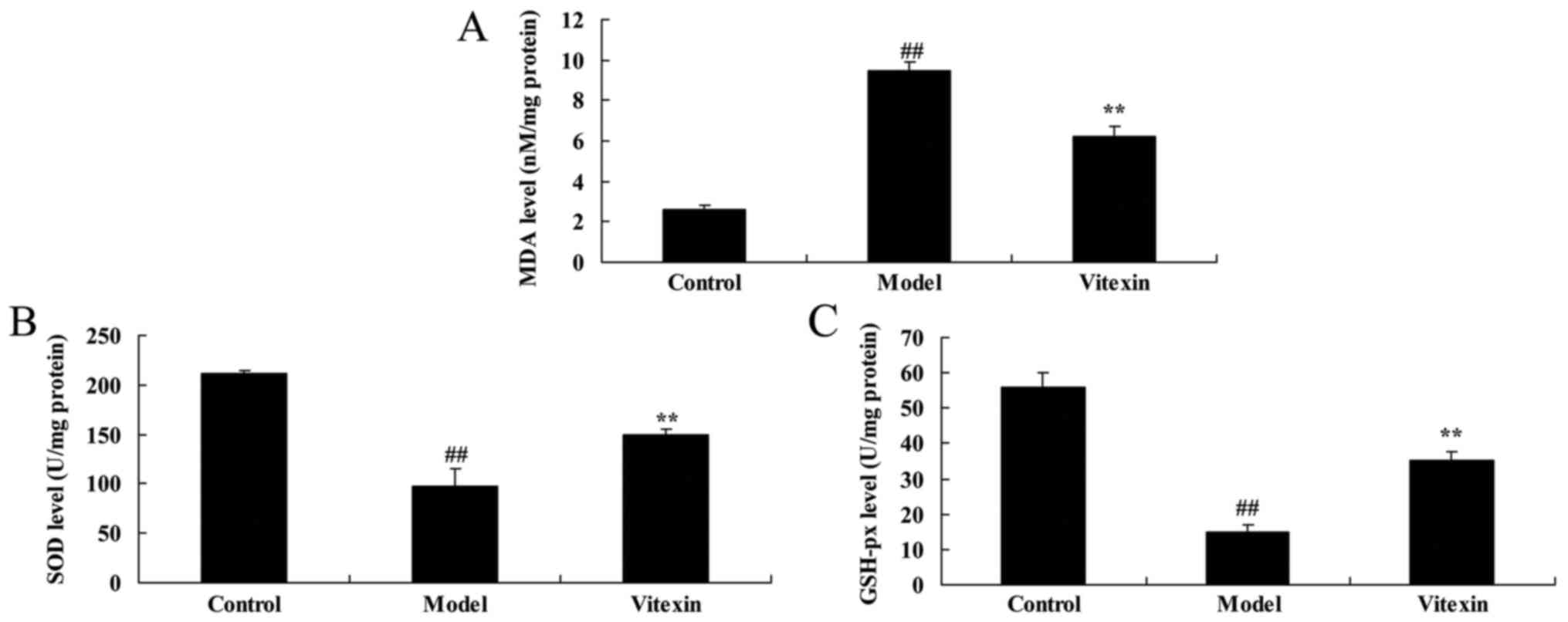
The effects of vitexin on MDA, SOD and
GSH-px activities of sevoflurane-induced newborn rat
Next, we examined the anti-oxidative effects of
Vitexin in sevoflurane-induced newborn rat. Fig. 3 showed that the increase of MDA level
and inhibition of SOD and GSH-px levels in sevoflurane-induced
newborn rat were significantly observed, compared with control
group. Vitexin treatment with significantly inhibited MDA level and
increased SOD and GSH-px levels in sevoflurane-induced newborn rat
(Fig. 3).
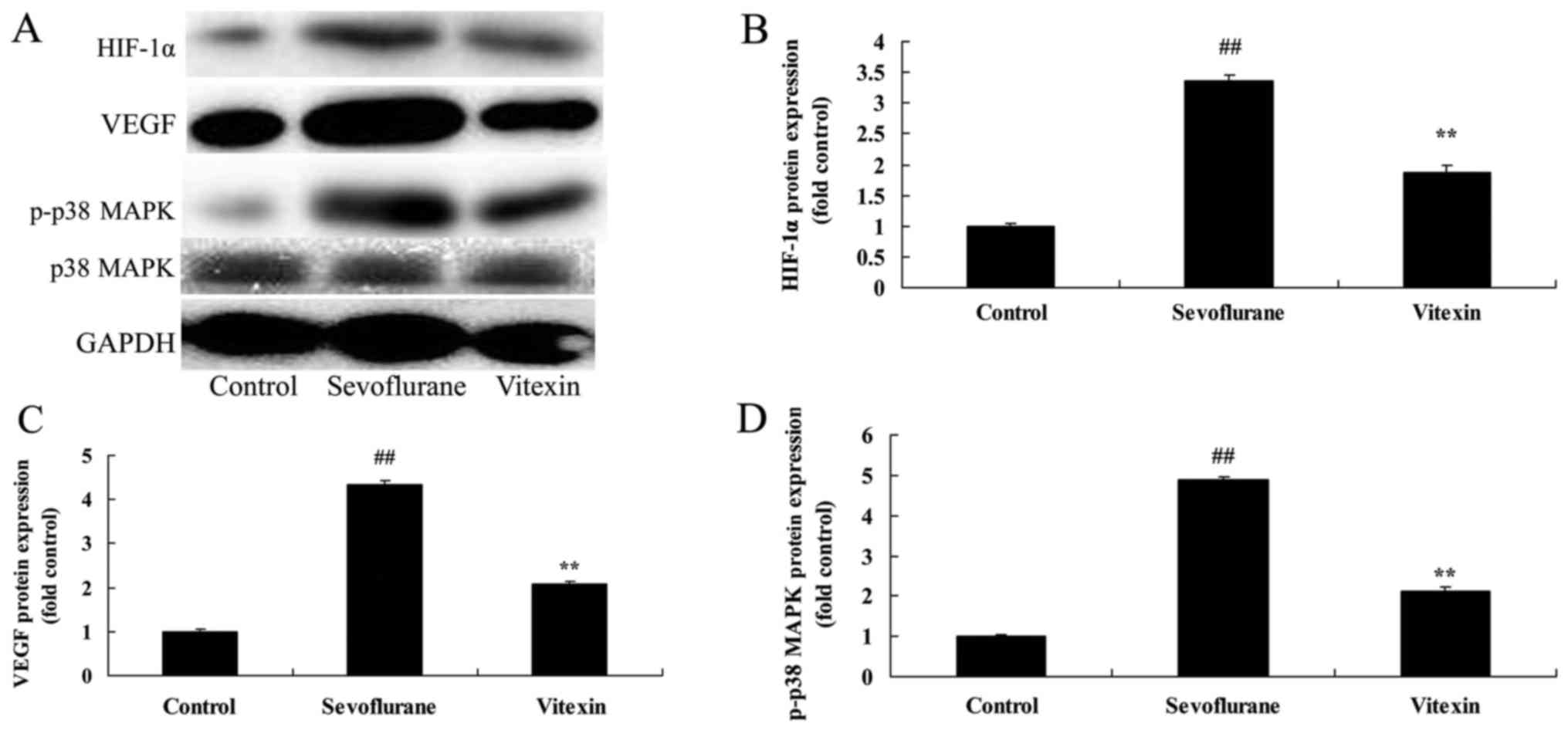
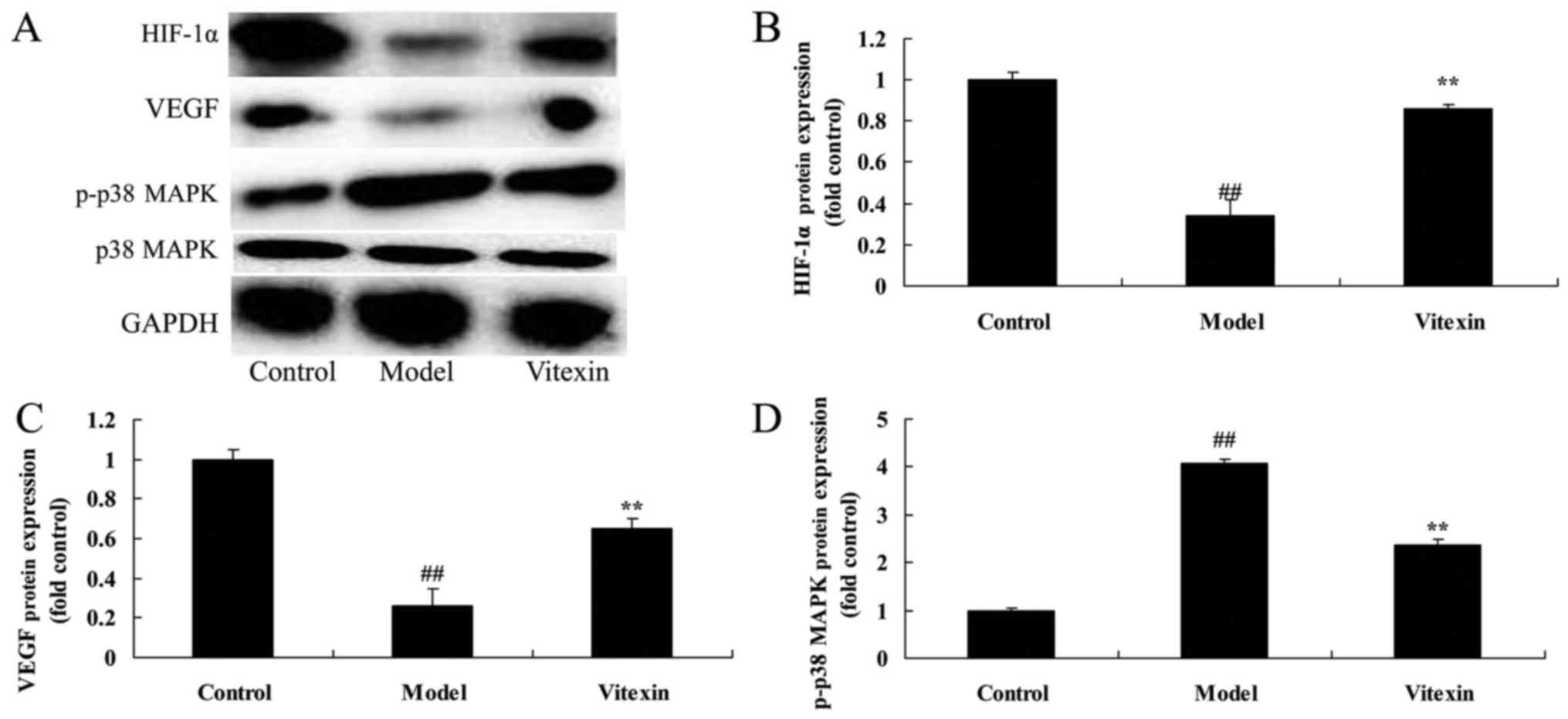
The effects of vitexin on HIF-1α, VEGF
and p38 MAPK protein expression of sevoflurane-induced newborn
rat
Thus, in the current study, we investigated the
possible mechanism of Vitexin on nerve apoptosis of
sevoflurane-induced newborn rat. The protein of HIF-1α, VEGF and
p38 MAPK expression in sevoflurane-induced newborn rat was very
higher than those of control group (Fig.
4). Treatment with Vitexin significantly suppressed HIF-1α,
VEGF and p38 MAPK expression, and promoted VEGF protein expression
in sevoflurane-induced newborn rat (Fig.
4).
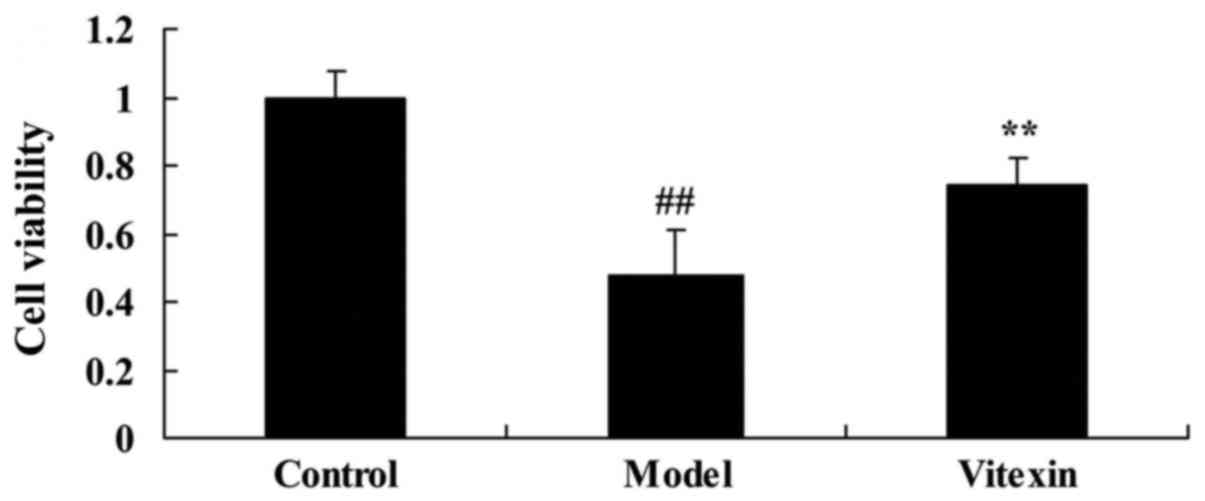
The effects of Vitexin on cell growth
of sevoflurane-induced human neuroglioma cells
Then, we needed to explore the effects of Vitexin on
cell growth of sevoflurane-induced human neuroglioma cells in
vitro model, so H4 cell was gassed with 4.1% sevoflurane for 15
min, and then treated with 100 µM vitexin for 24 h. These results
showed that cell proliferation of sevoflurane-induced human
neuroglioma cells was effectively inhibited, compared with control
group (Fig. 5). Treatment with
Vitexin increased the cell proliferation of sevoflurane-induced
human neuroglioma cells, compared with sevoflurane-induced human
neuroglioma cells group (Fig.
5).
The effects of Vitexin on caspase-3
activity and Bax protein expression of sevoflurane-induced human
neuroglioma cells
We investigated the effects of Vitexin on nerve
apoptosis of sevoflurane-induced human neuroglioma cells. There
were significant increases of caspase-3 activity and Bax protein
expression of sevoflurane-induced human neuroglioma cells, compared
with control group (Fig. 6).
Treatment with Vitexin significantly inhibited caspase-3 activity
and Bax protein expression of sevoflurane-induced human neuroglioma
cells, compared with sevoflurane-induced human neuroglioma cells
group (Fig. 6).
The effects of Vitexin on MDA, SOD and
GSH-px activities of sevoflurane-induced human neuroglioma
cells
To further investigate whether the effects of
Vitexin on oxidative stress of sevoflurane-induced human
neuroglioma cells, MDA, SOD and GSH-px activities were measured
using ELISA KITS. As showed in Fig.
7, the increase of MDA level and inhibition of SOD and GSH-px
levels were effectively in sevoflurane-induced human neuroglioma
cells, compared with control group. Treatment with Vitexin
significantly inhibited MDA level and increased SOD and GSH-px
levels in sevoflurane-induced human neuroglioma cells (Fig. 7).
The effects of Vitexin on HIF-1α, VEGF
and p38 MAPK protein expression of sevoflurane-induced human
neuroglioma cells
We performed western blot analysis to measure
HIF-1α, VEGF and p38 MAPK protein expression in sevoflurane-induced
human neuroglioma cells. As showed in Fig. 8, p38 MAPK protein expression were
effectively increased, HIF-1α, and VEGF protein expression was
effectively decreased in sevoflurane-induced human neuroglioma
cells, compared control group. However, treatment with Vitexin
significantly suppressed HIF-1α and p-p38 MAPK expression and
induced VEGF protein expression in sevoflurane-induced human
neuroglioma cells (Fig. 8).
Discussion
However, as we all know, the acting mechanisms of
both intravenous anesthetic and inhaled anesthetic have not been
clear so far (11). As for such a
drug in the central nervous system, the safety deserves more
attention when it facilitates surgical operation at the same time,
especially the problem whether young children's neurological
development is adversely affected, which is becoming urgent for
medical workers to solve (11). More
and more studies have gradually confirmed that there is a certain
degree of neurotoxicity in sevoflurane and other inhaled
anesthetics, which have significant impact on the development of
children's cognitive abilities (12). In the current study, we observed that
treatment with vitexin increased the cell proliferation of
sevoflurane-induced human neuroglioma cells. Treatment with Vitexin
significantly inhibited MDA level and increased SOD and GSH-px
levels in sevoflurane-induced newborn rat or human neuroglioma
cells. Sun et al suggested that vitexin possesses
cardioprotective action against DOX-induced cardiotoxicity through
anti-oxidative effects (10).
Bax is a Bcl-2 superfamily. Bcl-2 is an oncogene
isolated from follicular lymphoma, namely, B cell
lymphoma/Leukernia-2 (B cell lymphoma/Leukernia-2). Bcl-2 family
can be divided into two categories: One is to promote apoptosis,
namely, Bax, Bcl-xs, Bad, Bak, etc.; the other is to inhibit cell
apoptosis, namely, Bcl-2, Bcl-xl, Mcl-1, Bag-1, Ced-9 and some
viral genes (13). Once the cells
are induced by apoptotic factors, they can transduce to the
mitochondria, form a transmembrane channel in the mitochondrial
outer membrane by oligomerization, or open the PT pore of the
mitochondria, leading to the release of apoptotic factors in the
mitochondria, to activate caspases, thus leading to cell apoptosis
(14). These findings indicated that
vitexin suppressed Bax protein expression n sevoflurane-induced
newborn rat or human neuroglioma cells. Dong et al
demonstrated that vitexin protects against myocardial
ischemia/reperfusion injury through suppression of Bax expression
(15).
Caspases can be spontaneously activated or activated
by a similar cascade of specific enzymes (16). The cascade of hydrolysed proteins is
similar to the ‘waterfall effect’ of coagulation factor activation
(17). Recent studies have shown
that caspase family plays an important role in ischemic brain
injury, as the start and execution of mammalian apoptosis, in which
caspase-3 is the most important apoptotic protease caspasein the
downstream of cascade ‘waterfall’, and plays a pivotal role in the
apoptotic process initiated by a variety of factors (18). The present data demonstrated that
vitexin inhibited caspase-3 activity in sevoflurane-induced newborn
rat or human neuroglioma cells.
HIF-1 is an oxygen-sensitive transcriptional
regulator, which regulates more than 70 downstream target genes,
including VEGF, EPO and GLUT (12).
Both of them play an important role in the protection of cerebral
ischemic injury. HIF-1 has two subunits: HIF-1 and HIF-1β (6). Some studies have shown that the
upregulation of the expression of HIF-1 can play a neuroprotective
role in the focal cerebral ischemic injury model, but a few studies
suggest that its elevated expression may have some adverse effects
(6). The results of the present
study demonstrated that the protein of HIF-1α expression in
sevoflurane-induced newborn rat or human neuroglioma cells was
induced by vitexin. The function and implication of HIF-1α was
behaved in hypoxi, however, the implication of HIF-1α is limited
value in normoxic conditions in sevoflurane-induced newborn rat or
human neuroglioma cells. We need to be investigated in hypoxic and
normoxic conditions for the further study.
It is believed that VEGF has a direct role in
neurotrophy and protection in addition to the promoting effect on
the growth of new blood vessels (19). Recently, it has been found that VEGF
can protect injured neurons and promote vascular repair and nerve
regeneration in injured spinal cord tissue by acting on the
receptor, rapidly inducing nerve cell self-rescue, promoting nerve
axon regeneration and Schwann cell growth, etc (20). VEGF can also promote the
proliferation of spinal glial cells, especially astrocytes, and
secrete glial cell line-derived neurotrophic factor through VEGFR-2
receptor (20). GDNF can prevent
neuronal apoptosis and promote axon growth, thus indirectly
protecting nerve cells (21). In the
present study, vitexin induced VEGF protein expression in
sevoflurane-induced newborn rat or human neuroglioma cells. Min
et al indicated that vitexin reduces hypoxia-ischemia
neonatal brain injury through suppression of HIF-1α and VEGF
expression (22). Choi et al
reported that Vitexin has anti-metastatic potential in PC12 cells
through HIF-1alpha inhibitor and VEGF expression (23). These results suggest that vitexin
suppressed HIF-1α and VEGF expression to prevent
sevoflurane-induced neuronal apoptosis.
P38 mitogen-activated protein kinase (MAPK)
signaling pathway is involved in the late apoptosis of a large
number of neurons following ischemic brain injury. It is one of the
important signal transduction pathways of apoptosis induced by
ischemia reperfusion injury (8).
p-p38 MAPK activates the transcription factorsin downstream and
caspase family members (Caspase-3, −6 and −7) (24). In recent years, researchers use the
TUNEL method to label DNA fragments and have confirmed that the
hypoxia-ischemia in cultured neurons and in vivo can lead to
neuronal apoptosis (16). The
present findings indicated that vitexin suppressed p-p-38 MAPK
protein expression in sevoflurane-induced newborn rat or human
neuroglioma cells. Wang et al showed that vitexin alleviates
LPS-induced islet cell injury by inhibiting of p38 signaling
pathways (25).
In conclusion, this study indicated that the
protective effect of vitexin reduces sevoflurane-induced neuronal
apoptosis through HIF-1α, VEGF and p38 MAPK signaling pathway in
newborn rat or human cell. In this study, we used newborn rat model
and human neuroglioma H4 cells model, which showed that the
protective effect of vitexin reduces neuronal apoptosis in
vitro and in newborn rats by sevoflurane. Furthermore, our
results suggest that vitexin protects sevoflurane-induced neuronal
apoptosis in brain, which may be assisted adverse reactions during
anesthetic in clinical application.
References
|
1
|
Liu X, Song X, Yuan T, He J, Wang X and
Wang Q: Effects of calpain on sevoflurane-induced aged rats
hippocampal neuronal apoptosis. Aging Clin Exp Res. 28:633–639.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Istaphanous GK, Howard J, Nan X, Hughes
EA, McCann JC, McAuliffe JJ, Danzer SC and Loepke AW: Comparison of
the neuroapoptotic properties of equipotent anesthetic
concentrations of desflurane, isoflurane, or sevoflurane in
neonatal mice. Anesthesiology. 114:578–587. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Zhang DX, Zhang LM, Zhao XC and Sun W:
Neuroprotective effects of erythropoietin against
sevoflurane-induced neuronal apoptosis in primary rat cortical
neurons involving the EPOR-Erk1/2-Nrf2/Bach1 signal pathway. Biomed
Pharmacother. 87:332–341. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Wang LY, Tang ZJ and Han YZ:
Neuroprotective effects of caffeic acid phenethyl ester against
sevofluraneinduced neuronal degeneration in the hippocampus of
neonatal rats involve MAPK and PI3K/Akt signaling pathways. Mol Med
Rep. 14:3403–3412. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Bellanti F, Mirabella L, Mitarotonda D,
Blonda M, Tamborra R, Cinnella G, Fersini A, Ambrosi A, Dambrosio
M, Vendemiale G and Serviddio G: Propofol but not sevoflurane
prevents mitochondrial dysfunction and oxidative stress by limiting
HIF-1α activation in hepatic ischemia/reperfusion injury. Free
Radic Biol Med. 96:323–333. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Ye Z, Guo Q, Xia P, Wang N, Wang E and
Yuan Y: Sevoflurane postconditioning involves an up-regulation of
HIF-1α and HO-1 expression via PI3K/Akt pathway in a rat model of
focal cerebral ischemia. Brain Res. 1463:63–74. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Aminzadeh A, Dehpour AR, Safa M,
Mirzamohammadi S and Sharifi AM: Investigating the protective
effect of lithium against high glucose-induced neurotoxicity in
PC12 cells: Involvements of ROS, JNK and P38 MAPKs, and apoptotic
mitochondria pathway. Cell Mol Neurobiol. 34:1143–1150. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Ghasemi R, Zarifkar A, Rastegar K,
Maghsoudi N and Moosavi M: Insulin protects against Aβ-induced
spatial memory impairment, hippocampal apoptosis and MAPKs
signaling disruption. Neuropharmacology. 85:113–120. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Che X, Wang X, Zhang J, Peng C, Zhen Y,
Shao X, Zhang G and Dong L: Vitexin exerts cardioprotective effect
on chronic myocardial ischemia/reperfusion injury in rats via
inhibiting myocardial apoptosis and lipid peroxidation. Am J Transl
Res. 8:3319–3328. 2016.PubMed/NCBI
|
|
10
|
Sun Z, Yan B, Yu WY, Yao X, Ma X, Sheng G
and Ma Q: Vitexin attenuates acute doxorubicin cardiotoxicity in
rats via the suppression of oxidative stress, inflammation and
apoptosis and the activation of FOXO3a. Exp Ther Med. 12:1879–1884.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Zhang LM, Zhao XC, Sun WB, Li R and Jiang
XJ: Sevoflurane post-conditioning protects primary rat cortical
neurons against oxygen-glucose deprivation/resuscitation via
down-regulation in mitochondrial apoptosis axis of Bid, Bim,
Puma-Bax and Bak mediated by Erk1/2. J Neurol Sci. 357:80–87. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Liu XL, Lu J and Xing J: Stabilization of
HIF-1α modulates VEGF and Caspase-3 in the hippocampus of rats
following transient global ischemia induced by asphyxial cardiac
arrest. Life Sci. 151:243–249. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Zuo D, Lin L, Liu Y, Wang C, Xu J, Sun F,
Li L, Li Z and Wu Y: Baicalin attenuates ketamine-induced
neurotoxicity in the developing rats: Involvement of PI3K/Akt and
CREB/BDNF/Bcl-2 pathways. Neurotox Res. 30:159–172. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Nam YJ, Kim A, Lee MS, Shin YK, Sohn DS
and Lee CS: Lamotrigine attenuates proteasome inhibition-induced
apoptosis by suppressing the activation of the mitochondrial
pathway and the caspase-8- and bid-dependent pathways. Neurochem
Res. 41:2503–2516. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Dong L, Fan Y, Shao X and Chen Z: Vitexin
protects against myocardial ischemia/reperfusion injury in
Langendorff-perfused rat hearts by attenuating inflammatory
response and apoptosis. Food Chem Toxicol. 49:3211–3216. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
So EC, Lin YX, Tseng CH, Pan BS, Cheng KS,
Wong KL, Hao LJ, Wang YK and Huang BM: Midazolam induces apoptosis
in MA-10 mouse Leydig tumor cells through caspase activation and
the involvement of MAPK signaling pathway. Onco Targets Ther.
7:211–221. 2014.PubMed/NCBI
|
|
17
|
Liu HC, Zhang Y, Zhang S, Xin T, Li WH, Wu
WL, Pang Q and Chen YZ: Correlation research on the protein
expression (p75NTR, bax, bcl-2, and caspase-3) and cortical neuron
apoptosis following mechanical injury in rat. Eur Rev Med Pharmacol
Sci. 19:3459–3467. 2015.PubMed/NCBI
|
|
18
|
Chen B, Wang G, Li W, Liu W, Lin R, Tao J,
Jiang M, Chen L and Wang Y: Memantine attenuates cell apoptosis by
suppressing the calpain-caspase-3 pathway in an experimental model
of ischemic stroke. Exp Cell Res. 351:163–172. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Cui W, Li W, Han R, Mak S, Zhang H, Hu S,
Rong J and Han Y: PI3-K/Akt and ERK pathways activated by VEGF play
opposite roles in MPP+-induced neuronal apoptosis.
Neurochem Int. 59:945–953. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Cui W, Li W, Zhao Y, Mak S, Gao Y, Luo J,
Zhang H, Liu Y, Carlier PR, Rong J and Han Y: Preventing
H2O2-induced apoptosis in cerebellar granule
neurons by regulating the VEGFR-2/Akt signaling pathway using a
novel dimeric antiacetylcholinesterase bis(12)-hupyridone. Brain
Res. 1394:14–23. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Rong W, Wang J, Liu X, Jiang L, Wei F, Hu
X, Han X and Liu Z: Naringin treatment improves functional recovery
by increasing BDNF and VEGF expression, inhibiting neuronal
apoptosis after spinal cord injury. Neurochem Res. 37:1615–1623.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Min JW, Hu JJ, He M, Sanchez RM, Huang WX,
Liu YQ, Bsoul NB, Han S, Yin J, Liu WH, et al: Vitexin reduces
hypoxia-ischemia neonatal brain injury by the inhibition of
HIF-1alpha in a rat pup model. Neuropharmacology. 99:38–50. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Choi HJ, Eun JS, Kim BG, Kim SY, Jeon H
and Soh Y: Vitexin, an HIF-1alpha inhibitor, has anti-metastatic
potential in PC12 cells. Mol Cells. 22:291–299. 2006.PubMed/NCBI
|
|
24
|
Kwon SH, Kim JA, Hong SI, Jung YH, Kim HC,
Lee SY and Jang CG: Loganin protects against hydrogen
peroxide-induced apoptosis by inhibiting phosphorylation of JNK,
p38, and ERK 1/2 MAPKs in SH-SY5Y cells. Neurochem Int. 58:533–541.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Wang F, Yin J, Ma Y, Jiang H and Li Y:
Vitexin alleviates lipopolysaccharide-induced islet cell injury by
inhibiting HMGB1 release. Mol Med Rep. 15:1079–1086. 2017.
View Article : Google Scholar : PubMed/NCBI
|