Introduction
Food allergy is a pathological immune response
triggered by the exposure to allergenic foods and it results in
clinical symptoms, including gastrointestinal disorders (1). Allergenic reactions may be triggered by
dairy and other food products, including milk, peanuts, nuts,
shellfish and eggs (2). This broad
allergenic food spectrum suggests a high prevalence of food
allergies worldwide (3). A
retrospective study demonstrated that ≤6.7% of children in the
United States have allergies to different food (4). A similar or increased food allergy
prevalence in other countries has been reported in recent years
(5). A previous study in China
declared that the prevalence of food allergy in schoolchildren of
Guangzhou and Shaoguan was 4 and 3.5% respectively, in 2015
(6). Such prevalence results in an
increased number of food allergy anaphylaxis-associated hospital
admissions and high costs to healthcare systems worldwide (5,7,8). The elucidation of the mechanisms and
the development of preventive methods and novel therapies for
patients with food allergies are of great value.
Sensitization to food antigens can result in
inflammation and clinical symptoms similar to various common food
allergies, which can be mechanistically classified into
IgE-mediated, non-IgE-mediated and mixed-food allergies (9,10).
Generally, the immune system in mammals distinguishes pathogenic
antigens from harmless environmental antigens and maintains a state
of tolerance to common food antigens (11). Oral tolerance is an active process
that depends on diverse immune cell collaborations, including
resident CD103+ dendritic cells (DCs) and regulatory T
cells (Tregs) in the mucosa (12–14).
Epithelial damage or inflammation in the gut allows an increased
antigen entry and promotes epithelial cells to secrete inflammatory
cytokines, including interleukin (IL)-33, IL-25 and thymic stromal
lymphopoietin (15). These cytokines
reprogram the properties of mucosal DCs and Tregs and induce the
immune system towards a T helper 2 (Th2) cell response (16). The data from mouse models and
patients with allergies demonstrated that food allergy is
associated with a Th2 dominant response (17,18).
Therefore, Th2 cells mediate the immune response and Th2-derived
cytokine signaling pathways are potential targets in food allergy
treatment.
Mesenchymal stromal cells (MSCs) are multipotent
stem cells with self-renewing abilities, a differentiation
potential and they were identified by Friedenstein et al in
1968 (19). In the last decade,
accumulating data have suggested that MSCs have distinct immune
properties and these cells have gained considerable attention as
candidates for therapy in immune-associated diseases (20–22).
MSCs express the major histocompatibility complex (MHC) class I
molecule, but not MHC class II or co-stimulatory molecules,
including CD80 and CD86 (23). This
expression enables MSCs to avoid allogeneic rejection, which is
mediated by alloreactive T and natural killer cells (24). MSCs have been applied as
immunomodulators for autoimmune diseases and transplantation
rejection (23,25). In inflammation, MSCs can suppress T
cell-mediated responsiveness through the concerted action of
chemokines and nitric oxide, and can promote regulatory cell
differentiation (26,27). MSCs further regulate adaptive
immunity by reprogramming the maturation and phenotype of DCs
(28,29). Therefore, MSCs regulate the effects
of various immune cells through multiple mechanisms, which include
immunomodulatory soluble factor secretion and membrane-membrane
direct contact (20).
Bone marrow (BM)-derived MSCs were the first MSCs to
be identified and are best characterized. However, the special
handling of BM-MSCs limits their clinical application due to a low
frequency of cells and the invasive isolation procedure (30). Therefore, other tissues are now used
to isolate MSCs, including adipose tissue and the umbilical cord
(22,31). Human umbilical cord (hUC)-derived
MSCs exhibit an immunosuppressive capability, which is similar to
BM-MSCs, with regards to T cell activation and proliferation
(32). Increasing numbers of
experimental and clinic studies suggest that hUC-MSCs further
exhibit therapeutic effects in autoimmune diseases, including
diabetes, Crohn's disease and systemic lupus erythematosus
(33–35). As an alternative source of MSCs,
preparation from the hUC exhibits fewer ethical constraints and
increased maneuverability compared with human BM (30). Furthermore, hUC-MSCs can be prepared
in large quantities for therapeutic application in the clinic
(30).
In the current study, the therapeutic effects of
hUC-MSCs in food allergy treatment were investigated using an
ovalbumin (OVA)-induced mouse model. Following treatment with
hUC-MSCs, the main clinical symptoms and enteropathy in the allergy
model significantly improved. Simultaneously, the levels of Th2
cells and IgE in the blood, and IL-4 mRNA levels in the colon were
significantly decreased. The current experiments demonstrated a
potential therapeutic function of hUC-MSCs in the treatment of food
allergies.
Materials and methods
hUC-MSC culture
hUC-MSCs were isolated as previously described
(36). Fresh human umbilical cords
were obtained from 6 patients (age, 23–38 years) in the Fourth
Affiliated Hospital of Jiangsu University (Jiangsu, China) from
March 2016 to May 2017. Maternal blood samples were previously
screened and patients were screened and found negative for
infectious disease markers, including HIV I & II, HBV, HCV and
syphilis. Umbilical cord samples were cut into 1–2 mm3
pieces and were floated on Dulbecco's modified Eagle's medium
(DMEM)/F12 (1:1; L310KJ; Shanghai Basalmedia Technologies Co.,
Ltd., Shanghai, China) with 10% fetal bovine serum (FBS; Biowest,
Nuaillé, France), 100 U/ml penicillin and streptomycin at 37°C with
5% CO2. Medium was replaced every 3 days. When the
adherent cells reached 80–90% confluence, cultures were trypsinized
and transferred into a new flask for further expansion. The
phenotypes of hUC-MSCs were analyzed via flow cytometric analysis.
Following trypsinization and washing with PBS solution twice, the
single cell suspensions were blocked with 2% rabbit serum (Stemcell
Technologies, Vancouver, BC, Canada) at room temperature for 30
min. Samples were then labeled with different fluorescent
antibodies (1:200), from Thermo Fisher Scientific, Inc. (Waltham,
MA, USA) at 4°C for 30 min. Cells were further washed with PBS in
triplicate, re-suspended with PBS (1 mM EDTA) and analyzed with a
FACS Calibur flow cytometer (BD Bioscience, Franklin Lakes, NJ,
USA). All data were analyzed using FlowJo (version 8.7; Tree Star,
Inc., Ashland, OR, USA). The following fluorescent antibodies
(Thermo Fisher Scientific, Inc.) were utilized: Anti-human
CD73-Fluorescein isothiocyanate (FITC; cat. no. 11-0739-41),
anti-human CD90-FITC (cat. no. 11-0909-41), anti-human
CD105-phycoerythrin (PE; cat. no. 12-1057-41), anti-human CD34-FITC
(cat. no. 11-0349-41), anti-human CD45-FITC (cat. no. 11-0459-41),
anti-human-CD14-FITC (cat. no. 11-0149-42), anti-human-CD19-PE
(cat. no. 50-102-58) and anti-human HLA-DR-FITC (cat. no.
11-9952-41). The protocol for human tissue collection was approved
by the Ethical Committee of Jiangsu University and informed consent
for umbilical cord tissue collection was provided by the
patients.
hUC-MSC differentiation in vitro
Differentiation studies were performed as described
previously with few modifications (37). Briefly, for adipogenic
differentiation, cells were cultured at 37°C with 5% CO2
in DMEM/F12 (1:1) complete medium supplemented with 0.5 µM
isobutylmethylxanthine, 1 µM dexamethasone, 10 µM insulin and 60 µM
indomethacin (all Sigma-Aldrich; Merck KGaA, Darmstadt, Germany)
for 14 days. For osteogenic differentiation, cells were cultured at
37°C for 28 days with 5% CO2 in the complete medium with
0.1 µΜ dexamethasone, 10 mM β-glycerol phosphate and 50 µΜ
ascorbate (all Sigma-Aldrich; Merck KGaA) for 28 days. Oil Red O
and Alizarin Red staining was performed to visualize the adipogenic
and osteogenic differentiations, respectively. Differentiated
adipogenic cells were stained on day 14 with Oil Red O (ORO)
staining kit (Solarbio Technology Co., Ltd, Beijing, China)
according to the manufacturer's protocol. Cells were fixed with ORO
fixative buffer for 10–15 min at room temperature, freshly prepared
ORO staining solution was (mixed previously in the kit with
distilled water in a 3:2 ratio) and dipped for 15 min at room
temperature. The staining solution was then removed and cells were
rinsed with 60% isopropyl alcohol for 20–30 sec. Samples were
rinsed with PBS and stained with Mayer hematoxylin for 2 min at
room temperature. ORO buffer was then added for 1 min. Cells were
then washed with distilled water and dried. To perform Alizarin Red
staining on day 28, differentiated osteogenic cells were stained
using an Alizarin Red staining kit (Solarbio Technology Co., Ltd,
Beijing, china) according to the manufacturer's protocol. Cells
were fixed with 95% ethanol for 10–15 min at room temperature. The
fixative solution was added for alizarin red S staining and dipped
for 20 min. Following washing, cells were then dried. All stained
cells were observed under microscope (Nikon Corporation, Tokyo,
Japan) and photographs were obtained.
Food allergy model
A total of 20 mice Female BALB/c mice
(H-2d; age, 6–8 weeks; weight, 25±2 g) were obtained
from the Comparative Medicine Centre of Yangzhou University
(Yangzhou, China) and divided into four groups. Mice were housed in
microisolator units under a 12 h light/dark cycle at a temperature
of 24–26°C and a humidity of 30–50%, and with ad libitum
access to food and water. To induce food allergy, Mice (n=5 per
group, a total of 15 mice in three OVA challenged groups) were
intraperitoneally (i.p.) injected with 50 µg OVA (Grade V; A-5503;
Sigma-Aldrich; Merck KGaA) in 1 mg alum adjuvant on days 0 and 14
as previously described (38). From
day 28, the mice were challenged intragastrically with 50 mg of OVA
in 250 µl PBS every other day for 12 days (6 administrations). As
immune regulatory functions of MSCs depend on the cell-cell contact
and the secreted soluble factors in the medium, mice were treated
with MSCs and MSC conditioned medium (39,40).
Mice in the different groups (n=5 per group) were respectively
challenged with 500 µl of PBS, OVA, medium/OVA and MSC
supernatant/OVA (MSC/OVA group) by oral gavage on day 28, 30, 32,
34, 36 and 38. Subsequently, all mice were deprived of food for 3 h
to limit antigen degradation in the stomach. In addition, mice in
the MSC/OVA group were i.p. injected with 5×105 hUC-MSCs
in 200 µl PBS every other day from days 21–38. Symptoms of allergic
diarrhea were assessed 1 h following OVA oral challenge (day 28,
30, 32, 34, 36 and 38) and severity was classified as follows:
Normal, soft and loose stool, mild and severe diarrhea, and fluid
stool. Mice with profuse liquid stool (mild or severe diarrhea, or
fluid stool) were recorded as having diarrhea (41). On day 38, all mice were sacrificed
and their colons and mesenteric lymph nodes were collected. The
amounts of solid faeces within colon specimens were observed and
lymph nodes were weighed. All animal protocols have been approved
by the Animal Care and Use Committee of Jiangsu University.
Concanavalin (Con) A-induced
CD4+ T cell proliferation
To detect the immunosupression of hUC-MSC cells or
MSC cell culture supernatant on CD4+ T cell activation
and proliferation, 6-week old BALB/c mice were sacrificed via
CO2 exposure at a flow rate of 3 l per min and spleens
were collected to prepare cell suspensions for experiment as
previously described (42). Spleens
were collected, grinded and passed through strainers in PBS to
prepare cell suspensions. Red blood cells in these splenocyte
suspensions were removed using ACK lysis buffer. Cells were
subsequently mixed with 5 µM carboxyfluorescein succinimidyl ester
(Sigma-Aldrich; Merck KGaA) and incubated at 37°C for 10 min.
Following incubation in 1640 medium (PAA Laboratories; GE
Healthcare Life Sciences, Little Chalfont, UK) containing 10% FBS
at room temperature for 10 min, cells were washed with PBS (3X) and
plated at 2×106/well in 48-well plates with 5 µg/ml Con
A (Sigma-Aldrich; Merck KGaA). To assess the direct
immunosuppressive effects of MSC supernatants or hUC-MSC cells on
ConA activated T cells, half of the medium was replaced with
hUC-MSC culture supernatant or hUC-MSC cells
(2×105/well) in wells following splenocyte seeding,
which were then cultured at 37°C in 5% CO2 for 72 h.
Cells were collected and stained with fluorescein-labeled
anti-mouse CD4 (1:200; cat. no. 17-0041-81; eBioscience; Thermo
Fisher Scientific, Inc.) at 4°C for 30 min. 7-aminoactinomycin D
(7-AAD) was then added prior to analysis (cat. no. 00-6993-50;
eBioscience; Thermo Fisher Scientific, Inc.). The proliferation of
CD4+ cells was analyzed by flow cytometry. All the data
were analyzed using FlowJo (version 8.7; FlowJo LLC, Ashland, OR,
USA).
Intracellular cytokine staining
On day 38, mice were sacrificed and mesenteric lymph
nodes were collected and mononuclear cells were prepared by gently
pressing the tissues through a strainer (40 µm; BD biosciences) in
cold PBS containing 1% FBS. Cell suspensions were washed twice with
cold PBS. On day 38, prior to sacrifice, murine blood (300 µl) was
collected from the orbital sinus to use for intracellular cytokine
staining, or to prepare serum by storing samples at room
temperature for 1 h following centrifugation at 1,500 × g for 15
min at 4°C. For intracellular staining, red blood cells (RBCs) in
samples were removed using RBC lysis buffer (cat. no. 00-4333-57,
eBioscience; Thermo Fisher Scientific, Inc.) according to the
manufacturer's protocol. Cells were resuspended in the presence of
a Cell Stimulation Cocktail plus protein transport inhibitors
(eBioscience; Thermo Fisher Scientific, Inc.) at 37°C in 5%
CO2 for 4 h. Then, cells were incubated with anti-mouse
CD4-allophycocyanin (eBioscience; Thermo Fisher Scientific, Inc.)
at 4°C for 20 min. Following washing with cold PBS (2X), cells were
fixed with IC Fixation buffer (eBioscience; Thermo Fisher
Scientific, Inc.) at 4°C for 30 min. Cells were washed twice more
with permeabilization buffer (eBioscience; Thermo Fisher
Scientific, Inc.) and incubated with anti-mouse interleukin
(IL)-4-phycoerythrin (eBioscience; Thermo Fisher Scientific, Inc.)
for 45 min at 4°C. Cells were washed twice with PBS and suspended
in PBS-bovine serum albumin (1%; Sigma-Aldrich; Merck KGaA) for the
flow cytometry analysis as described above.
ELISA analysis
Total IgE serum levels were measured using an ELISA
kit (cat. no. 88-50460-22, eBioscience; Thermo Fisher Scientific,
Inc.) according to the manufacturer's protocols (eBioscience;
Thermo Fisher Scientific, Inc.). Briefly, 96-well plates were
coated overnight at 4°C with 100 µl/well capture antibody (provided
by the kit). Following two washes with wash buffer, plates were
blocked with 250 µl blocking buffer at room temperature for 2 h.
Plates were washed twice with wash buffer and 100 µl serum sample
(1:50) was added to each well. Plates were further incubated at
room temperature for 2 h. Next, 100 µl/well biotinylated detection
antibody (obtained from the kit) was added and incubated at room
temperature for 1 h. Following 4 washes, 100 µl/well
streptavidin-horseradish peroxidase was added and incubated at room
temperature for 30 min. Wells were further washed 4 times, 100 µl
substrate solution was added per well and samples were incubated
for 30 min at room temperature. Finally, stop solution was added
and OD values were measured at 450 nm.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR) analysis
For PCR analysis, the transverse colon was excised
following the last OVA challenge on day 38 and flushed with
ice-cold PBS. Tissue samples were snap-frozen in liquid nitrogen
and stored at −80°C for further experiments. Tissue was homogenized
and total RNA was extracted with RNAiso Plus (Takara Biotechnology
Co., Ltd., Dalian, China) according to the manufacturer's protocol.
The cDNA was synthesized using a PrimeScript RT Master mix (Takara
Biotechnology Co., Ltd.) for 15 min at 37°C and then 15 sec at 85°C
according to the manufacturer's protocol. qPCR amplification of the
target cDNA was performed using the SYBR Premix Ex Taq (Takara
Biotechnology Co., Ltd.) according to the manufacturer's protocol.
Thermocycling conditions were as follows: 30 sec at 95°C, followed
by 40 cycles of 5 sec at 95°C and 20 sec at 58°C. Alterations in
expression of target genes in treated vs. untreated samples were
observed; mRNA levels were quantified using the 2−∆∆Cq
method (43). Levels were normalized
to β-actin (internal control) and the PBS group with three
replicates was used as calibrator. Primer sequences are as follows:
IL-4 forward, 5′-GGTCTCAACCCCCAGCTAGT-3′ and reverse,
5′-GCCGATGATCTCTCTCAAGTGAT-3′; tumor necrosis factor (TNF)-α
forward, 5′-GAACTGGCAGAAGAGGCACT-3′ and reverse,
5′-GGTCTGGGCCATAGAACTGA-3′; and β-actin forward,
5′-TGGCGCTTTTGACTCAGGAT-3′ and reverse,
5′-GGGATGTTTGCTCCAACCAA-3′.
Histological staining
Transverse colons were collected as aforementioned
in PCR analysis. Following snap-freezing, tissues were fixed with
4% paraformaldehyde solution for 48 h at 4°C. Following ethanol
treatment for dehydration (70% ethanol for 120 min, 80% ethanol for
120 min, 90% ethanol for 60 min, 95% ethanol for 40 min, 100%
ethanol for 40 min, 100% ethanol for 40 min) at room temperature,
colons were then embedded in paraffin (first wax, 58°C for 1 h;
second wax, 58°C for 1 h) and cut into 4-µm-thick sections.
Subsequently, sections were stained with hematoxylin and eosin at
room temperature using the following procedure: Samples were washed
twice with Xylene for 10 min, then twice with 100% ethanol for 5
min, 95% ethanol for 2 min, 70% ethanol for 2 min and rinsed with
water. Hematoxylin solution was then added for 8 min, rinsed with
water and added to 1% acid ethanol for 30 sec. Following a further
rinse with water, 0.2% ammonia water was added for 1 min and
samples were rinsed. Samples were dipped 10 times in 95% ethanol,
stained with eosin-phloxine solution for 45 sec, added to 95%
ethanol and 100% ethanol 5 min each and finally washed with xylene
twice for 5 min. For the periodic acid-Schiff (PAS) staining,
sections of the colons were stained with the Glycogen PAS Staining
kit (Yeasen Biotechnology, Shanghai, China) (www.yeasen.com) according to the manufacturer's
protocol. Briefly, paraffin-embedded slices were deparaffinized
with xylene twice for 10 min, hydrated with 100% ethanol twice for
5 min, 95 and 70% ethanol for 2 min, washed with distilled water
and oxidized at room temperature in periodic acid solution for 10
min. All deparaffinization and hydration steps were performed at
room temperature. Subsequently, sections were placed in Schiff
reagent for 10 min and counterstained with Mayer's hematoxylin
stain for 20–30 sec at room temperature. All stained sections were
sealed overnight at room temperature using neutral resin and
digital photographs were taken using an automatic digital slide
scanner under a microscope (ECLIPSE Ti, Nikon corporation;
magnification, ×100, ×400, ×1,000).
Sequencing of 16S ribosomal (r)RNA in
the stool
The sequencing of 16S ribosomal (r)RNA was performed
by Zhenjiang Tianyi Biotechnology Co., Ltd. (Zhenjiang, China).
Each kit was used according to the manufacter's protocol. Stool in
the colon was collected on day 38 under sterile conditions and
genomic DNA was extracted and purified using the QIAamp DNA Stool
Mini kit and the QIAquick PCR Purification kit, respectively
(Qiagen GmbH, Hilden, Germany). The PCR amplification of the target
V4-V5 regions of the bacterial 16S rRNA genes was firstly performed
using the primers 515F (5′-GTGCCGCCGCCGCGGTAA-3′) and 806R
(5′-GGACTACHVGGGTWTCTAAT-3′) and 2× Taq Master Mix (Vazyme Biotech
Co., Ltd., Nanjing, China). The thermocycling conditions were as
follows: 3 min at 95°C, followed by 35 cycles of 30 sec at 95°C, 45
sec at 55°C and 60 sec at 72°C, and 5 min at 72°C. DNA products
were used for further PCR amplification using a 2X Taq Master Mix
(Vazyme Biotech Co., Ltd., Nanjing, China) and the following
primers (44,45): 515F forward primer, barcoded
[5′-AATGATACGGCGACCACCGAGATCTACACGCT (5′ Illumina adapter) XXX XXX
XXX XXX (barcode) TATGGTAATT (pad) GT (linker)
GTGYCAGCMGCCGCGGTAA-3′ (515F forward primer)] and 806R reverse
primer [5′-CAAGCAGAAGACGGCATACGAGAT (3′ Illumina adapter)
AGTCAGCCAG (pad) CC (linker) GGACTACNVGGGTWTCTAAT-3′ (806R reverse
primer)]. Barcodes in the 515F forward primer enables the usage of
various reverse primer constructs to obtain longer amplicons.
Additional degeneracy (such as Y in 515F and N in 806R) was used to
add bias against environmental archaea
Crenarachaeota/Thaumarchaeota and the aquatic bacteria SAR11
(44,45). The Thermocycling conditions for the
second PCR assay were as follows: 3 min at 95°C, followed by 35
cycles of 45 sec at 95°C, 60 sec at 55°C and 90 sec at 72°C, and
then 10 min at 72°C. Amplicons were then purified using the
Agencourt AMPure XP kit (Beckman Coulter, Inc., Brea, CA, USA) and
quantified using a Qubit 3.0 fluorometer (Thermo Fisher Scientific,
Inc.). DNA samples were further submitted to the Illumina
sequencing platform (www.illumina.com) for analysis. The raw reads were
filtered for noise deletion and then analyzed using the open source
software package, Quantitative Insights into Microbial Ecology
(version 1; http://qiime.org/). Operational taxonomy
units (OTUs) of 16S rRNA were identified based on an open reference
OTU pick using UCLUST (version 4.1.93; http://qiime.org/), which clustered at a threshold of
97% pair-wise identities and were classified taxonomically using
the Ribosomal Database Project classifier 2.3 (ttps://rdp.cme.msu.edu/) (46).
Statistical analysis
The experiments were repeated at least three times
and data are presented as the mean ± standard deviation. All
statistical comparisons were made using one-way analysis of
variance followed by the Newman-Keuls multiple comparison post-hoc
tests using GraphPad Prism 5.0 (GraphPad Software, Inc., La Jolla,
CA, USA). P<0.05 was considered to indicate a statistically
significant difference.
Results
Characterization of hUC-MSCs
Human umbilical cord tissues were cultured for
>15 days and spindle-shaped fibroblastic cells were observed to
grow to confluence (Fig. 1A).
hUC-MSCs were identified using a phenotype analysis and the
capability of adipogenic and osteogenic differentiation (36,47). The
phenotype of the cells was determined by flow cytometry. Fig. 1B demonstrates that cells were
positive for CD73, CD90 and CD105, and negative for CD34
(hematopoietic stem cell), CD45 (leukocyte), CD14 (myeloid cell),
CD19 (B cell) and HLA-DR (DC, macrophage and B cell) (47–49).
When the cells were cultured in adipogenic medium to analyze
adipocyte differentiation, a portion of the cells contained lipid
droplets and tested positive for Oil-Red-O staining (Fig. 1C). The differentiation of cells in
osteogenic medium revealed a portion of cells with positive
Alizarin Red staining (Fig. 1D).
Fig. 1E further revealed that the
hUC-MSCs alone and the hUC-MSC-cultured supernatant inhibited the
proliferation of CD4+ T cells. These results suggested
that the cells cultured from umbilical cord tissues present with
MSCs features.
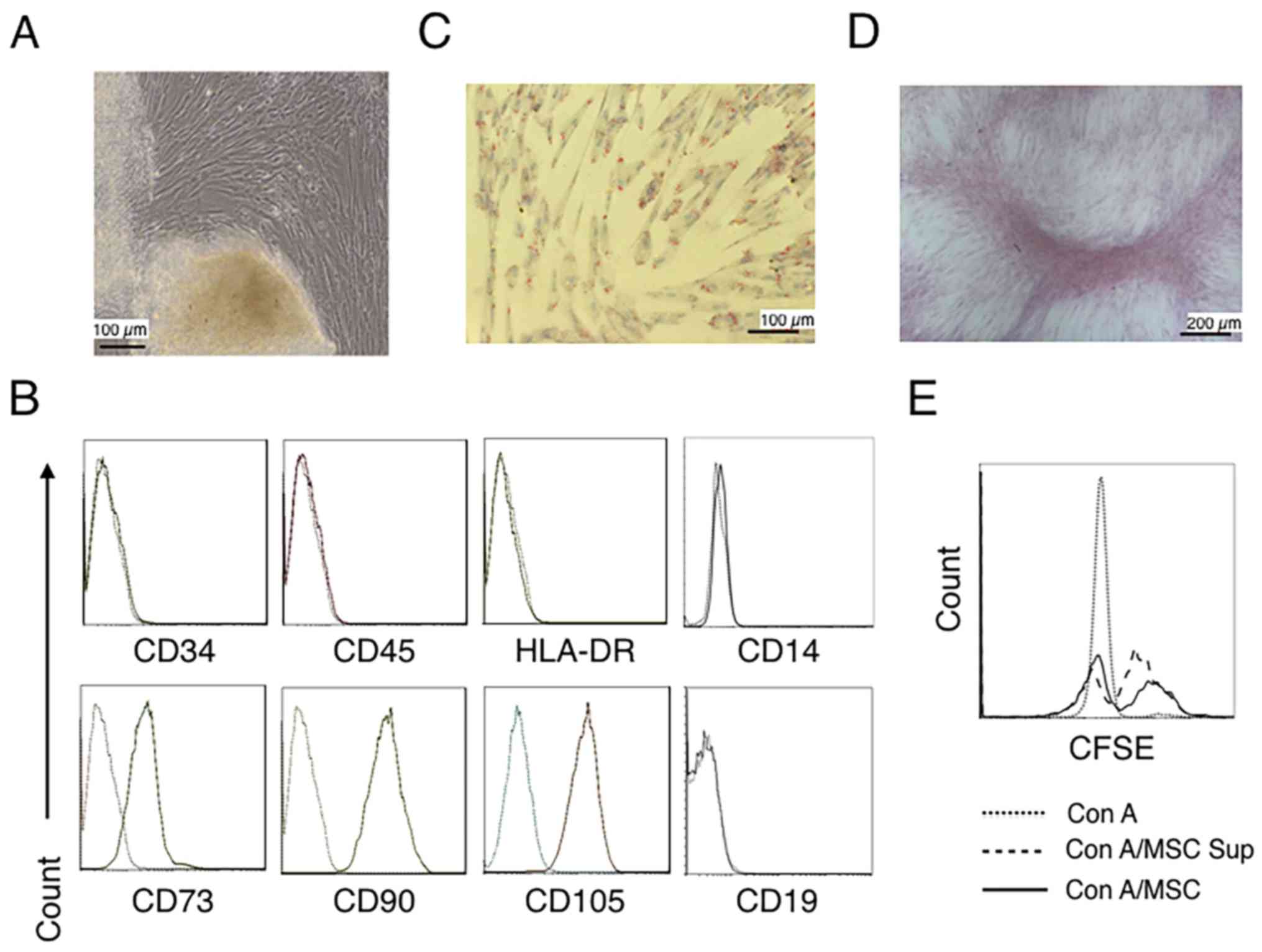 | Figure 1.Characterization of hUC-MSCs. (A)
Morphological observations of hUC-MSCs. Umbilical cord tissues were
cultured for >15 days and long spindle-shaded fibroblastic cells
were observed around the tissue using Zeiss light microscopy (scale
bar, 100 µm). (B) Phenotyping of hUC-MSCs. hUC-MSCs were stained
with a fluorescein-labeled antibodies (CD34, CD45, CD73, CD90,
CD105, CD14, CD19 and HLA-DR) and analyzed with a flow cytometer.
(C) Adipogenic and (D) osteogenic differentiation of hUC-MSCs.
hUC-MSCs were cultured in adipogenic and osteogenic medium,
respectively. Lipid droplets in the adipocytes are presented with
Oil Red O staining (scale bar, 100 µm). hUC-MSCs-derived
osteoblasts were detected with Alizarin Red staining (scale bar,
200 µm). (E) hUC-MSCs inhibit the proliferation of CFSE-labeled
CD4+ T cells, which were activated by Con A stimulation.
Experiments were repeated three times and representative graphs and
images are presented. hUC-MSC, human umbilical cord-derived
mesenchymal stem cell; MSC Sup, culture supernatant of hUC-MSCs;
Con A, concanavalin A; CFSE, carboxyfluorescein succinimidyl
ester. |
hUC-MSCs alleviate OVA-induced food
allergy symptoms
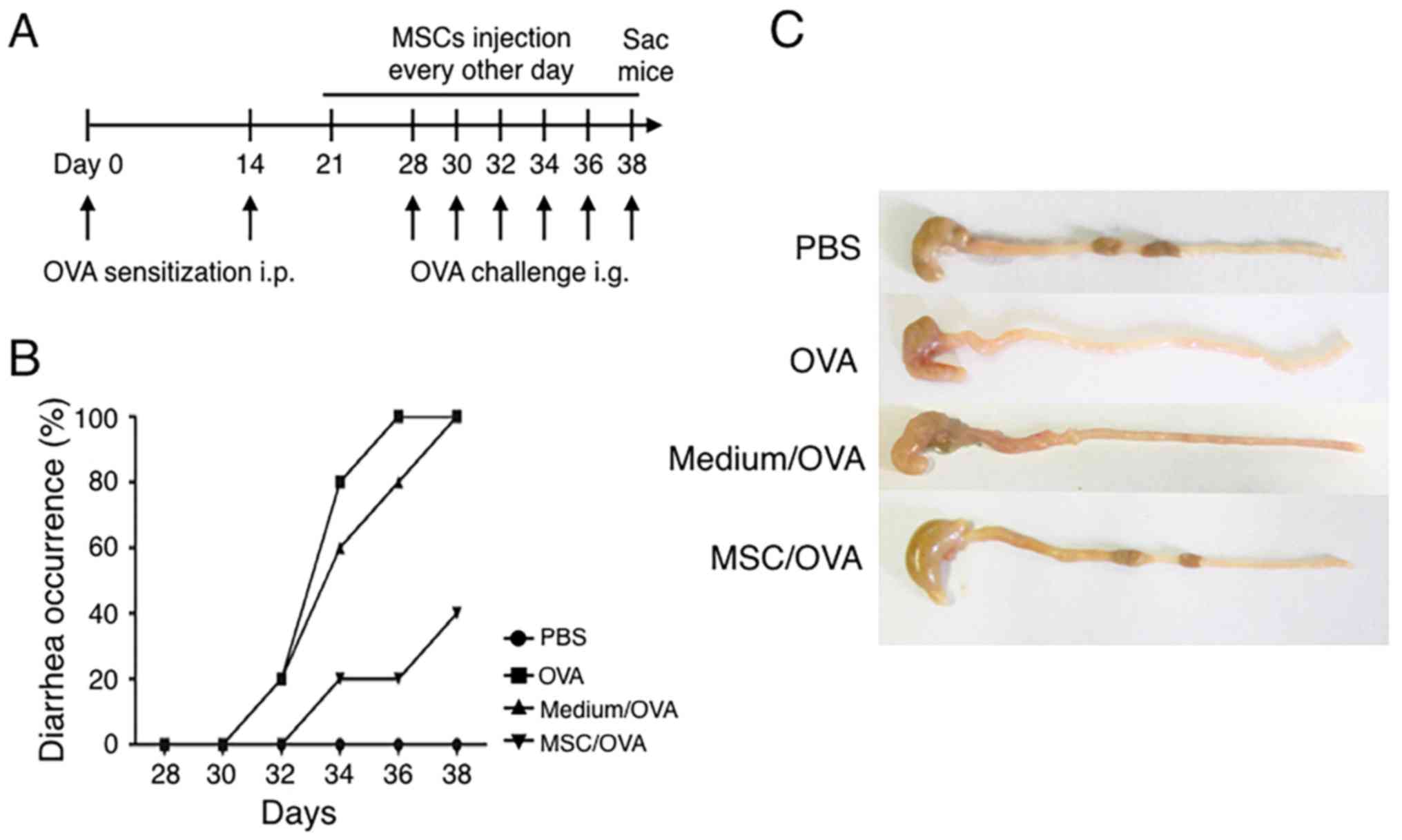
To explore effects of hUC-MSCs on food allergy, a
mouse model was prepared by OVA i.p. sensitization and oral
challenge (Fig. 2A). In the MSC/OVA
group, hUC-MSCs were i.p. injected every other day from day 21 to
day 38. These mice were also intragastrically administered with MSC
culture supernatant and simultaneously challenged with OVA antigen
on 6 alternating days. To account for medium effects, an equal
volume of medium was administered by oral gavage in conjunction
with OVA (medium/OVA group). The data in Fig. 2B demonstrated that OVA challenge
induced diarrhea on day 30, following 3 days of treatment, in the
PBS and medium/OVA groups. Following sacrifice on day 38, compared
with mice in the PBS and medium/OVA, an increased amount of solid
faeces was observed in the gut of the MSC/OVA mice (Fig. 2C). These results demonstrated that
hUC-MSC treatment attenuated the allergic diarrhea scores of
OVA-induced allergy in mice.
hUC-MSCs inhibit Th2 cells and IgE
production in mice
IgE-mediated degranulation of mast cells or
basophils results in the rapid manifestation of symptoms of food
allergy (50). The current study
detected the levels of IgE in the peripheral blood of mice. The
results indicated that IgE levels were increased in the PBS and
medium/OVA groups compared with the MSC/OVA mice (Fig. 3A). A Th2-type immune response,
stimulated by food allergens, contributes to the initiation and
development of allergies and associates with enteropathy (18). The percentage of IL-4+ Th2
cells in the peripheral blood and mesenteric lymph nodes were
further determined. Fig. 3B suggests
that OVA challenge markedly increased the percentage of
IL-4+ Th2 cells in CD4+ T cells in the blood.
Treatment with hUC-MSCs inhibited the frequency of IL-4+
Th2 cells. Comparable IL-4+ Th2 cell results were
detected in the mesenteric lymph nodes (Fig. 3C). No difference in the weight of the
lymph nodes was determined between the groups (Fig. 3D).
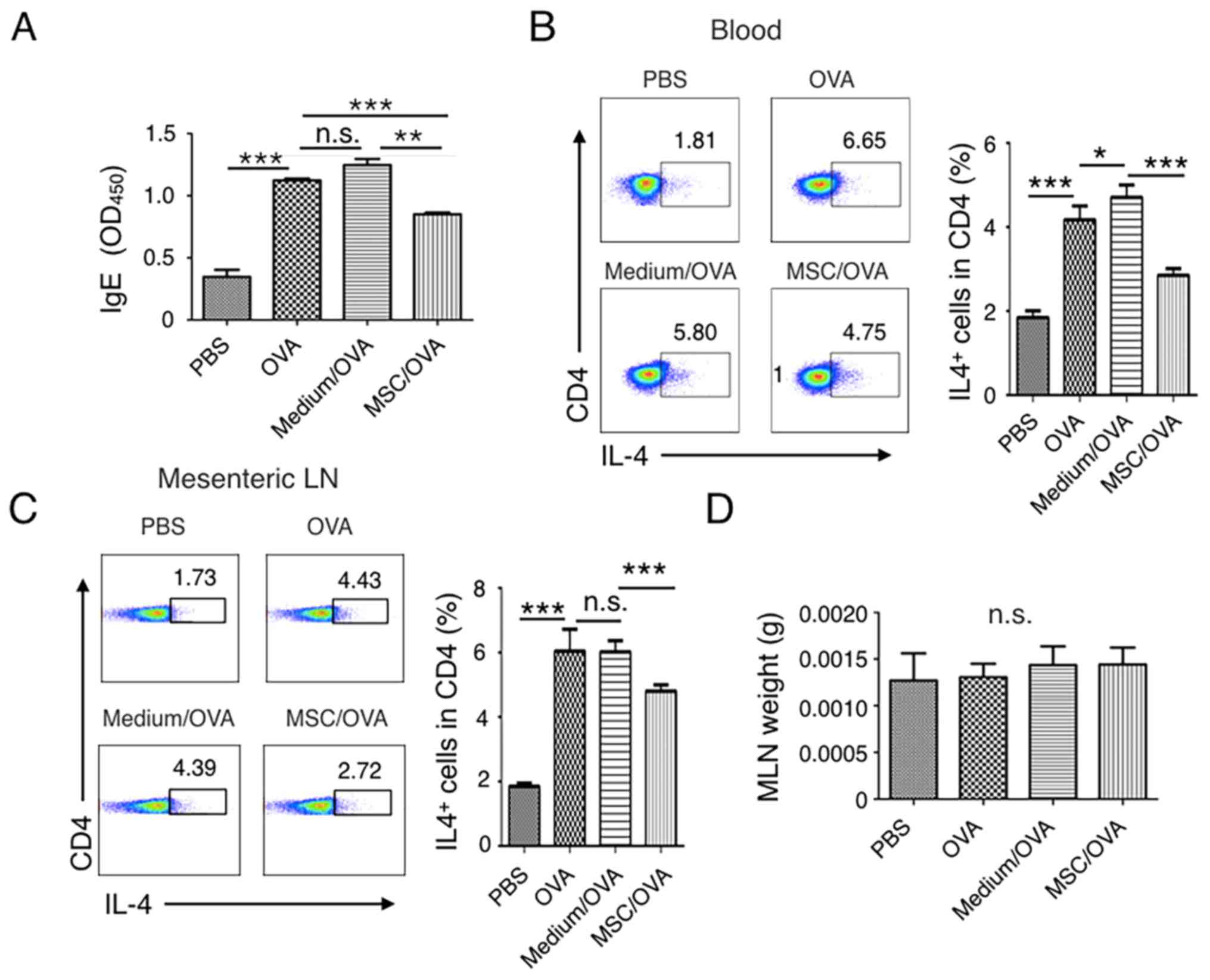 | Figure 3.Treatment with hUC-MSCs inhibits the
frequency of Th2 cells and IgE levels in the periphery. Mice were
divided into PBS, OVA, Medium/OVA and MSC/OVA treatment groups. (A)
Total IgE levels determined in the different groups. IgE levels in
the serum were detected by ELISA and OD450 values are presented.
The percentage of IL-4+ Th2 cells in (B) blood and (C)
MLNs were analyzed using a flow cytometer by gating the
CD4+ cells. Quantitative analysis on the percentage of
Th2 cells in (B) and (C) was also performed. Numbers in the graphs
represent the percentage of CD4+IL-4+ Th2
cells. (D) Weight of the MLNs of mice in the different treatment
groups, representative of four independent experiments. *P<0.05;
**P<0.01; ***P<0.001. hUC-MSC, human umbilical cord-derived
mesenchymal stem cell; Th2, T helper 2 cell; IL, interleukin; OD,
optical density; MLN, mesenteric lymph node; OVA, ovalbumin; n.s.,
not significant; Medium/OVA, group challenged with OVA and
administered Dulbecco's modified Eagle's medium/nutrient mixture
F12; MSC/OVA, group challenged with OVA, intraperitoneal injection
of hUC-MSCs and oral gavage of MSC culture supernatant. |
Treatment with hUC-MSCs reduces
inflammation and the numbers of goblet cells in the colons of mice
with food allergy
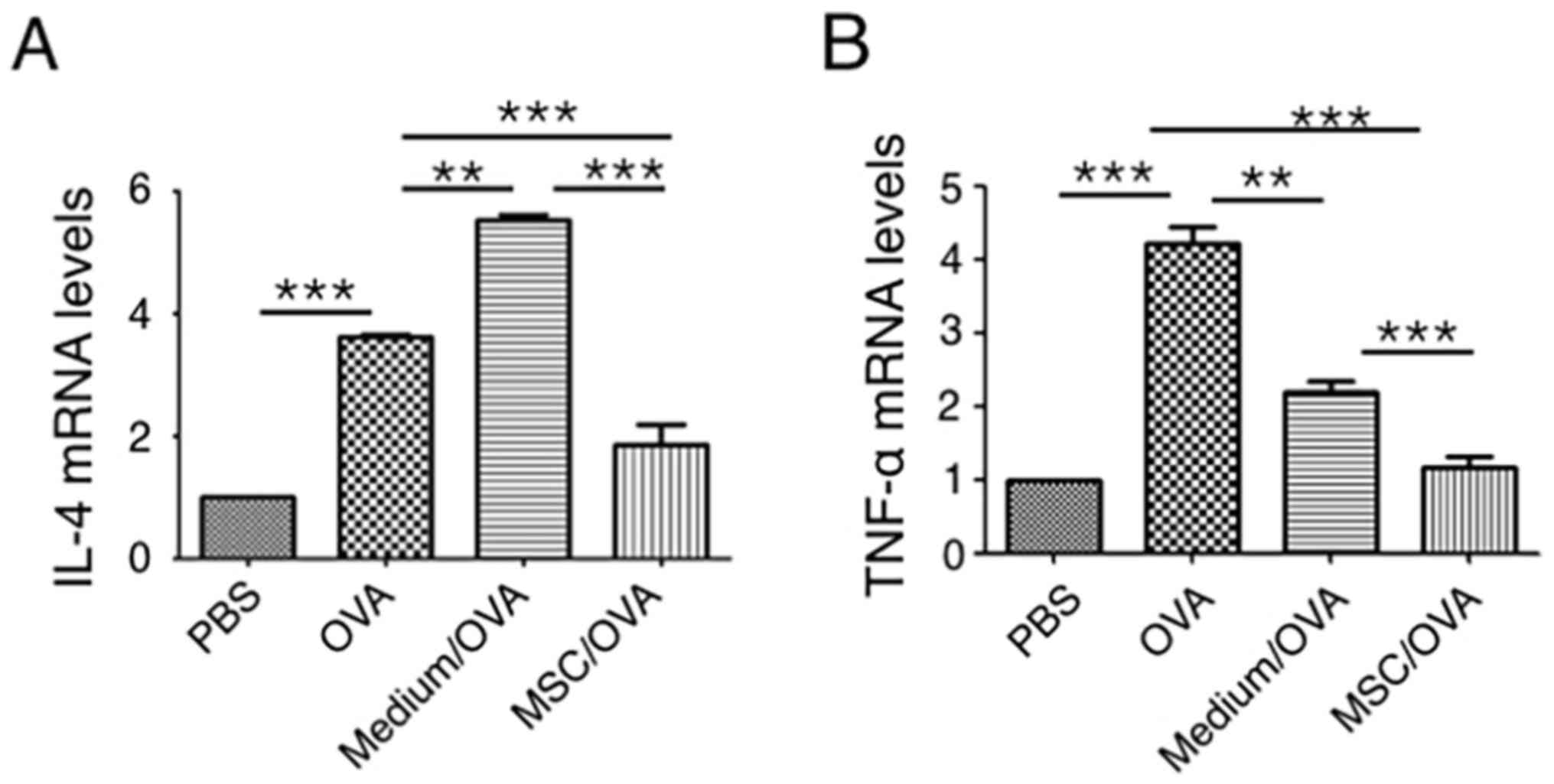
Cytokine profiles in mouse colons were analyzed
using RT-qPCR. The data in Fig. 4
indicates that OVA challenge significantly upregulates IL-4 and
TNF-α mRNA levels in the colons of the OVA group compared with the
PBS control. hUC-MSC treatment (MSC/OVA) significantly reversed the
OVA-induced higher mRNA levels of IL-4 and TNF-α in the OVA and the
medium/OVA groups. The data indicated that the inflammation in the
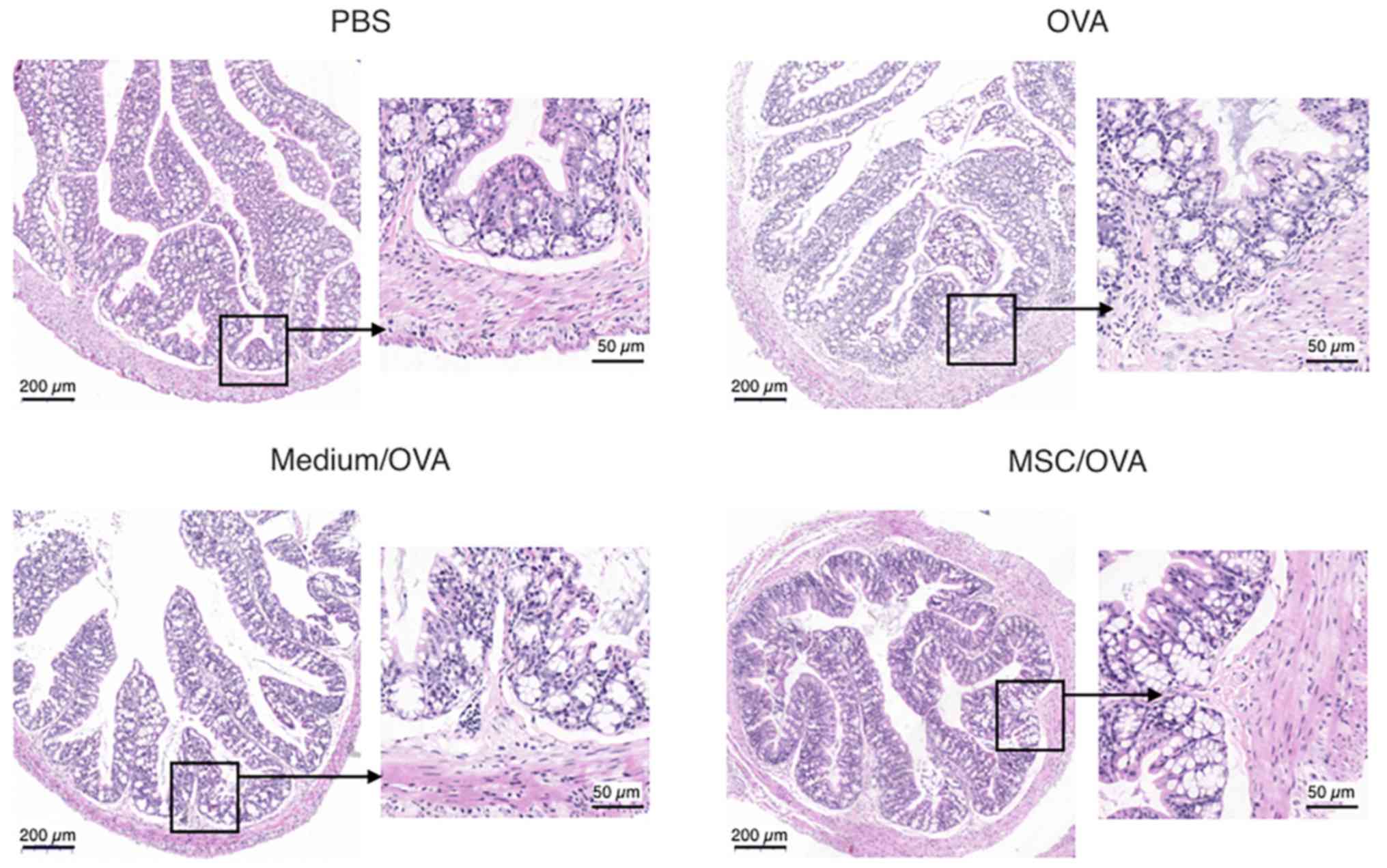
colon was suppressed by hUC-MSCs. This was further confirmed by
histological data of the colons (Fig.
5). Fewer infiltrating inflammatory cells were detected in the
lamina propria of the intestinal mucosa in the MSC/OVA group
compared with the OVA and medium/OVA groups (Fig. 5). By using PAS staining, more goblet
cells were detected in the OVA and medium/OVA group than in the PBS
control group (Fig. 6). If treated
with hUC-MSCs, the number of goblet cells was markedly decreased in
the colon compared with other OVA-challenged groups (Fig. 6). Collectively the data suggested
that treatment with hUC-MSCs inhibited the OVA-induced inflammation
and decreased the numbers of goblet cells in the colon.
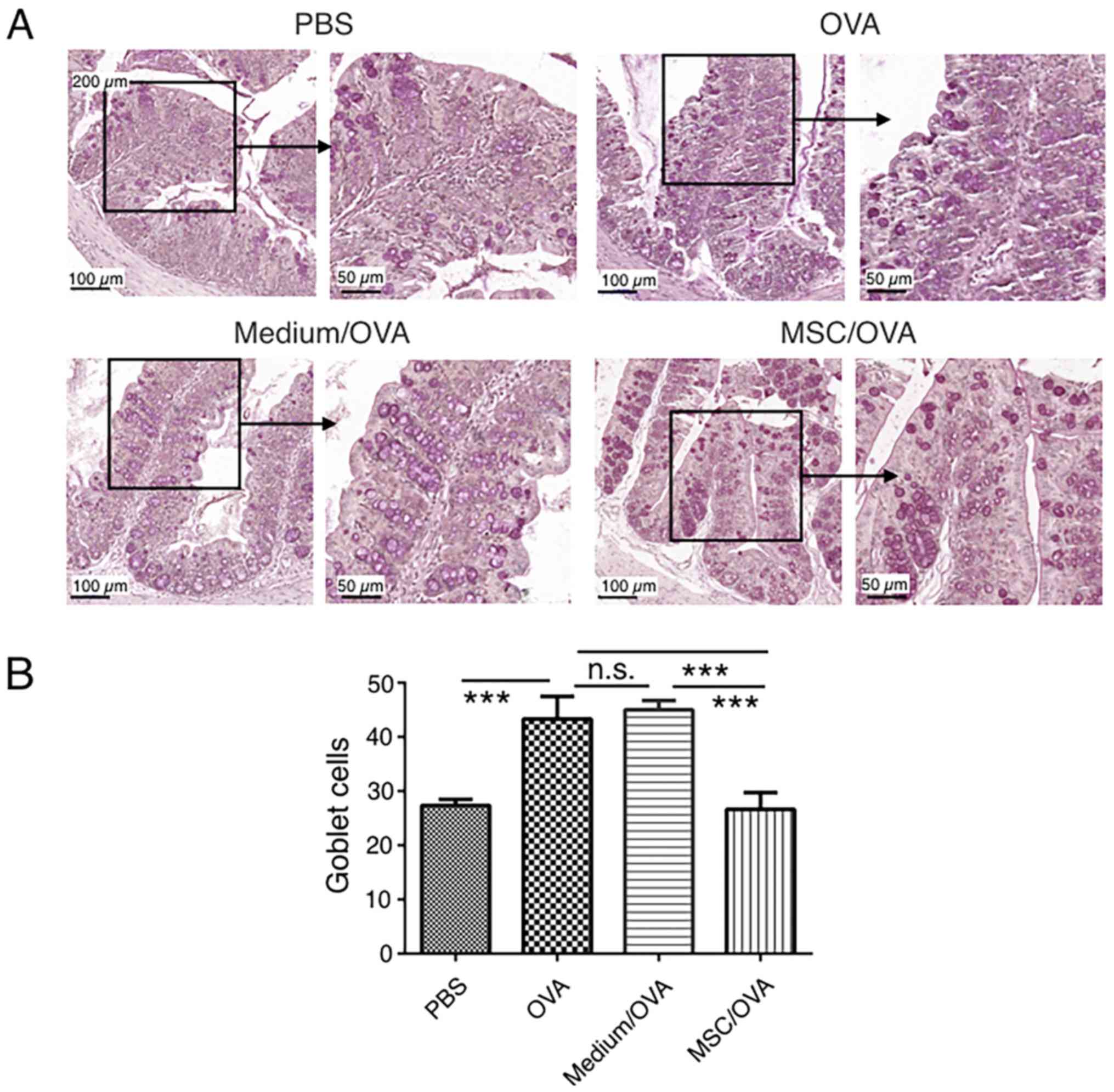 | Figure 6.Goblet cell staining in the colon.
Mice were divided into PBS, OVA, Medium/OVA and MSC/OVA treatment
groups. (A) Representative images of the goblet cells in the colon,
stained using periodic acid-Schiff stain. Scale bar, 100 and 50 µm.
(B) Quantification of the goblet cells in the fields under the
microscope. hUC-MSC, human umbilical cord-derived mesenchymal stem
cell; OVA, ovalbumin; Medium/OVA, group challenged with OVA and
administered Dulbecco's modified Eagle's medium/nutrient mixture
F12; MSC/OVAOVA/MSC, group challenged with OVA, intraperitoneal
injection of hUC-MSCs and oral gavage of MSC culture supernatant.
***P<0.001. ns, not significant. |
Treatment with hUC-MSCs recovers the
flora populations in the gut
Recently, accumulating evidence suggested that the
gut microbiota influences the development of allergic
manifestations (51). Effects of
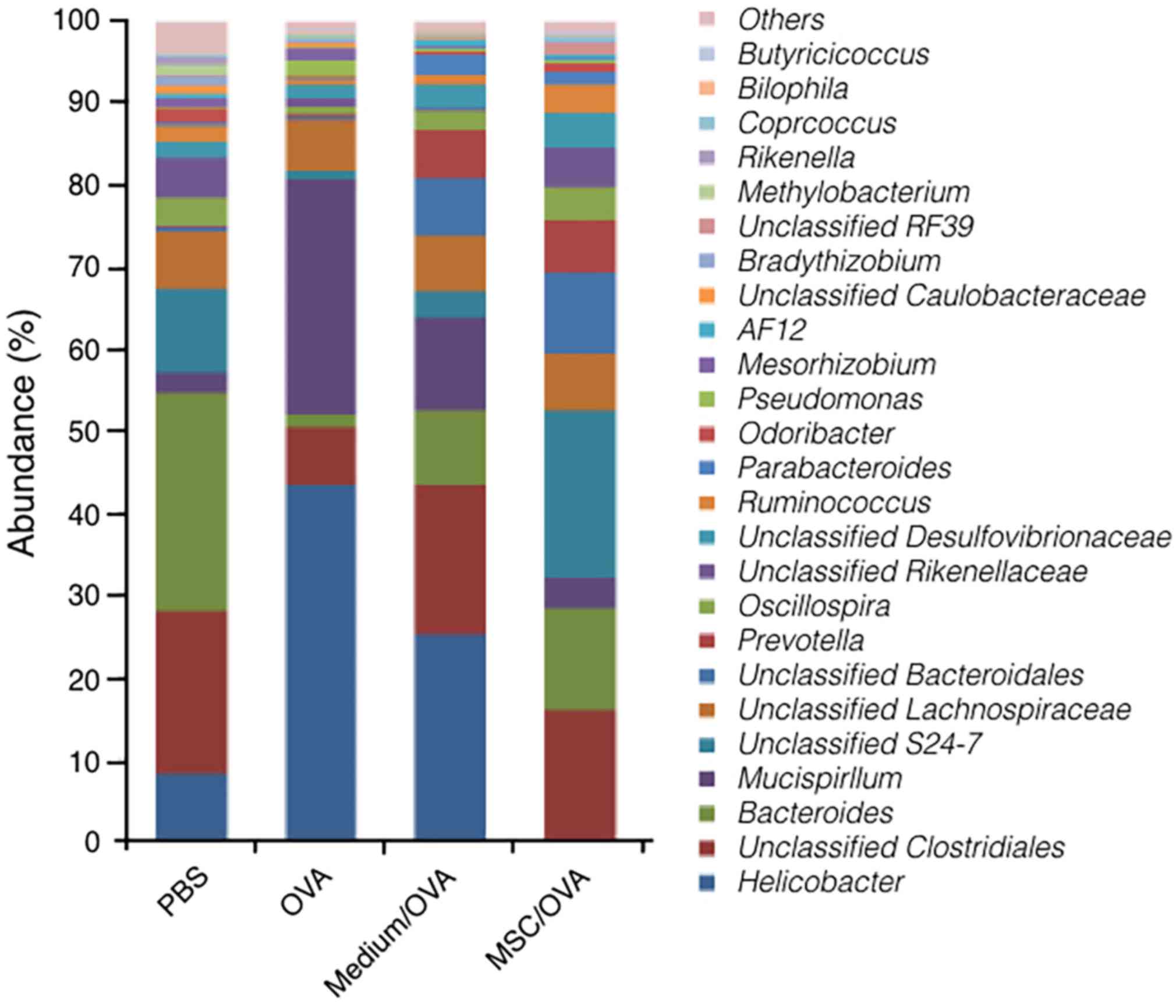
treatment with hUC-MSCs on gut microbiota populations were explored
using 16S rRNA gene sequencing. The data in Fig. 7 suggest that the population and
relative abundance of the microbiota in gut significantly changed
following OVA challenge. Following OVA challenge, the relative
abundance of the genus Helicobacter and Mucispirillum
significantly increased. At the same time, the relative abundance
of the genus Bacteroides, S24-7 and Lachnospiraceae,
which is known as butyrate-producing bacteria, significantly
decreased (52). Furthermore,
treatment with hUC-MSCs recovered the population and abundance of
commensal bacteria in the gut, particularly for S24-7 and
Lachnospiraceae. The results indicated that treatment with
hUC-MSCs may partially alleviate food allergy symptoms by
maintaining the population of gut microbiota.
Discussion
Due to their high potential expansion capacity ex
vivo, multi-lineage differentiation potential and immune
suppression functions, MSCs have emerged as attractive therapeutic
tools in transplantation, tissue regeneration and autoimmune
diseases (25). In the past decades,
MSCs were prepared from different tissues, including adipose
tissue, bone marrow, tonsils and umbilical cords, for clinical and
experimental disease therapies (21,49).
These experimental therapies include allergic diseases,
particularly experimental asthma, allergic rhinitis and allergic
contact dermatitis (25,26,53).
Compared with other tissues, the umbilical cord is an
extra-embryonic tissue and is easily obtained in large quantities
ex vivo (54). Therefore, the
umbilical cord provides a novel source of MSCs for clinical
therapy. Many different isolation and expansion procedures have
been explored to efficiently prepare hUC-MSCs ex vivo,
including enzymatic digestion and tissue explant culture (33,34,55). In
the current study, explants of umbilical cords were cultured
directly to avoid effects of enzymatic digestion on the biological
properties of the MSCs. Spindle-shaped fibroblastic cells were
observed to migrate from the tissue and adhered to the culture
dish, as previous reported (48,54).
Biological properties, including the transcriptomic profile of
hUC-MSCs were not compared with BM-MSCs. However, cells met the
basic criteria of multipotent MSCs, which are defined by the
International Society for Cellular Therapy (48). Using flow cytometry analysis, it was
observed that cells were positive for CD73, CD90 and CD105, and
negative for CD34, CD45, CD14 and HLA-DR. Additionally, these
fibroblastoid shaped cells differentiated into adipocytes and
osteoblasts in conditional differentiation medium and exhibited an
immune-suppressive function. Although, the immune suppression and
its associated mechanisms for various MSCs and MSC-conditioned
medium in autoimmune diseases have been widely explored (34,35,40,56),
effects on food allergy and on the gut microbiota population have
not been reported. In the current study, using hUC-MSC-conditioned
medium by oral gavage and hUC-MSCs by direct injection, it was
observed that treatment with hUC-MSCs alleviated allergy symptoms
and recovered the population of the gut flora. Unfortunately,
because no MSC supernatant/OVA control was utilized in the current
study, the exact role of MSC culture supernatant on allergy was
unlcear.
Food allergy in an adverse immunity to common food
and is associated with significant morbidity, particularly if
accidental ingestion is not prevented or adequately treated
(1). Previously, a number of
immunotherapeutic strategies have been investigated for the
treatment and prevention of this disease, including allergen
desensitization, anti-IgE antibodies injection for IgE blocking and
other non-antigen specific therapies (57). In the current study, it was observed
that the treatment with hUC-MSCs and MSC-conditioned medium
significantly alleviated clinical symptoms of food allergy, which
was further confirmed by histological data of the colon. As van
Halteren et al (50) first
described in 1997, this OVA intragastric challenge allergy model is
food allergen-specific, IgE-dependent and relevant to Th2
cytokines. Following OVA challenge by oral gavage six times, IgE
levels and Th2 cell percentages in the blood in OVA and Medium/OVA
groups were significantly increased in the mice compared with the
PBS group. Levels of IgE and percentage of Th2 cells in the blood
were significantly decreased in the mice treated with hUC-SCs
compared with other OVA-challenged groups. Protein levels of Th2
cytokines, including IL-4, were not determined in the blood.
However, IL-4 mRNA levels in the colon were increased in
OVA-challenged mice compared with the PBS-treated control.
Treatment with hUC-MSCs decreased these OVA-induced effects.
Published works declared that different mechanisms are involved in
the immune regulation of IL-4 expression mediated by various MSCs,
including TGF-β secretion and Treg cell differentiation (21,53,58).
Although, levels of TGF-β and Treg cells in the blood were not
detected here, the previous studies indicate that these
immune-suppression mechanisms mediated by direct injection of MSCs
may also serve potential roles in the results of the current
study.
Goblet cells are columnar epithelial cells in the
gastrointestinal, respiratory and reproductive tract. The basic
function of goblet cells is to secrete gel-forming mucins in order
to protect mucous membranes, including mucin-2 in the intestine and
mucin-5AC in the airway (59).
Generally, allergen-induced asthma promotes goblet cell metaplasia
to secrete more mucins protecting the mucosa (60). Mechanistically, published data
suggested that histamine and inflammatory cytokines, including
IL-13 and IL-7, contribute to this goblet cell hyperplasia in the
respiratory tract (61,62). In the intestinal tract, nutrition and
probiotics regulate the number of goblet cells (63). These goblet-like shaped cells further
act to preferentially deliver antigens to CD103+ DCs and
do contribution to oral tolerance in the gut (64). Similar to the goblet cell metaplasia
in asthma, an increased number of these cells in the intestinal
tract indicates its protection in food allergy (65).
Recently, by sequencing the bacterial 16S rRNA gene
to study the composition of the intestinal microbiota and the
microbial metabolism, evidence from experimental models and
clinical investigations suggested that a disturbed gut microbiota
is associated with the development of allergic manifestations
(51,66). The decreased gut microbiota richness,
the decreased Bacteroidaceae/Enterobacteriaceae ratio or a
lower relative abundance of Lachnospiraceae are associated
with sensitization to allergens in the gut or airway (67). This was further observed and
confirmed in the current study. Accumulating data demonstrated that
butyrate-producing bacteria use fiber to produce short fatty acids,
including butyrate, in order to maintain gut homeostasis and immune
tolerance by promoting differentiation of Treg and resident
CD103+ DCs (68,69). DC- or regulatory T cell-meditated
tolerance can be reprogrammed by inflammation-associated cytokines
to alter food-allergic immunity, which is mediated by Th2 cells or
type 2 innate lymphoid cells (48,54). In
an asthma model, MSCs were reported to alleviate clinical symptoms
through multiple mechanisms, including promoting Treg or inhibiting
Th1 cell differentiation, and modifying the phenotype of resident
DCs and macrophages (50,53,57,58).
However, the exact functions of the treatment with hUC-MSCs in the
current study on the population and abundance of the gut microbiota
requires further investigation. It is well known that a specific
diet may promote particular bacterial strains to grow through
altering the microbiota metabolism and drive the development of the
pathological flora microenvironment in the gut (42,70).
Additionally, published studies revealed that treatment with
induced pluripotent stem cell-derived MSCs and adipose-derived MSCs
partially restored the microbiome in mice with colitis (71). Similar to these reports, OVA by oral
gavage in the current study successfully modified the richness and
composition of the gut microbiota, which is associated with
allergies in the gut. Treatment with MSC culture supernatant by
oral gavage partially recovered the gut microbiota. The nutritional
ingredients from the MSC supernatant in the treatment group may
contribute to this restoration of the gut flora. In addition, it
has been demonstrated that immunomodulators, including
immunoglobulin and mammalian target of rapamycin inhibitors, affect
the immune regulation through directly altering the phenotype of
immune effector cells and by modifying the gut flora (72–74).
Supported by these studies, it is suggested that hUC-MSCs as strong
immune regulators may alter the gut flora. Collectively, though the
exact mechanism is yet to be elucidated, these results indicate
that treatment with hUC-MSCs may regulate the immune functions
partially through modifying the bacterial richness and composition
in the gut flora, particularly by increasing the abundance of
butyrate-producing bacteria, including S24-7,
Lachnospiraceae and Bacteroidaceae.
In conclusion, treatment with hUC-MSCs alleviated an
IgE-dependent food allergy by modifying the immune balance, which
included decreasing the IgE levels and the number of Th2 cells.
Furthermore, treatment with hUC-MSCs exhibited restorative effects
on goblet cells and commensal bacteria in the gut. Therefore, the
present study suggested that hUC-MSCs affected the regulation on
the intestinal immune system and the modification of the gut
microenvironment. hUC-MSCs may have a potential clinical
application in food allergy therapy.
Acknowledgements
The authors would like to thank the members of the
Shao and Zhou Lab (School of Medicine, Jiangsu University) for
helpful discussions and technical support.
Funding
The current study was funded by grants from the
National Natural Science Foundation of China (grant nos. 31570879
and 81871234) and the Key Research and Development Program of
Jiangsu Province, China (grant no. BE2017696).
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
NY performed experiments and analyzed the data. JX,
CZ and YW looked after the animals and performed flow cytometry.
FG, CL and WZ performed cell culturing and ELISA. TX, XZ and QS
performed tissue collections, histology and participated in
discussions. SX designed the project, analyzed the data and wrote
the manuscript. All authors read and approved the final
manuscript.
Ethics approval and consent to
participate
All the protocols were reviewed and approved by the
Ethics Committee of Jiangsu University and Animal Care and Use
Committee of Jiangsu University. Patients provided written informed
consent.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Yue D, Ciccolini A, Avilla E and Waserman
S: Food allergy and anaphylaxis. J Asthma Allergy. 11:111–120.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Sicherer SH and Leung DY: Advances in
allergic skin disease, anaphylaxis, and hypersensitivity reactions
to foods, drugs, and insects in 2010. J Allergy Clin Immunol.
127:326–335. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Parrish CP and Kim H: Food-induced
anaphylaxis: An update. Curr Allergy Asthma Rep. 18:412018.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Hill DA, Grundmeier RW, Ram G and Spergel
JM: The epidemiologic characteristics of healthcare
provider-diagnosed eczema, asthma, allergic rhinitis, and food
allergy in children: A retrospective cohort study. BMC Pediatr.
16:1332016. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Koplin JJ, Mills EN and Allen KJ:
Epidemiology of food allergy and food-induced anaphylaxis: Is there
really a western world epidemic? Curr Opin Allergy Clin Immunol.
15:409–416. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Yang Z, Zheng W, Yung E, Zhong N, Wong GW
and Li J: Frequency of food group consumption and risk of allergic
disease and sensitization in schoolchildren in urban and rural
china. Clin Exp Allergy. 45:1823–1832. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Mullins RJ, Dear KB and Tang ML: Time
trends in Australian hospital anaphylaxis admissions in 1998–1999
to 2011–2012. J Allergy Clin Immunol. 136:367–375. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Turner PJ, Gowland MH, Sharma V,
Ierodiakonou D, Harper N, Garcez T, Pumphrey R and Boyle RJ:
Increase in anaphylaxis-related hospitalizations but no increase in
fatalities: An analysis of United Kingdom national anaphylaxis
data, 1992–2012. J Allergy Clin Immunol. 135:956–963, e951. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Stone KD, Prussin C and Metcalfe DD: IgE,
mast cells, basophils, and eosinophils. J Allergy Clin Immunol. 125
Suppl 2:S73–S80. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Spergel JM: Nonimmunoglobulin e-mediated
immune reactions to foods. Allergy Asthma Clin Immunol. 2:78–85.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Castro-Sanchez P and Martin-Villa JM: Gut
immune system and oral tolerance. Br J Nutr. 109 Suppl 2:S3–S11.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Coombes JL, Siddiqui KR, Arancibia-Carcamo
CV, Hall J, Sun CM, Belkaid Y and Powrie F: A functionally
specialized population of mucosal CD103+ DCs induces Foxp3+
regulatory T cells via a TGF-beta and retinoic acid-dependent
mechanism. J Exp Med. 204:1757–1764. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Bakdash G, Vogelpoel LT, van Capel TM,
Kapsenberg ML and de Jong EC: Retinoic acid primes human dendritic
cells to induce gut-homing, IL-10-producing regulatory T cells.
Mucosal Immunol. 8:265–278. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Syed A, Garcia MA, Lyu SC, Bucayu R, Kohli
A, Ishida S, Berglund JP, Tsai M, Maecker H, O'Riordan G, et al:
Peanut oral immunotherapy results in increased antigen-induced
regulatory T-cell function and hypomethylation of forkhead box
protein 3 (FOXP3). J Allergy Clin Immunol. 133:500–510. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Wang YH: Developing food allergy: A
potential immunologic pathway linking skin barrier to gut.
F1000Res. 5:F10002016. View Article : Google Scholar
|
|
16
|
Rivas Noval M, Burton OT, Wise P,
Charbonnier LM, Georgiev P, Oettgen HC, Rachid R and Chatila TA:
Regulatory T cell reprogramming toward a Th2-cell-like lineage
impairs oral tolerance and promotes food allergy. Immunity.
42:512–523. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Turcanu V, Maleki SJ and Lack G:
Characterization of lymphocyte responses to peanuts in normal
children, peanut-allergic children, and allergic children who
acquired tolerance to peanuts. J Clin Invest. 111:1065–1072. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Nakajima-Adachi H, Ebihara A, Kikuchi A,
Ishida T, Sasaki K, Hirano K, Watanabe H, Asai K, Takahashi Y,
Kanamori Y, et al: Food antigen causes TH2-dependent enteropathy
followed by tissue repair in T-cell receptor transgenic mice. J
Allergy Clin Immunol. 117:1125–1132. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Friedenstein AJ, Petrakova KV, Kurolesova
AI and Frolova GP: Heterotopic of bone marrow. Analysis of
precursor cells for osteogenic and hematopoietic tissues.
Transplantation. 6:230–247. 1968. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Bernardo ME, Locatelli F and Fibbe WE:
Mesenchymal stromal cells. Ann N Y Acad Sci. 1176:101–117. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Cho KS, Park MK, Kang SA, Park HY, Hong
SL, Park HK, Yu HS and Roh HJ: Adipose-derived stem cells
ameliorate allergic airway inflammation by inducing regulatory T
cells in a mouse model of asthma. Mediators Inflamm.
2014:4364762014. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
de Aguiar CF, Castoldi A, Andrade-Oliveira
V, Ignacio A, da Cunha FF, Felizardo RJF, Bassi ÊJ, Câmara NOS and
de Almeida DC: Mesenchymal stromal cells modulate gut inflammation
in experimental colitis. Inflammopharmacology. 26:251–260. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Ryan JM, Barry FP, Murphy JM and Mahon BP:
Mesenchymal stem cells avoid allogeneic rejection. J Inflamm
(Lond). 2:82005. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Fujii S, Miura Y, Fujishiro A, Shindo T,
Shimazu Y, Hirai H, Tahara H, Takaori-Kondo A, Ichinohe T and
Maekawa T: Graft-versus-host disease amelioration by human bone
marrow mesenchymal stromal/stem cell-derived extracellular vesicles
is associated with peripheral preservation of naive t cell
populations. Stem Cells. 36:434–445. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Uccelli A, Moretta L and Pistoia V:
Mesenchymal stem cells in health and disease. Nat Rev Immunol.
8:726–736. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Ren G, Zhang L, Zhao X, Xu G, Zhang Y,
Roberts AI, Zhao RC and Shi Y: Mesenchymal stem cell-mediated
immunosuppression occurs via concerted action of chemokines and
nitric oxide. Cell Stem Cell. 2:141–150. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Ghannam S, Pene J, Moquet-Torcy G,
Jorgensen C and Yssel H: Mesenchymal stem cells inhibit human Th17
cell differentiation and function and induce a T regulatory cell
phenotype. J Immunol. 185:302–312. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Beyth S, Borovsky Z, Mevorach D,
Liebergall M, Gazit Z, Aslan H, Galun E and Rachmilewitz J: Human
mesenchymal stem cells alter antigen-presenting cell maturation and
induce T-cell unresponsiveness. Blood. 105:2214–2219. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Spaggiari GM, Abdelrazik H, Becchetti F
and Moretta L: MSCs inhibit monocyte-derived DC maturation and
function by selectively interfering with the generation of immature
DCs: Central role of MSC-derived prostaglandin E2. Blood.
113:6576–6583. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Li T, Xia M, Gao Y, Chen Y and Xu Y: Human
umbilical cord mesenchymal stem cells: An overview of their
potential in cell-based therapy. Expert Opin Biol Ther.
15:1293–1306. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Li T, Yan Y, Wang B, Qian H, Zhang X, Shen
L, Wang M, Zhou Y, Zhu W, Li W and Xu W: Exosomes derived from
human umbilical cord mesenchymal stem cells alleviate liver
fibrosis. Stem Cells Dev. 22:845–854. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Weiss ML, Anderson C, Medicetty S,
Seshareddy KB, Weiss RJ, VanderWerff I, Troyer D and McIntosh KR:
Immune properties of human umbilical cord Wharton's jelly-derived
cells. Stem Cells. 26:2865–2874. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Xie Z, Hao H, Tong C, Cheng Y, Liu J, Pang
Y, Si Y, Guo Y, Zang L, Mu Y and Han W: Human umbilical
cord-derived mesenchymal stem cells elicit macrophages into an
anti-inflammatory phenotype to alleviate insulin resistance in type
2 diabetic rats. Stem Cells. 34:627–639. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Kim HS, Shin TH, Lee BC, Yu KR, Seo Y, Lee
S, Seo MS, Hong IS, Choi SW, Seo KW, et al: Human umbilical cord
blood mesenchymal stem cells reduce colitis in mice by activating
NOD2 signaling to COX2. Gastroenterology. 145(1392–1403):
e1391–e1398. 2013.
|
|
35
|
Sun L, Wang D, Liang J, Zhang H, Feng X,
Wang H, Hua B, Liu B, Ye S, Hu X, et al: Umbilical cord mesenchymal
stem cell transplantation in severe and refractory systemic lupus
erythematosus. Arthritis Rheum. 62:2467–2475. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Qiao C, Xu W, Zhu W, Hu J, Qian H, Yin Q,
Jiang R, Yan Y, Mao F, Yang H, et al: Human mesenchymal stem cells
isolated from the umbilical cord. Cell Biol Int. 32:8–15. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Yen BL, Huang HI, Chien CC, Jui HY, Ko BS,
Yao M, Shun CT, Yen ML, Lee MC and Chen YC: Isolation of
multipotent cells from human term placenta. Stem Cells. 23:3–9.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Paula-Silva J, Santiago AF, Oliveira RP,
Rosa ML, Carvalho CR, Amaral JF and Faria AM: Effect of a
protein-free diet in the development of food allergy and oral
tolerance in BALB/c mice. Br J Nutr. 113:935–943. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Kay AG, Long G, Tyler G, Stefan A,
Broadfoot SJ, Piccinini AM, Middleton J and Kehoe O: Mesenchymal
stem cell-conditioned medium reduces disease severity and immune
responses in inflammatory arthritis. Sci Rep. 7:180192017.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Pouya S, Heidari M, Baghaei K, Aghdaei
Asadzadeh H, Moradi A, Namaki S, Zali MR and Hashemi SM: Study the
effects of mesenchymal stem cell conditioned medium injection in
mouse model of acute colitis. Int Immunopharmacol. 54:86–94. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Nagata Y, Yamamoto T, Hayashi M, Hayashi S
and Kadowaki M: Improvement of therapeutic efficacy of oral
immunotherapy in combination with regulatory T cell-inducer
kakkonto in a murine food allergy model. PLoS One. 12:e01705772017.
View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Cheng L, Jin H, Qiang Y, Wu S, Yan C, Han
M, Xiao T, Yan N, An H, Zhou X, et al: High fat diet exacerbates
dextran sulfate sodium induced colitis through disturbing mucosal
dendritic cell homeostasis. Int Immunopharmacol. 40:1–10. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Parada AE, Needham DM and Fuhrman JA:
Every base matters: Assessing small subunit rRNA primers for marine
microbiomes with mock communities, time series and global field
samples. Environ Microbiol. 18:1403–1414. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Metcalf JL, Xu ZZ, Weiss S, Lax S, Van
Treuren W, Hyde ER, Song SJ, Amir A, Larsen P, Sangwan N, et al:
Microbial community assembly and metabolic function during
mammalian corpse decomposition. Science. 351:158–162. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Ravussin Y, Koren O, Spor A, LeDuc C,
Gutman R, Stombaugh J, Knight R, Ley RE and Leibel RL: Responses of
gut microbiota to diet composition and weight loss in lean and
obese mice. Obesity (Silver Spring). 20:738–747. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Lindner U, Kramer J, Rohwedel J and
Schlenke P: Mesenchymal stem or stromal cells: Toward a better
understanding of their biology? Transfus Med Hemother. 37:75–83.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Dominici M, Le Blanc K, Mueller I,
Slaper-Cortenbach I, Marini F, Krause D, Deans R, Keating A,
Prockop Dj and Horwitz E: Minimal criteria for defining multipotent
mesenchymal stromal cells. The international society for cellular
therapy position statement. Cytotherapy. 8:315–317. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Gazit Z, Pelled G, Sheyn D, Kimelman N and
Gazit D: Mesenchymal Stem CellsEssentials of Stem Cell Biology
(third Edition). Lanza R and Atala A: Academic Press; Boston: pp.
255–266. 2014, View Article : Google Scholar
|
|
50
|
van Halteren AG, van der Cammen MJ,
Biewenga J, Savelkoul HF and Kraal G: IgE and mast cell response on
intestinal allergen exposure: A murine model to study the onset of
food allergy. J Allergy Clin Immunol. 99:94–99. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Sjodin Simonyte K, Vidman L, Ryden P and
West CE: Emerging evidence of the role of gut microbiota in the
development of allergic diseases. Curr Opin Allergy Clin Immunol.
16:390–395. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Vital M, Howe AC and Tiedje JM: Revealing
the bacterial butyrate synthesis pathways by analyzing
(meta)genomic data. MBio. 5:e008892014. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Nemeth K, Keane-Myers A, Brown JM,
Metcalfe DD, Gorham JD, Bundoc VG, Hodges MG, Jelinek I, Madala S,
Karpati S and Mezey E: Bone marrow stromal cells use TGF-beta to
suppress allergic responses in a mouse model of ragweed-induced
asthma. Proc Natl Acad Sci USA. 107:5652–5657. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Majore I, Moretti P, Stahl F, Hass R and
Kasper C: Growth and differentiation properties of mesenchymal
stromal cell populations derived from whole human umbilical cord.
Stem Cell Rev. 7:17–31. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Phillips CD, Wongsaisri P, Htut T and
Grossman T: Purified umbilical cord derived mesenchymal stem cell
treatment in a case of systemic lupus erythematosus. Clin Transl
Med. 6:312017. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Robinson AM, Sakkal S, Park A, Jovanovska
V, Payne N, Carbone SE, Miller S, Bornstein JC, Bernard C, Boyd R
and Nurgali K: Mesenchymal stem cells and conditioned medium avert
enteric neuropathy and colon dysfunction in guinea pig TNBS-induced
colitis. Am J Physiol Gastrointest Liver Physiol. 307:G1115–G1129.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Nowak-Wegrzyn A and Sampson HA: Future
therapies for food allergies. J Allergy Clin Immunol. 127:558–573;
quiz 574–555. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Miyagawa I, Nakayamada S, Nakano K,
Yamagata K, Sakata K, Yamaoka K and Tanaka Y: Induction of
regulatory T cells and its regulation with insulin-like growth
factor/insulin-like growth factor binding protein-4 by human
mesenchymal stem cells. J Immunol. 199:1616–1625. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Rubin BK: Secretion properties, clearance,
and therapy in airway disease. Transl Respir Med. 2:62014.
View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Lambrecht BN and Hammad H: The immunology
of asthma. Nat Immunol. 16:45–56. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Xia W, Bai J, Wu X, Wei Y, Feng S, Li L,
Zhang J, Xiong G, Fan Y, Shi J and Li H: Interleukin-17A promotes
MUC5AC expression and goblet cell hyperplasia in nasal polyps via
the Act1-mediated pathway. PLoS One. 9:e989152014. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Kuperman DA and Schleimer RP:
Interleukin-4, interleukin-13, signal transducer and activator of
transcription factor 6, and allergic asthma. Curr Mol Med.
8:384–392. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Dock-Nascimento DB, Junqueira K and
Aguilar-Nascimento JE: Rapid restoration of colonic goblet cells
induced by a hydrolyzed diet containing probiotics in experimental
malnutrition. Acta Cir Bras. 1 Suppl 22:S72–S76. 2007. View Article : Google Scholar
|
|
64
|
McDole JR, Wheeler LW, McDonald KG, Wang
B, Konjufca V, Knoop KA, Newberry RD and Miller MJ: Goblet cells
deliver luminal antigen to CD103+ dendritic cells in the small
intestine. Nature. 483:345–349. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Yamaki K and Yoshino S: Remission of food
allergy by the Janus kinase inhibitor ruxolitinib in mice. Int
Immunopharmacol. 18:217–224. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Clooney AG, Fouhy F, Sleator RD,
O'Driscoll A, Stanton C, Cotter PD and Claesson MJ: Comparing
apples and oranges?: Next generation sequencing and its impact on
microbiome analysis. PLoS One. 11:e01480282016. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Arrieta MC, Stiemsma LT, Dimitriu PA,
Thorson L, Russell S, Yurist-Doutsch S, Kuzeljevic B, Gold MJ,
Britton HM and Lefebvre DL: Early infancy microbial and metabolic
alterations affect risk of childhood asthma. Sci Transl Med.
7:307ra1522015. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Yamashiro Y: Gut microbiota in health and
disease. Ann Nutr Metab. 71:242–246. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Furusawa Y, Obata Y, Fukuda S, Endo TA,
Nakato G, Takahashi D, Nakanishi Y, Uetake C, Kato K, Kato T, et
al: Commensal microbe-derived butyrate induces the differentiation
of colonic regulatory T cells. Nature. 504:446–450. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Bibbo S, Ianiro G, Giorgio V, Scaldaferri
F, Masucci L, Gasbarrini A and Cammarota G: The role of diet on gut
microbiota composition. Eur Rev Med Pharmacol Sci. 20:4742–4749.
2016.PubMed/NCBI
|
|
71
|
Soontararak S, Chow L, Johnson V, Coy J,
Wheat W, Regan D and Dow S: Mesenchymal stem cells (MSC) derived
from induced pluripotent stem cells (iPSC) equivalent to
adipose-derived MSC in promoting intestinal healing and microbiome
normalization in mouse inflammatory bowel disease model. Stem Cells
Transl Med. 7:456–467. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Hurez V, Dao V, Liu A, Pandeswara S,
Gelfond J, Sun L, Bergman M, Orihuela CJ, Galvan V, Padrón Á, et
al: Chronic mTOR inhibition in mice with rapamycin alters T, B,
myeloid, and innate lymphoid cells and gut flora and prolongs life
of immune-deficient mice. Aging Cell. 14:945–956. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Henderson AL, Brand MW, Darling RJ, Maas
KJ, Detzel CJ, Hostetter J, Wannemuehler MJ and Weaver EM:
Attenuation of colitis by serum-derived bovine
immunoglobulin/protein isolate in a defined microbiota mouse model.
Dig Dis Sci. 60:3293–3303. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Carbonnel F, Soularue E, Coutzac C, Chaput
N, Mateus C, Lepage P and Robert C: Inflammatory bowel disease and
cancer response due to anti-CTLA-4: Is it in the flora? Semin
Immunopathol. 39:327–331. 2017. View Article : Google Scholar : PubMed/NCBI
|