Asthma is a heterogeneous disease that is usually
characterized by chronic airway inflammation with airway
hyper-responsiveness, airway remodelling and disordered mucosal
immunity (1–3). Most asthma-associated deaths occur in
low- and low-middle-income countries. According to the latest World
Health Organization estimates released in December 2016,
approximately 235 million individuals currently suffer from asthma,
and 383,000 deaths occurred due to asthma in 2015 (4). The strongest risk factors for
developing asthma are exposure to inhaled substances and particles,
such as pollen and house dust mites (HDM), which may provoke
allergic reactions or irritate the airways (5,6).
Patients with asthma suffer recurrent episodes of wheezing,
coughing, chest tightness and shortness of breath. These episodes
are usually associated with airflow obstruction within the lung,
which is often reversible either spontaneously or with treatment
(7). Additionally, the specific
pathogenesis of asthma remains unclear. Therefore, the
investigation of potential molecular mechanisms will improve our
understanding of the pathogenesis of asthma and help us to identify
new effective therapeutic targets.
Neuromedin U (NMU) is a multifunctional neuropeptide
with pleiotropic effects, including the mediation of intestinal
peristalsis and the modulation of the sense of satiety, body
weight, circadian oscillation, bone formation, insulin production,
cancer development, energy balance and metabolism (8–12).
However, these effects will not be addressed in the present review.
Recently, reports have demonstrated that the neuropeptide NMU
enhances ILC2-driven allergic lung inflammation (13–15).
Therefore, the effect of NMU on the pathogenesis of asthma will be
evaluated in the present review.
Asthma is an airway disease and is characterized by
four treatable traits: Airflow limitation, altered cough reflex
sensitivity, airway infection and airway inflammation (1,16)
(Table I). Airflow limitation is
caused by several factors, including the sensitization of airway
nerves, the accumulation of mast cells, the repeated obstruction of
airway smooth muscle, inflammatory mural oedema, the decreased
production of bronchoprotective factors and structural changes to
the airway (17,18). Airway hyper-responsiveness is another
important factor in the development of asthma and is a result of an
imbalance in the autonomic nervous system. It involves various
inflammatory cells, mediators and cytokines, damage to the
epithelial airway, and the exposure of the subepithelial nerve
terminals (19). Thirdly, viral
infections aggravate airway inflammation in asthma and can even
lead to asthma attacks (20).
Finally, airway inflammation is heterogeneous among asthma
patients. Eosinophilic airway inflammation is the predominant type
of granulocytic inflammation because it is recognizable and
treatable (21). In patients with
eosinophilic asthma, there are two different pathogenic pathways
(22). In allergic eosinophilic
airway inflammation, specialized dendritic cells present allergens
to steer the differentiation of naive T lymphocytes towards the
formation of Th2 cells, which produce cytokines such as
interleukin-4 (IL-4), IL-5, IL-9 and IL-13 that result in IgE
switching in B cells (23,24). The released IgE molecules bind to the
Fc epsilon receptor I (FcεRI) on mast cell surfaces (25). Exposure to allergens and the
cross-linking of receptors by allergens, which bind to
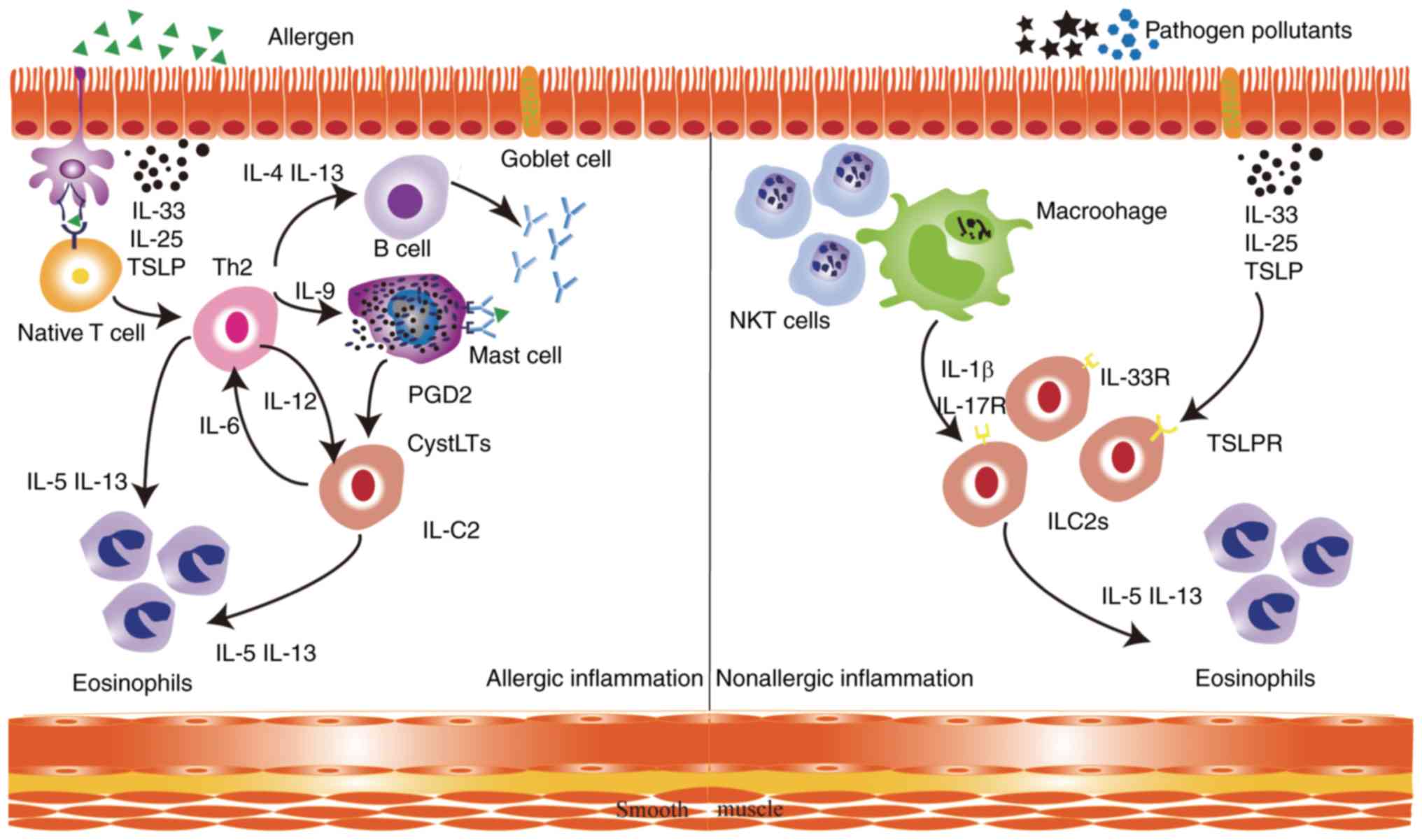
high-affinity IgE result in mast cell degranulation (26). Type 2 innate lymphoid cells (ILC2s)
are activated by cysteinyl leukotrienes (CysLTs) and prostaglandin
D2 (PGD2) secreted by activated mast cells (27). Once activated, ILC2s rapidly expand
and secrete large amounts of IL-5 and IL-13. ILC2s contribute to
allergic airway inflammation by directly interacting with Th2 cells
to promote the release of cytokines, mucus production and airway
eosinophilia (28–30). In nonallergic eosinophilic asthma,
air pollutants and pathogens induce the release of
epithelium-derived and macrophage-derived cytokines, including
IL-33, IL-25, TSLP and IL-1β, which activate ILC2s in an
antigen-independent manner via their respective receptors (Fig. 1) (31,32). It
was previously shown that IL-13 released by ILC2 disrupts the
bronchial barrier integrity of the airway epithelium in asthmatic
patients, therefore maintaining type 2 inflammation in the airways
(33). In summary, the role of ILC2s
in airway epithelium has received more attention.
In the treatment of asthma, inhaled
glucocorticosteroids are currently the most effective
anti-inflammatory medications for the therapy of persistent asthma.
Their efficacy in decreasing airway hyper-responsiveness has been
demonstrated, which decreases asthma symptoms and controls airway
inflammation (34). Additionally,
long-acting inhaled β2-agonists are more effective when combined
with inhaled glucocorticosteroids (35). However, long-term exposure to high
doses of inhaled glucocorticosteroids leads to adrenal suppression,
easy bruising and decreased bone mineral density (36–38). The
key role of IL-5 in eosinophilic airway inflammation make it a
crucial drug target, which has driven the clinical development of
two monoclonal antibodies against IL-5, reslizumab and mepolizumab
(39). Nevertheless, these
antibodies are ineffective for non-phenotyped patients with
persistent asthma who have already received treatment with inhaled
corticosteroids (Table I) (22).
Asthma is an inflammatory airway disease involving a
variety of cells and cytokines. Due to the phenotypic heterogeneity
of asthma, its pathogenesis is complex; the treatable traits are
not separate, but are rather interrelated. For this reason, airway
disease emerges as an intractable disease, without the development
of novel therapies. NMU acts as a multifunctional neuropeptide in
allergic responses. In the following sections, the discovery,
distribution and function of NMU are to be discussed. Furthermore,
the role of NMU in allergic airway inflammation and its potential
for clinical application will be discussed.
NMU, which was first isolated from porcine spinal
cord and named for its potent contractile effect on the rat uterus
in 1985, is a highly conserved peptide secreted by cholinergic
neurons (40). NMU was found in
rabbits, dogs, frogs and chickens; a 23-amino-acid version was
identified in rats, and nonapeptides were detected in guinea pigs
and chickens (41–43). NMU is therefore widely conserved
throughout the animal kingdom and shows almost complete
conservation of its amidated C-terminal pentapeptide, indicating
that there is a strong evolutionary pressure to conserve this
peptide. NMU also has widespread distribution in the peripheral and
central nervous system (9,44). NMU-like immunoreactivity (NMU-LI)
protein and mRNA are distributed in the stomach, ileum, spleen,
pancreas, heart, lung, kidney, prostate, pituitary gland, adipose
tissue, bone, bone marrow and lymphocytes in humans (45,46).
NMU-LI is also widely distributed in the central nervous system,
including the cingulate gyrus, thalamus, locus coeruleus, medulla
oblongata, hypothalamus, substantia nigra and medial frontal gyrus
in humans (45,46). Due to the high affinity of NMU and
the saturable and specific binding sites for NMU-23 in rats, NMU
has been previously characterized as a cognate ligand for the
designated ‘orphan’ class A G-protein-coupled receptors (GPCRs)
(47). Two different receptors exist
for NMU, termed NMUR1 (also known as GPR66 and FM-3) and NMUR2
(also known as TGR-1 and FM-4), which are encoded by genes located
in human chromosomes 2 and 5, respectively (48–50).
NMUR1 is mainly expressed in peripheral tissues, such as the
intestine, pancreas, uterus, lung and kidney. NMUR2 is
predominantly found in specific regions of the central nervous
system, including the spinal cord, dorsal root ganglia and medulla
oblongata (51). In the rat spinal
cord, NMU-like immunoreactivity protein levels are greater in the
dorsal than in the ventral horn, suggesting a sensory role for NMU
(41). More recently, using chimeric
G proteins, it was demonstrated that NMUR1 primarily signals
through the Gq/11 proteins, whereas NMUR2 signals through the Gi
proteins (50,52,53).
Moreover, NMU mRNA has been detected in antigen-presenting cells,
particularly monocytes and dendritic cells, and NMUR1 mRNA was
detected in T cells and natural killer cells (54,55).
Therefore, NMU has the potential to serve as a target for the
treatment of asthma.
Thus, NMU is a multifunctional neuropeptide with
several roles in different cells and tissue types, which relays
signals to the central nervous system (CNS) to stimulate organs.
NMU also affects cells directly, by increasing cell proliferation
and migration, and inducing the release of hormones and
autocrine/paracrine factors. These functions are mostly mediated
via the NMUR1 and NMUR2 receptors, although other alternative
receptors have been described. It is also possible that NMU may
also elicit its effects by binding and signalling through unknown
receptors. Furthermore, NMU may also serve as an ideal target for
the treatment of certain disorders, although its multiple roles
should be taken into consideration when inhibiting its functions
for therapeutic purposes (10).
ILC2s have recently been identified as effector
cells that are key early regulators of immune responses in airway
barrier surfaces (33). ILC2s are
activated by cell-derived exogenous cytokines, such as IL-25, IL-33
and thymic stromal lymphopoietin (TSLP). Activated ILC2s produce
type 2 cytokines, such as IL-5 and IL-13, to initiate allergic
inflammation at mucosal surfaces (28,56).
However, the specific molecular pathways that modulate the response
of ILC2s to alarmins such as IL-25, IL-33 and TSLP remain unclear
(29). Notably, NMUR1 is largely
specific to ILC2s, according to single-cell RNA sequencing and flow
cytometry (13). Moreover, it is
highly expressed in ILC2s at baseline and following the induction
of airway inflammation with HDM, in contrast with its expression in
other lung-resident cell populations. Furthermore, NMU expands
IL-25-driven inflammation. IL-25 combined with NMU led to increased
expression of IL-5 and IL-13 in the lung and bronchoalveolar lavage
fluid, whereas IL-25 alone only modestly increased their
expression. NMU combined with IL-25 alters a non-pathologic dose of
IL-25 into a pathogenic dose. However, the number of
ST2+ ILCs following HDM challenge was decreased in
NMU-knockout mice compared with the number in wild-type mice, which
indicates that NMU promotes ILC activation and effector function.
Moreover, in NMUR1-knockout mice, ILC2 frequency was markedly
reduced after HDM challenge compared to that induced by PBS,
reflecting the effects also observed in wild-type mice (13). In addition, NMUR1-knockout ILC2s
demonstrated lower average inflammatory score than wild-type ILC2s
following HDM challenge, which is consistent with the fact that
NMU-NMUR1 signalling promotes ILC2 responses in vivo
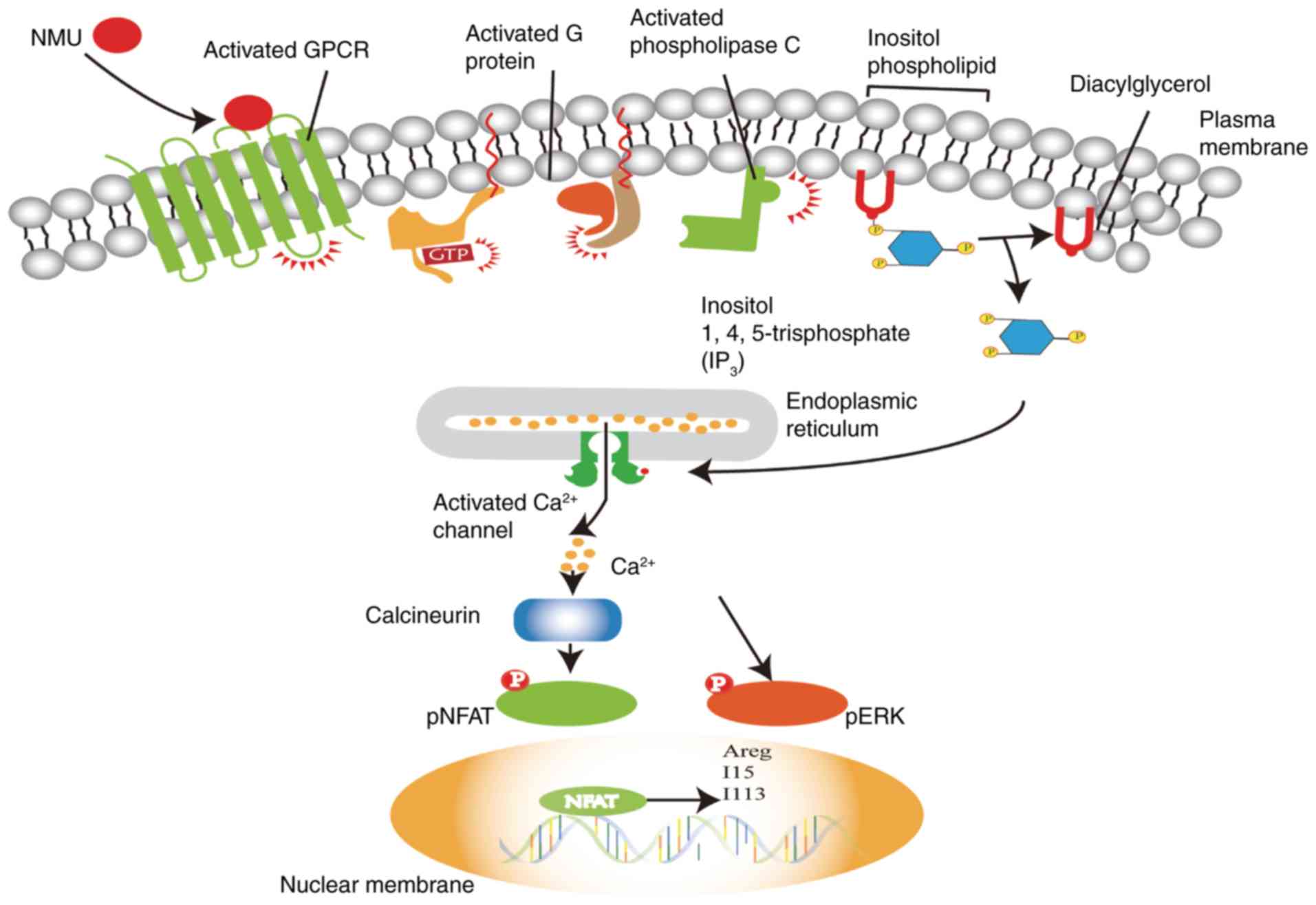
(13–15). In regard to the signalling pathway,
through which NMU activates ILC2s, it was shown that NMU activates
phospholipase C (PLC), which catalyses the conversion of the
phospholipid inositol to diacylglycerol (DAG) and inositol
1,4,5-trisphosphate (IP3). Subsequently, IP3 induces
Ca2+ release from intracellular stores (55). Cardoso et al (14) found that NMU triggered extracellular
signal-regulated kinase (ERK) phosphorylation and regulated innate
type-2 cytokines downstream of a Ca2+/calcineurin/NFAT
cascade, leading to the expression of the type 2 cytokine genes
IL-5, IL-13 and amphiregulin (Areg) in ILC2s (Fig. 2), (57). Thus, the neuropeptide NMU can
activate NMUR1 in an ILC2-mediated manner, resulting in the
uniquely potent and immediate production of innate type 2
cytokines.
Mast cells are important participants in the early
stage of allergic inflammation and are derived from haematopoietic
stem cells. Mast cells affect intercellular communication during
inflammation by secreting cytokines that contain several mediators
(58). In addition, the severity of
airway hyperresponsiveness was strongly correlated with mast cell
activation (59). IgE binds to FcεRI
on mast cells and is crosslinked with specific antigens on the cell
surface, which can induce mast cell degranulation and the synthesis
of chemokines and cytokines (60).
Specifically, mast cells proximal to nerve fibres contain, secrete
and respond to several neuropeptides (61). NMUR1 is also highly expressed in mast
cells. NMU combines with NMUR1, resulting in mast cell
degranulation and the subsequent release of preformed mediators,
most notably histamine and chemokines, which leads to early-phase
inflammation characterized by vasodilation, extravasation, edema
and smooth muscle contraction. Neutrophils also aggregate at
reaction sites as a result of the cytokines released by mast cells.
The levels of NMU-induced Ca2+ release and mast cell
degranulation are nearly comparable with those induced by IgE
receptor cross-linking. In contrast, the subsequent infiltration of
neutrophils is completely inhibited in NMU-deficient mice (62). NMU activates mast cell-mediated
inflammation; therefore, NMU receptor antagonists could be novel
targets for the pharmacological inhibition of mast cell-mediated
inflammatory diseases.
The activation of mast cells by NMU at an early
stage might trigger airway inflammation and the release of
eosinophilic chemotactic factors, which attract activated
eosinophils during allergen exposure and lead to progressive
allergic inflammation (63,64). NMUR1 is also expressed in a mouse
eosinophil cell line. NMU elevates intracellular Ca2+
levels and ERK phosphorylation and activation, which promotes cell
adhesion to components of the extracellular matrix (ECM), and
eotaxins contribute to eosinophil accumulation. Inflammatory cells
adhere and interact with components of the blood cell wall and the
ECM, which aids their ability to extravasate and migrate into
inflamed sites (65). NMU acts
directly on eosinophils to play an important role in cell
activation, adhesion and migration. On the contrary, it was
observed that the absence of Gq signalling in Gq-deficient mice
blocked the accumulation of eosinophils in the lungs, following an
allergic challenge (15,66). NMU directly acts on eosinophils to
induce cell adhesion to fibronectin and collagen type I.
Additionally, eosinophil chemotaxis is induced by NMU at high
concentration that is comparable to that induced by eotaxin, which
is also known to be involved in integrin activation and the
adhesion of eosinophils. Thus, NMU is an important mediator of
eosinophil-mediated inflammation, and a potential therapeutic
target for bronchial asthma/eosinophil-mediated inflammatory
diseases (67). To date, only
limited data have been obtained regarding the possible importance
of NMU and eosinophils. Therefore, the association between NMU and
eosinophils requires further study.
Evolution has generated multiple mechanisms to
defend against external and internal sources of danger. For
example, the immune system eliminates various threats through a
variety of immune cells and antibodies. The nervous system promptly
inputs information into the CNS and produces complex defence
behaviours. The immune system and the nervous system cooperate with
each other to defend against danger. In this, they share a common
‘language’ comprised of cytokines, receptors, and neuropeptides,
which enable mutual communication (68). Immune cells are found in close
proximity to the nerve terminal processes in the mucosal surfaces
of the airways, which are then poised for interaction (23). The lung is extensively innervated via
sensory fibres, most of which express markers of nociceptors
(69). Asthmatic patients have a
denser network of these fibres around small airways and a low
threshold for their activation in response to airborne irritants
(70,71). This indicates the excess activity of
peptidergic sensory fibres. Upon exposure to allergens, nociceptor
peripheral terminals release neuropeptides, such as substance P and
calcitonin gene-related peptide (CGRP), resulting in neuroimmunity
(68,72,73).
Furthermore, NMU has been found in the spinal cord, dorsal root
ganglia and medulla oblongata via radioimmunoassays and
immunohistochemistry (41,74). The distribution of NMU is consistent
with that of neurons involved in nociception. One of the
physiological roles of NMU may be its involvement in nociception
(75). Neurons that secret NMU can
be found in the ventromedial hypothalamic regions and in some
nuclei of the caudal brainstem regions, which are involved in
nociceptive transmission and pain modulation (44,45,76). NMU
has been demonstrated to markedly and selectively enhance the
excitability of nociceptive neurons in spinal dorsal horns in a
dose-dependent manner (77).
Hyperactivity of nociceptive dorsal horn neurons induced by NMU
could mediate pain-associated behavioural changes and several
neuroendocrine functions (49). This
expression pattern of NMUR2 mRNA corroborated with the hypothesis
that its ligand, NMU, is a sensory transmitter/modulator (77). Compared with the wild-type mice, the
nociceptive reflexes were decreased in the NMU KO mice, indicating
that endogenous NMU may play an important role in reflexes and in
adaptation to environmental stimuli (78). The mRNA expression NMU is increased
in the spinal cord but not in the hypothalamus following a pain
stimulus, suggesting that pain may stimulate the synthesis of NMU.
Thus, NMU is involved in nociceptive reflexes (78). Nociceptor activation upon allergen
exposure is a very early event in the development of inflammation.
Appreciation of the immune and nervous systems as part of a
holistic, coordinated defence system provides new insights into
inflammation and exciting opportunities for managing acute and
chronic inflammatory diseases (73).
In short, these studies suggest that NMU is important for
nociceptive reflexes and allergen exposure, although more detailed
genetic and mechanistic investigations of NMU and its role in
vivo and in vitro are still required.
In view of the importance of allergic airway
inflammation in the clinical manifestations of allergic diseases,
it is of note that NMU can induce immune cell-driven inflammation.
Upon inhaling pollutants, microbes and glycolipids, the nervous
system rapidly processes information and triggers these processes.
NMU acts as a mediator between sensory neurons and immune cells to
potentiate or initiate inflammation. Overall, NMU provides a novel
neuroimmune target for the treatment of asthma. At present, there
are no investigative reports regarding the development of NMUR
subtype-selective antagonists. The natural products EUK2010,
EUK2011 and EUK2012 have been identified as NMUR2-specific
agonists, and icariin from Herba epimedii has been
described as an NMUR2-selective agonist (79,80).
Structure-activity relationship study identified the more potent
hexapeptide 5d that exhibits NMUR1 agonist activity similar to that
of Hnmu (81). Moreover, scientists
have discovered two synthetic low molecular weight non-selective
NMU receptor agonists (45).
Regarding asthma therapy based on the function of
NMU, NMU amplifies allergic airway inflammation in an asthma model.
This indicates that NMU might be a meaningful therapeutic target
for the treatment of allergic airway inflammation in asthma. For
example, ketotifen, as a mast cell membrane stabilizer, can protect
mast cell membranes to decrease membrane metamorphosis and the
release of allergic inflammation mediators (82). Additionally, butyrate ameliorates
allergic airway inflammation by limiting eosinophil trafficking and
survival (83). Considering the
constant progress in pharmaceutical development and molecular
biology, screening for novel molecules that act on targets in the
human airway and immune cells will be conducted, leading to new
treatments for asthma.
In the context of the complexity and intricacy of
airway inflammation, NMU acts as a multifunctional neuropeptide in
the pathogenesis of asthma. An understanding of the function of NMU
aids in improving the understanding of the mechanism underlying the
pathogenesis of asthma. Considering the constant improvements in
organoid culture systems and transcriptomic techniques, NMU
receptor antagonists will likely be a novel target for the
pharmacological inhibition of asthma in the near future, which may
significantly improve the clinical outcomes of patients with
asthma. However, the precise regulation of NMU in asthma still
requires further study, in order to be used in clinical
applications.
Not applicable.
This work was supported by grants from the National
Natural Science Foundation of China (grant nos. 81673922, 81503663
and 81704167) and the Project of Educational Commission of
Guangdong Province of China (grant no. 2018KTSCX037).
Not applicable.
XR participated in the entire review process and
prepared the manuscript. FD and YZ contributed to collecting the
relevant literature. YW and WM conceived the review and modified
the manuscript. All authors read and approved the final version of
the manuscript.
Not applicable.
Not applicable.
The authors declare that they have no competing
interests.
|
1
|
Pavord ID, Beasley R, Agusti A, Anderson
GP, Bel E, Brusselle G, Cullinan P, Custovic A, Ducharme FM, Fahy
JV, et al: After asthma: Redefining airways diseases. Lancet.
391:350–400. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Bateman ED, Hurd SS, Barnes PJ, Bousquet
J, Drazen JM, FitzGerald M, Gibson P, Ohta K, O'Byrne P, Pedersen
SE, et al: Global strategy for asthma management and prevention:
GINA executive summary. Eur Respir J. 31:143–178. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Ray A, Raundhal M, Oriss TB, Ray P and
Wenzel SE: Current concepts of severe asthma. J Clin Invest.
126:2394–2403. 2016. View
Article : Google Scholar : PubMed/NCBI
|
|
4
|
Word Health Organiztion (2017), . Asthma
Fact Sheet. http://www.who.int/mediacentre/factsheets/fs307/en/May
26–2018
|
|
5
|
McInnes RN, Hernming D, Burgess P, Lyndsay
D, Osborne NJ, Skjøth CA, Thomas S and Vardoulakis S: Mapping
allergenic pollen vegetation in UK to study environmental exposure
and human health. Sci Total Environ. 599-600:483–499. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Stein MM, Hrusch CL, Gozdz J, Igartua C,
Pivniouk V, Murray SE, Ledford JG, dos Santos MM, Anderson RL,
Metwali N, et al: Innate immunity and asthma risk in amish and
hutterite farm children. N Engl J Med. 375:411–421. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Bleecker ER, FitzGerald JM, Chanez P, Papi
A, Weinstein SF, Barker P, Sproule S, Gilmartin G, Aurivillius M,
Werkström V, et al: Efficacy and safety of benralizumab for
patients with severe asthma uncontrolled with high-dosage inhaled
corticosteroids and long-acting β2-agonists (SIROCCO): A
randomised, multicentre, placebo-controlled phase 3 trial. Lancet.
388:2115–2127. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Chiu CN, Rihel J, Lee DA, Singh C, Mosser
EA, Chen SJ, Sapin V, Pham U, Engle J, Niles BJ, et al: A zebrafish
genetic screen identifies neuromedin U as a regulator of sleep/wake
states. Neuron. 89:842–856. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Schlegel P, Texada MJ, Miroschnikow A,
Schoofs A, Hückesfeld S, Peters M, Schneider-Mizell CM, Lacin H, Li
F, Fetter RD, et al: Synaptic transmission parallels
neuromodulation in a central food-intake circuit. Elife.
5:e167992016. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Martinez VG and O'Driscoll L: Neuromedin
U: A multifunctional neuropeptide with pleiotropic roles. Clin
Chem. 61:471–482. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Lee J, Snyder ER, Liu YH, Gu XY, Wang J,
Flowers BM, Kim YJ, Park S, Szot GL, Hruban RH, et al:
Reconstituting development of pancreatic intraepithelial neoplasia
from primary human pancreas duct cells. Nat Commun. 8:146862017.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Alfa RW, Park S, Skelly KR, Poffenberger
G, Jain N, Gu X, Kockel L, Wang J, Liu YH, Powers AC and Kim SK:
Suppression of insulin production and secretion by a decretin
hormone. Cell Metab. 21:323–333. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Wallrapp A, Riesenfeld SJ, Burkett PR,
Abdulnour RE, Nyman J, Dionne D, Hofree M, Cuoco MS, Rodman C,
Farouq D, et al: The neuropeptide NMU amplifies ILC2-driven
allergic lung inflammation. Nature. 549:351–356. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Cardoso V, Chesné J, Ribeiro H,
García-Cassani B, Carvalho T, Bouchery T, Shah K, Barbosa-Morais
NL, Harris N and Veiga-Fernandes H: Neuronal regulation of type 2
innate lymphoid cells via neuromedin U. Nature. 549:277–281. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Klose CSN, Mahlakõiv T, Moeller JB, Rankin
LC, Flamar AL, Kabata H, Monticelli LA, Moriyama S, Putzel GG,
Rakhilin N, et al: The neuropeptide neuromedin U stimulates innate
lymphoid cells and type 2 inflammation. Nature. 549:282–286. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Hargreave FE and Nair P: The definition
and diagnosis of asthma. Clin Exp Allergy. 39:1652–1658. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
O'Reilly R, Ullmann N, Irving S, Bossley
CJ, Sonnappa S, Zhu J, Oates T, Banya W, Jeffery PK, Bush A and
Saglani S: Increased airway smooth muscle in preschool wheezers who
have asthma at school age. J Allergy Clin Immunol. 131:1024–1032,
32.e1-16. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Noble PB, Pascoe CD, Lan B, Ito S,
Kistemaker LE, Tatler AL, Pera T, Brook BS, Gosens R and West AR:
Airway smooth muscle in asthma: Linking contraction and
mechanotransduction to disease pathogenesis and remodelling. Pulm
Pharmacol Ther. 29:96–107. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Nair P, Martin JG, Cockcroft DC, Dolovich
M, Lemiere C, Boulet LP and O'Byrne PM: Airway hyperresponsiveness
in asthma: Measurement and clinical relevance. J Allergy Clin
Immunol Pract. 5:649–659.e2. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Jackson DJ and Johnston SL: The role of
viruses in acute exacerbations of asthma. J Allergy Clin Immunol.
125:1178–1187; quiz 1188–1189. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Pascoe S, Locantore N, Dransfield MT,
Barnes NC and Pavord ID: Blood eosinophil counts, exacerbations,
and response to the addition of inhaled fluticasone furoate to
vilanterol in patients with chronic obstructive pulmonary disease:
A secondary analysis of data from two parallel randomised
controlled trials. Lancet Respir Med. 3:435–442. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Brusselle GG, Maes T and Bracke KR:
Eosinophils in the spotlight: Eosinophilic airway inflammation in
nonallergic asthma. Nat Med. 19:977–979. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Veres TZ, Shevchenko M, Krasteva G, Spies
E, Prenzler F, Rochlitzer S, Tschernig T, Krug N, Kummer W and
Braun A: Dendritic cell-nerve clusters are sites of T cell
proliferation in allergic airway inflammation. Am J Pathol.
174:808–817. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Furuhashi K, Chua YL, Wong KHS, Zhou Q,
Lee DCP, Liong KH, Teo GH, Hutchinson PE and Kemeny DM: Priming
with high and low respiratory allergen dose induces differential
CD4+ T helper type 2 cells and IgE/IgG1 antibody
responses in mice. Immunology. 151:227–238. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Suzuki R, Leach S, Liu WH, Ralston E,
Scheffel J, Zhang W, Lowell CA and Rivera J: Molecular editing of
cellular responses by the high-affinity receptor for IgE. Science.
343:1021–1025. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Cao PP, Zhang YN, Liao B, Ma J, Wang BF,
Wang H, Zeng M, Liu WH, Schleimer RP and Liu Z: Increased local IgE
production induced by common aeroallergens and phenotypic
alteration of mast cells in Chinese eosinophilic, but not
non-eosinophilic, chronic rhinosinusitis with nasal polyps. Clin
Exp Allergy. 44:690–700. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Moretti S, Renga G, Oikonomou V, Galosi C,
Pariano M, Iannitti RG, Borghi M, Puccetti M, De Zuani M, Pucillo
CE, et al: A mast cell-ILC2-Th9 pathway promotes lung inflammation
in cystic fibrosis. Nat Commun. 8:140172017. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Serafini N, Vosshenrich CA and Di Santo
JP: Transcriptional regulation of innate lymphoid cell fate. Nat
Rev Immunol. 15:415–428. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Licona-Limón P, Kim LK, Palm NW and
Flavell RA: TH2, allergy and group 2 innate lymphoid cells. Nat
Immunol. 14:536–542. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Mesnil C, Raulier S, Paulissen G, Xiao X,
Birrell MA, Pirottin D, Janss T, Starkl P, Ramery E, Henket M, et
al: Lung-resident eosinophils represent a distinct regulatory
eosinophil subset. J Clin Invest. 126:3279–3295. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Bal SM, Bernink JH, Nagasawa M, Groot J,
Shikhagaie MM, Golebski K, van Drunen CM, Lutter R, Jonkers RE,
Hombrink P, et al: IL-1β, IL-4 and IL-12 control the fate of group
2 innate lymphoid cells in human airway inflammation in the lungs.
Nat Immunol. 17:636–645. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Wenzel SE: Asthma phenotypes: The
evolution from clinical to molecular approaches. Nat Med.
18:716–725. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Sugita K, Steer CA, Martinez-Gonzalez I,
Altunbulakli C, Morita H, Castro-Giner F, Kubo T, Wawrzyniak P,
Ruckert B, Sudo K, et al: Type 2 innate lymphoid cells disrupt
bronchial epithelial barrier integrity by targeting tight junctions
through IL-13 in asthmatic patients. J Allergy Clin Immunol.
141:300–310.e11. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Pauwels RA, Löfdahl CG, Postma DS,
Tattersfield AE, O'Byrne P, Barnes PJ and Ullman A: Effect of
inhaled formoterol and budesonide on exacerbations of asthma.
formoterol and corticosteroids establishing therapy (FACET)
international study group. N Engl J Med. 337:1405–1411. 1997.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Gibson PG, Powell H and Ducharme FM:
Differential effects of maintenance long-acting beta-agonist and
inhaled corticosteroid on asthma control and asthma exacerbations.
J Allergy Clin Immunol. 119:344–350. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Weinstein RS: Clinical practice.
Glucocorticoid-induced bone disease. N Engl J Med. 365:62–70. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Mak VH, Melchor R and Spiro SG: Easy
bruising as a side-effect of inhaled corticosteroids. Eur Respir J.
5:1068–1074. 1992.PubMed/NCBI
|
|
38
|
Brown PH, Greening AP and Crompton GK:
Large volume spacer devices and the influence of high dose
beclomethasone dipropionate on hypothalamo-pituitary-adrenal axis
function. Thorax. 48:233–238. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Mukherjee M, Aleman Paramo F, Kjarsgaard
M, Salter B, Nair G, LaVigne N, Radford K, Sehmi R and Nair P:
Weight-adjusted intravenous reslizumab in severe asthma with
inadequate response to fixed-dose subcutaneous mepolizumab. Am J
Respir Crit Care Med. 197:38–46. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Minamino N, Kangawa K and Matsuo H:
Neuromedin U-8 and U-25: Novel uterus stimulating and hypertensive
peptides identified in porcine spinal cord. Biochem Biophys Res
Commun. 130:1078–1085. 1985. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Domin J, Ghatei MA, Chohan P and Bloom SR:
Neuromedin U--a study of its distribution in the rat. Peptides.
8:779–784. 1987. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Tanida M, Satomi J, Shen J and Nagai K:
Autonomic and cardiovascular effects of central neuromedin U in
rats. Physiol Behav. 96:282–288. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Austin C, Lo G, Nandha KA, Meleagros L and
Bloom SR: Cloning and characterization of the cDNA encoding the
human neuromedin U (NmU) precursor: NmU expression in the human
gastrointestinal tract. J Mol Endocrinol. 14:157–169. 1995.
View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Gevaert B, Wynendaele E, Stalmans S,
Bracke N, D'Hondt M, Smolders I, van Eeckhaut A and De Spiegeleer
B: Blood-brain barrier transport kinetics of the neuromedin
peptides NMU, NMN, NMB and NT. Neuropharmacology. 107:460–470.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Mitchell JD, Maguire JJ and Davenport AP:
Emerging pharmacology and physiology of neuromedin U and the
structurally related peptide neuromedin S. Br J Pharmacol.
158:87–103. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Szekeres PG, Muir AI, Spinage LD, Miller
JE, Butler SI, Smith A, Rennie GI, Murdock PR, Fitzgerald LR, Wu H,
et al: Neuromedin U is a potent agonist at the orphan G
protein-coupled receptor FM3. J Biol Chem. 275:20247–20250. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Alexander SP, Mathie A and Peters JA:
Guide to receptors and channels (GRAC), 3rd edition. Br J
Pharmacol. 153 (Suppl 2):S1–S209. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Hosoya M, Moriya T, Kawamata Y, Ohkubo S,
Fujii R, Matsui H, Shintani Y, Fukusumi S, Habata Y, Hinuma S, et
al: Identification and functional characterization of a novel
subtype of neuromedin U receptor. J Biol Chem. 275:29528–29532.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Fujii R, Hosoya M, Fukusumi S, Kawamata Y,
Habata Y, Hinuma S, Onda H, Nishimura O and Fujino M:
Identification of neuromedin U as the cognate ligand of the orphan
G protein-coupled receptor FM-3. J Biol Chem. 275:21068–21074.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Howard AD, Wang R, Pong SS, Mellin TN,
Strack A, Guan XM, Zeng Z, Williams DL Jr, Feighner SD, Nunes CN,
et al: Identification of receptors for neuromedin U and its role in
feeding. Nature. 406:70–74. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Kakarala KK and Jamil K:
Sequence-structure based phylogeny of GPCR Class A Rhodopsin
receptors. Mol Phylogenet Evol. 74:66–96. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Brighton PJ, Szekeres PG, Wise A and
Willars GB: Signaling and ligand binding by recombinant neuromedin
U receptors: Evidence for dual coupling to Galphaq/11 and Galphai
and an irreversible ligand-receptor interaction. Mol Pharmacol.
66:1544–1556. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Hsu SH and Luo CW: Molecular dissection of
G protein preference using Gsalpha chimeras reveals novel ligand
signaling of GPCRs. Am J Physiol Endocrinol Metab. 293:E1021–E1029.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Moriyama M, Matsukawa A, Kudoh S,
Takahashi T, Sato T, Kano T, Yoshimura A and Kojima M: The
neuropeptide neuromedin U promotes IL-6 production from macrophages
and endotoxin shock. Biochem Biophys Res Commun. 341:1149–1154.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Johnson EN, Appelbaum ER, Carpenter DC,
Cox RF, Disa J, Foley JJ, Ghosh SK, Naselsky DP, Pullen MA, Sarau
HM, et al: Neuromedin U elicits cytokine release in murine Th2-type
T cell clone D10.G4.1. J Immunol. 173:7230–7238. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Cording S, Medvedovic J, Aychek T and
Eberl G: Innate lymphoid cells in defense, immunopathology and
immunotherapy. Nat Immunol. 17:755–757. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Zhang Y, Jiang D, Zhang Y, Jiang X, Wang F
and Tao J: Neuromedin U type 1 receptor stimulation of A-type K+
current requires the βg subunits of Go protein, protein kinase A,
and extracellular signal-regulated kinase 1/2 (ERK1/2) in sensory
neurons. J Biol Chem. 287:18562–18572. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Gaudenzio N, Sibilano R, Marichal T,
Starkl P, Reber LL, Cenac N, McNeil BD, Dong XZ, Hernandez JD,
Sagi-Eisenberg R, et al: Different activation signals induce
distinct mast cell degranulation strategies. J Clin Invest.
126:3981–3998. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Mizutani N, Nabe T and Yoshino S:
IgE/antigen-mediated enhancement of IgE production is a mechanism
underlying the exacerbation of airway inflammation and remodelling
in mice. Immunology. 144:107–115. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Zhang N, Li H, Jia JH and He MQ:
Anti-inflammatory effect of curcumin on mast cell-mediated allergic
responses in ovalbumin-induced allergic rhinitis mouse. Cell
Immunol. 298:88–95. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Tore F and Tuncel N: Mast cells: Target
and source of neuropeptides. Curr Pharm Des. 15:3433–3445. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Moriyama M, Sato T, Inoue H, Fukuyama S,
Teranishi H, Kangawa K, Kano T, Yoshimura A and Kojima M: The
neuropeptide neuromedin U promotes inflammation by direct
activation of mast cells. J Exp Med. 202:217–224. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Smith SG, Chen R, Kjarsgaard M, Huang C,
Oliveria JP, O'Byrne PM, Gauvreau GM, Boulet LP, Lemiere C, Martin
J, et al: Increased numbers of activated group 2 innate lymphoid
cells in the airways of patients with severe asthma and persistent
airway eosinophilia. J Allergy Clin Immunol. 137:75–86.e8. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Mukherjee M, Bulir DC, Radford K,
Kjarsgaard M, Huang CM, Jacobsen EA, Ochkur SI, Catuneanu A,
Lamothe-Kipnes H, Mahony J, et al: Sputum autoantibodies in
patients with severe eosinophilic asthma. J Allergy Clin Immunol.
141:1269–1279. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Johansson MW: Eosinophil activation status
in separate compartments and association with asthma. Front Med
(Lausanne). 4:752017. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Borchers MT, Justice PJ, Ansay T, Mancino
V, McGarry MP, Crosby J, Simon MI, Lee NA and Lee JJ: Gq signaling
is required for allergen-induced pulmonary eosinophilia. J Immunol.
168:3543–3549. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Moriyama M, Fukuyama S, Inoue H, Matsumoto
T, Sato T, Tanaka K, Kinjyo I, Kano T, Yoshimura A and Kojima M:
The neuropeptide neuromedin U activates eosinophils and is involved
in allergen-induced eosinophilia. Am J Physiol Lung Cell Mol
Physiol. 290:L971–L977. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Talbot S, Foster SL and Woolf CJ:
Neuroimmunity: Physiology and pathology. Annu Rev Immunol.
34:421–447. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Ni D, Gu Q, Hu HZ, Gao N, Zhu MX and Lee
LY: Thermal sensitivity of isolated vagal pulmonary sensory
neurons: Role of transient receptor potential vanilloid receptors.
Am J Physiol Regul Integr Comp Physiol. 291:R541–R550. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Myers AC, Kajekar R and Undem BJ: Allergic
inflammation-induced neuropeptide production in rapidly adapting
afferent nerves in guinea pig airways. Am J Physiol Lung Cell Mol
Physiol. 282:L775–L781. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Canning BJ and Spina D: Sensory nerves and
airway irritability. Handb Exp Pharmacol. 139–183. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Hoogerwerf WA, Zou L, Shenoy M, Sun D,
Micci MA, Lee-Hellmich H, Xiao SY, Winston JH and Pasricha PJ: The
proteinase-activated receptor 2 is involved in nociception. J
Neurosci. 21:9036–9042. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Talbot S, Abdulnour RE, Burkett PR, Lee S,
Cronin SJ, Pascal MA, Laedermann C, Foster SL, Tran JV, Lai N, et
al: Silencing nociceptor neurons reduces allergic airway
inflammation. Neuron. 87:341–354. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Ballesta J, Carlei F, Bishop AE, Steel JH,
Gibson SJ, Fahey M, Hennessey R, Domin J, Bloom SR and Polak JM:
Occurrence and developmental pattern of neuromedin U-immunoreactive
nerves in the gastrointestinal tract and brain of the rat.
Neuroscience. 25:797–816. 1988. View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Yu XH, Cao CQ, Mennicken F, Puma C, Dray
A, O'Donnell D, Ahmad S and Perkins M: Pro-nociceptive effects of
neuromedin U in rat. Neuroscience. 120:467–474. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Wall PD and Melzack R: Textbook of Pain.
4th. Churchill Livingstone; London: 1999
|
|
77
|
Cao CQ, Yu XH, Dray A, Filosa A and
Perkins MN: A pro-nociceptive role of neuromedin U in adult mice.
Pain. 104:609–616. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Nakahara K, Kojima M, Hanada R, Egi Y, Ida
T, Miyazato M, Kangawa K and Murakami N: Neuromedin U is involved
in nociceptive reflexes and adaptation to environmental stimuli in
mice. Biochem Biophys Res Commun. 323:615–620. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
79
|
Fang L, Zhang M, Li C, Dong S and Hu Y:
Chemical genetic analysis reveals the effects of NMU2R on the
expression of peptide hormones. Neurosci Lett. 404:148–153. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
80
|
Zheng X, Hu Y, Liu J and Ouyang K:
Screening of active compounds as neuromedin U2 receptor agonist
from natural products. Bioorg Med Chem Lett. 15:4531–4535. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
81
|
Takayama K, Mori K, Sohma Y, Taketa K,
Taguchi A, Yakushiji F, Minamino N, Miyazato M, Kangawa K and
Hayashi Y: Discovery of potent hexapeptide agonists to human
neuromedin U receptor 1 and identification of their serum
metabolites. ACS Med Chem Lett. 6:302–307. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
82
|
Szczeklik A, Czerniawska-Mysik G,
Adamek-Guzik T, Woloszynski J and Koterba A: Ketotifen versus
sodium cromoglycate in the therapy of allergic (extrinsic)
bronchial asthma. Respiration. 39 (Suppl 1):S3–S9. 1980. View Article : Google Scholar
|
|
83
|
Theiler A, Barnthaler T, Platzer W,
Richtig G, Peinhaupt M, Rittchen S, Kargl J, Ulven T, Marsh LM,
Marsche G, et al: Butyrate ameliorates allergic airway inflammation
by limiting eosinophil trafficking and survival. J Allergy Clin
Immunol. 144:764–776. 2019. View Article : Google Scholar : PubMed/NCBI
|