Introduction
Lung cancer presents a major health threat to men
and women worldwide (1). The
mortality rate of lung cancer is the highest among all human
malignancies. Patients generally present at an advanced stage at
initial diagnosis and their estimated five-year survival rate is
10–20% (2,3). Lung adenocarcinoma, which is a typical
form of non-small cell lung cancer (NSCLC), accounts for a high
proportion of lung cancer cases, and the majority of primary
pulmonary adenocarcinomas are highly heterogeneous (4). Although new management strategies for
NSCLC, including chemotherapy and targeted therapy, have been
developed (5), the prognosis of
affected patients remains poor (6).
Gene expression microarrays facilitate the
identification of differentially expressed genes (DEGs) between
normal and tumor tissues. These genes may be promising biomarkers
and/or therapeutic targets for the diagnosis and treatment of
cancer (7). However, critical
molecular targets for lung adenocarcinomas have yet to be
identified.
With the aim of improving the diagnosis and
treatment of lung adenocarcinomas, in the present study, data
mining of the public Gene Expression Omnibus (GEO) database and
bioinformatics analysis was performed to identify differentially
expressed protein-coding genes between lung cancer and normal
tissue samples. The DEGs were assessed using Gene Ontology (GO)
functional enrichment analysis and Kyoto Encyclopedia of Genes and
Genomes (KEGG) pathway enrichment analysis to identify their
biological functions. A protein-protein interaction (PPI) network
of the DEGs was also constructed to identify hub genes. The results
indicated that DNA topoisomerase 2α (TOP2A), cell division cycle
protein 20 homolog (CDC20), mitotic checkpoint serine/threonine
kinase BUB1 (BUB1) and mitotic spindle assembly checkpoint MAD2A
(MAD2L1) may be potential biomarkers and therapeutic targets in
lung adenocarcinomas. In addition, reverse
transcription-quantitative PCR (RT-qPCR) analysis of the key genes
identified in 2 patient samples was used to verify the results of
the bioinformatics analysis.
Materials and methods
Bioinformatics prediction
The microarray dataset GSE27262, including 25 normal
tissue samples and 25 lung adenocarcinoma tissue samples, was
obtained from the GEO database (http://www.ncbi.nlm.nih.gov/geo) (8–11). The
GPL570 platform (Affymetrix; Thermo Fisher Scientific, Inc.) was
used to analyze the microarray data. The platform was an Affymetrix
Human Genome U133 Plus 2.0 Array. The annotation file was also
acquired.
Statistical computing and graphical representation
were performed using R software version 3.3.3 (The R Project for
Statistical Computing; The R Foundation). The R linear models for
microarray data (limma) package was used to accomplish all of the
data processing. After background subtraction and normalization
using robust multi-chip averaging, the samples from the GEO dataset
were divided into two groups: A disease group (25 tumor tissues)
and a control group (25 tumor-adjacent tissues). The limma
algorithm was used for differential analysis in disease. The
criteria to classify a gene as a DEG were a log fold change >2
and a significance of P<0.05 between tumor adjacent tissue
samples and tumor samples.
Co-expression network
The PPI network was established using the Search
Tool for the Retrieval of Interacting Genes/proteins (STRING;
version 11.0; http://string-db.org) and Cytoscape
software (version 3.6.1; www.cytoscape.org; The Cytoscape Consortium). Hub
genes were identified from the PPIs between differentially
expressed protein-coding genes and the edge lengths between nodes
from these hub genes were subsequently obtained.
Functional enrichment and pathway
analysis
DEG enriched GO terms and KEGG pathways were
determined to identify their biological function based on the
Database for Annotation, Visualization and Integrated Discovery
(DAVID; version 6.8; http://david.ncifcrf.gov).
Survival analysis
Kaplan-Meier curve and Cox regression analyses were
performed to identify the prognostic value of the key DEGs. The
patients from the dataset were divided into pairs of subgroups,
namely the ‘high expression group’ and the ‘low expression group’,
prior to performing the survival analysis. The Oncomine database
(version 4.5; http://www.oncomine.org) and Kaplan-Meier Plotter
database (www.kmplot.com) were used to perform the
survival analyses (12,13). Student's t-test was used to examine
the statistical significance between 2 groups. P<0.05 was
considered a statistically significant difference. The Oncomine and
Human Protein Atlas (HPA) databases (version 19; http://www.proteinatlas.org) are free open-access
databases (14), which were used to
validate the expression of TOP2A and BUB1 at the protein level in
lung adenocarcinoma tissues.
Verification of bioinformatics in
patient samples
Paired lung cancer tissues and adjacent normal
tissues were collected in January 2018 from two patients with lung
cancer, 1 male (age, 64; BMI, 18.9; no smoking history) and 1
female (age, 74; BMI, 19.9; no smoking history), that were treated
at the Affiliated Hospital of Qingdao University (Qingdao, China).
The patients included in the study were lung adenocarcinoma
patients who had received surgical treatment for lung
adenocarcinoma. To confirm the results of the present
bioinformatics analysis the expression levels of TOP2A, CDC20, BUB1
and MAD2L1 in tumor tissues of two lung adenocarcinoma patients
were quantified using RT-qPCR. Specimens were frozen in liquid
nitrogen immediately after resection and stored at −80°C. Total RNA
was extracted using NcmZol reagent (NCM Biotech), according to the
manufacturer's protocol. A FastQuant RT kit (with gDNase; Tiangen
Biotech Co., Ltd.) was used to reverse transcribe total RNA to
cDNA; the temperature of the RT process was 42°C. qPCR was
performed in an ABI StepOnePlus Thermocycler (Applied Biosystems;
Thermo Fisher Scientific, Inc.) using a SYBR Green PCR kit (Qiagen
GmbH). The primer sequences are listed in Table I. The following thermocycling
conditions were used for the qPCR: Initial denaturation temperature
is 95°C for 15 min; 40 cycles of 95°C for 10 sec, 55°C for 30 sec
and 72°C for 30 sec. All of the analyses were performed in
triplicate and the relative expression levels were calculated based
on the comparative 2−ΔΔCq method (15).
 | Table I.Primer sequences of β-actin, TOP2A,
BUB1, CDC20 and MAD2L1 used for reverse transcription-quantitative
PCR. |
Table I.
Primer sequences of β-actin, TOP2A,
BUB1, CDC20 and MAD2L1 used for reverse transcription-quantitative
PCR.
| Gene | Primer sequence
(5′→3′) |
|---|
| β-actin | F:
AAGAGAGGCATCCTGACCCT |
|
| R:
TACATGGCTGGGGTGTTGAA |
| TOP2A | F:
ACCATTGCAGCCTGTAAATGA |
|
| R:
GGGCGGAGCAAAATATGTTCC |
| BUB1 | F:
AGCCCAGACAGTAACAGACTC |
|
| R:
GTTGGCAACCTTATGTGTTTCAC |
| CDC20 | F:
GCTTTGAACCTGAACGGTTTTG |
|
| R:
TCTGGCGCATTTTGTGGTTTT |
| MAD2L1 | F:
GGACTCACCTTGCTTGTAACTAC |
|
| R:
GATCACTGAACGGATTTCATCCT |
Statistical analysis
Statistical analysis for comparison between the
tumor group and the control group was performed using unpaired
Student' t-test. The fold change >2 and P<0.05 were
considered to indicate a statistically significant difference.
Student's t-test was also used to examine the statistical
significance of survival analysis and the identification of hub
genes. P<0.05 was considered to indicate a statistically
significant difference. The expression of TOP2A and BUB1 in
different stages of lung cancer was analyzed using an F-test. F
test was performed as part of an ANOVA. Student-Newman-Keuls was
used as a post hoc test after ANOVA.
Results
Identification of DEGs and
bioinformatics analysis
In the present database study, a total of 303 DEGs
were identified in lung adenocarcinomas, including 247
downregulated (Table SI) and 56
upregulated DEGs (Table SII). The
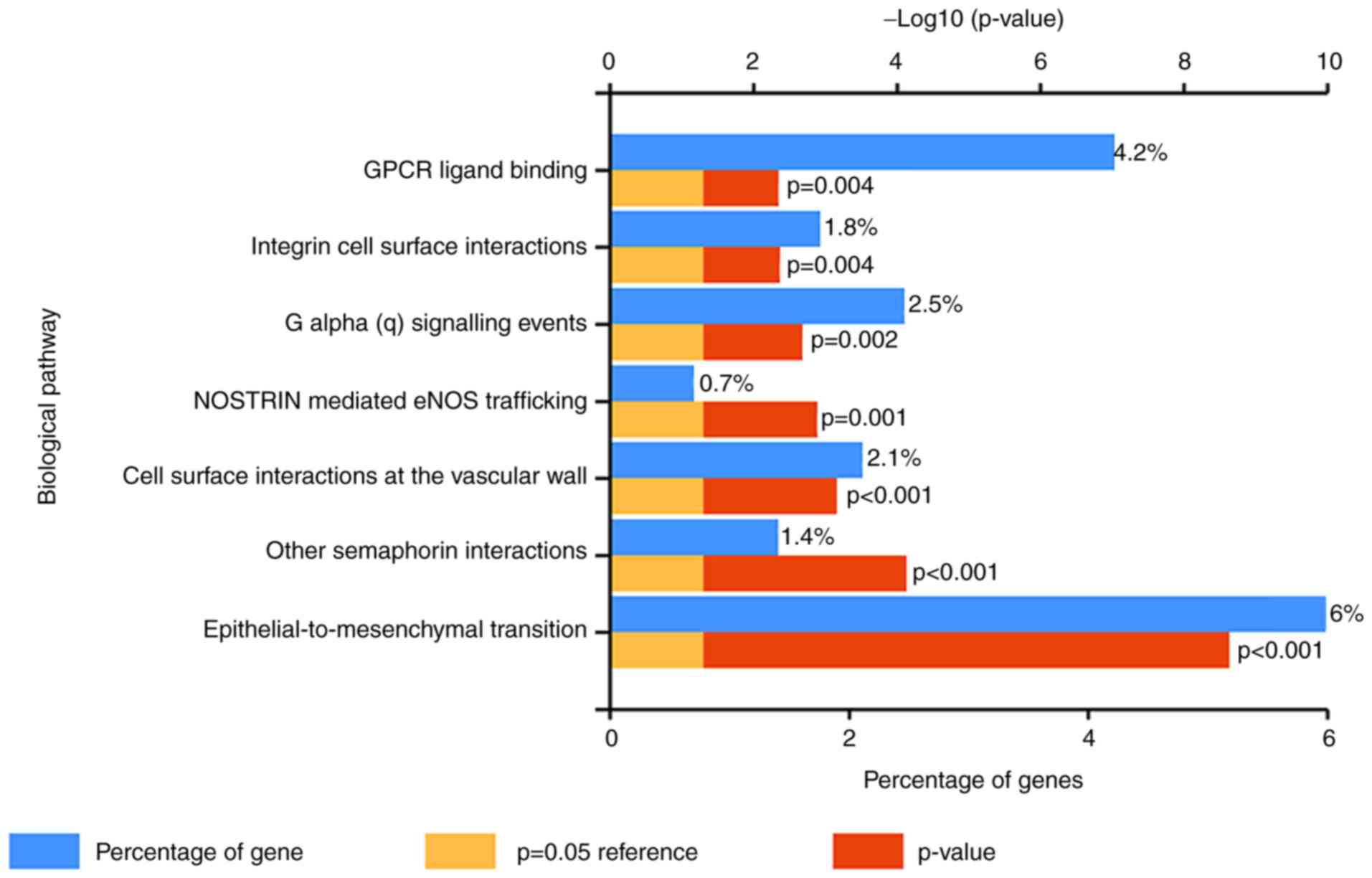
results of KEGG analysis indicated that these DEGs were enriched in
numerous cancer-associated biological pathways, including
epithelial-to-mesenchymal transition and G protein-coupled receptor
(GPCR) ligand binding (Fig. 1). The
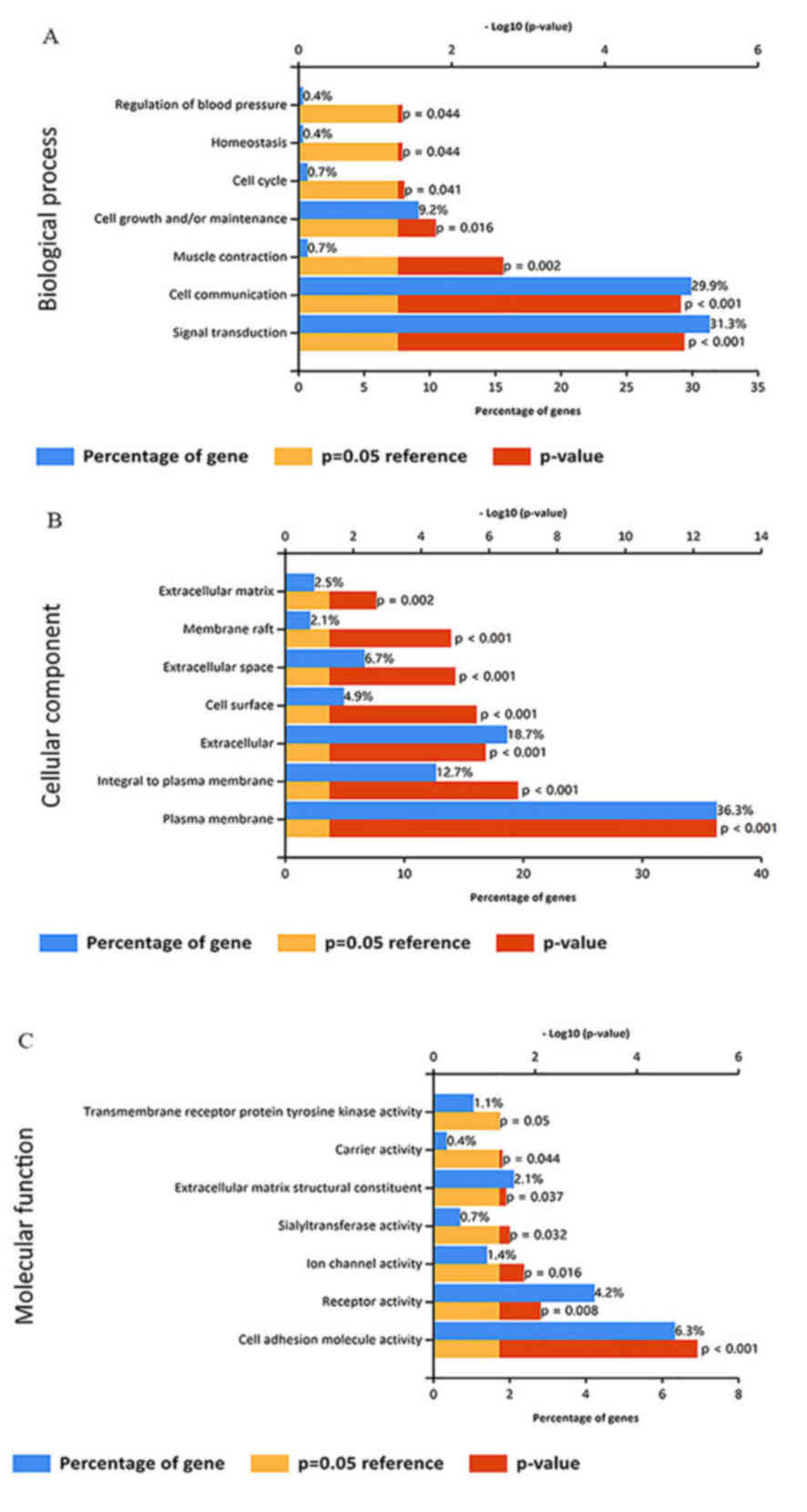
GO analyses indicated that the DEGs were enriched in numerous
cancer-associated terms in the category biological process,
including signal transduction, cell communication and cell growth
and/or maintenance (Fig. 2A). GO
enrichment analysis in the category cellular component indicated
that the DEGs are concentrated in the plasma membrane, and are
integral to the plasma membrane and extracellular matrix (Fig. 2B). In addition, the DEGs were
enriched in various cancer-associated GO terms in the category
molecular function, including cell adhesion molecule activity and
receptor activity (Fig. 2C).
TOP2A, CDC20, BUB1 and MAD2L1 are
significant hub genes in lung adenocarcinoma
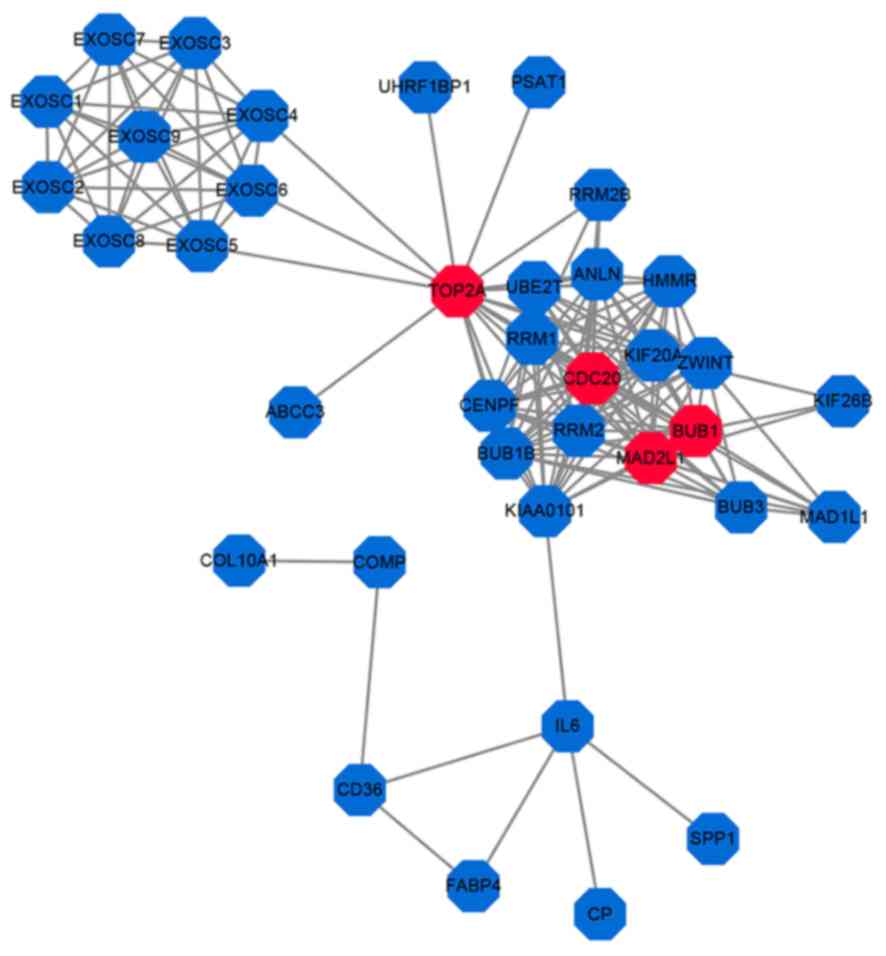
To identify the critical hub genes in lung
adenocarcinoma, a PPI network was generated for all 303 DEGs
(Fig. 3). Among the 303 DEGs, TOP2A,
CDC20, BUB1 and MAD2L1 were identified to have the highest degree
of interaction and were considered to be hub genes. In the present
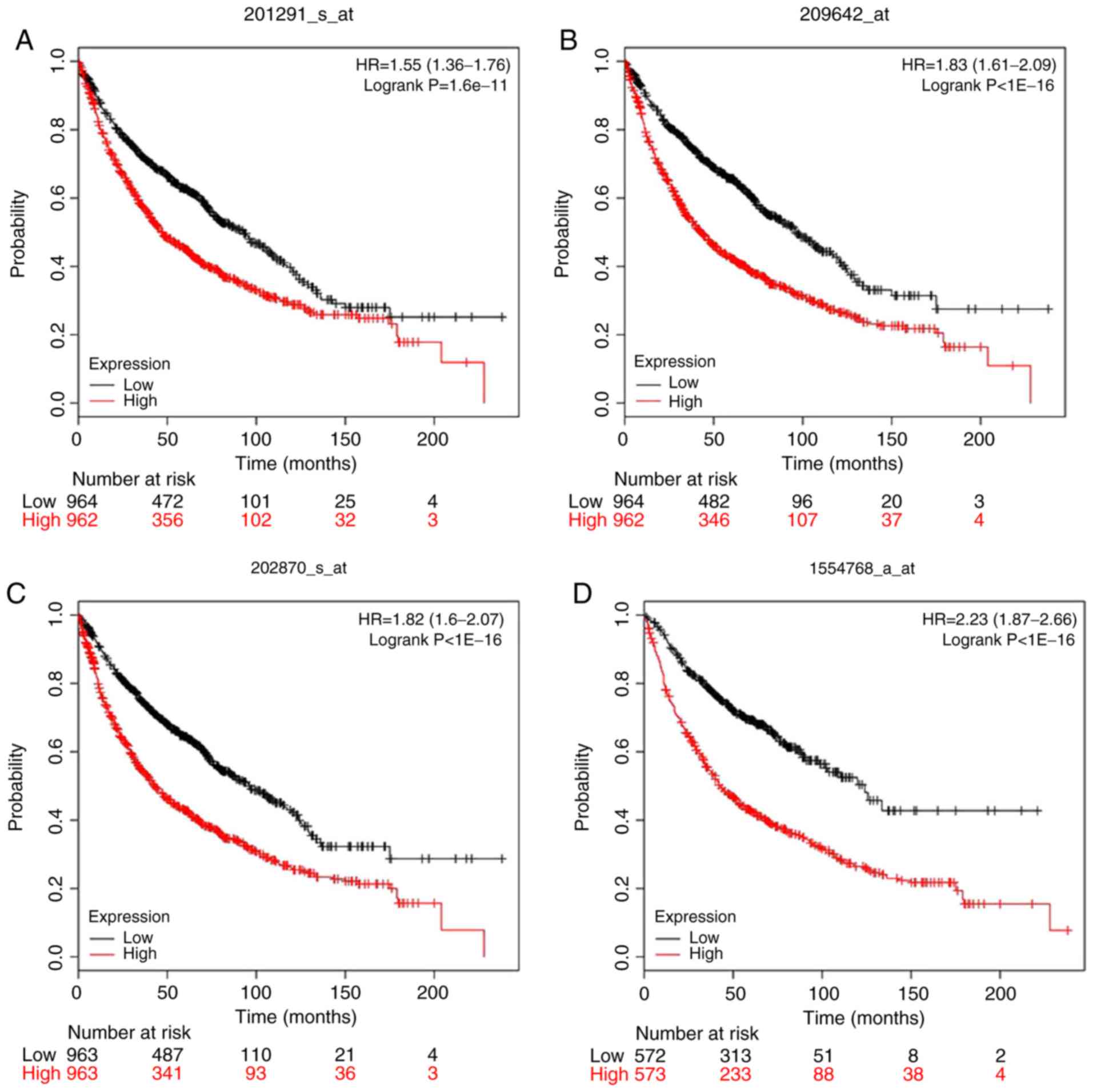
study, Kaplan-Meier curve and Cox regression analyses were
performed to determine the prognostic value of the four hub genes
identified from the PPI. The results indicated that TOP2A, CDC20,
BUB1 and MAD2L1 were all significantly associated with the survival
of patients with lung adenocarcinomas (hazard ratios: 1.55, 1.82,
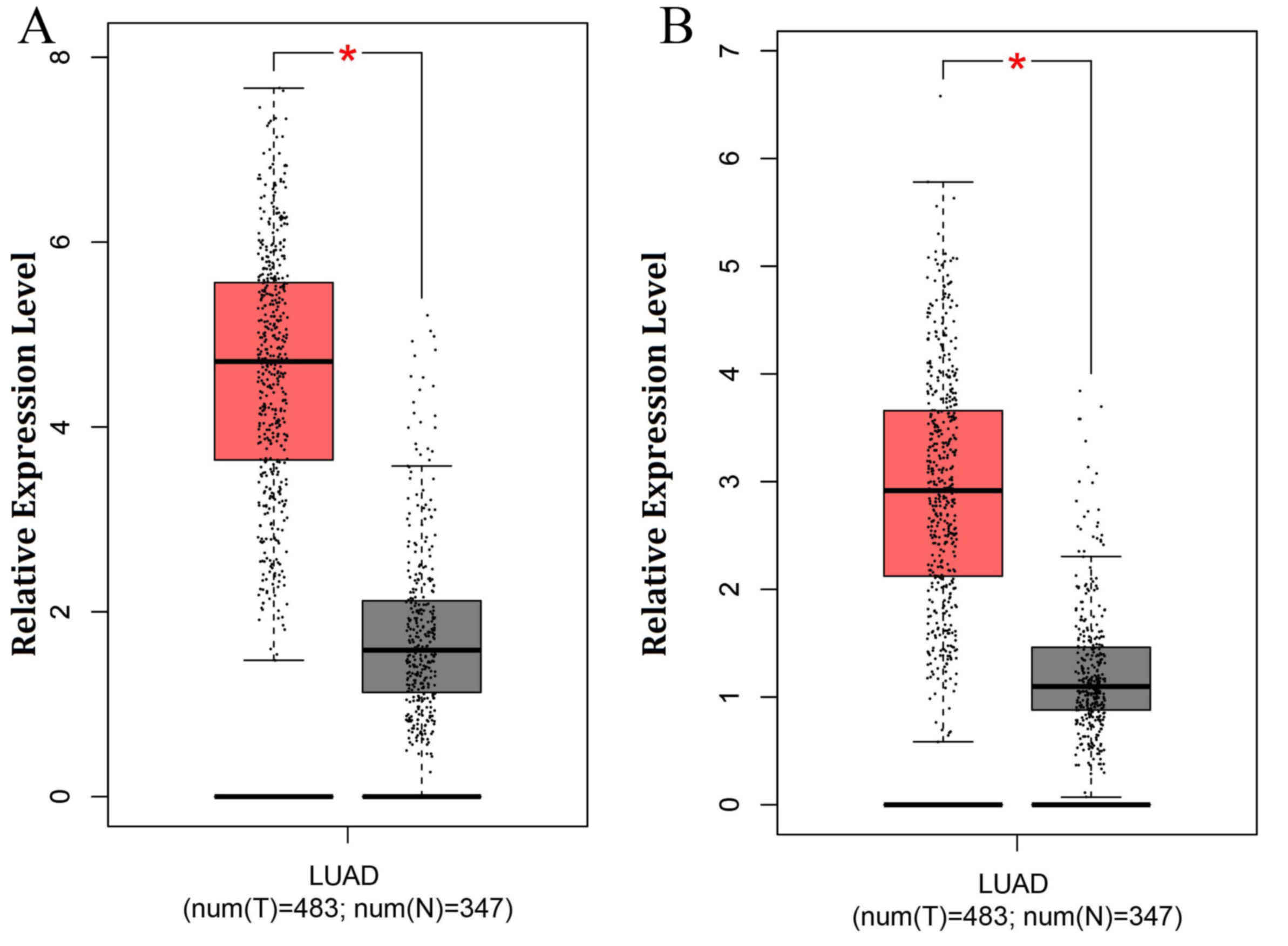
1.83 and 2.23, respectively) and were also highly expressed in lung
adenocarcinoma tissue (Fig. 4).
Therefore, these four DEGs may be potential biomarkers to predict
the survival of lung adenocarcinoma patients. In addition, TOP2A
and BUB1 were identified to be highly expressed in lung
adenocarcinoma tissues in patient samples from the Affiliated
Hospital of Qingdao University (Fig.
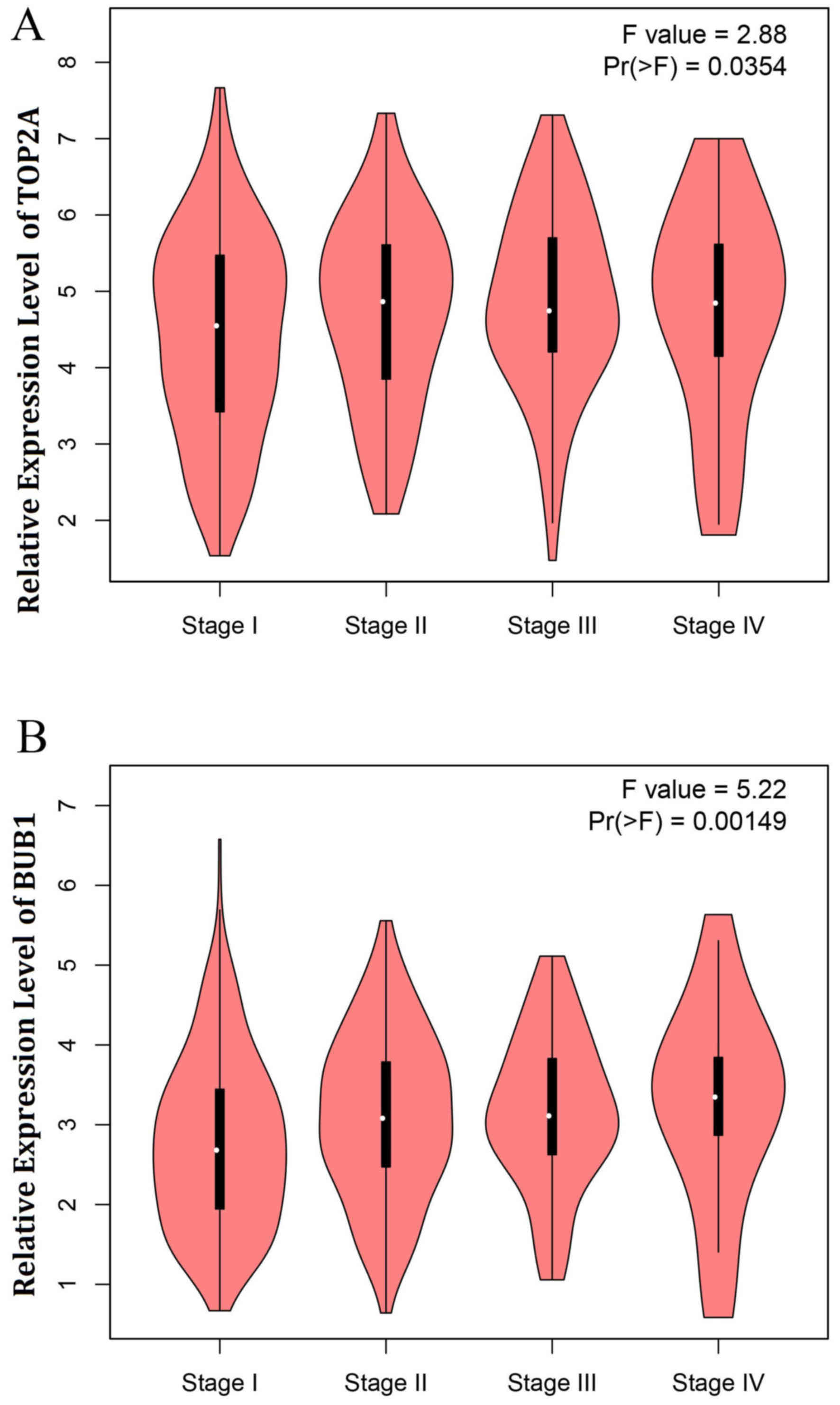
5). The expression of TOP2A and BUB1 in different stages of
lung cancer is presented in Fig. 6.
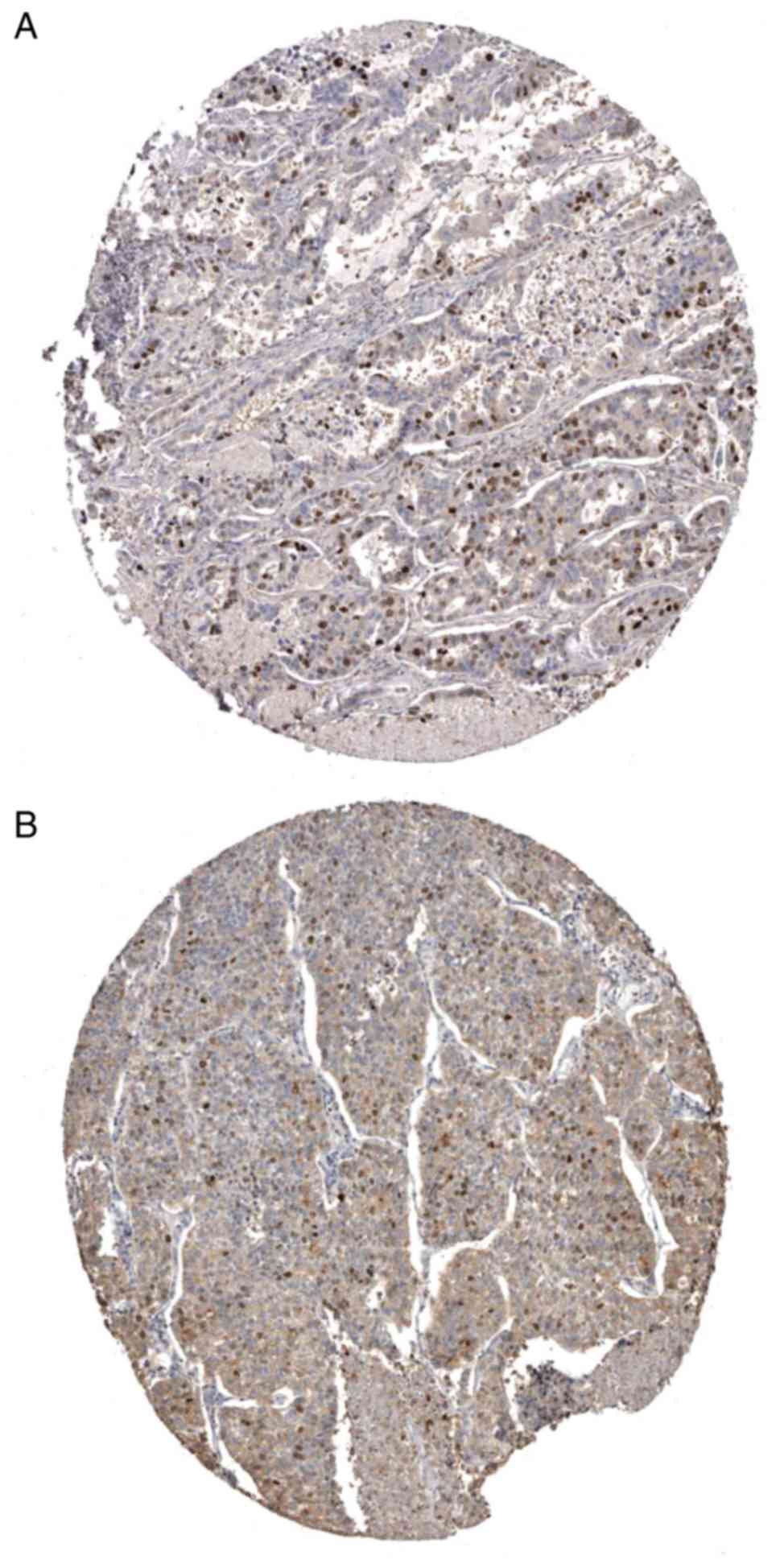
The Oncomine and HPA databases were used to analyze the validation
of the expression of TOP2A and BUB1 at the protein level in lung
adenocarcinoma tissues. The analysis results are presented in
Fig. 7.
Subsequent verification based on
RT-qPCR
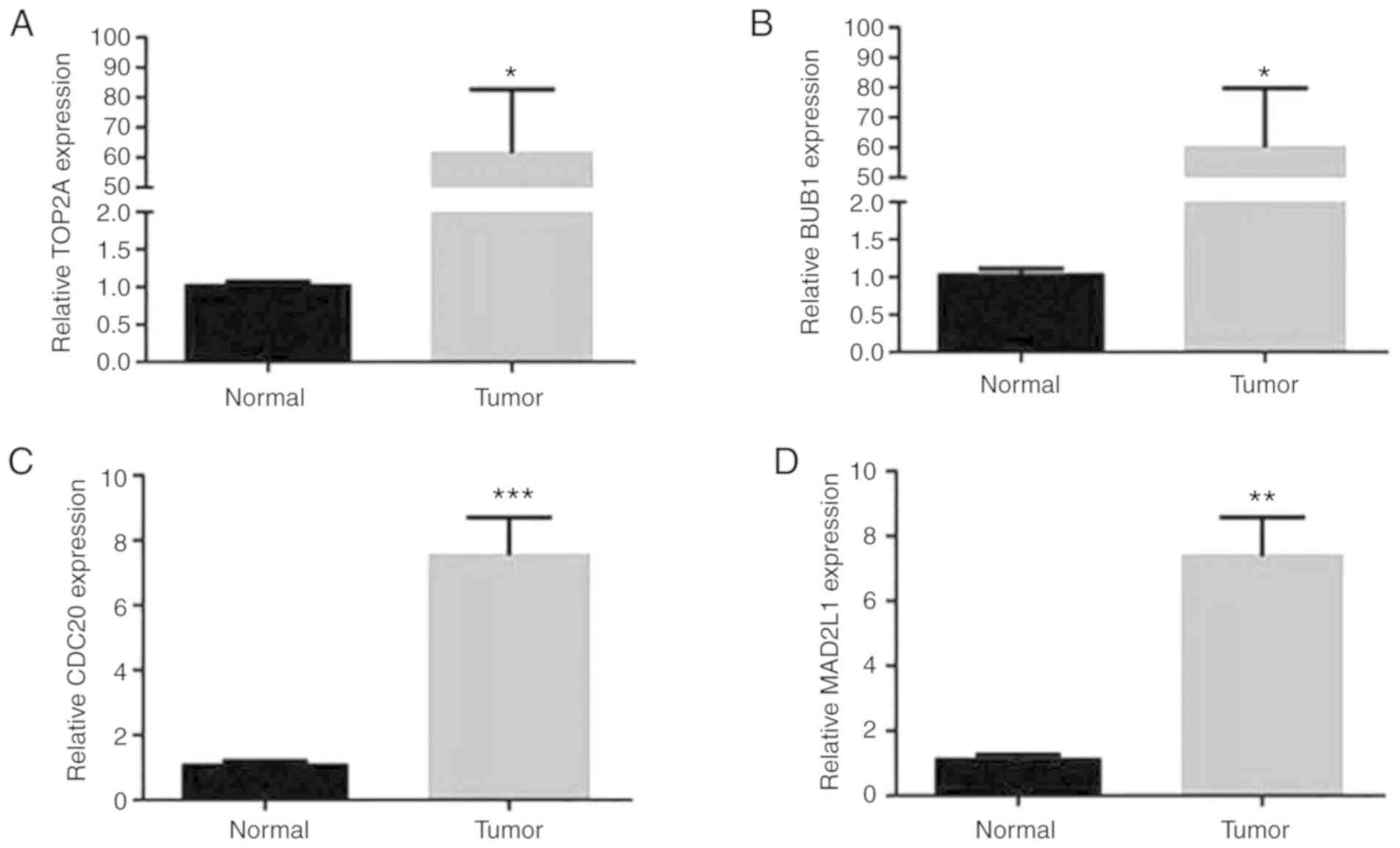
RT-qPCR was performed to experimentally confirm the
expression of TOP2A, CDC20, BUB1 and MAD2L1 in tumor tissues of
patients with lung adenocarcinoma, in order to verify the results
of the present bioinformatic analysis. As presented in Fig. 8, the mRNA expression of TOP2A, BUB1,
CDC20 and MAD2L1 in lung adenocarcinoma samples was significantly
increased compared with that in non-tumor tissues (all,
P<0.05).
Discussion
Despite advances in chemotherapy and targeted
therapies for NSCLC, lung adenocarcinoma remains a threat to human
life (16). The purpose of the
present study was to identify reliable biomarkers in lung
adenocarcinomas to improve the diagnosis and predicted survival of
patients and to provide novel therapeutic targets. The gene
expression profiles between lung cancer and normal tissues (25
pairs) from a GEO dataset were compared and 303 DEGs in lung NSCLC
tissues were identified; however, due to the number of samples in
the microarray dataset analyzed being limited to 25 pairs, future
studies should use a larger number of samples to verify the
findings of the present study. GO and KEGG pathway analyses may be
used to identify the biological functions of DEGs (17). The KEGG analysis performed in the
present study indicated that the DEGs were enriched in a number of
cancer-associated pathways, including epithelial-to-mesenchymal
transition and GPCR ligand binding. Furthermore, the GO enrichment
analysis in the category cellular component indicated that the DEGs
were concentrated in or/and are integral to the plasma membrane or
extracellular matrix. In the category biological process, the DEGs
were enriched in GO terms including signal transduction, cell
communication, and cell growth and/or maintenance, and in the
category molecular function, they were enriched in terms including
adhesion molecule activity and receptor activity. A PPI network was
generated to determine the critical hub genes and provide valuable
information for the analysis of cellular functions and biological
processes. TOP2A, CDC20, BUB1 and MAD2L1 displayed a high degree of
interaction in the PPI network and were considered as hub genes.
Survival analysis also indicated that these four key genes were
highly expressed in lung cancer and that their expression was
significantly associated with overall survival. Information,
including the TNM stage, performance status scores and the degree
of tumor histological differentiation for the patients was not
included in the dataset used to perform survival analysis, which is
a limitation of the present study. Taken together, the results of
the KEGG, GO and survival analyses demonstrated that TOP2A, CDC20,
BUB1 and MAD2L1 are significantly associated with lung
adenocarcinomas and may potentially serve as effective biomarkers
and therapeutic targets in this malignancy.
TOP2A is a highly evolutionarily conserved enzyme
with serves an important role in centromere biology (18). Previous studies have indicated that
TOP2A promotes tumor development and progression and may be used as
an early prognostic marker in numerous types of cancer,
particularly in breast cancer and ovarian cancer (19,20).
TOP2A protein has been considered as a marker that is associated
with increased tumor grade, since it is highly expressed in
proliferating cells (21).
Topoisomerase-targeting anti-cancer drugs have been successfully
used in the clinic, and their efficacy is based on topoisomerase
poisoning (22). Recently, it has
been reported that abnormal alterations and modifications of TOP2A
may be critical to chromosome instability in some cancer types
(23). Another recent study
indicated that miR-139 targets TOP2A and that defective miR-139
expression induces increased expression of TOP2A, and the increased
TOP2A then promotes the transcription of EMT-associated genes
through activating the β-catenin signaling pathway in cancer
(24).
BUB1 is a serine/threonine kinase protein that is
associated with kinase function and the spindle assembly checkpoint
(25,26). During mitosis and cell proliferation,
BUB1 is highly expressed. BUB1 is able to phosphorylate CDC20 in a
number of different ways. In early mitosis, BUB1 accumulates at
unattached kinetochores to promote the recruitment of core mitotic
checkpoint components, including mitotic spindle assembly
checkpoint MAD1, mitotic spindle assembly checkpoint MAD2A, mitotic
checkpoint serine/threonine kinase BUB1β, mitotic checkpoint
protein BUB3 and centromere protein E, which leads to inhibition of
CDC20 (27). It has been reported
that the expression of BUB1 may be a promising prognostic biomarker
in a number of different cancer types, including gastric
adenocarcinoma, breast cancer and ovarian cancer (20,28,29).
CDC20 is encoded by the CDC20 gene and serves an
essential role in the regulation of cell division (30). The most important function of CDC20
is thought to be the activation of a master cell cycle regulator
called the anaphase promoting complex/cyclosome (APC/C) by APC/C E3
ubiquitin ligase (31,32). Depletion of CDC20 has been indicated
to enhance the cytotoxicity of paclitaxel, indicating that CDC20 is
a potential marker that is associated with cancer (33). CDC20 has been reported to be
overexpressed in numerous human cancer cell lines and carcinoma
tissue types (34,35). Previous studies have also indicated
that high CDC20 expression is associated with poor prognosis in
multiple types of cancer (36–38).
Gene expression profiling studies have identified CDC20 as an
important gene that is associated with numerous types of cancer
(39,40). This evidence indicates that CDC20 may
be utilized as an effective potential biomarker and therapeutic
target in cancer.
MAD2L1 protein is a component of the mitotic spindle
assembly checkpoint and inhibits the onset of the anaphase by
sequestering CDC20 until all chromosomes are aligned at the
metaphase plate (41). It has been
reported that MAD2L1 is highly expressed in patients with lung
cancer (42), and furthermore,
MAD2L1 may have the potential to be used as a diagnostic marker and
therapeutic target in breast cancer (41). In addition, high intratumoral MAD2L1
levels may lead to reduced recurrence-free survival rates of
patients with cancer (41).
In the present study, a total of 56 upregulated
genes and 247 downregulated genes among a total of 303 DEGs were
identified using bioinformatics analysis. TOP2A, CDC20, BUB1 and
MAD2L1 were indicated to be significantly associated with survival
of patients with lung adenocarcinoma and may serve as potential
prognostic biomarkers for these patients; however, whether they can
accurately predict patient prognosis requires further research
because there are many other factors that can influence survival
rates of patients with cancer, including age and comorbidity
(43). The results of the present
study may provide valuable indications for further studies on lung
cancers. The absence of cell line-based analysis to confirm the
biological implication of altered expression of these genes is a
limitation of the present study. The absence of western blot to
measure the protein expressions of these genes and
immunohistochemistry to observe the expressions of these genes is
also a limitation. TOP2A and BUB1 were more significantly
upregulated than CDC20 and MAD2L1 in lung adenocarcinoma, according
to the results of RT-qPCR, so only the results of TOP2A and BUB1
were investigated in further experiments. The absence of further
data on CDC20 and MAD2L1 is an additional limitation of the current
study. The absence of data on the expression of these genes in
normal tissues and the expression of phospho-CDC20 are also
limitations of the present study. In the present study, the
relationship between the four identified hub genes and lung
adenocarcinoma was validated using RT-qPCR.
The present study may use the same bioinformatics
analysis tools as other microarray papers studying lung
adenocarcinoma, but the present study has focused on datasets that
have not, to the best of our knowledge, been analyzed previously in
this way and led to the identification of unique DEGs. Analysis of
further datasets may lead us to different conclusions. As the
EGFR-mutation in lung cancer may be a discrete entity, the
relationship between these four genes and the EGFR mutation should
be a focus of future study. In addition, the expression levels of a
number of genes in lung adenocarcinoma and non-lung adenocarcinoma
NSCLC are different; for example, the gene expression of
thymidylate synthase and excision repair cross complementing 1 in
lung adenocarcinoma patients are significantly higher compared with
patients with non-adenocarcinoma NSCLC (44). The data for bioinformatics analysis
in the present study is from patients with lung adenocarcinoma and
only four DEGs in patients with lung adenocarcinoma were detected;
however, there may be differences between lung adenocarcinoma and
non-adenocarcinoma NSCLC regarding the importance of these four
DEGs. Thus, the present study is limited because it lacks a
comparison of the expression of these four DEGs between lung
adenocarcinoma and non-lung adenocarcinoma NSCLC. Because there are
only two samples, the experimental results may be contingent, which
is another limitation of the present study. Finally, the Oncomine
and HPA database did not provide control samples of TOP2A and BUB1
expression levels, which is another limitation of the present
study.
In conclusion, TOP2A, CDC20, BUB1 and MAD2L1 may be
used as biomarkers and therapeutic targets in patients with lung
adenocarcinoma.
Supplementary Material
Supporting Data
Acknowledgements
Not applicable.
Funding
The present study was supported by The Zhejiang
Provincial Natural Science Foundation of China (grant no.
LQ16C090001).
Availability of data and materials
The dataset used and/or analyzed during the present
study is available in the GEO database (accession no. GSE27262;
http://www.ncbi.nlm.nih.gov/gds). All
other datasets used/or analyzed during the current study are
available from the corresponding author on reasonable request.
Authors' contributions
SL performed the data mining. LW performed RT-qPCR
and contributed to writing the manuscript. YW preserved and
extracted RNA and assisted with manuscript writing. ZT and CL
analyzed data and assisted with RT-qPCR. JL designed the
experiments and revised the manuscript. WJ recruited patients to
the study and harvested tissue. All of the authors read and
approved the final manuscript.
Ethics approval and consent to
participate
The present study was approved by the Ethics
Committee of the Affiliated Hospital of Qingdao University
(Qingdao, China; no. 2018020). Each patient was informed of the use
of their tissue specimens and provided a statement of written
informed consent.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Centers for Disease Control and
Prevention. Lung Cancer Statistics. simplecdc.gov/cancer/lung/statistics/index.htmMarch
26–2014
|
|
2
|
Jakobsen E, Rasmussen TR and Green A:
Mortality and survival of lung cancer in Denmark: Results from the
Danish lung cancer group 2000–2012. Acta Oncol. 55 (Suppl 2):S2–S9.
2016. View Article : Google Scholar
|
|
3
|
Richards TB, Henley SJ, Puckett MC, Weir
HK, Huang B, Tucker TC and Allemani C: Lung cancer survival in the
United States by race and stage (2001–2009): Findings from the
CONCORD-2 study. Cancer. 123 (Suppl 24):S5079–S5099. 2017.
View Article : Google Scholar
|
|
4
|
Travis WD: Pathology of lung cancer. Clin
Chest Med. 32:669–692. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Reck M, Heigener DF, Mok T, Soria JC and
Rabe KF: Management of non-small-cell lung cancer: Recent
developments. Lancet. 382:709–719. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Zhou C, Wu YL, Chen G, Feng J, Liu XQ,
Wang C, Zhang S, Wang J, Zhou S, Ren S, et al: Erlotinib versus
chemotherapy as first-line treatment for patients with advanced
EGFR mutation-positive non-small-cell lung cancer (OPTIMAL,
CTONG-0802): A multicentre, open-label, randomised, phase 3 study.
Lancet Oncol. 12:735–742. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Liang Y, Ma YY, Li LL, Shen XY, Xin T,
Zhao YW and Ma R: Effect of long non-coding RNA LINC01116 on
biological behaviors of non-small cell lung cancer cells via the
hippo signaling pathway. J Cell Biochem. 1192018.DOI:
10.1002/jcb.26711.
|
|
8
|
Barrett T and Edgar R: Mining microarray
data at NCBI's Gene Expression Omnibus (GEO)*. Methods Mol Biol.
338:175–190. 2006.PubMed/NCBI
|
|
9
|
Edgar R, Domrachev M and Lash AE: Gene
expression omnibus: NCBI gene expression and hybridization array
data repository. Nucleic Acids Res. 30:207–210. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Wei TY, Juan CC, Hisa JY, Su LJ, Lee YC,
Chou HY, Chen JM, Wu YC, Chiu SC, Hsu CP, et al: Protein arginine
methyltransferase 5 is a potential oncoprotein that upregulates G1
cyclins/cyclin-dependent kinases and the phosphoinositide
3-kinase/AKT signaling cascade. Cancer Sci. 103:1640–1650. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Wei TY, Hsia JY, Chiu SC, Su LJ, Juan CC,
Lee YC, Chen JM, Chou HY, Huang JY, Huang HM and Yu CT: Methylosome
protein 50 promotes androgen- and estrogen-independent
tumorigenesis. Cell Signal. 26:2940–2950. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Beer DG, Kardia SL, Huang CC, Giordano TJ,
Levin AM, Misek DE, Lin L, Chen G, Gharib TG, Thomas DG, et al:
Gene-expression profiles predict survival of patients with lung
adenocarcinoma. Nat Med. 8:816–824. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Rhodes DR, Yu J, Shanker K, Deshpande N,
Varambally R, Ghosh D, Barrette T, Pandey A and Chinnaiyan AM:
ONCOMINE: A cancer microarray database and integrated data-mining
platform. Neoplasia. 6:1–6. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Uhlén M, Fagerberg L, Hallström BM,
Lindskog C, Oksvold P, Mardinoglu A, Sivertsson Å, Kampf C,
Sjöstedt E, Asplund A, et al: Proteomics. Tissue-based map of the
human proteome. Science. 347:12604192015. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Lemjabbar-Alaoui H, Hassan OU, Yang YW and
Buchanan P: Lung cancer: Biology and treatment options. Biochim
Biophys Acta. 1856:189–210. 2015.PubMed/NCBI
|
|
17
|
Xing Z, Chu C, Chen L and Kong X: The use
of Gene Ontology terms and KEGG pathways for analysis and
prediction of oncogenes. Biochim Biophys Acta. 1860:2725–2734.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Wu KZ, Wang GN, Fitzgerald J, Quachthithu
H, Rainey MD, Cattaneo A, Bachi A and Santocanale C: DDK dependent
regulation of TOP2A at centromeres revealed by a chemical genetics
approach. Nucleic Acids Res. 44:8786–8798. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Klintman M, Buus R, Cheang MC, Sheri A,
Smith IE and Dowsett M: Changes in expression of genes representing
key biologic processes after neoadjuvant chemotherapy in breast
cancer, and prognostic implications in residual disease. Clin
Cancer Res. 22:2405–2416. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Ocaña A, Pérez-Peña J, Alcaraz-Sanabria A,
Sánchez-Corrales V, Nieto-Jiménez C, Templeton AJ, Seruga B,
Pandiella A and Amir E: In silico analyses identify gene-sets,
associated with clinical outcome in ovarian cancer: Role of mitotic
kinases. Oncotarget. 7:22865–22872. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Chen T, Sun Y, Ji P, Kopetz S and Zhang W:
Topoisomerase IIα in chromosome instability and personalized cancer
therapy. Oncogene. 34:4019–4031. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Delgado JL, Hsieh CM, Chan NL and Hiasa H:
Topoisomerases as anticancer targets. Biochem J. 475:373–398. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
An X, Xu F, Luo R, Zheng Q, Lu J, Yang Y,
Qin T, Yuan Z, Shi Y, Jiang W and Wang S: The prognostic
significance of topoisomerase II alpha protein in early stage
luminal breast cancer. BMC Cancer. 18:3312018. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Pei YF, Yin XM and Liu XQ: TOP2A induces
malignant character of pancreatic cancer through activating
β-catenin signaling pathway. Biochim Biophys Acta. 1864:197–207.
2018. View Article : Google Scholar
|
|
25
|
Lara-Gonzalez P, Westhorpe FG and Taylor
SS: The spindle assembly checkpoint. Curr Biol. 22:R966–R980. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Raaijmakers JA, van Heesbeen RGHP, Blomen
VA, Janssen LME, van Diemen F, Brummelkamp TR and Medema RH: BUB1
is essential for the viability of human cells in which the spindle
assembly checkpoint is compromised. Cell Rep. 22:1424–1438. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Ricke RM, Jeganathan KB, Malureanu L,
Harrison AM and van Deursen JM: Bub1 kinase activity drives error
correction and mitotic checkpoint control but not tumor
suppression. J Cell Biol. 199:931–949. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Mukherjee A, Joseph C, Craze M,
Chrysanthou E and Ellis IO: The role of BUB and CDC proteins in
low-grade breast cancers. Lancet. 385 (Suppl 1):S722015. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Stahl D, Braun M, Gentles AJ, Lingohr P,
Walter A, Kristiansen G and Gütgemann I: Low BUB1 expression is an
adverse prognostic marker in gastric adenocarcinoma. Oncotarget.
8:76329–76339. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Hartwell LH, Culotti J and Reid B: Genetic
control of the cell-division cycle in yeast. I. Detection of
mutants. Proc Natl Acad Sci USA. 66:352–359. 1970. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Quek LS, Grasset N, Jasmen JB, Robinson KS
and Bellanger S: Dual role of the anaphase promoting
Complex/Cyclosome in regulating Stemness and differentiation in
human primary keratinocytes. J Invest Dermatol. 138:1851–1861.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Kapanidou M, Curtis NL and Bolanos-Garcia
VM: Cdc20: At the Crossroads between chromosome segregation and
mitotic exit. Trends Biochem Sci. 42:193–205. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Taniguchi K, Momiyama N, Ueda M, Matsuyama
R, Mori R, Fujii Y, Ichikawa Y, Endo I, Togo S and Shimada H:
Targeting of CDC20 via small interfering RNA causes enhancement of
the cytotoxicity of chemoradiation. Anticancer Res. 28:1559–1563.
2008.PubMed/NCBI
|
|
34
|
Jiang J, Jedinak A and Sliva D:
Ganodermanontriol (GDNT) exerts its effect on growth and
invasiveness of breast cancer cells through the down-regulation of
CDC20 and uPA. Biochem Biophys Res Commun. 415:325–329. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Chang DZ, Ma Y, Ji B, Liu Y, Hwu P,
Abbruzzese JL, Logsdon C and Wang H: Increased CDC20 expression is
associated with pancreatic ductal adenocarcinoma differentiation
and progression. J Hematol Oncol. 5:152012. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Choi JW, Kim Y, Lee JH and Kim YS: High
expression of spindle assembly checkpoint proteins CDC20 and MAD2
is associated with poor prognosis in urothelial bladder cancer.
Virchows Arch. 463:681–687. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Kato T, Daigo Y, Aragaki M, Ishikawa K,
Sato M and Kaji M: Overexpression of CDC20 predicts poor prognosis
in primary non-small cell lung cancer patients. J Surg Oncol.
106:423–430. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Karra H, Repo H, Ahonen I, Löyttyniemi E,
Pitkänen R, Lintunen M, Kuopio T, Söderström M and Kronqvist P:
Cdc20 and securin overexpression predict short-term breast cancer
survival. Br J Cancer. 110:2905–2913. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Han Y, Jin X, Zhou H and Liu B:
Identification of key genes associated with bladder cancer using
gene expression profiles. Oncol Lett. 15:297–303. 2018.PubMed/NCBI
|
|
40
|
He X, Zhang C, Shi C and Lu Q:
Meta-analysis of mRNA expression profiles to identify
differentially expressed genes in lung adenocarcinoma tissue from
smokers and non-smokers. Oncol Rep. 39:929–938. 2018.PubMed/NCBI
|
|
41
|
Wang Z, Katsaros D, Shen Y, Fu Y, Canuto
EM, Benedetto C, Lu L, Chu WM, Risch HA and Yu H: Biological and
clinical significance of MAD2L1 and BUB1, genes frequently
appearing in expression signatures for breast cancer prognosis.
PLoS One. 10:e01362462015. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Kato T, Daigo Y, Aragaki M, Ishikawa K,
Sato M, Kondo S and Kaji M: Overexpression of MAD2 predicts
clinical outcome in primary lung cancer patients. Lung Cancer.
74:124–131. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Asmis TR, Ding K, Seymour L, Shepherd FA,
Leighl NB, Winton TL, Whitehead M, Spaans JN, Graham BC and Goss
GD; National Cancer Institute of Canada Clinical Trials Group, :
Age and comorbidity as independent prognostic factors in the
treatment of non small-cell lung cancer: A review of National
Cancer Institute of Canada Clinical Trials Group trials. J Clin
Oncol. 26:54–59. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Zou ZQ, Du YY, Sui G and Xu SN: Expression
of TS, RRM1, ERCC1, TUBB3 and STMN1 genes in tissues of non-small
cell lung cancer and its significance in guiding postoperative
adjuvant chemotherapy. Asian Pac J Cancer Prev. 16:3189–3194. 2015.
View Article : Google Scholar : PubMed/NCBI
|