Introduction
Parkinson's disease (PD) is a common, progressive
degenerative disease of the central nervous system that causes loss
of steady motor function (1). PD is
clinically characterized by resting tremor, bradykinesia, rigidity,
impairment of balance, akinesia, postural reflex loss and other
non-motor symptoms (1-4).
The pathophysiological changes of PD are associated with the death
of dopaminergic neurons in the substantia nigra pars compacta
(SNpc), which subsequently leads to a lack of dopamine production
and/or release in the striatum and motor impairment (5).
In addition to traditional pathological findings,
such as intracellular inclusions containing α-synuclein during
post-mortem diagnosis (6,7), there is evidence to indicate that PD is
an inflammatory disease. Microglia are activated and express human
leukocyte antigen DR isotype molecules in patients with PD
(8). T-cell infiltration, which is
absent in normal brain tissue, has been observed in samples from
patients with PD (9,10), indicating that T cells might play a
role during the inflammatory pathogenesis of PD.
Inflammation observed in patients with PD has raised
questions, such as how T cells migrate through the blood-brain
barrier, and whether blocking T-cell infiltration can attenuate and
slow the progression of PD (11,12).
Cellular adhesion molecules such as intercellular adhesion molecule
1 (ICAM1 or CD54) are upregulated during inflammation in the
central nervous system and play an important role in T-cell
recruitment (11,12). However, the involvement of ICAM1 in
the pathogenesis of PD remains to be elucidated. The present study
investigated whether ICAM1 and its ligand lymphocyte
function-associated antigen 1 (LFA-1) interact with each other to
mediate T-cell infiltration. The results revealed that T-cell
infiltration was evident in patients with PD and mice with
experimental PD, depletion of CD4+ T cells or
CD8+ T cells attenuated the severity of experimental PD,
ICAM1 was upregulated in the brain tissue of experimental PD mice,
and blocking the ICAM1-LFA1 axis reduced T-cell infiltration and
the severity of PD in experimental mice.
Materials and methods
Ethics consideration
The ethics committee of Ningbo First Hospital
approved and supervised the proposed experimental protocol
(approval no: 2017-R044). Written consent forms were explained,
agreed to and signed by the next-of-kin of deceased patients with
PD and patients without central nervous system disease.
Histological diagnosis of PD
Brain tissues were obtained from post-mortem
individuals (PD patients, 4 males and 4 females; age range, 67-89
years) admitted to Ningbo First Hospital between January and June
2017. Sex- and age-matched patients without PD served as controls.
Brain tissues were processed as frozen sections. Serial sections
(4-6 µm) were cut. Slides were fixed with 10% formalin for 30 min
at room temperature (RT), and stained with hematoxylin and eosin
(Sigma-Aldrich; Merck KGaA) according to the manufacturer's
instructions. PD diagnosis was confirmed for patients with PD or
excluded for control patients in line with a published criterion by
two experienced pathologists (13).
Immunofluorescence staining
Brain tissue obtained from the SNpc of human or
mouse samples was processed as frozen sections and cut into serial
sections of thickness 4-6 µm. Slides were fixed in 10% formalin for
30 min at RT and blocked with 10% goat serum (Gibco; Thermo Fisher
Scientific, Inc.) for 30 min at RT. Samples were then incubated
with fluorescence-conjugated antibodies for 30 min at RT, which are
listed in Table I. The number of
tyrosine hydroxylase (TH)-producing cells in the lesions of the
brain was used as an indicator for disease severity as previously
reported (10). Briefly,
histological sections were stained with anti-TH primary antibodies
for 30 min at RT (Table I). After
washing with PBS, slices were stained with anti-rabbit
AF647-conjugated secondary antibodies (1:1,000, Thermo Fisher
Scientific, Inc.; cat. no. A32733) for 30 min at RT.
 | Table IAntibodies used for immunostaining
and flow cytometry. |
Table I
Antibodies used for immunostaining
and flow cytometry.
| Antibody
(clone) | Manufacturer (cat.
no.) | Utilization | Dilution |
|---|
| Tyrosine
hydroxylase (TH) | Abcam (ab112) | IF | 1:1,000 |
| CD8-PE
(53-6.7) | eBioscience; Thermo
Fisher Scientific, Inc. (12-0081-82) | IF, FC | 1:100 |
| CD4-PE (RM4-5) | eBioscience; Thermo
Fisher Scientific, Inc. (12-0042-82) | IF, FC | 1:100 |
| CD45-APC
(30-F11) | eBioscience; Thermo
Fisher Scientific, Inc. (17-0451-82) | FC | 1:100 |
| CD31-PE (390) | eBioscience; Thermo
Fisher Scientific, Inc. (12-0311-82) | FC | 1:100 |
| ICAM1-FITC
(YN1/1.7.4) | eBioscience; Thermo
Fisher Scientific, Inc. (11-0541-82) | FC | 1:100 |
| CD3-PE
(145-2C11) | eBioscience; Thermo
Fisher Scientific, Inc. (12-0031-82) | FC | 1:100 |
| LFA1-FITC
(M18/2) | eBioscience; Thermo
Fisher Scientific, Inc. (11-0181-82) | FC | 1:100 |
Experimental PD induction
A total of 395 Male C57/Bl6 mice, aged 10-12 weeks
old, were purchased from the Jackson Laboratory. Mice were kept in
a specific pathogen-free environment with controlled temperature
(23˚C) and humidity (40-60%), under a 12-h light/dark cycle. Mice
were allowed ad libitum access to food and water.
Experimental PD was induced by intraperitoneally injecting
1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine (MPTP; Sigma-Aldrich;
Merck KGaA) according to a published protocol (10). Briefly, MPTP (20 mg/kg) in 100 ml of
saline was intraperitoneally injected 4 times at 2-h intervals, and
mice were sacrificed by cervical dislocation under general
anesthesia by isoflurane inhalation (oxygen flow, 0.8 l/min;
isoflurane vaporizer, 3%) at designated time points (4 days after
MPTP injection) following the last injection. Control mice received
100 ml saline via intraperitoneal injection. A total of 9 mice per
group were utilized in each experiment, which was repeated in
triplicate.
Open field tests
Behavioral assessments were performed 4 days
following MPTP injection using open field tests, which are the most
common tests for behavior of MPTP-treated PD mice (14). The experimental procedure was adapted
from a previous publication (15).
All tests were performed between 12:00 and 2:00 p.m. in normal
lighting. Briefly, a white plastic rectangular box with dimensions
of 80x40x20 cm was utilized for open field tests. The bottom of the
box was drawn with a grid of 5x5-cm2 squares. The total
distance that a mouse moved was manually measured by counting the
number of squares. Rearing, which represents exploratory activity,
and grooming, which indicates displacement response, were scored
when a mouse was placed in the open field for 5 min (16).
Neutralizing antibodies
The abundance of CD8+ and CD4+
T cells as well as the efficacy of deletion of these two cell types
in the brain tissue were determined by immunofluorescence.
Circulating CD8+ and CD4+ T cells were also
measured by flow cytometry when neutralizing antibodies were
applied. To deplete CD8+ T cells, an anti-CD8 antibody
(clone 2.43; Bio X cell; cat. no. BP0061) at a dose of 250 µg in
100 µl PBS was intraperitoneally injected at days -7 and -3 before
MPTP injection (day 0). Control mice received isotype control
antibodies (rat anti-mouse IgG2b; Bio X Cell; cat. no. BP0090)
following the same regimen as CD8+ T cell depletion.
To deplete CD4+ T cells, anti-CD4
antibodies (cloneGK1.5; Bio X Cell; cat. no. BP0003-1) were
administered at a dose of 250 µg in 100 µl PBS by intraperitoneal
injection at days-7 and -3. Control mice received isotype control
antibodies (rat anti-mouse IgG2b; Bio X Cell; cat. no. BP0090)
using the same regimen of anti-CD4 antibodies.
Cell type-specific expression of ICAM-1 and LFA1 was
determined by flow cytometry. For blocking of ICAM1, anti-ICAM1
antibodies (clone YN1/1.7.4; Thermo Fisher Scientific, Inc.) were
administered at a dose of 4 mg/kg by intraperitoneal injection at
days-7 and -3. Control mice received mouse IgG2bκ isotype control
antibodies (Thermo Fisher Scientific, Inc.) following the same
regimen as ICAM-1 blockade. For blocking of LFA1, anti-CD11a
antibodies (clone M17/4; Thermo Fisher Scientific, Inc.) were
intraperitoneally injected at a dose of 100 µg at days-7 and -3.
Control mice received mouse IgG2bκ isotype control antibodies
following the same regimen as LFA-1blockade. A total of 9 mice per
group were utilized in each experiment, which was repeated in
triplicate.
Flow cytometry
Mouse brain tissue was processed using a commercial
kit for brain tissue dissociation (Miltenyi Biotec, Inc.; cat. no.
130-107-677) to obtain single-cell suspensions according to the
manufacturer's instructions. Circulating T cells were obtained by
lysing red blood cells for 5 min at RT and centrifugation at 300 x
g for 10 min at RT. Splenic T cells were obtained by mechanical
isolation using a cell strainer and centrifugation at 300 x g for
10 min at RT.
Cells were treated with anti-CD16/32 antibody
(1:200; Thermo Fisher Scientific, Inc.; cat. no. 14-0161) for 10
min at RT, and then stained with the designated antibodies listed
in Table I. Following incubation
with the corresponding antibodies, cells were analyzed with an
Attune NxT flow cytometer (Thermo Fisher Scientific, Inc.). Data
were presented and analyzed using Kaluza 1.3 (Beckman Coulter,
Inc.).
Statistical analysis
Data are presented as the mean ± SD. Data were
compared using Student's t-test for two groups or one-way ANOVA
with Bonferroni's post hoc test for multiple groups. Associations
between the abundance of CD8+ or CD4+ T cells
and severity of PD were analyzed using linear regression. P<0.05
was considered to indicate a statistically significant difference.
Statistical analysis was performed using GraphPad Prism 7 (GraphPad
Software, Inc.).
Results
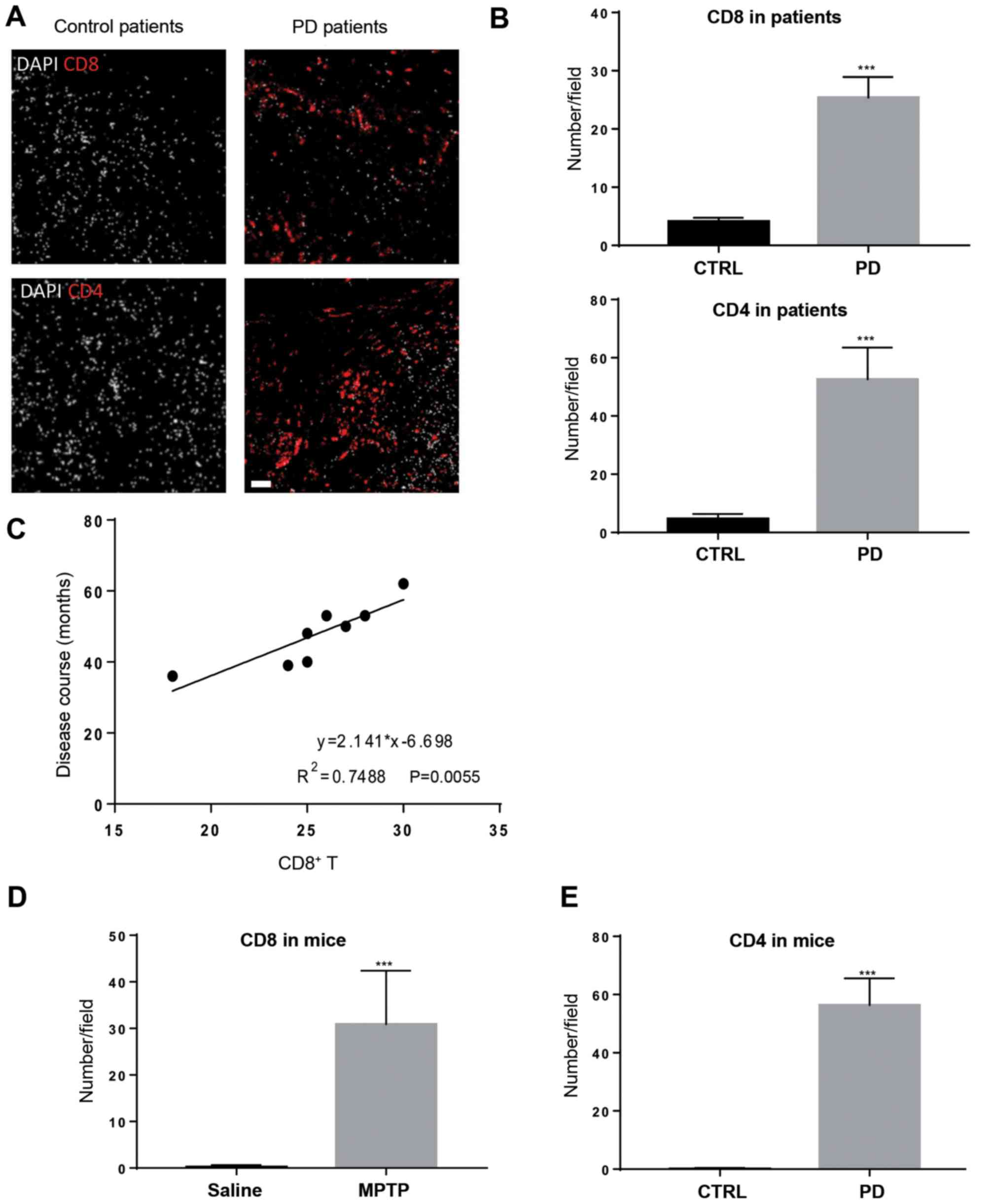
T-cell infiltration is observed in the
brain tissue of PD patients and MPTP-intoxicated mice
To explore T-cell infiltration in the brain tissue
of patients with PD, post-mortem samples were used for
immunofluorescence staining. In agreement with previous reports
(10,17), CD8+ T cells and
CD4+ T cells were observed in the brain tissue of
patients with PD, but were expressed at a significantly lower level
in patients without PD (Fig. 1A and
B). CD8+ T-cell
infiltration positively associated with disease duration (Fig. 1C), suggesting that CD8+
T-cell infiltration might contribute to disease severity.
Subsequently, the MPTP-intoxicated mouse model of PD was used to
study T-cell recruitment in an experimental setting. Mice treated
with MPTP gradually manifested symptoms of hypoactivity beginning
at 12 h after MPTP injection. Four days after MPTP injection, mice
treated with MPTP also harbored CD4+ and CD8+
T cells in the brain tissue, mainly in the SNpc and striatum
regions (Figs. 1D and E; S1);
whereas these two types of immune cells were not evident in
saline-treated mice. These results indicate that T-cell
infiltration is a feature in patients with PD and experimental PD
mice.
Depletion of CD4+ T cells
or CD8+ T cells attenuates experimental PD
Next, it was determined whether the disease severity
in the mouse model of PD was CD4+ T cell- or
CD8+ T cell-dependent. Neutralizing antibodies or
matched isotype control antibodies were used to deplete
CD4+ T cells or CD8+ T cells in MPTP-treated
mice. The efficacy of CD4+ or CD8+ T-cell
depletion was determined by measuring circulating T cells using
flow cytometry (data not shown). Depletion of CD4+ T
cells and CD8+ T cells in the lesions of MPTP-treated
mice was also confirmed by immunofluorescence (Fig. 2A). As shown in Fig. 2B and C, depletion of CD4+ T cells
attenuated disease severity, as indicated by the increased number
of TH+ cells at 4 days after MPTP injection compared
with isotype control antibody-treated mice. The behaviour of the
mice was also observed using open field tests. As shown in Fig. 2D-F, depletion of CD4+
attenuated behavioral dysfunctions in MPTP-treated mice compared
with isotype control antibody-treated mice. Similar results were
also observed in mice treated with anti-CD8 neutralizing antibodies
(Fig. 2C-F). These data suggest that
CD4+ and CD8+ T cells play a detrimental role
in the pathogenesis of experimental PD.
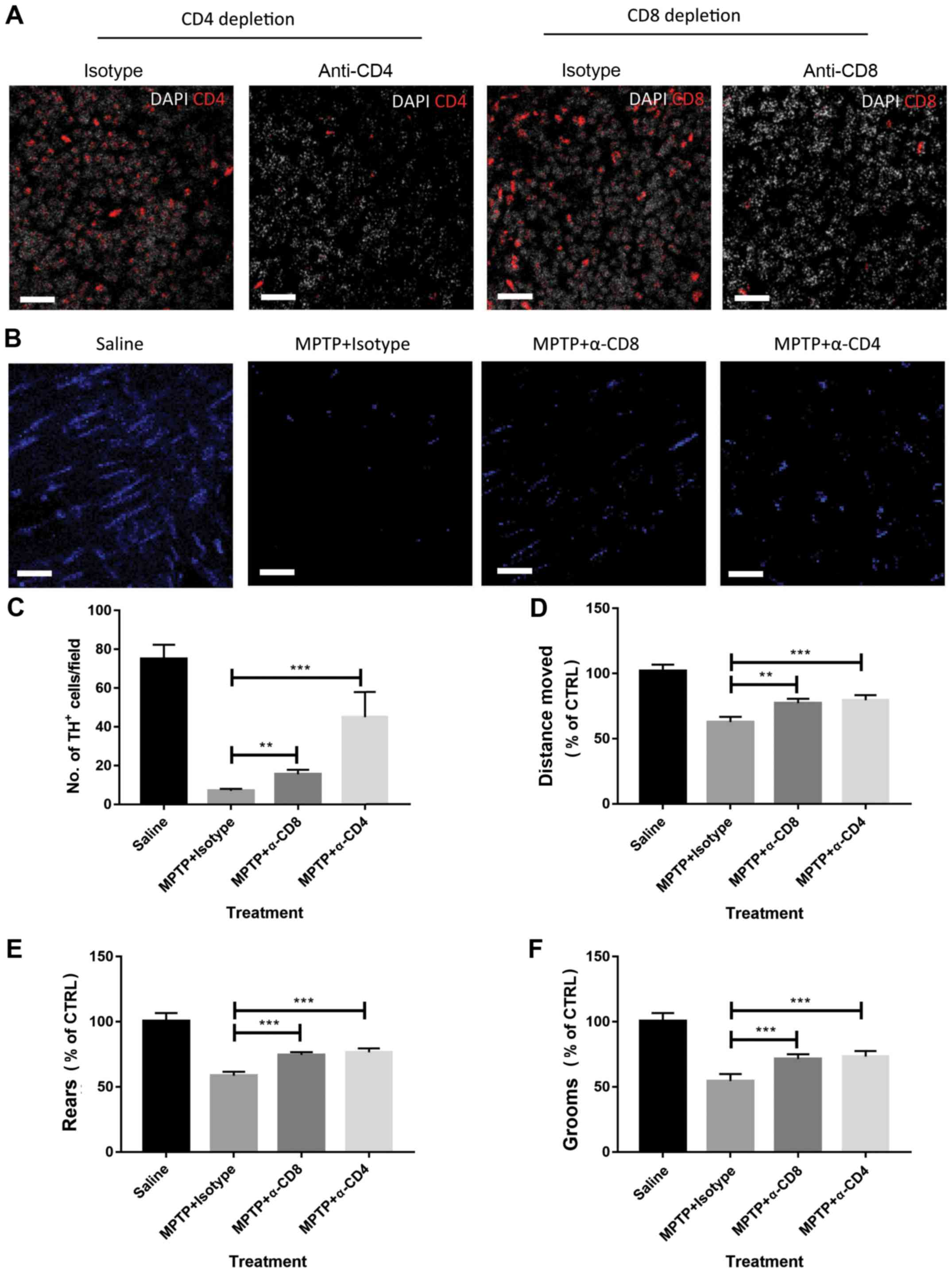 | Figure 2Depletion of CD8+ or
CD4+ T cells attenuates MPTP-induced brain damage. (A)
Depletion of CD4+ and CD8+ T cells in
MPTP-treated mice was confirmed by immunofluorescence. Scale bars,
20 µm. (B) Representative images of TH+ cell staining
(visualized by AF-647 secondary antibodies, blue in the images) in
the brain tissue of mice. Scale bars, 20 µm. (C) Depletion of
CD8+ or CD4+ T cells attenuated the severity
of MPTP-induced Parkinson's disease in mice as measured by the
abundance of TH+ cells. Mouse behavior, including (D)
distance moved, (E) rears and (F) grooms were assessed by open
field tests. MPTP-treated mice showed impaired behavior, but
depletion of CD4+ or CD8+ T cells attenuated
behavioral dysfunction. n=9. Experiments were performed in
triplicate. **P<0.01 and ***P<0.001.
MPTP, 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine; CTRL, control;
TH, tyrosine hydroxylase; α-CD4, anti-CD4 antibodies; α-CD8,
anti-CD8 antibodies. |
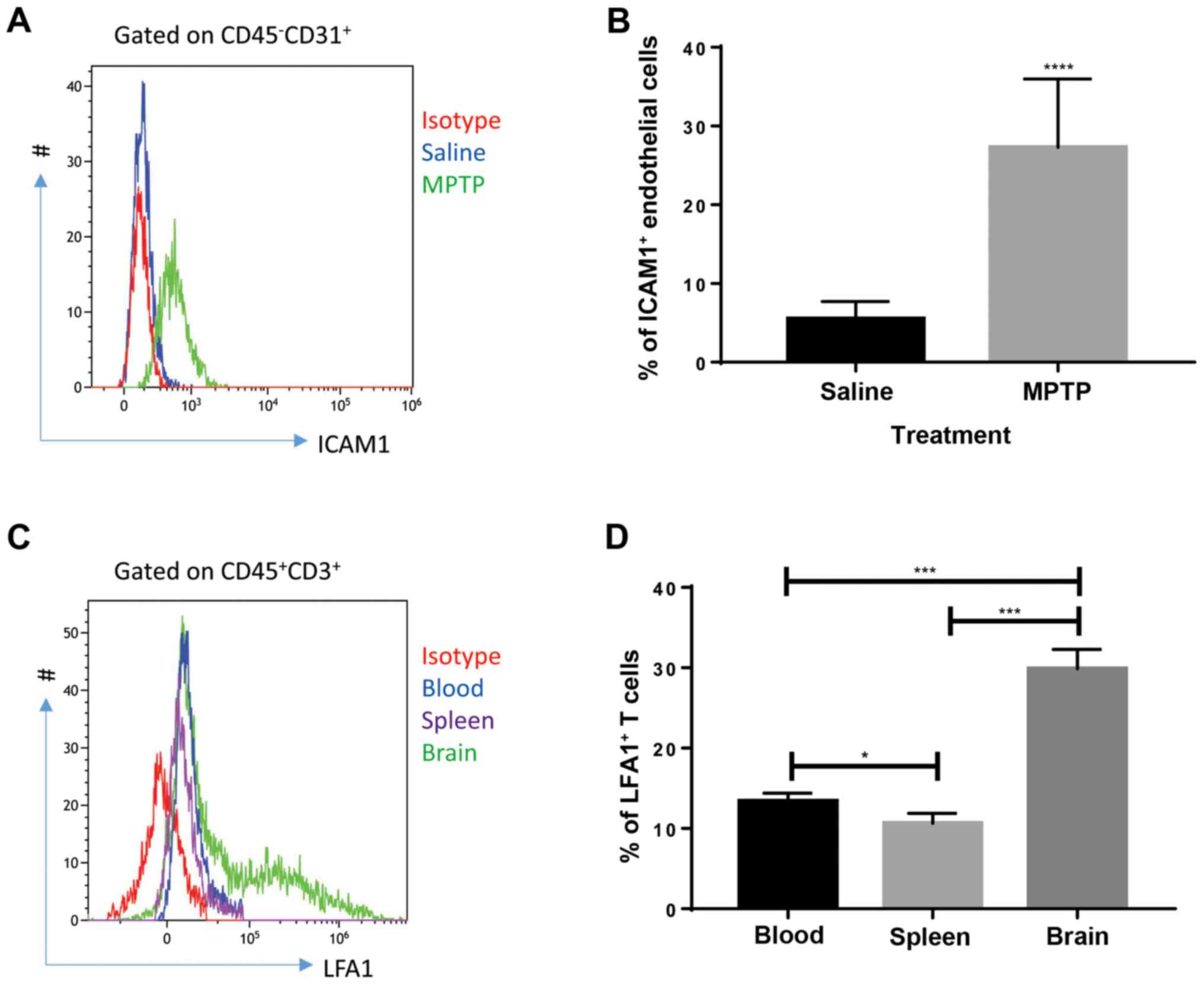
ICAM1 expression in endothelial cells
and LFA1 expression in T cells are upregulated in MPTP-treated
mice
Cellular adhesion molecules on endothelial cells
play a role in the initiation of leukocyte infiltration through the
blood-brain barrier (18). Thus, the
expression of cellular adhesion molecules was investigated.
According to the literature, ICAM1 is critical for T-cell
recruitment in inflammatory brain conditions; LFA1, a major ligand
of ICAM1, expressed by T cells interacts with ICAM1 to mediate
T-cell infiltration in the brain during a number of conditions
(11,12). Therefore, the expression of ICAM1 in
endothelial cells and LFA1 in T cells were investigated. Flow
cytometry results showed that endothelial cells (defined as
CD45-CD31+ cells) collected from the brain of
MPTP-treated mice expressed high levels of ICAM1 (Fig. 3A and B) compared with saline-treated mice at day
4 after MPTP injection. Since saline-treated mice presented with
very low T-cell infiltration in the brain, T cells collected from
the brain, blood and spleen of MPTP-treated mice were analyzed. As
shown in Fig. 3C and D, T cells collected from the brain
expressed increased levels of LFA1 compared with circulating and
splenic T cells, indicating that upregulation of LFA1 on T cells
was specific to brain inflammation at day 4 after MPTP injection.
These results indicate that the interaction between endothelial
ICAM1 and LFA1 expressed by T cells might be involved in the
process of MPTP-induced PD.
Blocking ICAM1 or LFA1 alleviates
MPTP-induced PD in mice
Whether blocking the ICAM1/LFA1 axis could be used
as a therapeutic strategy for experimental PD was investigated.
Antibodies neutralizing ICAM1 or CD11a (a subunit of LFA1) were
administered to MPTP-treated mice, while matched isotype control
antibodies were used as controls. Blocking ICAM1 or its ligand LFA1
significantly reduced disease severity in MPTP-challenged mice as
measured by the number of TH+ cells (Figs. 4A; S2) and behavior (Fig. 4B-D) compared with isotype
antibody-administered mice. Subsequently, anti-ICAM1 and anti-CD11a
antibodies were co-administered to experimental animals; no
synergistic effects were observed (data not shown), indicating the
possibility that the interaction between ICAM1 and LFA1 in this
setting might be specific with no involvement of any other
receptors or ligands. Therefore, blocking ICAM1 or LFA1 may have
therapeutic potential in PD.
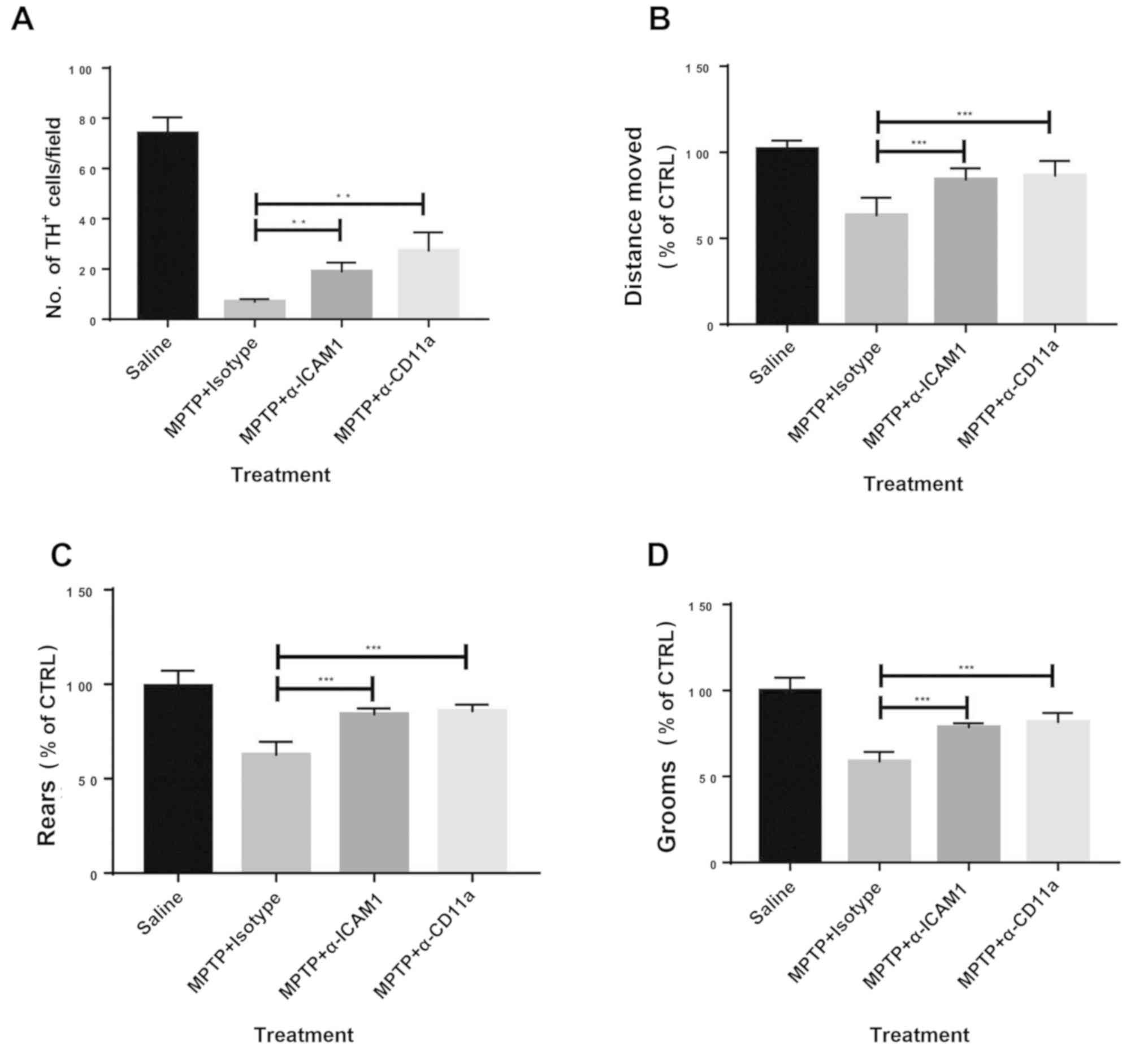 | Figure 4Blocking of ICAM1 or CD11a attenuates
the severity of MPTP-induced PD in mice. (A) Blocking of ICAM1 or
CD11a increased the levels of TH+ cells at day 4 after
MPTP injection. Mouse behaviour assessed by open field tests showed
improved clinical presentation in those treated with ICAM1 or CD11a
blockade at day 4 after MPTP injection, including (B) distance
moved, (C) rears and (D) grooms. n=9. Experiments were performed in
triplicate. **P<0.01 and ***P<0.001.
ICAM1, intercellular adhesion molecule 1; MPTP,
1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine; TH, tyrosine
hydroxylase; CTRL, control; α-ICAM1, anti-ICAM1 antibodies;
α-CD11a, anti-CD11a antibodies. |
Discussion
The present study demonstrated that T cells play a
role during the pathogenesis of PD. The results indicate that
T-cell infiltration is a universal feature in patients with PD and
experimental PD mice. Depletion of CD4+ or
CD8+ T cells by the administration of neutralizing
antibodies attenuated the severity of experimental PD in mice as
assessed by the abundance of TH+ cells in the brain
tissue and behavioral assessment by open field tests. Moreover,
endothelial ICAM1 and LFA1 expressed on T cells appear to be
involved during the process of T cell recruitment, as inhibiting
T-cell recruitment by blocking the ICAM1-LFA1 axis alleviated
experimental PD. Therefore, the present study highlighted the
therapeutic potential of using strategies where the ICAM1-LFA1 axis
is inhibited for treatment of PD.
PD is a progressive degenerative disease of the
central nervous system that affects millions of individuals
worldwide (1-4).
The precise etiology of PD has not yet been fully elucidated, where
the loss of dopamine-producing neurons in the SNpc mediates the
pathogenesis (10). A growing body
of evidence, including findings in the present study, indicate that
aberrant immune responses in the brain contribute to the
pathogenesis of PD (1-4,10).
It has been reported that T cells are recruited into the SNpc
during experimental PD (9,10). T-cell infiltration was also observed
in the brain tissue of patients with PD and mice with experimental
PD in the present study. Considering that dysregulated inflammation
in the brain occurs in other central nervous system diseases such
as Alzheimer's disease and multiple sclerosis (1-4),
more research should be focused on the immunopathogenesis of these
brain conditions and how to manipulate the aberrant inflammatory
responses in order to treat these diseases.
Interactions between ICAM1 and LFA1 are critical
during the recruitment of leukocytes to inflamed sites (19). The ICAM1-LFA1 axis is overactivated
in patients with PD (20). Although
ICAM1 or LFA1 expression was not assessed in the human cohort of
the present study, it is possible that PD patients in the present
cohort have high expressions of ICAM1 and LFA1, although this
warrants further examination. The present study found that
endothelial ICAM1 is upregulated in mice with experimental PD
compared with control mice. The role of the ICAM1-LFA1 axis in PD
was also investigated. Notably, the aforementioned study (20) did not conduct functional assays in
terms of disease severity when the axis is inhibited, whereas the
present data indicated the roles of the ICAM1-LFA1 axis in the
pathogenesis of PD. T cells in the SNpc of mice treated with MPTP
expressed increased levels of LFA1 compared with T cells harvested
from the peripheral circulation or the spleen, indicating that T
cells in the brain tissue of mice with PD are more activated. In
the process of experimental PD, endothelial cells in the brain
tissue harbor increased ICAM1 expression. Circulating T cells in
the inflamed area of the brain upregulate the expression of LFA1 to
interact with ICAM1 expressed by endothelial cells. This
interaction further activates T cells to begin the multiple-step
process of T-cell recruitment (21),
including capture, rolling, adhesion and transmigration, which
involves the interaction between ICAM1 and LFA1. However, the
present study did not investigate in which of the aforementioned
steps the interaction between ICAM1 and LFA1 plays the most
prominent effect. Advanced imaging techniques such intravital
microscopy will provide a means to trace each step of leukocyte
recruitment in order to elucidate the precise influence of the
ICAM1-LFA1 axis on PD pathogenesis.
T-cell infiltration is a hallmark of PD in humans
and experimental PD in mice (10).
Depletion of CD4+ or CD8+ T cells resulted in
improved disease status, indicating that T cells in the brain
tissue are detrimental. The current study speculates that there
might be several theories as to how T cells execute pathological
effects in PD. First, CD8+ T cells exert cytotoxic
effects in the setting of PD to mediate cell death in the inflamed
brain tissue. Second, CD4+ T cells, possibly T helper
(Th) 1 and Th17 cells, produce pro-inflammatory cytokines such as
tumor necrosis factor (TNF)-α to increase apoptosis of neurons.
Pathogenic Th17 cells expressing TNF-α and IL17 have been observed
in other inflammatory conditions (22,23), and
might also contribute to the pathogenesis of PD. Chemokines
secreted by T cells might recruit other leukocytes such monocytes
and neutrophils to perform cell-killing effects. It is worth noting
that more studies are warranted for the precise detrimental roles
of T cells in the pathogenesis of PD.
A limitation of the present study is that the levels
of CD4+ or CD8+ T cells in the central
nervous system when the ICAM1-LFA1 axis is inhibited were not
measured. However, the present study showed that when the
ICAM1-LFA1 axis is inhibited or when T cells are depleted, the
disease severity is attenuated. Hence, it is hypothesized that the
abundance of T cells in the brain lesions would decrease when the
ICAM1-LFA1 axis is inhibited. The roles of the axis in T-cell
recruitment during the pathogenesis of PD are worthy of
exploration.
Intervention in the ICAM1-LFA1 axis has possible
therapeutic benefits for patients with PD. The depletion of T cells
and also inhibition of T-cell recruitment attenuated the severity
of PD, highlighting the potential of intervention in the ICAM1-LFA1
axis in a clinical setting. Blocking T-cell or other leukocyte
recruitment has been shown to be effective in a number of
inflammatory conditions, including inflammatory bowel disease and
multiple sclerosis, and so it may also be a possible therapeutic
strategy for patients with PD.
In conclusion, ICAM1 expressed in endothelial cells
and LFA1 expressed in T cells play an essential role in T cell
recruitment during experimental PD in mice.
Supplementary Material
Representative immunofluorescence
images of CD8+ and CD4+ T cells in the brain
of saline (left panels) and
1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine (Right panels) treated
mice. (A) CD8+ and (B) CD4+ T cells. Scale
bars, 25 μm.
Representative immunofluorescence
image of TH+ cells (stained blue) in MPTP-treated mice
during the blocking of ICAM1 or CD11a. Scale bars, 20 μm. ICAM1,
intercellular adhesion molecule 1; MPTP,
1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine; TH, tyrosine
hydroxylase; α-ICAM1, anti-mouse ICAM1 antibodies; α-CD11a,
anti-mouse CD11a antibodies.
Acknowledgements
Not applicable.
Funding
The current study was supported by Zhejiang Province
Chinese Medicine Science Projects (grant no. 2018ZA108).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
WL performed the majority of experiments and
prepared the first draft of the manuscript. SC and YL performed
histological, immunofluorescence and flow cytometry experiments. YX
and QM performed statistical analysis. QY and JW conceptualized and
supervised the current study. All authors read and approved the
final manuscript.
Ethics approval and consent to
participate
The ethics committee of Ningbo First Hospital
approved and supervised the proposed experimental protocol
(approval no: 2017-R044). Written informed consent was provided by
the next-of-kin of the participants.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Jankovic J: Parkinson's disease: Clinical
features and diagnosis. J Neurol Neurosurg Psychiatry. 79:368–376.
2008.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Noorian AR, Rha J, Annerino DM, Bernhard
D, Taylor GM and Greene JG: Alpha-synuclein transgenic mice display
age-related slowing of gastrointestinal motility associated with
transgene expression in the vagal system. Neurobiol Dis. 48:9–19.
2012.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Chaudhuri KR and Schapira AH: Non-motor
symptoms of Parkinson's disease: Dopaminergic pathophysiology and
treatment. Lancet Neurol. 8:464–474. 2009.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Magerkurth C, Schnitzer R and Braune S:
Symptoms of autonomic failure in Parkinson's disease: Prevalence
and impact on daily life. Clin Auton Res. 15:76–82. 2005.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Fasano M and Lopiano L: Alpha-synuclein
and Parkinson's disease: A proteomic view. Expert Rev Proteomics.
5:239–248. 2008.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Tofaris GK, Razzaq A, Ghetti B, Lilley KS
and Spillantini MG: Ubiquitination of alpha-synuclein in Lewy
bodies is a pathological event not associated with impairment of
proteasome function. J Biol Chem. 278:44405–44411. 2003.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Spillantini MG, Schmidt ML, Lee VM,
Trojanowski JQ, Jakes R and Goedert M: Alpha-synuclein in Lewy
bodies. Nature. 388:839–880. 1997.PubMed/NCBI View
Article : Google Scholar
|
|
8
|
Tansey MG, McCoy MK and Frank-Cannon TC:
Neuroinflammatory mechanisms in Parkinson's disease: Potential
environmental triggers, pathways, and targets for early therapeutic
intervention. Exp Neurol. 208:1–25. 2007.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Saunders JA, Estes KA, Kosloski LM, Allen
HE, Dempsey KM, Torres-Russotto DR, Meza JL, Santamaria PM, Bertoni
JM, Murman DL, et al: CD4+ regulatory and effector/memory T cell
subsets profile motor dysfunction in Parkinson's disease. J
Neuroimmune Pharmacol. 7:927–938. 2012.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Brochard V, Combadière B, Prigent A,
Laouar Y, Perrin A, Beray-Berthat V, Bonduelle O, Alvarez-Fischer
D, Callebert J, Launay JM, et al: Infiltration of CD4+ lymphocytes
into the brain contributes to neurodegeneration in a mouse model of
Parkinson disease. J Clin Invest. 119:182–192. 2009.PubMed/NCBI View
Article : Google Scholar
|
|
11
|
Jiang L, Hu J, Feng J, Han D and Yang C:
Substrate stiffness of endothelial cells directs LFA-1/ICAM-1
interaction: A physical trigger of immune-related diseases? Clin
Hemorheol Microcirc. 61:633–643. 2016.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Harrer A, Pilz G, Wipfler P, Oppermann K,
Sellner J, Hitzl W, Haschke-Becher E, Afazel S, Rispens T, van der
Kleij D, et al: High interindividual variability in the CD4/CD8 T
cell ratio and natalizumab concentration levels in the
cerebrospinal fluid of patients with multiple sclerosis. Clin Exp
Immunol. 180:383–392. 2015.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Dickson DW: Parkinson's disease and
parkinsonism: neuropathology. Cold Spring Harb Perspect Med.
2(a009258)2012.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Meredith GE and Rademacher DJ: MPTP mouse
models of Parkinson's disease: An update. J Parkinsons Dis.
1:19–33. 2011.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Luchtman DW, Shao D and Song C: Behavior,
neurotransmitters and inflammation in three regimens of the MPTP
mouse model of Parkinson's disease. Physiol Behav. 98:130–138.
2009.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Deacon RM, Koros E, Bornemann KD and
Rawlins JN: Aged Tg2576 mice are impaired on social memory and open
field habituation tests. Behav Brain Res. 197:466–468.
2009.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Chandra G, Rangasamy SB, Roy A, Kordower
JH and Pahan K: Neutralization of RANTES and eotaxin prevents the
loss of dopaminergic neurons in a mouse model of parkinson disease.
J Biol Chem. 291:15267–15281. 2016.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Krummel MF, Bartumeus F and Gérard A: T
cell migration, search strategies and mechanisms. Nat Rev Immunol.
16:193–201. 2016.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Walling BL and Kim M: LFA-1 in T cell
migration and differentiation. Front Immunol. 9(952)2018.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Miklossy J, Doudet DD, Schwab C, Yu S,
McGeer EG and McGeer PL: Role of ICAM-1 in persisting inflammation
in Parkinson disease and MPTP monkeys. Exp Neurol. 197:275–283.
2006.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Rivera-Nieves J, Gorfu G and Ley K:
Leukocyte adhesion molecules in animal models of inflammatory bowel
disease. Inflamm Bowel Dis. 14:1715–1735. 2008.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Ivanov II, Atarashi K, Manel N, Brodie EL,
Shima T, Karaoz U, Wei D, Goldfarb KC, Santee CA, Lynch SV, et al:
Induction of intestinal Th17 cells by segmented filamentous
bacteria. Cell. 139:485–498. 2009.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Ivanov II, Frutos Rde L, Manel N,
Yoshinaga K, Rifkin DB, Sartor RB, Finlay BB and Littman DR:
Specific microbiota direct the differentiation of IL-17-producing
T-helper cells in the mucosa of the small intestine. Cell Host
Microbe. 4:337–349. 2008.PubMed/NCBI View Article : Google Scholar
|