Intracellular pH (pHi) is usually maintained between
6.9-7.2; it is important to maintain a normal pHi as a decrease in
pHi may influence the normal functioning of proteins, ion channels
and several other physiological processes involved in cell
proliferation, division and differentiation (1). Some ion exchanger families regulate
proton flux at the plasma membrane, such as
Na+/H+ exchanger (NHE), Cl-/HCO3-
exchanger, Na+/HCO3- cotransporter and
Na+-driven Cl-/HCO3- exchanger
(2). The present review focused on
the NHE family. NHEs are pH-regulated membrane proteins that
exchange extracellular Na+ for intracellular H+ with a
stoichiometry of one for one (2).
The inward Na+ gradient produced by
Na+/K+-ATPase provides a constant driving
force for H+ efflux (3).
Moreover, NHEs are activated by decreases in pHi, and thus are
likely to respond to an increase in proton load during acute acid
stimulation (4).
NHEs are evolutionarily conserved membrane
transporters; the solute carrier 9A (SLC9A) family contains the
well-characterized plasma membrane and intracellular NHE isoforms
(NHE1-9), and the SLC9B subgroup consists of
Na+/H+ antiporter 1 and
Na+/H+ antiporter 2(5). Different NHE isoforms are positioned
differently. NHE1 is the ‘housekeeping’ isoform in NHE family
(6) and is nearly ubiquitous in the
plasma membrane of almost all tissues (7), while the other isoforms have more
restricted localization and function. The specific localization of
NHE1 may vary depending on cell type. In fibroblasts, NHE1 is
mainly localized in lamellae and participates in migration and
anchoring (8). However, in
epithelial cells, NHE1 is distributed in the basolateral membrane
(9). NHE2-5 are also localized to
the plasma membrane, but have more restricted tissue distributions
(10). For example, NHE2 is an
apical membrane protein found mainly in the stomach and intestines
(11). Both NHE3 and NHE4 are highly
expressed in the kidney and gastrointestinal tract (12). In the gastrointestinal tract, NHE3 is
mainly expressed in the intestine, while NHE4 is expressed in the
stomach (12). Moreover, NHE5 is
expressed primarily in the brain (13). The isoforms NHE6-NHE9 exist in
intracellular organelles, where they participate in the maintenance
of pHi (14). NHE6 is expressed in
early recycling endosomes and mitochondria, NHE7 is located in the
trans-Golgi network, NHE8 is in the mid- to trans-Golgi and NHE9 is
expressed in late recovered endosomes (15,16).
Various subtypes of NHEs are related to the
pathogenesis of digestive diseases, such as Barrett's esophagus
(17), gastric cancer (18), IBD (19), colon cancer (20) or liver diseases. Currently, the vast
majority of review articles have focused on the role of the NHE
family members in IBD (21),
intestinal infectious diarrhea (22)
and digestive system tumorigenesis (23), and to the best of our knowledge, only
a few have reported the role of NHEs in liver disease. The liver is
the largest digestive gland in the human digestive system, and
plays an important role in metabolism, deoxygenation, glycogen
storage and secretory protein synthesis (24). Disease development in the liver
seriously affects the normal function of the body (25). Thus, it is important to study the
physiological and pathological regulation of the liver. However,
the role of NHEs in liver function is not fully understood,
although all NHEs except NHE5 have been detected in this organ
(26).
Therefore, the present review details the physiology
and pathology of NHEs in the liver, including the regulation of
hepatocyte volume, hepatocyte growth, regeneration, proliferation,
apoptosis, bile formation and other physiological activities. The
pathologies discussed include non-alcoholic fatty liver disease
(NAFLD), liver fibrosis, liver cancer and other liver diseases.
The sequences of the nine subtypes of NHEs in the
SLC9A subfamily are significantly different, with amino acid
identities ranging between <12% (NHE1 vs. NHE9) and >70%
(NHE6 vs. NHE7) (27). Despite these
differences, silico-predicted transmembrane protein domains have
suggested very similar structural arrangements for all nine
isoforms, which have a high degree of similarity in the
NH2-terminal hydrophobic domain, which contains multiple predicted
membrane-spanning segments (27).
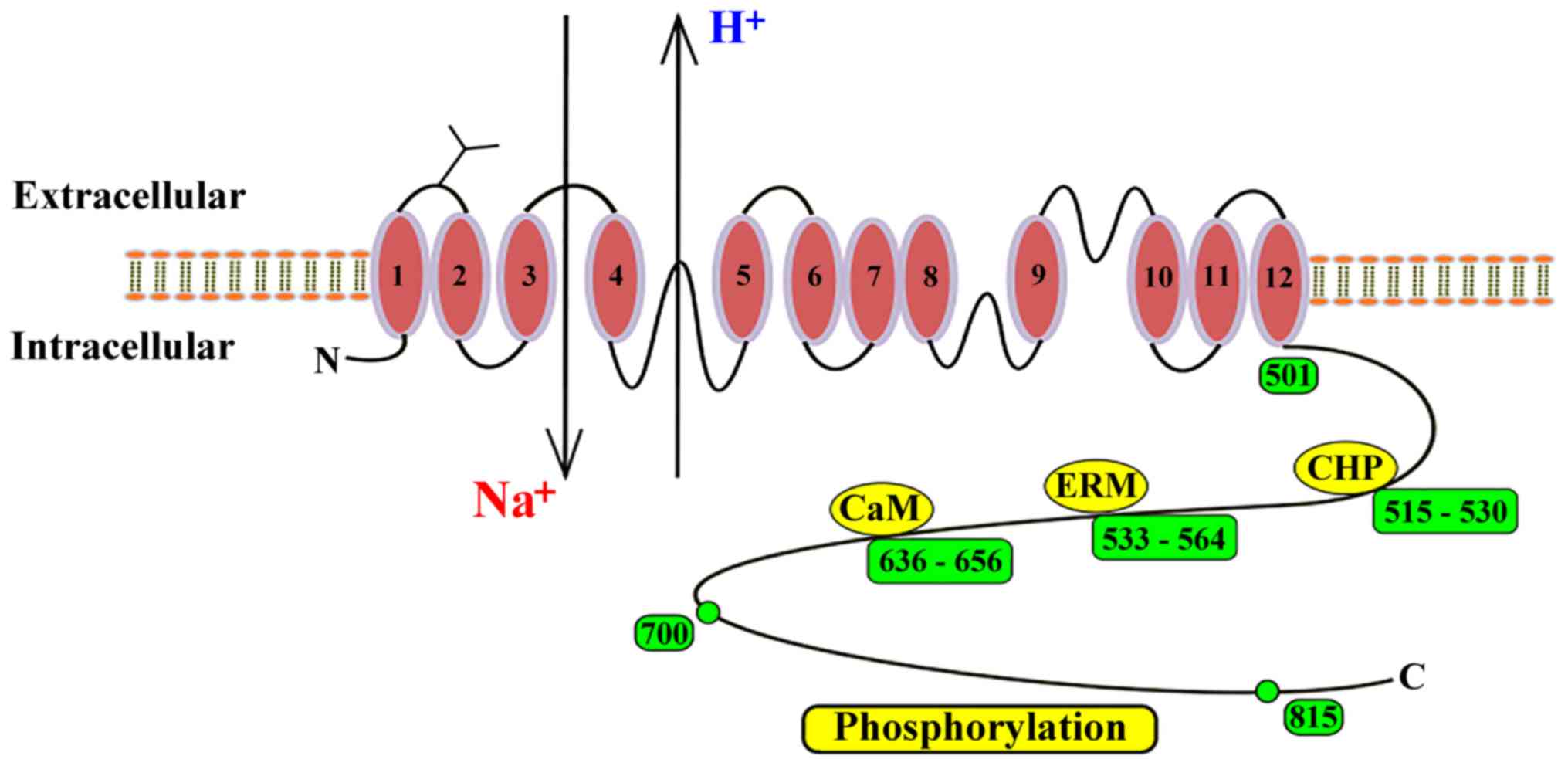
However, it is important to note that NHE membrane topology has
been most extensively studied in the NHE1 isoform (28). The complete membrane protein consists
of 815 amino acids, and the first 500 amino acids of the protein
are speculated to consist of 12 transmembrane hydrophobic domains
(29). A C-terminal hydrophilic
cytosolic domain of ~315 amino acids regulates the protein and
mediates cytoskeletal interactions (30,31).
Moreover, NHE2, 3, 4 and 5 have been reported to have 42, 39, 42
and 39% amino acid homology to NHE1, respectively (12). The NHE1 protein contains N- and
O-glycosylated residues (32) and
the N- and C- termini of NHE1 are found in the cytosol (6). Growth factors, hormones, integrins,
osmotic stress and other signaling pathways regulate the activity
of NHE1 via the mediation of the C-terminal domain, thus
determining the pHi (33). In
addition, there are some binding sites in the C-terminus, such as
calmodulin (CaM), CaM homologous protein (CHP) and
esrin/radixin/moesin (ERM) (4). When
CaM binds to NHE1, it eliminates self-inhibition and activates
NHE1. CHP AND ERM are bound to the cytosolic regulatory tail and
also support the physiological activity of NHE1(4) (Fig.
1).
NHEs are the most widely studied pHi regulators in
various animal cells, including hepatocytes (34). Intracellular acid load produced by
normal hepatocyte metabolism activates NHE proteins to catalyze the
electroneutral exchange of one extracellular Na+ with
one intracellular H+, thus constituting a key component
that prevents cell acidosis (4).
Furthermore, this exchange process depends on the inward-directed
Na+ gradient produced by the Na+/K+-ATPase to
excrete H+ from the cytoplasm (9,35). NHEs
are also involved in regulating the volume of hepatocytes (36), hepatocyte growth, regeneration,
proliferation, apoptosis and bile formation, and a series of
physiological activities, which are described below.
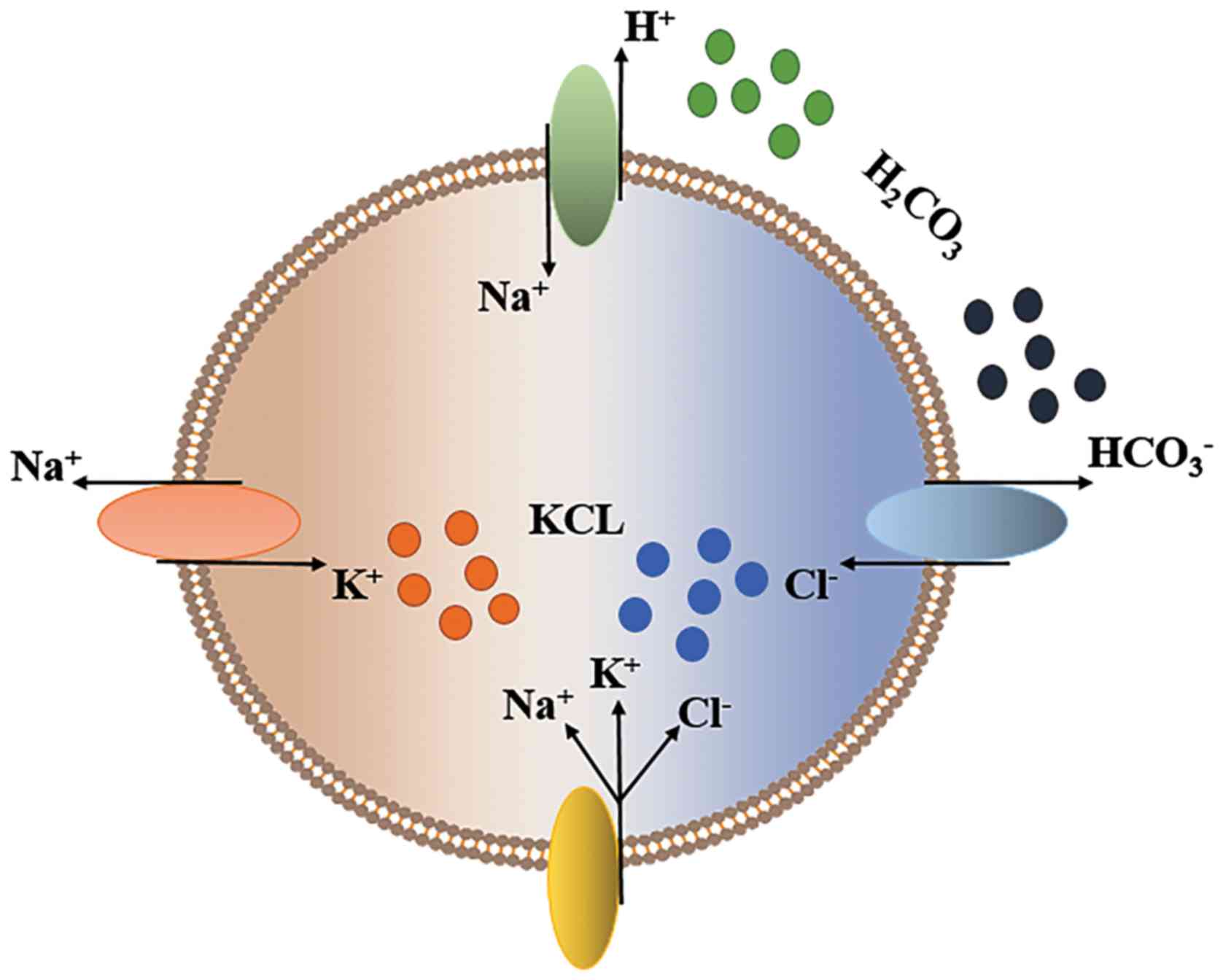
Regulation of cell volume is critical for liver
function in healthy and disease states (37). Shrinkage or swelling of cells may
result in disruption of the integrity of the cell membrane and
cytoskeletal structure. To survive, ions must pass via certain ion
transporters to avoid excessive changes in cell volume (38,39).
When cells are exposed to hypertonic extracellular media, cell
contraction triggers a regulated cell volume increase, which is
largely accomplished by cellular ion uptake (40,41).
Cellular contraction also stimulates
Na+/K+/2Cl- cotransporters (NKCC)
and/or NHEs in parallel with
Cl-/HCO3- exchangers (40). H+ excretion via NHEs and
HCO3- exiting via the
Cl-/HCO3- exchanger are
replenished in the cell by H2CO3, which is
readily produced by CO2; this process achieves NaCl entry (42). On the other hand, Na+,
which enters the cell via the NKCC and NHE, is pumped out by the
Na+/K+-ATPase in exchange for K+,
which eventually causes KCl uptake by the cells (42) (Fig.
2). Moreover, the NKCC isoforms NKCC1 and NKCC2(43) and the NHE isomers NHE1, 2 and 4 are
activated by cell contraction, while NHE3 is inhibited by cell
contraction (43).
The effects of cell proliferation may be mediated by
enhancing cell survival or inhibiting apoptosis (55). Intracellular acidification and cell
volume reduction are markers of apoptosis, while NHE1 intracellular
alkalization and the regulatory volume increases may be
antiapoptotic signals (2,56). TGF-β, which induces apoptosis of
hepatic parenchymal cells (57), has
a significant inhibitory effect on NHE activity in short-term
cultured rat hepatocytes, especially in cells isolated from
perivenular regions, in which apoptosis is more frequently observed
(58).
Hepatocytes and bile duct epithelial cells are
involved in bile secretion and absorption (59). Bile duct epithelial cells also serve
a role in the transport of water, electrolytes, sugar, bile and
amino acids, and express several transport proteins to modify the
primary production of hepatocyte bile (60). In biliary cells, four isoforms
(NHE1-4) have been identified (61,62).
Basolateral NHE1 is generally speculated to be involved in pHi,
cell volume homeostasis and fluid and electrolyte transport,
particularly secretin-induced bile secretion (63,64). In
addition, NHE3 has been detected in cholangiocarcinoma cells in
rats (65,66) and in gallbladder epithelial cells in
calves (67) and prairie dogs
(68). Targeted destruction of the
NHE3 gene results in inhibition of fluid reabsorption in isolated
bile duct units. For example, a study in mice found that decreased
gallbladder absorption of bile may be the result of a decrease in
NHE3 activity caused by an increased level of NHE3 phosphorylated
at serine-552; this increase in phosphorylation is hypothesized to
lead to a higher turnover of NHE3, which leads to a decrease in the
gallbladder's concentrating function (69). Moreover, prairie dogs represent a
good animal model for human gallstone formation, and their
gallbladder epithelial cells exhibit H+
gradient-dependent Na+ uptake via NHE1 (~6% of total intake), NHE2
(~66% of total intake) and NHE3 (~28% of total intake), indicating
a significant contribution of NHEs to epithelial Na+
absorption (68,70). Along with the findings showing
increased absorption of Na+ and liquid in the early
stage of gallstones (71), it has
been proposed that apical membrane NHEs may be involved in the
pathogenesis of gallstones (69). In
conclusion, the aforementioned results suggested that decreases in
NHE activity affects the absorption capacity of bile duct
cells.
NHE1 may also regulate cell differentiation, as the
absence or inhibition of NHE1 impairs the differentiation pathway
(72). Furthermore, NHE1 function is
important in cytoskeletal tissue and cell migration (22). The cytoplasmic tail of NHE1 acts as
an anchor for actin filaments via the binding of ezrin, Radixin and
moesin proteins, and the destruction of these interactions or
inhibition of NHE1 activity leads to the inhibition of cell
migration and the formation of external adhesions (2).
NAFLD is closely related to liposome imbalance, and
hepatic steatosis is considered to be the first stage in the
development of NAFLD (73). With the
development of fibrosis and inflammation, NAFLD can progress to
non-alcoholic steatohepatitis (NASH) and eventually lead to liver
fibrosis, cirrhosis and cancer (74,75). NHE
activity is associated with steatosis in NAFLD. Previous studies
(76) have compared the expression
of NHE1 in the livers of normal diet mice and high-fat diet mice,
and revealed that the expression of NHE1 in the livers of high-fat
diet mice was nearly tripled and the long-term ablation of NHE1
activity in mice weakened high lipid diet-induced liver lipid
accumulation, which suggests that NHE activity plays a role in the
development of NAFLD (76).
Farnesoid X receptor (FXR) is a nuclear hormone receptor. It has
been reported that activation of FXR attenuates the development of
hepatic steatosis (77,78), and FXR agonists reduce
NASH-associated fibrosis (79,80). The
prolipogenic liver X receptor (LXRa) is an important regulator of
lipid metabolism. LXR activation promotes hepatic steatosis
(81,82), and treatment with liver-specific LXR
inhibitors reduced the development of hepatic steatosis (83). Prasad et al (76) revealed that in the livers of
NHE1-null (KO) mice, increased expression of FXR and downregulation
of LXRa expression were consistent with the results of long-term
NHE1 deletion in reducing liver lipid accumulation induced by
high-fat diet. In addition, the key regulators of adipogenesis,
acetyl-CoA carboxylase α (Acc1) and Acc β (Acc2), are downregulated
in NHE1-KO liver (76). Moreover,
downregulation of Acc1 and Acc2 expression levels can reverse
hepatic steatosis (84). Based on
these findings, it was speculated that the loss of NHEs in the
liver may lead to increased expression of FXR, downregulation of
LXRa and downregulation of Acc1/Acc2 expression, which may reduce
or reverse liver steatosis during the occurrence of fatty liver
diseases such as NAFLD, which slows the development of NAFLD.
Hepatic fibrosis is a common response to chronic
liver injury of variable origins, such as viruses and metabolism
(85). The mechanisms of liver
fibrosis include activation of hepatic stellate cells (HSCs) and
extracellular matrix (ECM) protein deposition, including various
collagens and matrix glycoconjugates (86). During the development of fibrosis,
HSCs proliferate and the activation process is characterized by the
appearance of myofibroblast-like phenotypes that accumulate near
necrotic areas (87). Activated HSCs
are characterized by the expression of α-smooth muscle actin (SMA)
(87), increased cell numbers, loss
of retinoic acid (88,89) and increased expression of collagen
fibrin (90). Currently, oxidative
stress (91,92), paracrine stimulation of damaged
hepatocytes, cytokines (93,94) and mitogens, such as platelet-derived
growth factor (PDGF), TGF-β and insulin-like growth factor I, have
been reported to promote the proliferation of HSCs and matrix
synthesis. All of these factors activate NHEs, mostly likely the
subtype 1, in the liver, and NHE activation is one of the earliest
responses to mitogens and growth factors in most cell types
(95). Previous studies have shown
that NHE1 is the main pH regulator in HSCs, and its activity
increases with the activation process of HSCs (96,97).
When different growth factors and oxidative stress stimulate HSC
proliferation and collagen type synthesis, the activity of NHE1
protein increases (96,98-100).
Furthermore, the mechanism of HSCs proliferation is related to the
increase in cell volume caused by NHE activation. For example, an
increase in cell volume itself induces multiple changes in cellular
function and gene expression by activating the osmosignaling
pathway, and is a prerequisite for cell division and proliferation
(38,101). It has also been reported that an
increase in cell volume is parallel to the process via which
fibroblasts transition from G1 to S phase (102).
Hepatocellular carcinoma (HCC) is one of the most
common malignant tumors in the world and has a poor prognosis
(106). The occurrence and
development of cancer are closely related to dysregulation of cell
energy metabolism, which is known as the Warburg effect (107). As a supportive anticancer therapy,
glucose restriction (GR) inhibits enhanced glycolytic activity in
cancer cells via energy-dependent signaling pathways, including the
insulin like growth factor-1/PI3K/Akt/mTOR pathway (108). Previous studies have reported that
intracellular alkalization is a major transformation event for
cancer cells. For instance, one study showed that glioblastoma (MG)
is a highly glycolytic malignant tumor that has a strong dependence
on pH, and NHE1 activation drives cytoplasmic alkalization
(109). However, inhibition of NHE1
in MG can acidify tumor cells, while healthy astrocytes are not
affected; this finding may facilitate the development of treatment
for malignant tumors (109).
Targeting proton dynamics associated with pHi gradients has been
proposed as a potential cancer prevention strategy and treatment
(110). Kim et al demonstrated that
curcumin treatment or GR slightly inhibited NHE1, while the
combined treatment of curcumin and GR further enhanced the
inhibitory effect on NHE1 and reduced pHi (110). Since the activation of NHE1 depends
on energy and Akt (111), GR
enhances the ability of curcumin to synergistically inhibit NHE1.
Therefore, the combined treatment of GR and curcumin may have an
important role in the regulation of pH in human hepatoma cells.
The expression of NHE1 in tumor tissues is not only
related to the tumor size in HCC, but also venous invasion and
pathological TNM staging (112).
Previous studies have reported that inhibition of NHE1 blocks the
invasion and metastasis of SMMC-7721 and HepG2 liver cancer cells
(113,114). Invasion and migration of malignant
tumor cells require destruction of the basement membrane and ECM
proteolysis (115). The acidic
extracellular pH (pHe) of tumor cells is also crucial in the
activation of extracellular proteinases and the degradation of ECM
in tumors (116,117). In the early stages of cell
migration, invasion and metastasis, ECM decomposition is mainly
mediated by matrix metalloproteinases (MMP)-2, -3 and -9 (114,118).
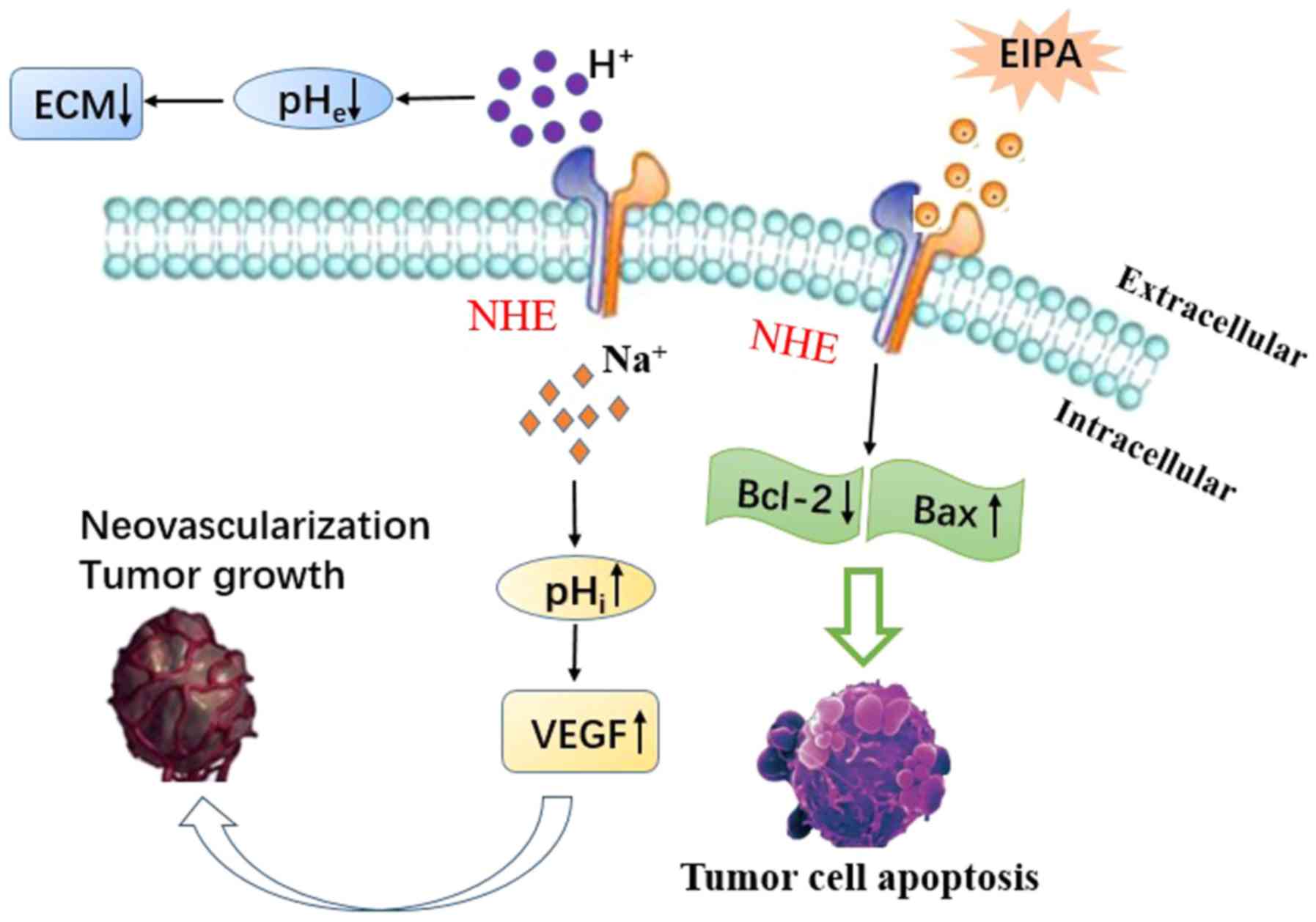
Moreover, alkaline pHi promotes the expression of vascular
endothelial growth factor (VEGF), which plays an important role in
inducing neovascularization and promoting tumor growth and
metastatic potential (Fig. 3)
(119-121).
Hypoxia is also a common feature of the tumor microenvironment and
has been shown to stimulate invasion and metastasis (122,123).
Hypoxia activates ERK1/2, a family of mitogen-activated protein
kinases that play a major role in signaling pathways that are
involved in cell scatter, motion, invasion, proliferation and
survival (124). ERK1/2 also
regulates the expression of MMPs and VEGF (125,126).
Yang et al (114)
demonstrated that inhibition of NHE1 by ethyl-isopropyl-amiloride
(EIPA) inhibited HepG2 cell invasion and metastasis, and that EIPA
inhibition acts by downregulating MMP-2, MMP-9 and VEGF in an
ERK1/2-dependent manner. Moreover, NHE1 not only affects the
migration and invasion of tumor cells, but is also related to the
apoptosis of tumor cells (127).
Bcl-2 family members, such as Bcl-2, Bcl-xL and Bax, play a crucial
role in controlling apoptosis (127). Previous studies have reported that
the NHE1 inhibitor EIPA downregulates Bcl-2, and upregulates Bax
expression in HepG2 cells, leading to tumor cell apoptosis
(128) (Fig. 3).
Interleukin 6 (IL6) is a key cytokine involved in
the development and progression of inflammation-associated HCC. For
example. Xu et al (128)
found that IL6 activates the functional activity of NHE1, induces
the interaction of NHE1, Na+/Ca2+ exchanger1
(NCX1) and calmodulin (CaM), and upregulates the expression of NHE1
in human hepatoma cells. Benzo[a]pyrene (B[a]P) is a prototype of
polycyclic aromatic hydrocarbons, and is a human carcinogen
(107). In addition to triggering
apoptotic signals, B[a]P may induce survival signals and
participate in the promotion of cancer (107). Previous studies have also reported
that B[a]P induces metabolic reprogramming, which involves the
activation of NHE1, and it leads to epithelial-mesenchymal
transition (129-131).
Ginsenoside Rg3, the main pharmacologically active compound
extracted from Chinese ginseng, has been widely recognized as
having antitumor properties in various cancer types (132,133),
including inhibition of HCC cell proliferation, induction of
apoptosis and inhibition of angiogenesis (134) and metastasis (135). Recently, it has been reported that
Rg3 inhibits HCC cell proliferation and induces apoptosis by
decreasing NHE1 expression and activity, and Rg3-mediated NHE1
inhibition is dependent on the EGF/EGFR/ERK1/2/hypoxia-inducible
factor-1α signaling pathway (136).
As a common ion transporter, NHEs are regulated by numerous
substances. For example, IL6 and B[a]P can activate the activity of
NHE1, while Rg3 can inhibit the activity of NHEs (107,128,132).
Therefore, based on the aforementioned description of IL6, B[a]P
and Rg3 in HCC, it was suggested that NHEs may be a potential
therapeutic target in HCC. However, further studies are required to
identify the potential mechanisms.
Cholangiocellular carcinoma (CCC) is the second most
common primary liver cancer after HCC, with an increased incidence
(137). Furthermore, chronic
inflammation and oxidative stress play a key role in the
development of CCC (138). NHEs
form a potential link between controlling pHi and tumor
development. Therefore, Elsing et al (62) determined the effect of oxidative
stress on NHEs using tert-Butyl hydroperoxide (t-BOOH), a hydrogen
peroxide, in the biliary epithelial cancer cell line Mz-Cha-1.
These authors demonstrated that t-BOOH reduced NHE activity in a
dose-dependent manner; at 4 mM t-BOOH, the NHE activity was almost
absent and glutathione supplementation and intracellular
Ca2+ chelation partially restored NHE activity.
Moreover, in Mz-Cha-1 cells, inhibition of NHE by oxidative stress
depends in part on the presence of intracellular Ca2+
and intracellular glutathione levels (62).
Hepatic failure (HF) is a life-threatening disease
with a very high mortality rate (139), and hepatocyte apoptosis leading to
HF is an important event in hepatocyte death. Tumor necrosis
factor-α (TNF-α) is an inflammatory factor and an inducer of
hepatocyte apoptosis (140). It has
been reported that TNF-α induces NHE activity in hepatocytes in a
time-dependent manner (141).
Activation of NHEs increases intracellular Na+, promotes
Na+/Ca2+ exchange and causes Ca2+
overload (142), which is
considered to be the key factor in cell damage. Moreover, an
increase in intracellular Ca2+ concentration
automatically activates calpain, a calcium-dependent protease
(143). The antiapoptotic family
member Bcl-xL is a natural substrate for calpain, and NHE mediates
TNF-α-induced hepatocyte apoptosis via
Ca2+/calpain-dependent degradation of Bcl-xL (141). However, the NHE inhibitor
cariporide reverses the effects induced by TNF-α and has a
protective effect on acute HF (141).
Endotoxin-mediated production of proinflammatory
cytokines plays an important role in the pathogenesis of liver
disease (144). Previous studies
have reported that lipopolysaccharide (LPS) causes liver damage by
increasing the release of TNF-α in a NASH model (145). Therefore, interfering with
LPS-induced inflammatory responses may help to alleviate
inflammation associated with liver disease. Heat shock proteins
(Hsp70) play an important role in LPS-mediated inflammatory
responses (144). NHE1 is
considered to be a mediator of inflammatory responses in
macrophages (146-148)
and has been reported to interact with Hsp70(149). Inhibition of Hsp70 substrate
binding activity in vivo reduces induction of proinflammatory
factors (144). Huang et al
(144) treated macrophages and
livers with LPS and revealed a significant increase in the
association of NHE1-Hsp70, suggesting that the formation of the
NHE1-Hsp70 complex is essential for the induction of
proinflammatory factors. Therefore, LPS-induced liver damage may be
prevented by disrupting the NHE1-Hsp70 interaction.
In addition, the beneficial effects of the NHE
inhibitor EIPA in blocking NHEs in a partial hepatic ischemia rat
model suggested a positive role for NHE1 in oxidative liver injury,
and indicated that inhibition of NHEs is a potential strategy for
preventing or reducing ischemic liver injury (27).
NHEs are ion transporters that are widely present in
a variety of organisms and are important regulators at the
cellular, tissue and systemic levels (150). To the best of our knowledge, the
current review is the first detailed description of the physiology
and pathology of NHEs in the liver. While NHEs are common targets
for various inflammatory stimuli, the effect of selective targeted
therapy of NHEs in the liver are inconclusive, and thus further
studies are required. Numerous experimental models currently show
that NHE inhibitors lack major toxic effects, and several, such as
cariporide, have been used in preclinical and clinical trials
(151). However, a large number of
studies have only analyzed the effects of single factors, and have
not considered that their function in physiological and
pathological conditions may mainly be the result of the interaction
of various transporters. Therefore, more comprehensive methods are
required to alter the function, and regulate and target NHEs in
liver-related pathology.
The authors would like to thank Professor Jiaxing
An (Department of Gastroenterology, Affiliated Hospital to Zunyi
Medical University) for their highly professional services.
This study was supported by grant from the Clinical
Medical Research Center for Digestive Diseases,Guizhou
province,China
Not applicable.
TL wrote the manuscript and participated in
information collection, analysis, organization and figure design.
BT primarily revised and final editing. All authors read and
approved the final manuscript.
Not applicable.
Not applicable.
The authors declare that they have no competing
interests.
|
1
|
Kraut JA and Madias NE: Treatment of acute
metabolic acidosis: A pathophysiologic approach. Nat Rev Nephrol.
8:589–601. 2012.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Putney LK, Denker SP and Barber DL: The
changing face of the Na+/H+ exchanger, NHE1:
Structure, regulation, and cellular actions. Annu Rev Pharmacol
Toxicol. 42:527–552. 2002.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Kondapalli KC, Prasad H and Rao R: An
inside job: How endosomal Na(+)/H(+) exchangers link to autism and
neurological disease. Front Cell Neurosci. 8(172)2014.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Malo ME and Fliegel L: Physiological role
and regulation of the Na+/H+ exchanger. Can J
Physiol Pharmacol. 84:1081–1095. 2006.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Gurney MA, Laubitz D, Ghishan FK and Kiela
PR: Pathophysiology of Intestinal Na+/H+
exchange. Cell Mol Gastroenterol Hepatol. 3:27–40. 2017.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Sardet C, Franchi A and Pouysségur J:
Molecular cloning, primary structure, and expression of the human
growth factor-activatable Na+/H+ antiporter.
Cell. 56:271–280. 1989.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Xu H, Chen H, Li J, Zhao Y and Ghishan FK:
Disruption of NHE8 expression impairs Leydig cell function in the
testes. Am J Physiol Cell Physiol. 308:C330–C338. 2015.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Denker SP and Barber DL: Cell migration
requires both ion translocation and cytoskeletal anchoring by the
Na-H exchanger NHE1. J Cell Biol. 159:1087–1096. 2002.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Orlowski J and Grinstein S: Diversity of
the mammalian sodium/proton exchanger SLC9 gene family. Pflugers
Arch. 447:549–565. 2004.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Zhao H, Carney KE, Falgoust L, Pan JW, Sun
D and Zhang Z: Emerging roles of Na+/H+
exchangers in epilepsy and developmental brain disorders. Prog
Neurobiol. 138-140:19–35. 2016.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Malakooti J, Dahdal RY, Schmidt L, Layden
TJ, Dudeja PK and Ramaswamy K: Molecular cloning, tissue
distribution, and functional expression of the human Na(+)/H(+)
exchanger NHE2. Am J Physiol. 277:G383–G390. 1999.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Loo SY, Chang MK, Chua CS, Kumar AP,
Pervaiz S and Clement MV: NHE-1: A promising target for novel
anti-cancer therapeutics. Curr Pharm Des. 18:1372–1382.
2012.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Kurata T, Rajendran V, Fan S, Ohta T,
Numata M and Fushida S: NHE5 regulates growth factor signaling,
integrin trafficking, and degradation in glioma cells. Clin Exp
Metastasis. 36:527–538. 2019.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Pescosolido MF, Stein DM, Schmidt M, El
Achkar CM, Sabbagh M, Rogg JM, Tantravahi U, McLean RL, Liu JS,
Poduri A, et al: Genetic and phenotypic diversity of NHE6 mutations
in Christianson syndrome. Ann Neurol. 76:581–593. 2014.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Nakamura N, Tanaka S, Teko Y, Mitsui K and
Kanazawa H: Four Na+/H+ exchanger isoforms
are distributed to Golgi and post-Golgi compartments and are
involved in organelle pH regulation. J Biol Chem. 280:1561–1572.
2005.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Ko M, Quiñones-Hinojosa A and Rao R:
Emerging links between endosomal pH and cancer. Cancer Metastasis
Rev: Apr 6, 2020 (Epub ahead of print).
|
|
17
|
Laczkó D, Rosztóczy A, Birkás K, Katona M,
Rakonczay Z Jr, Tiszlavicz L, Róka R, Wittmann T, Hegyi P and
Venglovecz V: Role of ion transporters in the bile acid-induced
esophageal injury. Am J Physiol Gastrointest Liver Physiol.
311:G16–G31. 2016.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Hosogi S, Miyazaki H, Nakajima K, Ashihara
E, Niisato N, Kusuzaki K and Marunaka Y: An inhibitor of Na(+)/H(+)
exchanger (NHE), ethyl isopropyl amiloride (EIPA), diminishes
proliferation of MKN28 human gastric cancer cells by decreasing the
cytosolic Cl(-) concentration via DIDS sensitive pathways. Cell
Physiol Biochem. 30:1241–1253. 2012.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Khan I and Khan K: Uncoupling of Carbonic
Anhydrase from Na H exchanger 1 in Experimental Colitis: A Possible
Mechanistic Link with Na H Exchanger. Biomolecules.
9(700)2019.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Xu H, Li J, Chen H and Ghishan FK: NHE8
Deficiency Promotes Colitis-Associated Cancer in Mice via Expansion
of Lgr5-Expressing Cells. Cell Mol Gastroenterol Hepatol. 7:19–31.
2018.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Magalhães D, Cabral JM, Soares-da-Silva P
and Magro F: Role of epithelial ion transports in inflammatory
bowel disease. Am J Physiol Gastrointest Liver Physiol.
310:G460–G476. 2016.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Das S, Jayaratne R and Barrett KE: The
role of ion transporters in the pathophysiology of infectious
diarrhea. Cell Mol Gastroenterol Hepatol. 6:33–45. 2018.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Cao L, Yuan Z, Liu M and Stock C:
(Patho-)Physiology of Na+/H+ Exchangers
(NHEs) in the Digestive System. Front Physiol.
10(1566)2020.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Yong W: Image diagnosis of common liver
lesions Image research and medical application(on the column). J
Imaging Res Med Applic, 2018 (In Chinese).
|
|
25
|
Lowry SF and Brennan MF: Abnormal liver
function during parenteral nutrition: Relation to infusion excess.
J Surg Res. 26:300–307. 1979.
|
|
26
|
Laohapitakworn S, Thongbunchoo J,
Nakkrasae LI, Krishnamra N and Charoenphandhu N: Parathyroid
hormone (PTH) rapidly enhances CFTR-mediated
HCO3- secretion in intestinal epithelium-like
Caco-2 monolayer: A novel ion regulatory action of PTH. Am J
Physiol Cell Physiol. 301:C137–C149. 2011.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Xu H, Ghishan FK and Kiela PR: SLC9 Gene
Family: Function, expression, and regulation. Compr Physiol.
8:555–583. 2018.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Kemp G, Young H and Fliegel L: Structure
and function of the human Na+/H+ exchanger
isoform 1. Channels (Austin). 2:329–336. 2008.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Fuster DG and Alexander RT: Traditional
and emerging roles for the SLC9 Na+/H+
exchangers. Pflugers Arch. 466:61–76. 2014.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Stock C and Schwab A: Role of the Na/H
exchanger NHE1 in cell migration. Acta Physiol (Oxf). 187:149–157.
2006.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Fliegel L: Structural and Functional
Changes in the Na(+)/H(+) Exchanger Isoform 1, Induced by Erk1/2
Phosphorylation. Int J Mol Sci. 20(2378)2019.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Fuster DG and Alexander RT: Traditional
and emerging roles for the SLC9 Na+/H+
exchangers. Pflugers Arch. 466:61–76. 2014.PubMed/NCBI View Article : Google Scholar
|
|
33
|
Amith SR and Fliegel L: Regulation of the
Na+/H+ Exchanger (NHE1) in Breast Cancer
Metastasis. Cancer Res. 73:1259–1264. 2013.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Lee CH, Cragoe EJ Jr and Edwards AM:
Control of hepatocyte DNA synthesis by intracellular pH and its
role in the action of tumor promoters. J Cell Physiol. 195:61–69.
2003.PubMed/NCBI View Article : Google Scholar
|
|
35
|
Aharonovitz O, Zaun HC, Balla T, York JD,
Orlowski J and Grinstein S: Intracellular pH regulation by
Na(+)/H(+) exchange requires phosphatidylinositol 4,5-bisphosphate.
J Cell Biol. 150:213–224. 2000.PubMed/NCBI View Article : Google Scholar
|
|
36
|
Ahmed KH, Pelster B and Krumschnabel G:
Signalling pathways involved in hypertonicity- and
acidification-induced activation of Na+/H+
exchange in trout hepatocytes. J Exp Biol. 209:3101–3113.
2006.PubMed/NCBI View Article : Google Scholar
|
|
37
|
Haussinger D: Osmosensing and
osmosignaling in the liver. Wiener medizinische Wochenschrift
(1946). 158:549–552. 2008.PubMed/NCBI View Article : Google Scholar
|
|
38
|
Lang F, Busch GL, Ritter M, Völkl H,
Waldegger S, Gulbins E and Häussinger D: Functional significance of
cell volume regulatory mechanisms. Physiol Rev. 78:247–306.
1998.PubMed/NCBI View Article : Google Scholar
|
|
39
|
Jo AO, Ryskamp DA, Phuong TT, Verkman AS,
Yarishkin O, MacAulay N and Križaj D: TRPV4 and AQP4 channels
synergistically regulate cell volume and calcium homeostasis in
retinal Müller Glia. J Neurosci. 35:13525–13537. 2015.PubMed/NCBI View Article : Google Scholar
|
|
40
|
Lang F, Föller M, Lang K, Lang P, Ritter
M, Vereninov A, Szabo I, Huber SM and Gulbins E: Cell volume
regulatory ion channels in cell proliferation and cell death.
Methods Enzymol. 428:209–225. 2007.PubMed/NCBI View Article : Google Scholar
|
|
41
|
Mongin AA: Volume-regulated anion channel
- a frenemy within the brain. Pflugers Arch. 468:421–441.
2016.PubMed/NCBI View Article : Google Scholar
|
|
42
|
Lang PA, Graf D, Boini KM, Lang KS,
Klingel K, Kandolf R and Lang F: Cell volume, the serum and
glucocorticoid inducible kinase 1 and the liver. Z Gastroenterol.
49:713–719. 2011.PubMed/NCBI View Article : Google Scholar
|
|
43
|
Hoffmann EK, Lambert IH and Pedersen SF:
Physiology of cell volume regulation in vertebrates. Physiol Rev.
89:193–277. 2009.PubMed/NCBI View Article : Google Scholar
|
|
44
|
Häussinger D and Lang F: Cell volume and
hormone action. Trends Pharmacol Sci. 13:371–373. 1992.PubMed/NCBI View Article : Google Scholar
|
|
45
|
Lee MJ: Hormonal regulation of
adipogenesis. Compr Physiol. 7:1151–1195. 2017.PubMed/NCBI View Article : Google Scholar
|
|
46
|
O'Connor McCourt M, Soley M, Hayden LJ and
Hollenberg MD: Receptors for epidermal growth factor (urogastrone)
and insulin in primary cultures of rat hepatocytes maintained in
serum free medium. Biochem Cell Biol. 64:803–810. 1986.PubMed/NCBI View Article : Google Scholar
|
|
47
|
Dykes SS, Steffan JJ and Cardelli JA:
Lysosome trafficking is necessary for EGF-driven invasion and is
regulated by p38 MAPK and Na+/H+ exchangers.
BMC Cancer. 17(672)2017.PubMed/NCBI View Article : Google Scholar
|
|
48
|
Steffan JJ, Williams BC, Welbourne T and
Cardelli JA: HGF-induced invasion by prostate tumor cells requires
anterograde lysosome trafficking and activity of
Na+-H+ exchangers. J Cell Sci. 123:1151–1159.
2010.PubMed/NCBI View Article : Google Scholar
|
|
49
|
Coaxum SD, Blanton MG, Joyner A, Akter T,
Bell PD, Luttrell LM, Raymond JR Sr, Lee MH, Blichmann PA,
Garnovskaya MN, et al: Epidermal growth factor-induced
proliferation of collecting duct cells from Oak Ridge polycystic
kidney mice involves activation of Na+/H+
exchanger. Am J Physiol Cell Physiol. 307:C554–C560.
2014.PubMed/NCBI View Article : Google Scholar
|
|
50
|
Mead JE and Fausto N: Transforming growth
factor alpha may be a physiological regulator of liver regeneration
by means of an autocrine mechanism. Proc Natl Acad Sci USA.
86:1558–1562. 1989.PubMed/NCBI View Article : Google Scholar
|
|
51
|
Kaneko A, Hayashi N, Tanaka Y, Horimoto M,
Ito T, Sasaki Y, Fusamoto H and Kamada T: Activation of
Na+/H+ exchanger by hepatocyte growth factor
in hepatocytes. Hepatology. 22:629–636. 1995.PubMed/NCBI
|
|
52
|
Goodrich AL and Suchy FJ: Na(+)-H+
exchange in basolateral plasma membrane vesicles from neonatal rat
liver. Am J Physiol. 259:G334–G339. 1990.PubMed/NCBI View Article : Google Scholar
|
|
53
|
Dällenbach A, Marti U and Renner EL:
Hepatocellular Na+/H+ exchange is activated
early, transiently and at a posttranscriptional level during rat
liver regeneration. Hepatology. 19:1290–1301. 1994.PubMed/NCBI View Article : Google Scholar
|
|
54
|
Moule SK and McGivan JD: Epidermal growth
factor and cyclic AMP stimulate Na+/H+
exchange in isolated rat hepatocytes. Eur J Biochem. 187:677–682.
1990.PubMed/NCBI View Article : Google Scholar
|
|
55
|
Love MR, Palee S, Chattipakorn SC and
Chattipakorn N: Effects of electrical stimulation on cell
proliferation and apoptosis. J Cell Physiol. 233:1860–1876.
2018.PubMed/NCBI View Article : Google Scholar
|
|
56
|
Cardoso VG, Gonçalves GL, Costa-Pessoa JM,
Thieme K, Lins BB, Casare FAM, de Ponte MC, Camara NOS and
Oliveira-Souza M: Angiotensin II-induced podocyte apoptosis is
mediated by endoplasmic reticulum stress/PKC-δ/p38 MAPK pathway
activation and trough increased Na+/H+
exchanger isoform 1 activity. BMC Nephrol. 19(179)2018.PubMed/NCBI View Article : Google Scholar
|
|
57
|
Gaitantzi H, Meyer C, Rakoczy P, Thomas M,
Wahl K, Wandrer F, Bantel H, Alborzinia H, Wölfl S, Ehnert S, et
al: Ethanol sensitizes hepatocytes for TGF-β-triggered apoptosis.
Cell Death Dis. 9(51)2018.PubMed/NCBI View Article : Google Scholar
|
|
58
|
Benedetti A, Di Sario A, Svegliati Baroni
G and Jezequel AM: Transforming growth factor beta 1 increases the
number of apoptotic bodies and decreases intracellular pH in
isolated periportal and perivenular rat hepatocytes. Hepatology.
22:1488–1498. 1995.PubMed/NCBI
|
|
59
|
Martínez-Ansó E, Castillo JE, Díez J,
Medina JF and Prieto J: Immunohistochemical detection of
chloride/bicarbonate anion exchangers in human liver. Hepatology.
19:1400–1406. 1994.PubMed/NCBI
|
|
60
|
Marin JJ, Macias RI, Briz O, Banales JM
and Monte MJ: Bile Acids in Physiology, Pathology and Pharmacology.
Curr Drug Metab. 17:4–29. 2015.PubMed/NCBI View Article : Google Scholar
|
|
61
|
Marti U, Elsing C, Renner EL,
Liechti-Gallati S and Reichen J: Differential expression of Na+
H(+)-antiporter mRNA in biliary epithelial cells and in
hepatocytes. J Hepatol. 24:498–502. 1996.PubMed/NCBI View Article : Google Scholar
|
|
62
|
Elsing C, Voss A, Herrmann T, Kaiser I,
Huebner CA and Schlenker T: Oxidative stress reduces
Na+/H+ exchange (NHE) activity in a biliary
epithelial cancer cell line (Mz-Cha-1). Anticancer Res. 31:459–465.
2011.PubMed/NCBI
|
|
63
|
Hirata K and Nathanson MH: Bile duct
epithelia regulate biliary bicarbonate excretion in normal rat
liver. Gastroenterology. 121:396–406. 2001.PubMed/NCBI View Article : Google Scholar
|
|
64
|
Hübner C, Stremmel W and Elsing C: Sodium,
hydrogen exchange type 1 and bile ductular secretory activity in
the guinea pig. Hepatology. 31:562–571. 2000.PubMed/NCBI View Article : Google Scholar
|
|
65
|
Roussa E, Bertram J, Berge KE, Labori KJ,
Thévenod F and Raeder MG: Differential regulation of vacuolar
H+ -ATPase and Na+/H+ exchanger 3
in rat cholangiocytes after bile duct ligation. Histochem Cell
Biol. 125:419–428. 2006.PubMed/NCBI View Article : Google Scholar
|
|
66
|
Mennone A, Biemesderfer D, Negoianu D,
Yang CL, Abbiati T, Schultheis PJ, Shull GE, Aronson PS and Boyer
JL: Role of sodium/hydrogen exchanger isoform NHE3 in fluid
secretion and absorption in mouse and rat cholangiocytes. Am J
Physiol Gastrointest Liver Physiol. 280:G247–G254. 2001.PubMed/NCBI View Article : Google Scholar
|
|
67
|
Bazzini C, Bottà G, Meyer G, Baroni MD and
Paulmichl M: The presence of NHE1 and NHE3
Na+-H+ exchangers and an apical
cAMP-independent Cl- channel indicate that both
absorptive and secretory functions are present in calf gall bladder
epithelium. Exp Physiol. 86:571–583. 2001.PubMed/NCBI View Article : Google Scholar
|
|
68
|
Narins SC, Park EH, Ramakrishnan R, Garcia
FU, Diven JN, Balin BJ, Hammond CJ, Sodam BR, Smith PR and Abedin
MZ: Functional characterization of Na(+)/H(+) exchangers in primary
cultures of prairie dog gallbladder. J Membr Biol. 197:123–134.
2004.PubMed/NCBI View Article : Google Scholar
|
|
69
|
Chen Y, Wu S, Tian Y and Kong J:
Phosphorylation and subcellular localization of
Na+/H+ exchanger isoform 3 (NHE3) are
associated with altered gallbladder absorptive function after
formation of cholesterol gallstones. J Physiol Biochem. 73:133–139.
2017.
|
|
70
|
Saier MH Jr, Yen MR, Noto K, Tamang DG and
Elkan C: The Transporter Classification Database: Recent advances.
Nucleic Acids Res. 37 (Database):D274–D278. 2009.PubMed/NCBI View Article : Google Scholar
|
|
71
|
Giurgiu DI, Saunders-Kirkwood KD, Roslyn
JJ and Abedin MZ: Sequential changes in biliary lipids and
gallbladder ion transport during gallstone formation. Ann Surg.
225:382–390. 1997.PubMed/NCBI View Article : Google Scholar
|
|
72
|
Li X, Karki P, Lei L, Wang H and Fliegel
L: Na+/H+ exchanger isoform 1 facilitates
cardiomyocyte embryonic stem cell differentiation. Am J Physiol
Heart Circ Physiol. 296:H159–H170. 2009.PubMed/NCBI View Article : Google Scholar
|
|
73
|
Manne V, Handa P and Kowdley KV:
Pathophysiology of Nonalcoholic Fatty Liver Disease/Nonalcoholic
Steatohepatitis. Clin Liver Dis. 22:23–37. 2018.PubMed/NCBI View Article : Google Scholar
|
|
74
|
Baffy G, Brunt EM and Caldwell SH:
Hepatocellular carcinoma in non-alcoholic fatty liver disease: An
emerging menace. J Hepatol. 56:1384–1391. 2012.PubMed/NCBI View Article : Google Scholar
|
|
75
|
Friedman SL, Neuschwander-Tetri BA,
Rinella M and Sanyal AJ: Mechanisms of NAFLD development and
therapeutic strategies. Nat Med. 24:908–922. 2018.PubMed/NCBI View Article : Google Scholar
|
|
76
|
Prasad V, Chirra S, Kohli R and Shull GE:
NHE1 deficiency in liver: Implications for non-alcoholic fatty
liver disease. Biochem Biophys Res Commun. 450:1027–1031.
2014.PubMed/NCBI View Article : Google Scholar
|
|
77
|
Cipriani S, Mencarelli A, Palladino G and
Fiorucci S: FXR activation reverses insulin resistance and lipid
abnormalities and protects against liver steatosis in Zucker
(fa/fa) obese rats. J Lipid Res. 51:771–784. 2010.PubMed/NCBI View Article : Google Scholar
|
|
78
|
Ma Y, Huang Y, Yan L, Gao M and Liu D:
Synthetic FXR agonist GW4064 prevents diet-induced hepatic
steatosis and insulin resistance. Pharm Res. 30:1447–1457.
2013.PubMed/NCBI View Article : Google Scholar
|
|
79
|
Mudaliar S, Henry RR, Sanyal AJ, Morrow L,
Marschall HU, Kipnes M, Adorini L, Sciacca CI, Clopton P, Castelloe
E, et al: Efficacy and safety of the farnesoid X receptor agonist
obeticholic acid in patients with type 2 diabetes and nonalcoholic
fatty liver disease. Gastroenterology. 145:574–82.e1.
2013.PubMed/NCBI View Article : Google Scholar
|
|
80
|
Tully DC, Rucker PV, Chianelli D, Williams
J, Vidal A, Alper PB, Mutnick D, Bursulaya B, Schmeits J, Wu X, et
al: Discovery of tropifexor (LJN452), a highly potent non-bile acid
FXR agonist for the treatment of cholestatic liver diseases and
nonalcoholic steatohepatitis (NASH). J Med Chem. 60:9960–9973.
2017.PubMed/NCBI View Article : Google Scholar
|
|
81
|
Baranowski M, Zabielski P, Blachnio
Zabielska AU, Harasim E, Chabowski A and Gorski J: Insulin
sensitizing effect of LXR agonist T0901317 in high fat fed rats is
associated with restored muscle GLUT4 expression and insulin
stimulated AS160 phosphorylation. Cell Physiol Biochem.
33:1047–1057. 2014.PubMed/NCBI View Article : Google Scholar
|
|
82
|
Ducheix S, Montagner A, Theodorou V,
Ferrier L and Guillou H: The liver X receptor: A master regulator
of the gut-liver axis and a target for non alcoholic fatty liver
disease. Biochem Pharmacol. 86:96–105. 2013.PubMed/NCBI View Article : Google Scholar
|
|
83
|
Griffett K, Solt LA, El-Gendy BD,
Kamenecka TM and Burris TP: A liver-selective LXR inverse agonist
that suppresses hepatic steatosis. ACS Chem Biol. 8:559–567.
2013.PubMed/NCBI View Article : Google Scholar
|
|
84
|
Savage DB, Choi CS, Samuel VT, Liu ZX,
Zhang D, Wang A, Zhang XM, Cline GW, Yu XX, Geisler JG, et al:
Reversal of diet-induced hepatic steatosis and hepatic insulin
resistance by antisense oligonucleotide inhibitors of acetyl-CoA
carboxylases 1 and 2. J Clin Invest. 116:817–824. 2006.PubMed/NCBI View Article : Google Scholar
|
|
85
|
Dewidar B, Meyer C, Dooley S and Meindl
Beinker AN: TGF beta in hepatic stellate cell activation and liver
fibrogenesis-Updated 2019. Cells. 8(1419)2019.PubMed/NCBI View Article : Google Scholar
|
|
86
|
Yang C, Zeisberg M, Mosterman B, Sudhakar
A, Yerramalla U, Holthaus K, Xu L, Eng F, Afdhal N and Kalluri R:
Liver fibrosis: Insights into migration of hepatic stellate cells
in response to extracellular matrix and growth factors.
Gastroenterology. 124:147–159. 2003.PubMed/NCBI View Article : Google Scholar
|
|
87
|
Li J, Zhao YR and Tian Z: Roles of hepatic
stellate cells in acute liver failure: From the perspective of
inflammation and fibrosis. World J Hepatol. 11:412–420.
2019.PubMed/NCBI View Article : Google Scholar
|
|
88
|
Tsuchida T and Friedman SL: Mechanisms of
hepatic stellate cell activation. Nat Rev Gastroenterol Hepatol.
14:397–411. 2017.PubMed/NCBI View Article : Google Scholar
|
|
89
|
Mak KM, Leo MA and Lieber CS: Alcoholic
liver injury in baboons: Transformation of lipocytes to
transitional cells. Gastroenterology. 87:188–200. 1984.PubMed/NCBI
|
|
90
|
Chen Z, Jain A, Liu H, Zhao Z and Cheng K:
Targeted drug delivery to hepatic stellate cells for the treatment
of liver fibrosis. J Pharmacol Exp Ther. 370:695–702.
2019.PubMed/NCBI View Article : Google Scholar
|
|
91
|
Lan T, Kisseleva T and Brenner DA:
Deficiency of NOX1 or NOX4 prevents liver inflammation and fibrosis
in mice through inhibition of hepatic stellate cell activation.
PLoS One. 10(e0129743)2015.PubMed/NCBI View Article : Google Scholar
|
|
92
|
Ou Q, Weng Y, Wang S, Zhao Y, Zhang F,
Zhou J and Wu X: Silybin alleviates hepatic steatosis and fibrosis
in NASH mice by inhibiting oxidative stress and involvement with
the Nf-κB Pathway. Dig Dis Sci. 63:3398–3408. 2018.PubMed/NCBI View Article : Google Scholar
|
|
93
|
Syn WK, Agboola KM, Swiderska M,
Michelotti GA, Liaskou E, Pang H, Xie G, Philips G, Chan IS, Karaca
GF, et al: NKT-associated hedgehog and osteopontin drive
fibrogenesis in non-alcoholic fatty liver disease. Gut.
61:1323–1329. 2012.PubMed/NCBI View Article : Google Scholar
|
|
94
|
Syn WK, Choi SS, Liaskou E, Karaca GF,
Agboola KM, Oo YH, Mi Z, Pereira TA, Zdanowicz M, Malladi P, et al:
Osteopontin is induced by hedgehog pathway activation and promotes
fibrosis progression in nonalcoholic steatohepatitis. Hepatology.
53:106–115. 2011.PubMed/NCBI View Article : Google Scholar
|
|
95
|
Grinstein S, Rotin D and Mason MJ:
Na+/H+ exchange and growth factor-induced
cytosolic pH changes. Role in cellular proliferation. Biochim
Biophys Acta. 988:73–97. 1989.PubMed/NCBI View Article : Google Scholar
|
|
96
|
Di Sario A, Baroni GS, Bendia E,
D'Ambrosio L, Ridolfi F, Marileo JR, Jezequel AM and Benedetti A:
Characterization of ion transport mechanisms regulating
intracellular pH in hepatic stellate cells. Am J Physiol.
273:G39–G48. 1997.PubMed/NCBI View Article : Google Scholar
|
|
97
|
Trappoliere M, Caligiuri A, Schmid M,
Bertolani C, Failli P, Vizzutti F, Novo E, di Manzano C, Marra F,
Loguercio C, et al: Silybin, a component of sylimarin, exerts
anti-inflammatory and anti-fibrogenic effects on human hepatic
stellate cells. J Hepatol. 50:1102–1111. 2009.PubMed/NCBI View Article : Google Scholar
|
|
98
|
Di Sario A, Bendia E, Svegliati Baroni G,
Ridolfi F, Bolognini L, Feliciangeli G, Jezequel AM, Orlandi F and
Benedetti A: Intracellular pathways mediating
Na+/H+ exchange activation by
platelet-derived growth factor in rat hepatic stellate cells.
Gastroenterology. 116:1155–1166. 1999.PubMed/NCBI View Article : Google Scholar
|
|
99
|
Svegliati-Baroni G, Di Sario A, Casini A,
Ferretti G, D'Ambrosio L, Ridolfi F, Bolognini L, Salzano R,
Orlandi F and Benedetti A: The Na+/H+
exchanger modulates the fibrogenic effect of oxidative stress in
rat hepatic stellate cells. J Hepatol. 30:868–875. 1999.PubMed/NCBI View Article : Google Scholar
|
|
100
|
Svegliati Baroni G, D'Ambrosio L, Ferretti
G, Casini A, Di Sario A, Salzano R, Ridolfi F, Saccomanno S,
Jezequel AM and Benedetti A: Fibrogenic effect of oxidative stress
on rat hepatic stellate cells. Hepatology. 27:720–726.
1998.PubMed/NCBI View Article : Google Scholar
|
|
101
|
Häussinger D and Schliess F: Osmotic
induction of signaling cascades: Role in regulation of cell
function. Biochem Biophys Res Commun. 255:551–555. 1999.PubMed/NCBI View Article : Google Scholar
|
|
102
|
Pendergrass WR, Angello JC, Kirschner MD
and Norwood TH: The relationship between the rate of entry into S
phase, concentration of DNA polymerase alpha, and cell volume in
human diploid fibroblast-like monokaryon cells. Exp Cell Res.
192:418–425. 1991.PubMed/NCBI View Article : Google Scholar
|
|
103
|
Vairo G, Cocks BG, Cragoe EJ Jr and
Hamilton JA: Selective suppression of growth factor-induced cell
cycle gene expression by Na+/H+ antiport
inhibitors. J Biol Chem. 267:19043–19046. 1992.PubMed/NCBI
|
|
104
|
Fontecave M, Lepoivre M, Elleingand E,
Gerez C and Guittet O: Resveratrol, a remarkable inhibitor of
ribonucleotide reductase. FEBS Lett. 421:277–279. 1998.PubMed/NCBI View Article : Google Scholar
|
|
105
|
Benedetti A, Di Sario A, Casini A, Ridolfi
F, Bendia E, Pigini P, Tonnini C, D'Ambrosio L, Feliciangeli G,
Macarri G, et al: Inhibition of the NA(+)/H(+) exchanger reduces
rat hepatic stellate cell activity and liver fibrosis: An in vitro
and in vivo study. Gastroenterology. 120:545–556. 2001.PubMed/NCBI View Article : Google Scholar
|
|
106
|
Huang Q, Li J, Zheng J and Wei A: The
carcinogenic role of the Notch signaling pathway in the development
of hepatocellular carcinoma. J Cancer. 10:1570–1579.
2019.PubMed/NCBI View Article : Google Scholar
|
|
107
|
Hardonnière K, Saunier E, Lemarié A,
Fernier M, Gallais I, Héliès-Toussaint C, Mograbi B, Antonio S,
Bénit P, Rustin P, et al: The environmental carcinogen
benzo[a]pyrene induces a Warburg-like metabolic reprogramming
dependent on NHE1 and associated with cell survival. Sci Rep.
6(30776)2016.PubMed/NCBI View Article : Google Scholar
|
|
108
|
Thyfault JP and Rector RS: Exercise
Combats Hepatic Steatosis: Potential Mechanisms and Clinical
Implications. Diabetes. 69:517–524. 2020.PubMed/NCBI View Article : Google Scholar
|
|
109
|
Harguindey S, Polo Orozco J, Alfarouk KO
and Devesa J: Hydrogen ion dynamics of cancer and a new molecular,
biochemical and metabolic approach to the etiopathogenesis and
treatment of brain malignancies. Int J Mol Sci.
20(4278)2019.PubMed/NCBI View Article : Google Scholar
|
|
110
|
Kim SW, Cha MJ, Lee SK, Song BW, Jin X,
Lee JM, Park JH and Lee JD: Curcumin treatment in combination with
glucose restriction inhibits intracellular alkalinization and tumor
growth in hepatoma cells. Int J Mol Sci. 20(2375)2019.PubMed/NCBI View Article : Google Scholar
|
|
111
|
Meima ME, Webb BA, Witkowska HE and Barber
DL: The sodium-hydrogen exchanger NHE1 is an Akt substrate
necessary for actin filament reorganization by growth factors. J
Biol Chem. 284:26666–26675. 2009.PubMed/NCBI View Article : Google Scholar
|
|
112
|
Yang X, Wang D, Dong W, Song Z and Dou K:
Over-expression of Na+/H+ exchanger 1 and its
clinicopathologic significance in hepatocellular carcinoma. Med
Oncol. 27:1109–1113. 2010.PubMed/NCBI View Article : Google Scholar
|
|
113
|
Yang X, Wang D, Dong W, Song Z and Dou K:
Suppression of Na+/H+ exchanger 1 by RNA
interference or amiloride inhibits human hepatoma cell line
SMMC-7721 cell invasion. Med Oncol. 28:385–390. 2011.PubMed/NCBI View Article : Google Scholar
|
|
114
|
Yang X, Wang D, Dong W, Song Z and Dou K:
Inhibition of Na(+)/H(+) exchanger 1 by 5-(N-ethyl-N-isopropyl)
amiloride reduces hypoxia-induced hepatocellular carcinoma invasion
and motility. Cancer Lett. 295:198–204. 2010.PubMed/NCBI View Article : Google Scholar
|
|
115
|
He X, Lee B and Jiang Y: Cell-ECM
Interactions in Tumor Invasion. Adv Exp Med Biol. 936:73–91.
2016.PubMed/NCBI View Article : Google Scholar
|
|
116
|
Stüwe L, Müller M, Fabian A, Waning J,
Mally S, Noël J, Schwab A and Stock C: pH dependence of melanoma
cell migration: Protons extruded by NHE1 dominate protons of the
bulk solution. J Physiol. 585:351–360. 2007.PubMed/NCBI View Article : Google Scholar
|
|
117
|
Reshkin SJ, Cardone RA and Harguindey S: :
Na+-H+ exchanger, pH regulation and cancer.
Recent Patents Anticancer Drug Discov. 8:85–99. 2013.PubMed/NCBI View Article : Google Scholar
|
|
118
|
Keurhorst D, Liashkovich I, Frontzek F,
Nitzlaff S, Hofschröer V, Dreier R and Stock C: MMP3 activity
rather than cortical stiffness determines NHE1-dependent
invasiveness of melanoma cells. Cancer Cell Int.
19(285)2019.PubMed/NCBI View Article : Google Scholar
|
|
119
|
He B, Deng C, Zhang M, Zou D and Xu M:
Reduction of intracellular pH inhibits the expression of VEGF in
K562 cells after targeted inhibition of the
Na+/H+ exchanger. Leuk Res. 31:507–514.
2007.PubMed/NCBI View Article : Google Scholar
|
|
120
|
Apte RS, Chen DS and Ferrara N: VEGF in
Signaling and Disease: Beyond Discovery and Development. Cell.
176:1248–1264. 2019.PubMed/NCBI View Article : Google Scholar
|
|
121
|
Alfarouk KO: Tumor metabolism, cancer cell
transporters, and microenvironmental resistance. J Enzyme Inhib Med
Chem. 31:859–866. 2016.PubMed/NCBI View Article : Google Scholar
|
|
122
|
Gilkes DM, Semenza GL and Wirtz D: Hypoxia
and the extracellular matrix: Drivers of tumour metastasis. Nat Rev
Cancer. 14:430–439. 2014.PubMed/NCBI View Article : Google Scholar
|
|
123
|
Vaupel P, Kallinowski F and Okunieff P:
Blood flow, oxygen and nutrient supply, and metabolic
microenvironment of human tumors: A review. Cancer Res.
49:6449–6465. 1989.PubMed/NCBI
|
|
124
|
Sun Y, Liu WZ, Liu T, Feng X, Yang N and
Zhou HF: Signaling pathway of MAPK/ERK in cell proliferation,
differentiation, migration, senescence and apoptosis. J Recept
Signal Transduct Res. 35:600–604. 2015.PubMed/NCBI View Article : Google Scholar
|
|
125
|
Chen LC, Shibu MA, Liu CJ, Han CK, Ju DT,
Chen PY, Viswanadha VP, Lai CH, Kuo WW and Huang CY: ERK1/2
mediates the lipopolysaccharide-induced upregulation of FGF-2, uPA,
MMP-2, MMP-9 and cellular migration in cardiac fibroblasts. Chem
Biol Interact. 306:62–69. 2019.PubMed/NCBI View Article : Google Scholar
|
|
126
|
Wang JC, Chen SY, Wang M, Ko JL, Wu CL,
Chen CC, Lin HW and Chang YY: Nickel-induced VEGF expression via
regulation of Akt, ERK1/2, NFκB, and AMPK pathways in H460 cells.
Environ Toxicol. 34:652–658. 2019.PubMed/NCBI View Article : Google Scholar
|
|
127
|
Yang X, Wang D, Dong W, Song Z and Dou K:
Expression and modulation of Na(+) /H(+) exchanger 1 gene in
hepatocellular carcinoma: A potential therapeutic target. J
Gastroenterol Hepatol. 26:364–370. 2011.PubMed/NCBI View Article : Google Scholar
|
|
128
|
Xu J, Ji B, Wen G, Yang Y, Jin H, Liu X,
Xie R, Song W, Song P, Dong H, et al: Na+/H+
exchanger 1, Na+/Ca2+ exchanger 1 and
calmodulin complex regulates interleukin 6-mediated cellular
behavior of human hepatocellular carcinoma. Carcinogenesis.
37:290–300. 2016.PubMed/NCBI View Article : Google Scholar
|
|
129
|
Huc L, Sparfel L, Rissel M,
Dimanche-Boitrel MT, Guillouzo A, Fardel O and Lagadic-Gossmann D:
Identification of Na+/H+ exchange as a new
target for toxic polycyclic aromatic hydrocarbons. FASEB J.
18:344–346. 2004.PubMed/NCBI View Article : Google Scholar
|
|
130
|
Hardonnière K, Saunier E, Lemarié A,
Fernier M, Gallais I, Héliès-Toussaint C, Mograbi B, Antonio S,
Bénit P, Rustin P, et al: The environmental carcinogen
benzo[a]pyrene induces a Warburg-like metabolic reprogramming
dependent on NHE1 and associated with cell survival. Sci Rep.
6(30776)2016.PubMed/NCBI View Article : Google Scholar
|
|
131
|
Dendelé B, Tekpli X, Hardonnière K, Holme
JA, Debure L, Catheline D, Arlt VM, Nagy E, Phillips DH, Ovrebø S,
et al: Protective action of n-3 fatty acids on
benzo[a]pyrene-induced apoptosis through the plasma membrane
remodeling-dependent NHE1 pathway. Chem Biol Interact. 207:41–51.
2014.PubMed/NCBI View Article : Google Scholar
|
|
132
|
Wang J, Tian L, Khan MN, Zhang L, Chen Q,
Zhao Y, Yan Q, Fu L and Liu J: Ginsenoside Rg3 sensitizes hypoxic
lung cancer cells to cisplatin via blocking of NF-κB mediated
epithelial-mesenchymal transition and stemness. Cancer Lett.
415:73–85. 2018.PubMed/NCBI View Article : Google Scholar
|
|
133
|
Zhang C, Liu L, Yu Y, Chen B, Tang C and
Li X: Antitumor effects of ginsenoside Rg3 on human hepatocellular
carcinoma cells. Mol Med Rep. 5:1295–1298. 2012.PubMed/NCBI View Article : Google Scholar
|
|
134
|
Zhou B, Wang J and Yan Z: Ginsenoside Rg3
attenuates hepatoma VEGF overexpression after hepatic artery
embolization in an orthotopic transplantation hepatocellular
carcinoma rat model. OncoTargets Ther. 7:1945–1954. 2014.PubMed/NCBI View Article : Google Scholar
|
|
135
|
Zhou B, Yan Z, Liu R, Shi P, Qian S, Qu X,
Zhu L, Zhang W and Wang J: Prospective Study of Transcatheter
Arterial Chemoembolization (TACE) with Ginsenoside Rg3 versus TACE
Alone for the Treatment of Patients with Advanced Hepatocellular
Carcinoma. Radiology. 280:630–639. 2016.PubMed/NCBI View Article : Google Scholar
|
|
136
|
Li X, Tsauo J, Geng C, Zhao H, Lei X and
Li X: Ginsenoside Rg3 Decreases NHE1 Expression via Inhibiting
EGF-EGFR-ERK1/2-HIF-1 [Formula: see text] Pathway in Hepatocellular
Carcinoma: A Novel Antitumor Mechanism. Am J Chin Med.
46:1915–1931. 2018.PubMed/NCBI View Article : Google Scholar
|
|
137
|
Kloeckner R, Ruckes C, Kronfeld K, Wörns
MA, Weinmann A, Galle PR, Lang H, Otto G, Eichhorn W,
Schreckenberger M, et al: Selective internal radiotherapy (SIRT)
versus transarterial chemoembolization (TACE) for the treatment of
intrahepatic cholangiocellular carcinoma (CCC): study protocol for
a randomized controlled trial. Trials. 15(311)2014.PubMed/NCBI View Article : Google Scholar
|
|
138
|
Uchida D, Takaki A, Ishikawa H, Tomono Y,
Kato H, Tsutsumi K, Tamaki N, Maruyama T, Tomofuji T, Tsuzaki R, et
al: Oxidative stress balance is dysregulated and represents an
additional target for treating cholangiocarcinoma. Free Radic Res.
50:732–743. 2016.PubMed/NCBI View Article : Google Scholar
|
|
139
|
Grek A and Arasi L: Acute liver failure.
AACN Adv Crit Care. 27:420–429. 2016.PubMed/NCBI View Article : Google Scholar
|
|
140
|
Ezquerro S, Mocha F, Frühbeck G,
Guzmán-Ruiz R, Valentí V, Mugueta C, Becerril S, Catalán V,
Gómez-Ambrosi J, Silva C, et al: Ghrelin Reduces TNF-α-Induced
Human Hepatocyte Apoptosis, Autophagy, and Pyroptosis: Role in
Obesity-Associated NAFLD. J Clin Endocrinol Metab. 104:21–37.
2019.PubMed/NCBI View Article : Google Scholar
|
|
141
|
Liu Z, Wang S, Zhou H, Yang Y and Zhang M:
Na+/H+ exchanger mediates TNF-alpha-induced
hepatocyte apoptosis via the calpain-dependent degradation of
Bcl-xL. J Gastroenterol Hepatol. 24:879–885. 2009.PubMed/NCBI View Article : Google Scholar
|
|
142
|
Alexander RT, Dimke H and Cordat E:
Proximal tubular NHEs: Sodium, protons and calcium? Am J Physiol
Renal Physiol. 305:F229–F236. 2013.PubMed/NCBI View Article : Google Scholar
|
|
143
|
Shi M, Zhang T, Sun L, Luo Y, Liu DH, Xie
ST, Song XY, Wang GF, Chen XL, Zhou BC and Zhang YZ: Calpain, Atg5
and Bak play important roles in the crosstalk between apoptosis and
autophagy induced by influx of extracellular calcium. Apoptosis.
18:435–451. 2013.PubMed/NCBI View Article : Google Scholar
|
|
144
|
Huang C, Wang J, Chen Z, Wang Y and Zhang
W: 2-phenylethynesulfonamide Prevents Induction of Pro-inflammatory
Factors and Attenuates LPS-induced Liver Injury by Targeting
NHE1-Hsp70 Complex in Mice. PLoS One. 8(e67582)2013.PubMed/NCBI View Article : Google Scholar
|
|
145
|
Ceccarelli S, Panera N, Mina M, Gnani D,
De Stefanis C, Crudele A, Rychlicki C, Petrini S, Bruscalupi G,
Agostinelli L, et al: LPS-induced TNF-α factor mediates
pro-inflammatory and pro-fibrogenic pattern in non-alcoholic fatty
liver disease. Oncotarget. 6:41434–41452. 2015.PubMed/NCBI View Article : Google Scholar
|
|
146
|
Liu CL, Zhang X, Liu J, Wang Y, Sukhova
GK, Wojtkiewicz GR, Liu T, Tang R, Achilefu S, Nahrendorf M, et al:
Na+-H+ exchanger 1 determines atherosclerotic
lesion acidification and promotes atherogenesis. Nat Commun.
10(3978)2019.PubMed/NCBI View Article : Google Scholar
|
|
147
|
Guan X, Hasan MN, Begum G, Kohanbash G,
Carney KE, Pigott VM, Persson AI, Castro MG, Jia W and Sun D:
Blockade of Na/H exchanger stimulates glioma tumor immunogenicity
and enhances combinatorial TMZ and anti-PD-1 therapy. Cell Death
Dis. 9(1010)2018.PubMed/NCBI View Article : Google Scholar
|
|
148
|
Rotstein OD, Houston K and Grinstein S:
Control of cytoplasmic pH by Na+/H+ exchange
in rat peritoneal macrophages activated with phorbol ester. FEBS
Lett. 215:223–227. 1987.PubMed/NCBI View Article : Google Scholar
|
|
149
|
Ye Y, Jia X, Bajaj M and Birnbaum Y:
Dapagliflozin Attenuates Na+/H+ Exchanger-1
in Cardiofibroblasts via AMPK Activation. Cardiovasc Drugs Ther.
32:553–558. 2018.PubMed/NCBI View Article : Google Scholar
|
|
150
|
Ryuichi O, Masafumi M and Hiroshi K:
Localization, ion transport activity, and physiological function of
mammalian organellar NHEs. Seikagaku. J Jpn Biochem Soc.
82:2010.PubMed/NCBI(In Japanese).
|
|
151
|
Karmazyn M: Pharmacology and clinical
assessment of cariporide for the treatment coronary artery
diseases. Expert Opin Investig Drugs. 9:1099–1108. 2000.PubMed/NCBI View Article : Google Scholar
|