1. Introduction
Nicotinamide adenine dinucleotide (NAD+)
is considered the precursor for the pyridine nucleotide family
constituted by NADH and two phosphorylated forms, nicotinamide
adenine dinucleotide phosphate in its oxidized (NADP+)
and reduced (NADPH) forms (1). All
of these nucleotides are well-known cofactors in numerous cellular
processes. NAD+ is the substrate for major redox
transformations of nutrients during the catabolic phase of
metabolism. Its reduced form, NADH, is the main source of electrons
for mitochondrial oxidative chain and production of adenosine
triphosphate through oxidative phosphorylation. NAD+ is
also involved in DNA repair, as a substrate for poly(ADP-ribose)
polymerases (PARP) (2), epigenetic
modification of gene expression, also having an important influence
on immunological function (3).
NAD+ also has a critical role in calcium homeostasis, as
the substrate for NAD+ glycohydrolases, enzymes related
to the production of cyclic ADP-ribose, a calcium efflux effector
(4).
Since the original discovery of NAD+ by
Harden and Young in 1906 in cell-free yeast juices as a factor that
enhanced the rate of fermentation, many studies have been developed
to reveal the NAD+ biosynthetic pathway as a key player
in cellular metabolism (5).
NAD+ is currently regarded as an important target with
which to extend lifespan and health span. Accurate evaluation of
the NAD+ metabolome is of great interest due to its
association with cognitive impairment, cancer, normal aging and
age-related disorders (1).
Research conducted to date has elucidated the
signaling pathways and cellular processes that contribute to the
maintenance of pyridine nucleoside and nucleotide homeostasis; the
regulation of molecular mechanisms underlying NAD+
metabolism is known but the use of its precursors in therapy are
still incompletely explained.
2. NAD+ biosynthesis, metabolism
and effects on the human body
NAD+ biosynthesis in mammals involves
five important intermediates including tryptophan, niacin,
nicotinamide riboside (NR) and nicotinamide mononucleotide (NMN),
through three biosynthetic pathways: i) Nucleotide salvage from
nicotinamide (NAM), NR and NMN; ii) de novo synthesis from
tryptophan and iii) Preiss-Handler pathway from nicotinic acid (NA)
(6).
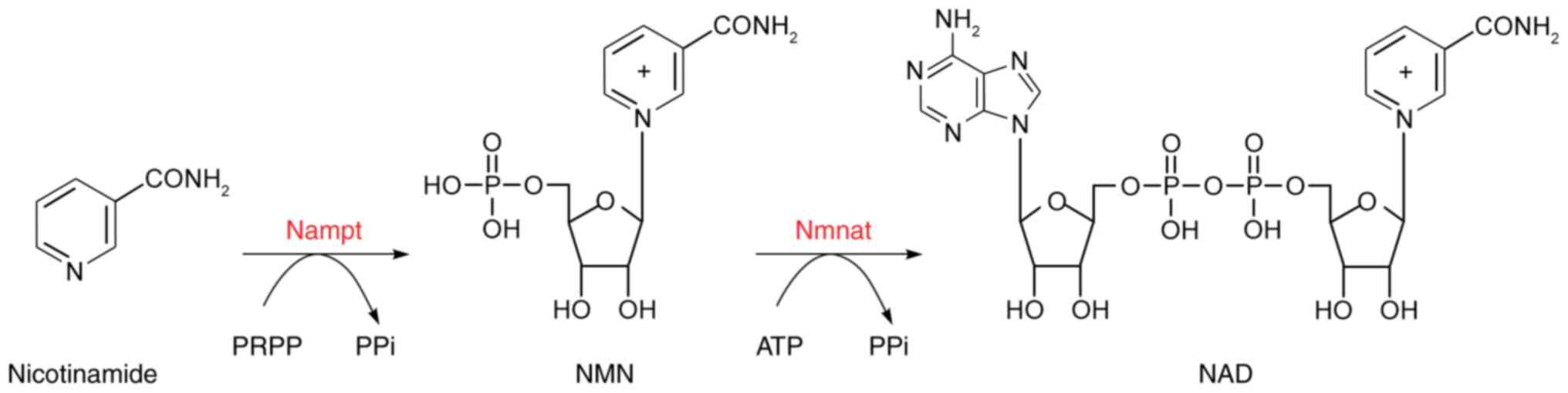
In mammals, the salvage pathway is the major route
to generate NAD+. Conversion of NMN, one of the forms of
the water-soluble vitamin B3, to NAD+ is catalyzed by
nicotinamide mononucleotide adenylyltransferase (Nmnat) (Fig. 1). NMN can be synthesized by
nicotinamide phosphoribosyltransferase (Nampt) from NAM.
Bieganowski and Brenner revealed an alternative NAD+
salvage biosynthesis pathway, in which NR, another form of vitamin
B3 that enters cells through nucleoside transporters, could be
converted to NAD+ by nicotinamide riboside kinases
(NMRK1 and NMRK2) and Nmnat via phosphorylation reactions (7).
In recent years, NMN and NR have been extensively
investigated in various experiments on rodents and humans and
evidence suggests that with age NAD+ levels decline at a
systemic level (8,9), causing profound metabolic changes
(10). These precursors of
NAD+ have an essential influence on the elevation of
NAD+ concentration in a variety of tissues,
automatically suggesting a beneficial therapeutic effect (8,11,12).
The metabolic balance of NAD+ in the
cellular environment can be accomplished in several ways and
involves multiple metabolites in an oxidized and reduced form such
as NADP+, NADH, NADPH, and nicotinic acid adenine
dinucleotide phosphate (NAADP), all with an essential role in
energy production and cellular metabolism, acting as one
proton-accepting or donating coenzymes. NAD+ derivatives
are important for regulating cellular redox status, intracellular
Ca2+ pools, DNA damage and repair, cell cycle timing and
lipid and energy metabolism (13).
NAD+ is a major cofactor for mitochondrial ATP
production and for NAD-dependent enzymes including sirtuins and
poly-ADP-ribosylpolymerases, essential players in fundamental
processes such as cell division and proliferation, apoptosis,
aging, senescence and stress resistance (14). Lin and Guarente argued that sirtuins
can sense the NADH/NAD+ ratio in cells by the
recognition of the oxidized dinucleotides, but it seems that other
cellular factors also respond to the alterations within NADH levels
(15).
For many years it was believed that NAD+
breakdown is a nonspecific process but in the last decade evidence
has shown that NAD+ consumption is linked with the
signaling reactions inside and outside the cells (16). For example, the oxidation of glucose
and fatty acids lead to the reduction of oxidized NAD+
to NADH and alterations in its concentration were found to be
involved in multiple pathogenic signaling pathways, including heart
failure (17). The last two decades
generated multiple discoveries regarding NAD+ and its
precursors, identifying the important role in DNA repair, immune
activation and epigenetic control through protein deacetylation
(3). Even if the metabolism of
NAD+ has been the subject of numerous research studies
in various cells, tissues, and organs, those regarding pyridine
nucleotide metabolism in eye structures and its involvement in
visual dysfunctions are scarce.
3. Diet and NAD+ level
Dietary supplementation is the only way to increase
body NAD+ levels. Since NAD+ administration
is not efficient enough to increase NAD+ levels, its
precursors, such as NAM and NA known as niacin, NMN and NR could be
useful to increase NAD+ levels in animal models and
humans (18).
Most raw foods provide these substances which can
regulate cellular activities and the timing of changes that lead to
aging phenotypes (13). Another
important precursor of NAD+ is the amino acid tryptophan
which is synthetized via the kynurenine pathway. While NR is the
form of vitamin B3 found in humans and cow milk and other foods, NA
is produced by plants and algae, and tryptophan is the most
abundant amino acid found in animal and plant proteins (19).
Many of the biochemical studies performed recently
have shown that a poor diet lacking niacin and tryptophan intake or
a chronic immune activation can lead to inefficient production of
NAD+, where catabolism exceeds anabolism producing a
consequent cellular dysfunction (20). By contrast, a reduced energy load
due to activities such exercise, calorie restriction, fasting and
glucose deprivation can increase the concentration of
NAD+ (8).
Although studies on rodents have shown that both NMN
and NR enhance NAD+ biosynthesis and have beneficial
effects in multiple disorders (21-23),
it still remains unclear what mechanisms mediate their beneficial
effect. Presently, the pharmacokinetics and metabolic fates of
NAD+ precursors are still under investigation, multiple
human clinical trials being conducted to study and understand the
safety of NAD+ precursor supplementation, mainly NR and
NMN (24-26).
The concentrations of NAD+ and NAM in blood are in the
micromolar range and can be boosted by oral administration of their
precursors, NR and NMN. In their study, Trammell et al
showed that a daily dose of 1000 mg NR leads to a 2.7-fold rise in
blood NAD+ after one dose of NR (24). Another clinical study demonstrated
that daily doses of NR up to 1000 mg are well tolerated and
efficient to increase NAD+ levels by almost 60% in
peripheral blood mononuclear cells (25). Irie et al conducted a human
study regarding the safety of a single oral NMN dose (100, 250 or
500 mg) in healthy Japanese men by investigating the
pharmacokinetics of NMN metabolites for 5 h after each intervention
and found that NMN supplementation is safe without causing any
significant deleterious effects (27).
4. NAD+ and diseases
Multiple studies strongly confirm that deficiencies
in NAD+ levels can lead to degenerative diseases and
multiple pathologies such as metabolic disorders, heart and renal
diseases, cognitive impairment and even cancer (28,29).
In the past few years, research on the biology of
NAD+ has provided many critical insights into the
pathogenesis of age-associated functional decline (30).
Studies on rodents have demonstrated that under
normal and pathophysiological conditions, systemic NMN
administration enhanced NAD+ biosynthesis in various
peripheral tissues such as the pancreas, liver and white adipose
tissue of diabetic mice (21),
heart (31,32), skeletal muscle (33), kidney (34,35),
eyes (36) and blood vessels
(37,38). Recent studies on mouse and rat
models suggest that NMN improves the neuronal function with visible
benefits on cognition and memory (39-41).
NR is also a direct factor involved in
NAD+ metabolism with a recognized efficacy in metabolic
and skeleton-muscular disorders. Similar to NMN, NR also exhibits
protective effects against aging and age-related diseases, improves
liver health and protects against diabetic and chemotherapy induced
neuropathy (42).
In the past few years, metabolic syndrome has become
a global health concern due to its critical risk in regards to the
development of various life-threatening conditions such as
cardiovascular disease, stroke and cancer (43). Obesity, diabetes, dyslipidemia,
fatty liver and hypertension are common diseases linked with the
metabolic syndrome and are closely associated with impaired
nutrient status and lifestyle (44). A number of studies have demonstrated
that NAD+ levels decline with aberrant nutritional
status and its prevention is regarded as a promising strategy with
which to combat metabolic disorders (18). Administration of NMN and NR can
ameliorate diet-associated weight gain, dietary intervention, their
use being a promising treatment against obesity (45). In addition, the administration of NR
in mice was found to increase fatty acid oxidation and energy
expenditure and also to improve insulin sensitivity (45). On other hand, studies on various
diabetic rodent models, have revealed that NAM administration
ameliorates hyperglycemia by increasing β-cell proliferation
(46).
Recent studies indicate that NAD+ is
involved in tumor cell progression being considered a promising
therapeutic target for cancer (47-49).
NAD+-metabolizing enzymes play an important role in
conditioning several aspects of cancer and immune cell fate and
functions (50). Cancer cells
display a unique energy metabolism and the continuous replenishment
of NAD+ promotes the proliferation and survival of
fast-dividing cancer cells as elevated NAD+ levels
enhance anaerobic glycolysis (51).
Increased intracellular NAD+ levels accompanied by
overexpression of NAD+ salvage biosynthesis pathway
enzymes sustain tumor cell proliferation and promote survival
against anticancer cell agents (52).
Overexpression of Nampt is frequently observed in
several types of malignant tumors, for example, breast, colorectal,
gastric, ovarian, prostate cancers, and malignant lymphomas
(48,52). A recent study showed that high
expression of Nampt and Naprt predicts a poor prognosis in
colorectal cancer and was associated with vascular invasion. This
finding suggests that these two enzymes of the NAD+
salvage synthesis pathway may be novel markers for the diagnosis
and treatment of colorectal cancer (53).
5. NAD+ in normal and abnormal
function of the retina
The retina is a highly organized eye structure,
light-sensitive, intensively innervated and vascularized. It
consists of numerous cell types that function in a coordinated
manner to give rise to a signal transmitted to the brain. To create
an image, light photons are sensed by the retinal receptors and the
signal from the photoreceptors is then transmitted to secondary
neurons before being transmitted to the retinal ganglion cells.
Retinal photoreceptor death is a common cause of blindness in
retinal degenerative diseases (54-56).
Retinal degenerative diseases are a major cause of
morbidity because visual impairment significantly decreases the
quality of life of patients. These have different etiologies, being
either acquired, clinical manifestation of a systemic problem or
inherited (54).
Despite their diversity, these diseases focus on the
death of photoreceptors. The huge metabolic and energetic
requirements of the retinal photoreceptors and their progressive
impairment in various retinal degenerative diseases have been for a
long time the subject of research.
Given the key metabolic role of NAD+ and
the high metabolism in photoreceptor cells, there is a growing
interest in debating the role of the ‘NADome’ in controlling
retinal metabolism and mediating the pathogenesis of retinal
degeneration.
The involvement of NAD+ biosynthesis
pathways in retinal photoreceptor dysfunction has been previously
investigated. The mutation of Nmnat1, one of the key enzymes
involved in NAD+ biosynthesis, was identified as a cause
of a childhood blinding disease (57). Rodent studies that replicated this
mutation showed the same alterations (for example, rapidly
photoreceptor degeneration, retinal pigment epithelial cell loss,
retinal vasculature attenuation) and confirm its importance for
retinal survival (58).
Data reported by Kaja and colleagues showed that
altered circulating levels of Nampt were also significantly
associated with risk for retinal vein occlusion, ischemia and
metabolic alteration (59). Other
studies confirmed the relevance of Nampt for retinal function
proving its importance for maintenance of functional retinal
pigment epithelial and endothelial cells. For example, inhibition
of Nampt with pharmacological inhibitors or deficits in
NAD+ bioavailability promoted an early senescent
phenotype in these cells (60,61).
NAD+ metabolism is also important for
cell longevity. Age-dependent decline in NAD+ levels
reported for multiple human organs has been demonstrated to occur
also in the retina, especially in photoreceptor, ganglion,
endothelial, and retinal pigment epithelial cells (62). That is why extensive research has
focused on the role of NAD+ as a modulator of sirtuin
activity, known that sirtuins play important roles in the retina,
confer protection against oxidative stress and retinal degeneration
being able to increase lifespan (63). All seven mammal sirtuins (SRT1-SRT7)
are highly expressed in the mouse retina (64) and could enhance retinal metabolism,
reduce photoreceptor death, and improve vision.
Given the roles of sirtuins in retinal function,
pharmacological modulation of sirtuin activation may be a
therapeutic strategy for preventing retinal degeneration. The
supply with NAD+ precursors, such as NMN and NR,
represent an interesting approach to this condition. In a mouse
model of light-induced acute retinal degeneration, Zhang and
colleagues found that NR treatment prevented NAD+
diminution following toxic light exposure (65). Maintenance of NAD+ levels
allowed continued activity of enzymes for which NAD+ is
a substrate, for example mitochondrial sirtuins, on which optimal
activity relies NAD+-dependent retinal homeostasis
(66). Supplementation with these
precursors was found to lead to increased intracellular
NAD+ bioavailability and enhanced sirtuin function,
especially in contexts of cellular stress when NAD+
requirements may be increased and have been shown to be critical to
retinal health. Further studies are important to ensure the
development of new safe and efficient therapeutic strategies.
6. Conclusions
While the first century of NAD+ research
has revealed multiple discoveries which prove that NAD+
plays a key role in the regulation of cell metabolism, stress and
immune responses to physiological or pathological signals, it is
widely recognized that additional research is required to fully
reveal the complex metabolic fate of the ‘NADome’ in order to
sustain the paramount importance of the supplementation of
NAD+ precursors to promote health and improve
therapeutic safety in various diseases, including retinal
disturbances.
Acknowledgements
Not applicable.
Funding
Funding: No funding was received.
Availability of data and materials
All information discussed in this review is
documented by relevant references.
Authors' contributions
ASP, AMA, ECS, IMB, CGP, SJ and RC made equal
contributions to the conception and editing of this manuscript.
ASP, AMA, ECS, CGP, SJ and RC contributed to the acquisition of the
data. ASP, AMA, ECS, CGP, IMB, SJ and RC selected, processed the
data acquired and wrote appropriate parts of this manuscript. SJ,
RC, CGP and IMB revised its contents. All authors read and approved
the final manuscript to be published.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Clement J, Wong M, Poljak A, Sachdev P and
Braidy N: The plasma NAD+ metabolome is dysregulated in
‘normal’ aging. Rejuvenation Res. 22:121–130. 2019.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Surjana D, Halliday GM and Damian DL: Role
of nicotinamide in DNA damage, mutagenesis, and DNA repair. J
Nucleic Acids. 2010(157591)2010.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Braidy N, Beg J, Clement J, Khorshidi F,
Poljak A, Jayasena T, Grant R and Sachdev P: Role of nicotinamide
adenine dinucleotide and related precursors as therapeutic targets
for age-related degenerative diseases: Rationale, biochemistry,
pharmacokinetics, and outcomes. Antioxid Redox Signal. 30:251–294.
2019.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Horenstein AL, Sizzano F, Lusso R, Besso
FG, Ferrero E, Deaglio S, Corno F and Malavasi F: CD38 and CD157
ectoenzymes mark cell subsets in the human corneal limbus. Mol Med.
15:76–84. 2009.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Croft T, Venkatakrishnan P and Lin SJ:
NAD+ metabolism and regulation: Lessons from yeast.
Biomolecules. 10(330)2020.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Wherry EJ and Kurachi M: Molecular and
cellular insights into T cell exhaustion. Nat Rev Immunol.
15:486–499. 2015.PubMed/NCBI View
Article : Google Scholar
|
|
7
|
Bieganowski P and Brenner C: Discoveries
of nicotinamide riboside as a nutrient and conserved NRK genes
establish a Preiss-Handler independent route to NAD+ in
fungi and humans. Cell. 117:495–502. 2004.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Canto C, Menzies KJ and Auwerx J:
NAD+ metabolism and the control of energy homeostasis: A
balancing act between mitochondria and the nucleus. Cell Metab.
22:31–53. 2015.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Verdin E: NAD+ in aging,
metabolism, and neurodegeneration. Science. 350:1208–1213.
2015.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Finley LW and Haigis MC: The coordination
of nuclear and mitochondrial communication during aging and calorie
restriction. Age Res Rev. 8:173–188. 2009.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Chini CC, Tarrago MG and Chini EN: NAD and
the aging process: Role in life, death and everything in between.
Mol Cell Endocrinol. 455:62–74. 2017.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Fang EF, Lautrup S, Hou Y, Demarest TG,
Croteau DL, Mattson MP and Bohr VA: NAD+ in aging:
Molecular mechanisms and translational implications. Trends Mol
Med. 23:899–916. 2017.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Xu P and Sauve A: Vitamin B3, the
nicotinamide adenine dinucleotides and aging. Mech Ageing Dev.
131:287–298. 2010.PubMed/NCBI View Article : Google Scholar
|
|
14
|
van de Ven RA, Santos D and Haigis MC:
Mitochondrial sirtuins and molecular mechanisms of aging. Trends
Mol Med. 23:320–331. 2017.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Lin SJ and Guarente L: Nicotinamide
adenine dinucleotide, a metabolic regulator of transcription,
longevity and disease. Curr Opin Cell Biol. 15:241–246.
2003.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Belenky P, Bogan KL and Brenner C: NAD+
metabolism in health and disease. Trends Biochem Sci. 32:12–19.
2007.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Mericskay M: Nicotinamide adenine
dinucleotide homeostasis and signalling in heart disease:
Pathophysiological implications and therapeutic potential. Arch
Cardiovasc Dis. 109:207–215. 2016.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Okabe K, Yaki K, Tobe K and Nakagawa T:
Implications of altered NAD metabolism in metabolic disorders. J
Biomed Sci. 26(34)2019.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Trammell SA, Yu L, Redpath P, Migaud ME
and Brenner C: Nicotinamide riboside is a major NAD+ precursor
vitamin in cow milk. J Nutr. 146:957–963. 2016.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Tang K, Sham H, Hui E and Kirkland JB:
Niacin deficiency causes oxidative stress in rat bone marrow cells
but not through decreased NADPH or glutathione status. J Nutr
Biochem. 19:746–753. 2008.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Yoshino J, Mills KF, Yoon MJ and Imai SI:
Nicotinamide mononucleotide, a key NAD(+) intermediate, treats the
pathophysiology of diet- and age-induced diabetes in mice. Cell
Metab. 14:528–536. 2011.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Zhang H, Ryu D, Wu Y, Gariani K, Wang X,
Luan P, D'Amico D, Ropelle ER, Lutolf MP, Aebersold R, et al:
NAD+ repletion improves mitochondrial and stem cell
function and enhances life span in mice. Science. 352:1436–1443.
2016.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Mills KF, Yoshida S, Stein LR, Grozio A,
Kubota S, Sasaki Y, Redpath P, Migaud ME, Apte RS, Uchida K, et al:
Long-term administration of nicotinamide mononucleotide mitigates
age-associated physiological decline in mice. Cell Metab.
24:795–806. 2016.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Trammell SAJ, Schmidt MS, Weidemann BJ,
Redpath P, Jaksch F, Dellinger RW, Li Z, Dale Abel E, Migaud ME and
Brenner C: Nicotinamide riboside is uniquely and orally
bioavailable in mice and humans. Nat Commun.
7(12948)2016.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Martens CR, Denman BA, Mazzo MR, Armstrong
ML, Reisdorph N, McQueen MB, Chonchol M and Seals DR: Chronic
nicotinamide riboside supplementation is well-tolerated and
elevates NAD+ in healthy middle-aged and older adults.
Nat Commun. 9(1286)2018.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Elhassan YS, Kluckova K, Fletcher RS,
Schmidt MS, Garten A, Doig CL, Cartwright DM, Oakey L, Burley CV,
Jenkinson N, et al: Nicotinamide riboside augments the aged human
skeletal muscle NAD+ metabolome and induces
transcriptomic and anti-inflammatory signatures. Cell Rep.
28:1717–1728.e6. 2019.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Irie J, Inagaki E, Fujita M, Nakaya H,
Mitsuishi M, Yamaguchi S, Yamashita K, Shigaki S, Ono T, Yukioka H,
et al: Effect of oral administration of nicotinamide mononucleotide
on clinical parameters and nicotinamide metabolite levels in
healthy Japanese men. Endocr J. 67:153–160. 2020.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Ralto KM, Rhee EP and Parikh SM:
NAD+ homeostasis in renal health and disease. Nat Rev
Nephrol. 16:99–111. 2020.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Katsyuba E, Romani M, Hofer D and Auwerx
J: NAD+ homeostasis in health and disease. Nat Metab.
2:9–31. 2020.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Johnson S and Imai S: NAD+
biosynthesis, aging, and disease. F1000Res. 7(132)2018.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Yamamoto T, Byun J, Zhai P, Ikeda Y, Oka S
and Sadoshima J: Nicotinamide mononucleotide, an intermediate of
NAD+ synthesis, protects the heart from ischemia and
reperfusion. PLoS One. 9(e98972)2014.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Martin AS, Abraham DM, Hershberger KA,
Bhatt DP, Mao L, Cui H, Liu J, Liu X, Muehlbauer MJ, Grimsrud PA,
et al: Nicotinamide mononucleotide requires SIRT3 to improve
cardiac function and bioenergetics in a Friedreich's ataxia
cardiomyopathy model. JCI Insight. 2(e93885)2017.PubMed/NCBI View Article : Google Scholar
|
|
33
|
Ryu D, Zhang H, Ropelle ER, Sorrentino V,
Mázala DAG, Mouchiroud L, Marshall PL, Campbell MD, Ali AS, Knowels
GM, et al: NAD+ repletion improves muscle function in muscular
dystrophy and counters global PARylation. Sci Transl Med.
8(361ra139)2016.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Guan Y, Wang SR, Huang XZ, Xie QH, Xu YY,
Shang D and Hao CM: Nicotinamide mononucleotide, an NAD+
precursor, rescues age-associated susceptibility to AKI in a
sirtuin 1-dependent manner. J Am Soc Nephrol. 28:2337–2352.
2017.PubMed/NCBI View Article : Google Scholar
|
|
35
|
Yoshino J, Baur JA and Imai SI:
NAD+ intermediates: The biology and therapeutic
potential of NMN and NR. Cell Metab. 27:513–528. 2018.PubMed/NCBI View Article : Google Scholar
|
|
36
|
Lin JB, Kubota S, Ban N, Yoshida M,
Santeford A, Sene A, Nakamura R, Zapata N, Kubota M, Tsubota K, et
al: NAMPT-mediated NAD(+) biosynthesis is essential for vision in
mice. Cell Rep. 17:69–85. 2016.PubMed/NCBI View Article : Google Scholar
|
|
37
|
Zhao C, Li W, Duan H, Li Z, Jia Y, Zhang
S, Wang X, Zhou Q and Shi W: NAD+ precursors protect
corneal endothelial cells from UVB-induced apoptosis. Cell Physiol.
318:C796–C805. 2020.PubMed/NCBI View Article : Google Scholar
|
|
38
|
de Picciotto NE, Gano LB, Johnson LC,
Martens CR, Sindler AL, Mills KF, Imai S and Seals DR: Nicotinamide
mononucleotide supplementation reverses vascular dysfunction and
oxidative stress with aging in mice. Aging Cell. 15:522–530.
2016.PubMed/NCBI View Article : Google Scholar
|
|
39
|
Wang X, Hu X, Yang Y, Takata T and Sakurai
T: Nicotinamide mononucleotide protects against β-amyloid
oligomer-induced cognitive impairment and neuronal death. Brain
Res. 1643:1–9. 2016.PubMed/NCBI View Article : Google Scholar
|
|
40
|
Tarantini S, Valcarcel-Ares MA, Toth P,
Yabluchanskiy A, Tucsek Z, Kiss T, Hertelendy P, Kinter M, Ballabh
P, Süle Z, et al: Nicotinamide mononucleotide (NMN) supplementation
rescues cerebromicrovascular endothelial function and neurovascular
coupling responses and improves cognitive function in aged mice.
Redox Biol. 24(101192)2019.PubMed/NCBI View Article : Google Scholar
|
|
41
|
Wang X, Hu X, Zhang L, Xu X and Sakurai T:
Nicotinamide mononucleotide administration after sever hypoglycemia
improves neuronal survival and cognitive function in rats. Brain
Res Bull. 160:98–106. 2020.PubMed/NCBI View Article : Google Scholar
|
|
42
|
Aman Y, Qiu Y, Tao J and Fang EF:
Therapeutic potential of boosting NAD+ in aging and
age-related diseases. Transl Med Aging. 2018:30–37. 2018.PubMed/NCBI View Article : Google Scholar
|
|
43
|
Saklayen MG: The global epidemic of the
metabolic syndrome. Curr Hypertens Rep. 20(12)2018.PubMed/NCBI View Article : Google Scholar
|
|
44
|
O'Neill S and O'Driscoll L: Metabolic
syndrome: A closer look at the growing epidemic and its associated
pathologies. Obes Rev. 16:1–12. 2015.PubMed/NCBI View Article : Google Scholar
|
|
45
|
Canto C, Houtkooper RH, Pirinen E, Youn
DY, Oosterveer MH, Cen Y, Fernandez-Marcos PJ, Yamamoto H, Andreux
PA, Cettour-Rose P, et al: The NAD(+) precursor nicotinamide
riboside enhances oxidative metabolism and protects against
high-fat diet-induced obesity. Cell Metab. 15:838–847.
2012.PubMed/NCBI View Article : Google Scholar
|
|
46
|
Shima K, Zhu M and Kuwajima M: A role of
nicotinamide-induced increase in pancreatic beta-cell mass on blood
glucose control after discontinuation of the treatment in partially
pancreatectomized OLETF rats. Diabetes Res Clin Pract. 41:1–8.
1998.PubMed/NCBI View Article : Google Scholar
|
|
47
|
Park SY, Lee KB, Lee MJ, Bae SC and Jang
JJ: Nicotinamide inhibits the early stage of carcinogen-induced
hepatocarcinogenesis in mice and suppresses human hepatocellular
carcinoma cell growth. J Cell Physiol. 227:899–908. 2012.PubMed/NCBI View Article : Google Scholar
|
|
48
|
Hong SM, Hwang SW, Wang T, Park CW, Ryu
YM, Jung JH, Shin JH, Kim SY, Lee JL, Kim CW, et al: Increased
nicotinamide adenine dinucleotide pool promotes colon cancer
progression by suppressing reactive oxygen species level. Cancer
Sci. 110:629–638. 2019.PubMed/NCBI View Article : Google Scholar
|
|
49
|
Galbraith AR, Seabloom DE, Wuertz BR,
Antonides JD, Steele VE, Wattenberg LW and Ondrey FG:
Chemoprevention of lung carcinogenesis by dietary nicotinamide and
inhaled budesonide. Cancer Prev Res. 12:69–78. 2019.PubMed/NCBI View Article : Google Scholar
|
|
50
|
Audrito V, Managò A, Gaudino F, Sorci L,
Messana VG, Raffaelli N and Deaglio S: NAD-biosynthetic and
consuming enzymes as central players of metabolic regulation of
innate and adaptive immune responses in cancer. Front Immunol.
10(1720)2019.PubMed/NCBI View Article : Google Scholar
|
|
51
|
Yamamoto M, Inohara H and Nakagawa T:
Targeting metabolic pathways for head and neck cancers
therapeutics. Cancer Metastasis Rev. 36:503–514. 2017.PubMed/NCBI View Article : Google Scholar
|
|
52
|
Yaku K, Okabe K, Hikosaka K and Nakagawa
T: NAD metabolism in cancer therapeutics. Front Oncol.
8(622)2018.PubMed/NCBI View Article : Google Scholar
|
|
53
|
Li X, Lei J, Mao L, Wang Q, Xu F, Ran T,
Zhou Z and He S: NAMPT and NAPRT, key enzymes in NAD salvage
synthesis pathway, are of negative prognostic value in colorectal
cancer. Front Oncol. 9(736)2019.PubMed/NCBI View Article : Google Scholar
|
|
54
|
Lin JB and Apte RS: NAD+ and
sirtuins in retinal degenerative diseases: A look at future
therapies. Prog Retin Eye Res. 67:118–129. 2018.PubMed/NCBI View Article : Google Scholar
|
|
55
|
Jurja S, Hincu M, Dobrescu MA, Golu AE,
Balasoiu AT and Coman M: Ocular cells and light: Harmony or
conflict? Rom J Morphol Embryol. 55:257–261. 2014.PubMed/NCBI
|
|
56
|
Jurja S, Coman M and Hincu M: The
ultraviolet influence upon soft eye tissues. Rom J Morphol Embryol.
58:45–52. 2017.PubMed/NCBI
|
|
57
|
Koenekoop RK, Wang H, Majewski J, Wang X,
Lopez I, Ren H, Chen Y, Li Y, Fishman GA, Genead M, et al:
Mutations in NMNAT1 cause Leber congenital amaurosis and identify a
new disease pathway for retinal degeneration. Nat Genet.
44:1035–1039. 2012.PubMed/NCBI View Article : Google Scholar
|
|
58
|
Greenwald SH, Charette JR, Staniszewska M,
Shi LY, Brown SD, Stone L, Liu Q, Hicks WL, Collin GB, Bowl MR, et
al: Mouse models of NMNAT1-Leber congenital Amaurosis (LCA9)
recapitulate key features of the human disease. Am J Pathol.
186:1925–1938. 2016.PubMed/NCBI View Article : Google Scholar
|
|
59
|
Kaja S, Shah A, Haji S, Patel KB, Naumchuk
Y, Zabaneh A, Gerdes BC, Kunjukunju N, Sabates NR, Cassell MA, et
al: Nampt/PBEF/visfatin serum levels: A new biomarker for retinal
blood vessel occlusions. Clin Ophthalmol. 9:611–618.
2015.PubMed/NCBI View Article : Google Scholar
|
|
60
|
Zabka TS, Singh J, Dhawan P, Liederer BM,
Oeh J, Kauss MA, Xiao Y, Zak M, Lin T, McCray B, et al: Retinal
toxicity, in vivo and in vitro, associated with inhibition of
nicotinamide phosphoribosyltransferase. Toxicol Sci. 144:163–172.
2015.PubMed/NCBI View Article : Google Scholar
|
|
61
|
Jadeja RN, Powell FL and Jones MA: Loss of
NAMPT in aging retinal pigment epithelium reduces NAD+
availability and promotes cellular senescence. Aging. 10:1306–1323.
2018.PubMed/NCBI View Article : Google Scholar
|
|
62
|
Jadeja RN, Thounaojam MC, Bartoli M and
Martin PM: Implications of NAD+ metabolism in the aging
retina and retinal degeneration. Oxid Med Cell Longev.
2020(2692794)2020.PubMed/NCBI View Article : Google Scholar
|
|
63
|
Balaiya S, Abu-Amero KK, Kondkar AA and
Chalam KV: Sirtuins expression and their role in retinal diseases.
Oxid Med Cell Longev. 2017(3187594)2017.PubMed/NCBI View Article : Google Scholar
|
|
64
|
Ban N, Ozawa Y, Inaba T, Miyake S,
Watanabe M, Shinmura K and Tsubota K: Light-dark condition
regulates sirtuin mRNA levels in the retina. Experimental Gerontol.
48:1212–1217. 2013.PubMed/NCBI View Article : Google Scholar
|
|
65
|
Zhang X, Henneman NF, Girardot PE, Sellers
JT, Chrenek MA, Li Y, Wang J, Brenner C, Nickerson JM and Boatright
JH: Systemic treatment with nicotinamide riboside is protective in
a mouse model of light-induced retinal degeneration. Invest
Ophthalmol Vis Sci. 61(47)2020.PubMed/NCBI View Article : Google Scholar
|
|
66
|
Lin JB, Kubota S, Mostoslavsky R and Apte
RS: Role of sirtuins in retinal function under basal conditions.
Adv Exp Med Biol. 1074:561–567. 2018.PubMed/NCBI View Article : Google Scholar
|