Introduction
Ossification of the posterior longitudinal ligament
(OPLL) is a common musculoskeletal disease, characterized by
progressive ectopic ossification in the tissues of the spinal
ligament and leading to hyperostotic conditions (1-3).
Patients with OPLL usually develop various degrees of neurological
symptoms, from discomfort to severe myelopathy, due to compression
of the spinal cord and nerve roots by the progressively calcified
posterior longitudinal ligament (PLL). The development of OPLL
depends on multiple factors, including diet, obesity, age, diabetes
mellitus, and genetic factors (4);
however, the role of fibroblastic cells, the most common cells in
connective tissues, including PLL (5), is largely unknown.
Recently, Wang et al (6) analyzed the long non-coding RNAs
(lncRNA) in OPLL tissues and identified 143 differentially
expressed lncRNAs compared with tissues from healthy controls,
which indicated an extensive molecular change in the development of
OPLL. The silencing of lncRNA SNHG1, which is highly expressed in
OPLL tissues, significantly inhibited the osteogenic
differentiation of ligament fibroblastic cells (6). Dickkopf-1 (Dkk1) is a secreted
inhibitor of the Wnt pathway, and its expression level was
significantly decreased in OPLL patients and ligament cells
compared with that in non-OPLL controls (7). Treatment with recombinant Dkk1
markedly reversed the osteogenic differentiation of OPLL ligament
cells via inhibition of the Wnt pathway (7). In addition, microRNA (miRNA/miR)-563
levels were found to be significantly higher in OPLL ligament
cells, and the silencing of miR-563 considerably inhibited the
osteogenic differentiation of OPLL ligament cells (8). These findings indicate that
fibroblastic cells from PLL are implicated in the progression of
ectopic ossification. Therefore, understanding the underlying
molecular mechanism of the osteogenic differentiation of ligament
cells is expected to provide a promising therapeutic target for
OPLL treatment.
Circular RNAs (circRNAs) are a large endogenous
group of non-coding RNAs that form a closed loop structure due to a
covalent phosphodiester bond between the 3' and 5' ends (9,10).
CircRNAs are thought to have a significant role in bone metabolism
(11). A previous study has shown
that circ19142 and circ5846 are associated with osteoblast
differentiation, and that bone morphogenetic protein (BMP) 2 might
induce osteogenic differentiation through the
circ19142/circ5846-targeted miRNA-mRNA axis (12). CircRNAs hsa_circ_0032462,
hsa_circ_0028173, and hsa_circ_0005909 are known to promote cell
adhesion molecule 1 expression by functioning as miRNA sponges in
human osteosarcoma (13).
Additionally, the silencing of circIGSF11 promoted osteoblast
differentiation and increased the expression of miR-199b-5p in bone
marrow stem cells (14). To date,
whether circRNA is implicated in the osteoblast differentiation of
ligament cells remains unknown.
In the present study, control and OPLL patient
ligament samples were collected and the expression levels of
circSKIL, which was upregulated in osteogenically differentiated
periodontal ligament stem cells (15), were assessed. The role of circSKIL
in the proliferation and osteogenic differentiation of ligament
cells was analyzed, and the JNK/STAT3 pathway was also
examined.
Materials and methods
Sample collection
Cervical posterior longitudinal ligament tissue
samples were collected from 24 patients diagnosed with cervical
spondylotic myelopathy (CSM) without OPLL (control group) and from
another 24 CSM patients with OPLL (OPLL group) between June 2017
and October 2019. The clinicopathological characteristics of the
patients are presented in Table I.
The diagnosis of CSM and OPLL was based on the clinical data
obtained from computer tomography (CT) and magnetic resonance
imaging (MRI). Patients with diffuse idiopathic skeletal
hyperostosis or diabetes mellitus were excluded. The modified
Japanese Orthopedic Association (mJOA) guidelines were used to
evaluate the severity of myelopathy (16), and only patients with moderate
symptoms (mJOA score, 12-14) were considered. The OPLL type was
diagnosed as segmental, according to a widely used OPLL
classification (17). All patients
underwent cervical anterior decompression surgery, and the
posterior longitudinal ligaments were collected intraoperatively.
The current study was approved by the Ethical Committee of the
First Affiliated Hospital, Xiamen University (Xiamen, China). The
study protocols were in accordance with The Declaration of
Helsinki. Informed consent was obtained from all patients prior to
sample collection for the study.
 | Table IClinicopathological characteristics
of control and OPLL patients. |
Table I
Clinicopathological characteristics
of control and OPLL patients.
| Feature | Control | OPLL |
|---|
| Sex | | |
|
Male | 14 | 10 |
|
Female | 9 | 13 |
| Age | 49.22±6.43 | 52.37±5.28 |
| Diagnosis | CSM | CSM and OPLL |
| OPLL type | NA | segmental |
| mJOA score | 12.55±0.74 | 12.75±0.83 |
Cell culturing of OPLL cells and
osteogenic induction
For primary cell isolation, three control ligament
tissues and three OPLL tissues from the aforementioned 46 patients
were collected and minced to an approximate size of 0.5
mm3. The fragments were washed twice with PBS and plated
on a 90-mm culture dish with low-glucose DMEM (Thermo Fisher
Scientific, Inc.) supplemented with 10% fetal bovine serum (FBS;
Gibco; Thermo Fisher Scientific, Inc.), 1% L-glutamine (Gibco;
Thermo Fisher Scientific, Inc.) and 1% penicillin/streptomycin
(Gibco; Thermo Fisher Scientific, Inc.). The medium was changed
every three days. Passage number 3 cells were used for the
experimental studies. For osteogenic differentiation, cells were
cultured in differentiation medium, which is an osteogenic
inductive DMEM containing various supplements (10% FBS, 50 µg/ml
ascorbate-2 phosphate, 100 nM dexamethasone, and 10 mM
β-glycerophosphate; Sigma-Aldrich; Merck KGaA). Following 7 or 14
days of osteogenic induction, the cells were used for subsequent
assays. All cells were incubated in a humidified atmosphere
containing 95% air and 5% CO2 at 37˚C.
Alizarin Red S (ARS) staining and
quantification
To visualize the mineralization potential of
ligament cells, ARS staining was performed, as described previously
(18). ARS selectively binds to
calcium, yielding a dark red color. Before ARS staining, OPLL cells
(1x105 cells/well) were cultured in a 12-well plate with
osteogenic induction medium for 7 days, as aforementioned. The
cultured cells were fixed with 95% ethanol for 30 min at room
temperature, and then stained with 0.1% ARS (Sciencell Research
Laboratories, Inc.) for 20 min at room temperature. Images of the
calcified matrices were photographed using a conventional
camera.
An ARS Staining Quantification Assay kit (Sciencell
Research Laboratories, Inc.) was used for the quantification of
matrix mineralization. Basically, the stain was eluted using 100 mM
cetylpyridinium chloride for 1 h and spectrophotometric
quantification was performed at 562 nm.
Cell proliferation assay
The cell proliferation rate was determined using a
CCK-8 assay. Cells were cultured in 96-well plates
(3.5x103 cells/well) for 24 h at 37˚C. The cells in
various wells overexpressed either circSKIL or the mock plasmid for
the control cells group, and either the small interfering (si-)
circSKIL or negative control siRNA (si-NC) for the OPLL cells
group. Cell proliferation was assessed every day, from the 1st to
the 5th day. After each incubation time, 10 µl of CCK-8 reagent
(Beyotime Institute of Biotechnology) was added to each well and
cultured for 1 h at 37˚C. The absorbance was measured at 450 nm
using a microplate reader. All tests were performed in
triplicates.
RNA isolation and reverse
transcription-quantitative PCR (RT-qPCR)
Total RNA extraction was performed using
TRIzol® reagent (Thermo Fisher Scientific, Inc.),
according to the manufacturer's instructions. In total, 1 µg RNA
was reverse transcribed into cDNA using the iScriptcDNA synthesis
kit (Bio-Rad Laboratories, Inc.). The reaction was started at 25˚C
for 5 min, followed by 42˚C for 60 min and 95˚C for 5 min. Random
primers (Toyobo Life Science) were used for circRNA analysis.
ABsolute qPCR SYBR Green Master Mix (Thermo Fisher Scientific,
Inc.) was used for qPCR analysis. The total reaction volume (20 µl)
of buffered solution contained 1 µl of the diluted (1:5) reverse
transcription product and 10 pM sense and antisense primers.
Thermocycling was performed as follows: 95˚C for 5 min, then 42
cycles of 95˚C for 5 sec and 60˚C for 1 min. GAPDH was used as the
housekeeping gene to normalize the expression data. To calculate
the fold change in each experiment, the 2-ΔΔCq method
was used (19). The primers used in
the quantitative PCR analysis are listed in Table II. The gene transcription levels of
different osteogenic differentiation markers, namely Runt-related
transcription factor 2 (RUNX2), alkaline phosphatase (ALP),
collagen I (COL I), osteocalcin (OCN), and osteopontin (OPN), were
assessed using different primers. The transcription levels of
circSKIL were assessed in control cells in which circSKIL was
overexpressed and in OPLL cells in which circSKIL expression was
downregulated.
| Table IISequences of primers used for reverse
transcription-quantitative PCR. |
Table II
Sequences of primers used for reverse
transcription-quantitative PCR.
| Gene | Forward primer
(‘5-3’) | Reverse primer
(‘5-3’) |
|---|
| circSKIL |
AGGAGAAGTTTAGCATGAGAAGTG |
AATTGCCCAGTTATCTTCAAGGATT |
| RUNX2 |
GGGAACCAAGAAGGCACAGA |
GGATGAGGAATGCGCCCTAA |
| ALP |
ATCGACGTGATCATGGGTGG |
TGGGAATGCTTGTGTCTGGG |
| COL I |
GGACACAATGGATTGCAAGGCCGC |
TAACCACTGCTCCACTCTGGATGG |
| OCN |
ACCTCACAGATGCCAAGCC |
GCCGGAGTCTGTTCACTACC |
| OPN |
CTCCATTGACTCGAACGACTC |
CAGGTCTGCGAAACTTCTTAGAT |
| GAPDH |
GTCTCCTCTGACTTCAACAGCG |
ACCACCCTGTTGCTGTAGCCAA |
Vector construction and cell
transfection
To overexpress circSKIL in control cells, the
full-length cDNA of circSKIL was amplified and cloned into the
pcircRNA 1.2 vector (Guangzhou Boxin Biotechnology Co., Ltd.). The
empty plasmid pcircRNA 1.2 without the circSKIL cDNA served as a
control vector. To knockdown circSKIL in OPLL cells, siRNAs
targeting the back-splice junction of circSKIL (si-circSKIL,
5'-AUCCAAGAUUAAUUAAAAGAA-3') and its negative control (si-NC,
5'-UUCUCCGAACGUGUCACGUTT-3') were designed and synthesized
(Guangzhou RiboBio Co., Ltd.).
Cells were seeded in 6-well plates, and 2 µg/ml of
plasmids or 50 nM siRNAs were transfected into the corresponding
cells using Lipofectamine 2000 (Thermo Fisher Scientific, Inc.) at
37˚C for 48 h. For the circSKIL overexpression, cells
overexpressing plasmids were selected using 300 µg/ml neomycin for
10 days at 37˚C and confirmed by RT-qPCR, then used for subsequent
experiments. For the circSKIL knockdown, expression levels in cells
transfected with siRNAs were confirmed by RT-qPCR, and the cells
were then used for subsequent experiments.
Western blot analysis
The total protein from the experimental cells was
obtained using RIPA lysis buffer (Beyotime Institute of
Biotechnology). The protein concentration was determined using a
bicinchoninic acid protein assay kit (Biosharp Life Sciences),
using bovine serum albumin as a standard. Equal amounts of total
protein (20 µg) were separated using 12% sodium dodecyl sulfate
polyacrylamide gel electrophoresis (SDS-PAGE). Proteins were
transferred onto PVDF membranes (EMD Millipore). After blocking the
membranes with 5% non-fat milk for 1 h at room temperature, they
were incubated with primary antibodies against collagen type I α 1
chain (COL1A1; cat. no. 39952; 1:1,000; Cell Signaling Technology,
Inc.), RUNX2 (cat. no. 12556; 1:1,000; Cell Signaling Technology,
Inc.), OCN (cat. no. A18241; 1:1,000; ABclonal Biotech Co., Ltd.),
OPN (cat. no. A1361; 1:1,000; ABclonal Biotech Co., Ltd.), ALP
(cat. no. A0514; 1:1,000; ABclonal Biotech Co., Ltd.),
phosphorylated (p-) JNK (cat. no. 4668; 1:1,000; Cell Signaling
Technology, Inc.), JNK (cat. no. 9252; 1:1,000; Cell Signaling
Technology, Inc.), p-STAT3 (cat. no. 9145; 1:2,000; Cell Signaling
Technology, Inc.), STAT3 (cat. no. 4904; 1:2,000; Cell Signaling
Technology, Inc.), and GAPDH (cat. no. 2118; 1:1,000; Cell
Signaling Technology, Inc.) at 4˚C overnight. GAPDH was used as the
loading control. After washing with TBST (1% Tween) twice, the
membranes were incubated for 2 h with HRP-conjugated secondary
antibodies (cat. no. 7074; 1:2,000; Cell Signaling Technology,
Inc.) at room temperature. The protein signals were detected and
visualized using an ECL detection kit (Beyotime Institute of
Biotechnology). Densitometry quantification analysis was performed
using ImageJ v1.8.0 software (National Institutes of Health).
Statistical Analysis
Quantitative data were expressed as mean ± standard
deviation from at least three independent experiments. All analyses
were performed using SPSS 16.0 (SPSS, Inc.). Student's t-test was
used to evaluate the statistical significance of differences
between two groups. P<0.05 was considered to indicate a
statistically significant difference.
Results
circSKIL is upregulated in OPLL cells
and tissues
Primary cervical posterior longitudinal ligament
cells were isolated from three patients with CSM without
ossification (control) and another three patients with CSM with
significant ossification (OPLL), diagnosed according to their MRI
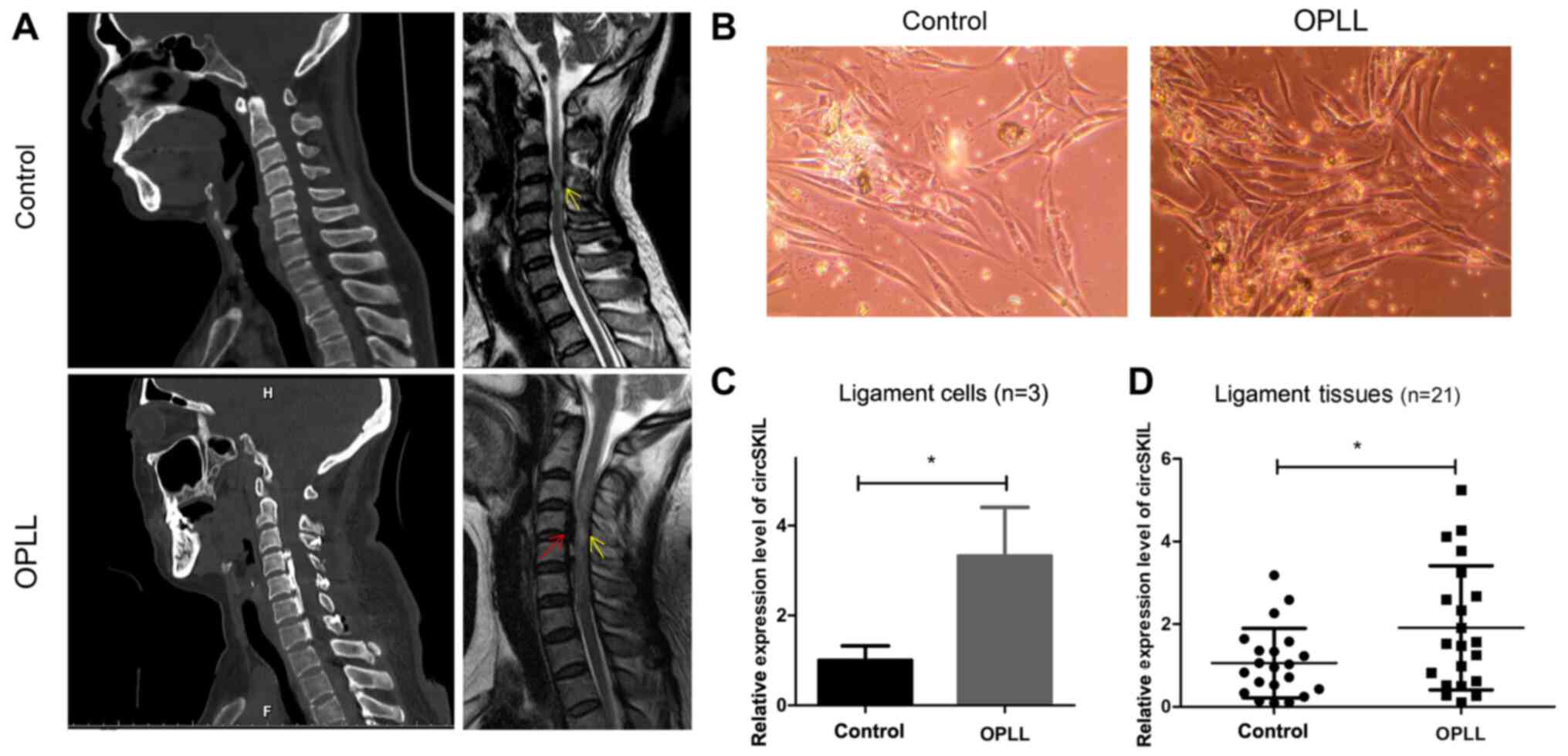
and CT data (Fig. 1A).
Representative phase-contrast microscopy images from the two groups
are presented in Fig. 1B, showing
that the cells had a fibroblast-like morphology. The expression
levels of circSKIL were detected, and the results indicated that
its relative expression levels were significantly higher in the
OPLL group compared with the control group (Fig. 1C). For further confirmation,
circSKIL expression levels were also detected in 21 tissue samples
from patients with OPLL and 21 samples from the control patients,
and the results were consistent; circSKIL expression levels were
significantly elevated in OPLL tissues compared with control
tissues (Fig. 1D).
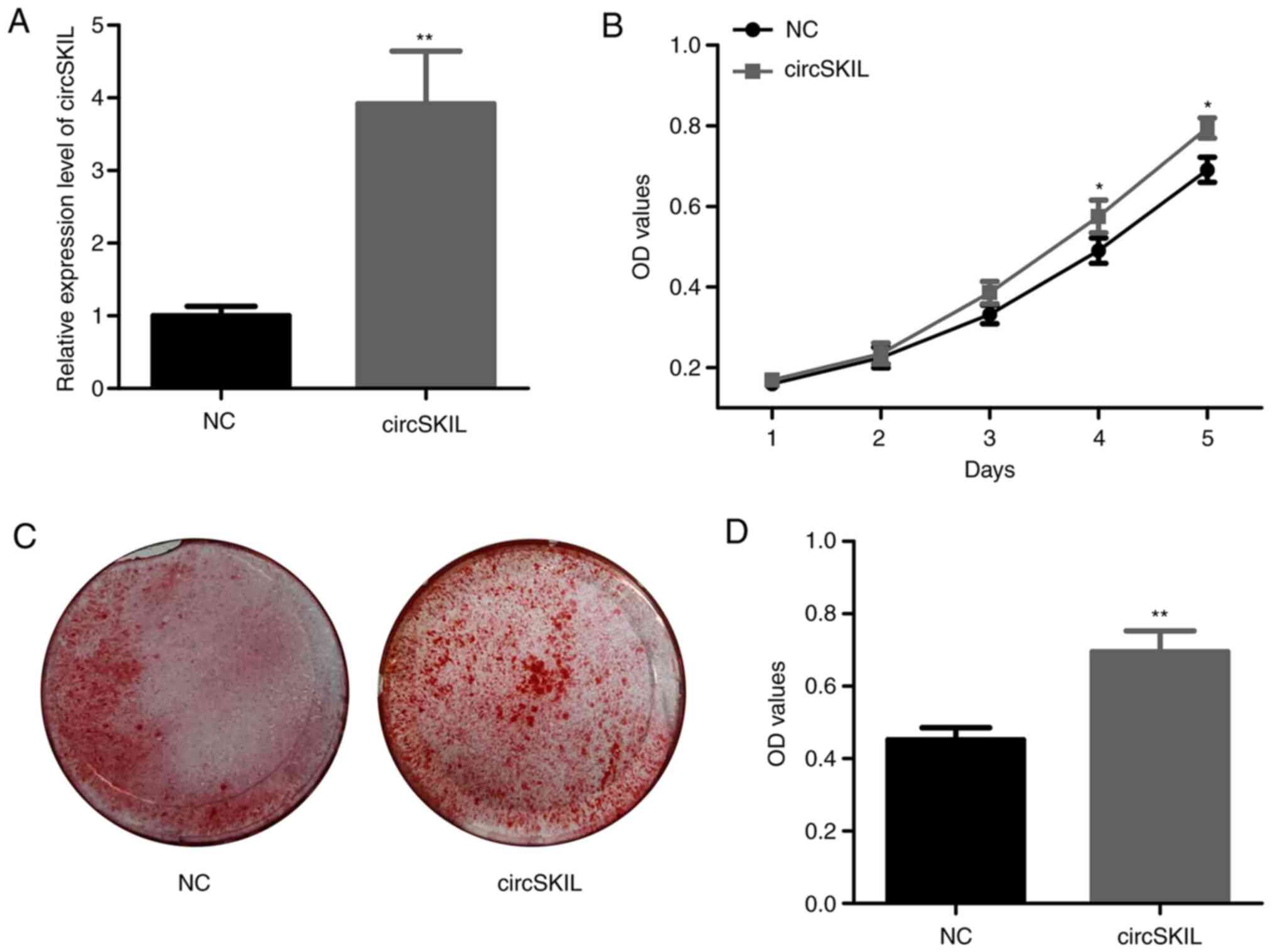
circSKIL overexpression accelerates
the proliferation and mineralization of control ligament cells
Next, the function of circSKIL was investigated in
control cells by overexpressing circSKIL. Following transfection
with an overexpressing plasmid, the levels of circSKIL were
significantly higher compared with cells transfected with an empty
control vector (NC group; Fig. 2A).
The proliferation assay demonstrated a significant increase in the
proliferation of circSKIL-overexpressing cells on the 4th and 5th
days, compared with that of the NC cells (Fig. 2B). In addition, mineralization was
significantly higher in the circSKIL-overexpressing cells compared
with that of NC cells, based on ARS staining (Fig. 2C and D).
circSKIL overexpression promotes
osteogenic differentiation and activates the JNK/STAT3 pathway in
control ligament cells
For further confirmation of osteogenic
differentiation, the expression of specific markers was evaluated
in control ligament cells overexpressing circSKIL. circSKIL
overexpression significantly increased the mRNA expression levels
of various osteogenic markers, including RUNX2, ALP, COL I, OCN,
and OPN compared with the empty vector-transfected cells (Fig. 3A). Western blot analysis of the
circSKIL-overexpressing cells indicated that the protein levels of
osteogenic markers were all increased (Fig. 3B), which was consistent with the
RT-qPCR results. The JNK/STAT3 pathway was also evaluated, and the
results revealed that phosphorylation of both JNK and STAT3 was
significantly induced following circSKIL overexpression (Fig. 3C).
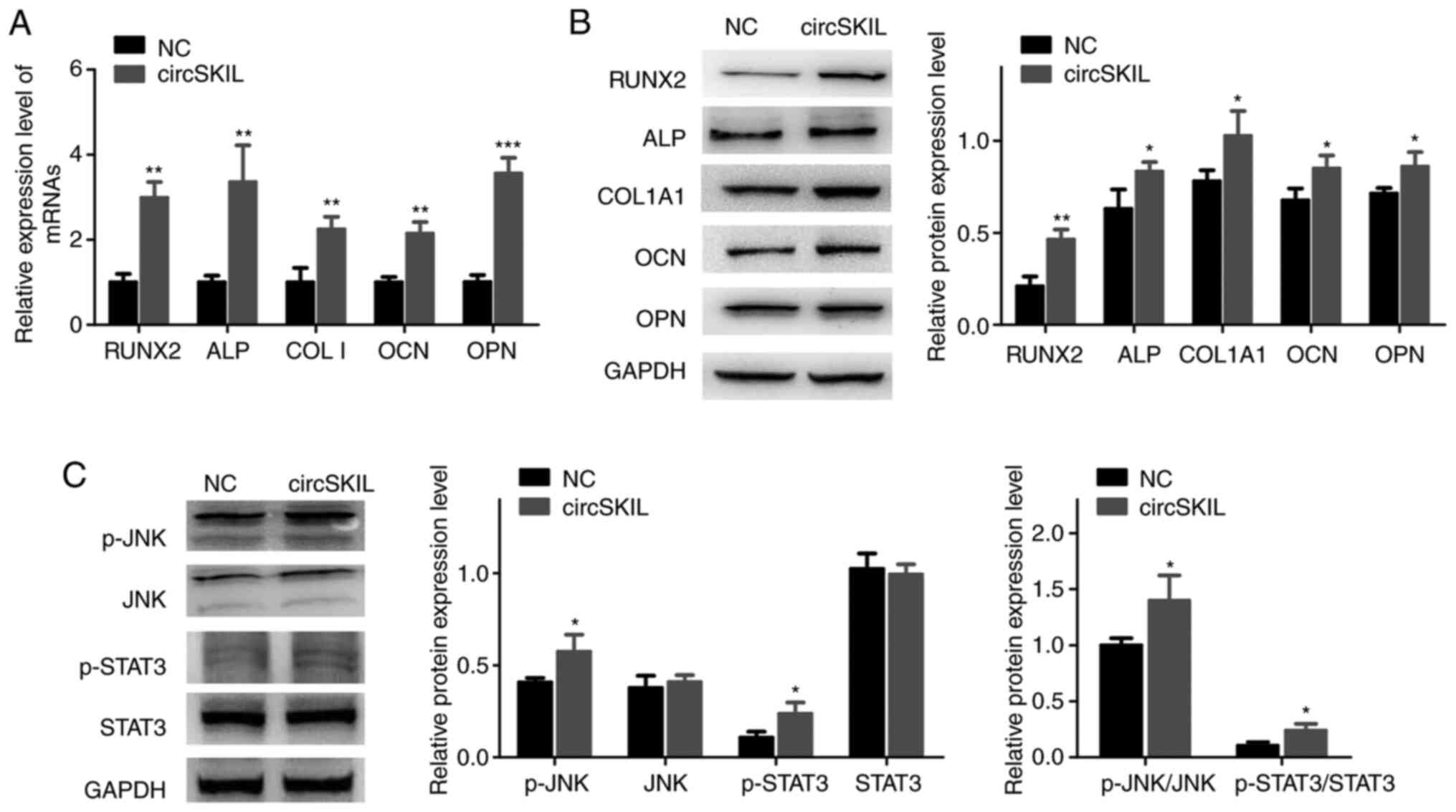 | Figure 3Overexpression of circSKIL promotes
the osteogenic differentiation and activates the JNK/STAT3 pathway
in control cells. Control cells were transfected with either a
circSKIL-overexpressing plasmid or an empty vector (NC). (A) mRNA
expression levels of RUNX2, ALP, COL I, OCN, and OPN (n=3). (B)
Western blot analysis of the protein expression levels of RUNX2,
ALP, COL1A1, OCN, and OPN (n=3). (C) Western blot analysis of the
protein levels of p-JNK, JNK, p-STAT3, and STAT3 (n=3).
*P<0.05, **P<0.01 and
***P<0.001 vs. NC. circ, circular RNA; NC, negative
control; RUNX2, runt-related transcription factor 2; ALP, alkaline
phosphatase; COL I, collagen I; OCN, osteocalcin; OPN, osteopontin;
COL1A1, collagen type I α 1 chain; p-, phosphorylated. |
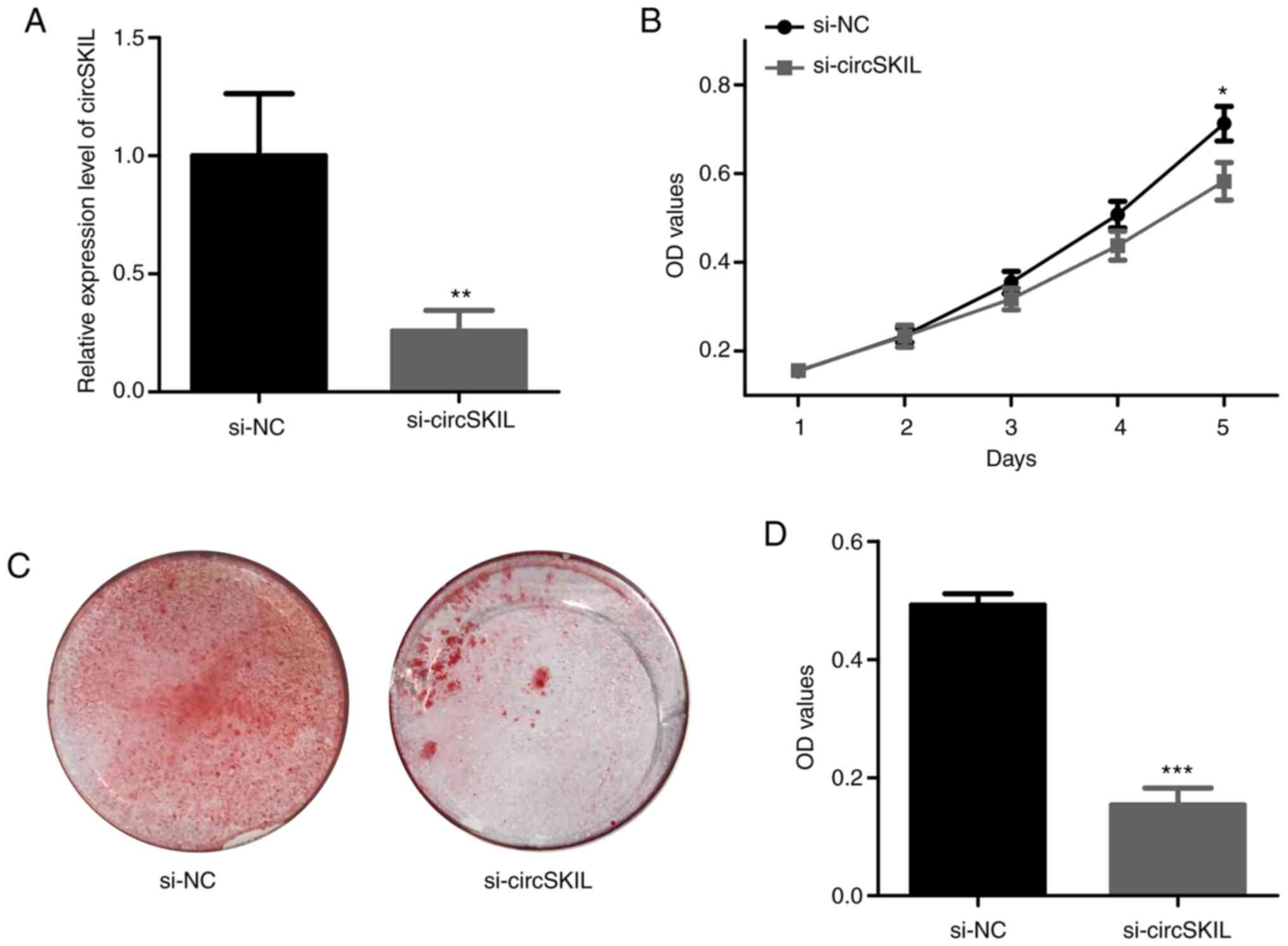
circSKIL downregulation inhibits the
proliferation and mineralization of OPLL cells
To further investigate the role of circSKIL, its
expression was silenced by siRNA in OPLL cells. Following siRNA
silencing, the levels of circSKIL were significantly reduced in
si-circSKIL cells compared to those in si-NC cells (Fig. 4A). The proliferation assay indicated
a significant decrease in proliferation after circSKIL was
downregulated in OPLL cells (Fig.
4B). In addition, mineralization in the circSKIL-silenced cells
was significantly abrogated compared with that in si-NC cells,
based on ARS staining (Fig. 4C and
D).
circSKIL downregulation suppresses
osteogenic differentiation and inhibits the JNK/STAT3 pathway in
OPLL cells
RT-qPCR and western blot results demonstrated that
the mRNA and proteins expression levels, respectively, of the
different osteogenic markers were significantly reduced in OPLL
cells silenced with si-circSKIL compared with si-NC-transfected
cells (Fig. 5A and B). In addition, the JNK/STAT3 pathway was
significantly inhibited in circSKIL-silenced cells, as evident by a
significant decrease in both JNK and STAT3 phosphorylation
(Fig. 5C).
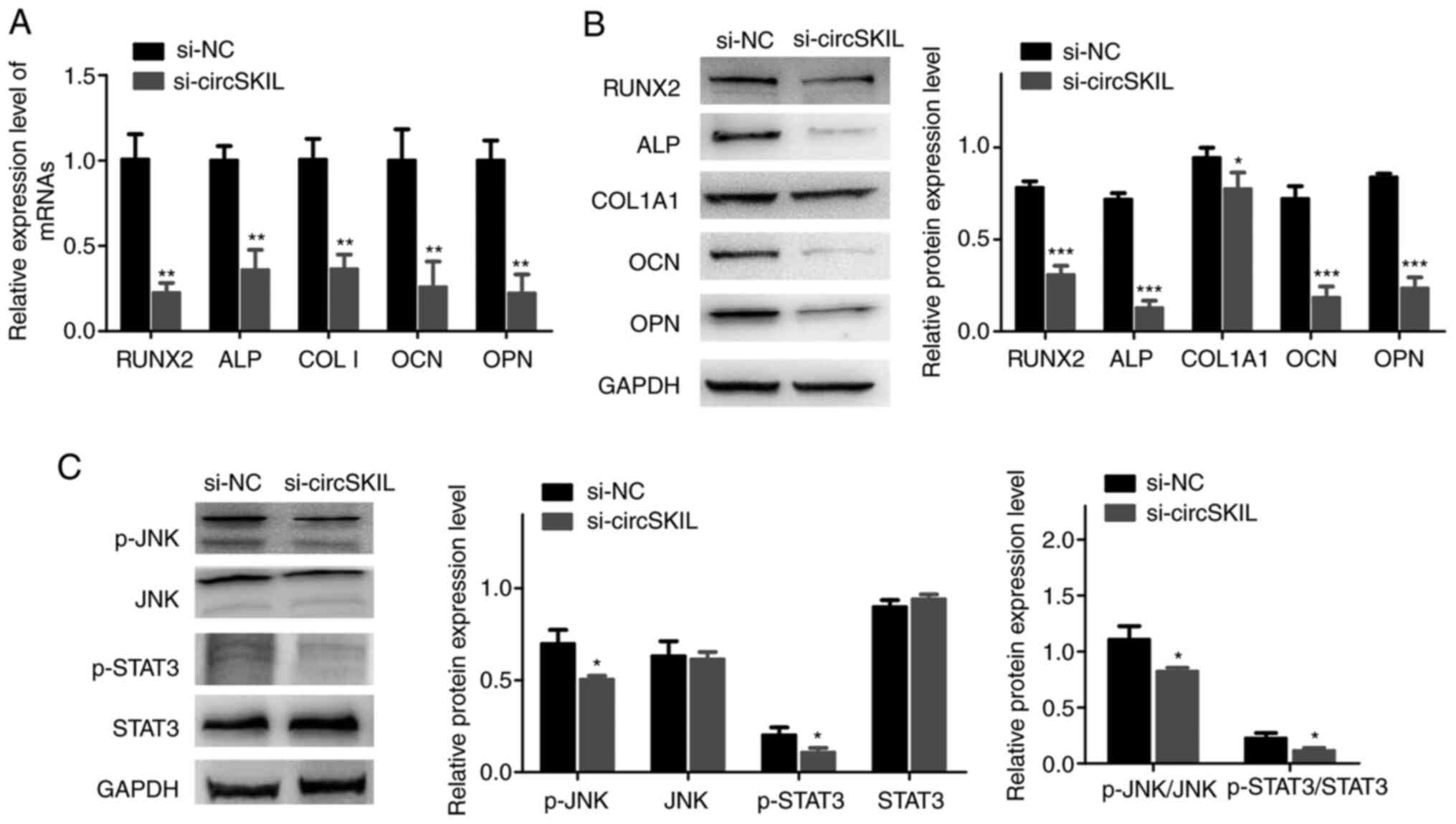 | Figure 5Downregulation of circSKIL suppresses
osteogenic differentiation and inhibits the JNK/STAT3 pathway in
OPLL cells. OPLL cells were transfected with either a
circSKIL-targeting siRNA or a negative control siRNA (si-NC). (A)
mRNA expression levels of RUNX2, ALP, COL I, OCN, and OPN (n=3).
(B) Western blot analysis of the protein expression levels of
RUNX2, ALP, COL1A1, OCN, and OPN (n=3). (C) Western blot analysis
of the protein levels of p-JNK, JNK, p-STAT3, and STAT3 (n=3).
*P<0.05, **P<0.01 and
***P<0.001 vs. si-NC. circ, circular RNA; OPLL,
ossification of the posterior longitudinal ligament; si, small
interfering; NC, negative control; RUNX2, runt-related
transcription factor 2; ALP, alkaline phosphatase; COL I, collagen
I; OCN, osteocalcin; OPN, osteopontin; COL1A1, collagen type I α 1
chain; p-, phosphorylated. |
Discussion
OPLL is a challenging spinal disease, with its high
incidence affecting the normal life of patients. Anterior and
posterior approaches are currently the two major OPLL therapies,
but their outcomes remain not satisfactory (20). Multiple factors, such as diet,
obesity, age, and diabetes mellitus, are reported to be involved in
the progression of OPLL but the underlying molecular mechanism of
OPLL is largely unknown.
Emerging evidence has revealed that non-coding RNAs,
including lncRNAs and microRNAs (miRNAs), can regulate osteogenic
differentiation of ligament cells (6,8);
however, the role of circRNA, a rising star in non-coding RNAs, in
OPLL progression remains unknown. CircRNA SKIL was first identified
as upregulated in osteogenically differentiated periodontal
ligament stem cells compared with undifferentiated cells using
RNA-seq analysis, and its upregulation was confirmed using RT-qPCR
(15). The presents study detected
its levels in OPLL tissues and cells to explore the expression
differences between OPLL and control samples. The present RT-qPCR
analyses demonstrated that circSKIL expression levels were
significantly increased in OPLL ligament cells and tissues compared
with control cells and tissues, respectively, which suggested that
an upregulation of circSKIL might play a role in the osteogenic
differentiation of ligament cells.
RUNX2 is a key transcription factor that has a
crucial role in the early stage of osteoblastic differentiation
(21) and is also associated with
OPLL (22). ALP is an
osteoblast-specific phenotypic marker that is attached to membrane
phospholipids of the matrix vesicles and whose activity is
increased during the matrix maturation phase (23). COL I is involved in the formation of
bone matrix and it is synthesized and secreted by osteoblasts
(24). OCN is the most abundant
non-collagenous protein in bone matrix and has a crucial role in
the process of biomineralization during osteogenic maturation
(25). Alterations in the
expression levels of these markers are commonly used to identify
the osteogenic differentiation level of ligament cells, as
previously reported (6,7,26). In
the present study, circSKIL was overexpressed in control ligament
cells to investigate its role in osteogenic differentiation. The
results demonstrated that overexpression of circSKIL significantly
induced the mineralization process and increased the expression
levels of RUNX2, ALP, COL I, OCN, and OPN. By contrast, the
downregulation of circSKIL significantly inhibited the levels of
the five osteogenic differentiation markers and the mineralization
of OPLL ligament cells.
STAT3 belongs to the signal transducer and activator
of transcription family, and the activated STAT3 forms dimers and
translocates to the nucleus to regulate the transcription of
various genes involved in cellular proliferation and
differentiation (27), as well as
in osteogenic differentiation (28). The inhibition of STAT3 suppressed
osteogenic differentiation-related gene expression in BMSCs induced
by hypoxia (29). JNK is an
important subfamily of MAPKs, and the activation of JNK has been
reported to enhance the osteogenic differentiation of mesodermal
progenitor cells (30) and
periodontal ligament stem cells (PDLSCs) (31). Hence, the phosphorylation levels of
JNK and STAT3 were detected in ligament cells in the present study.
The current results indicated that overexpression of circSKIL
upregulated p-JNK and p-STAT3 in control ligament cells, whereas
silencing of circSKIL inhibited the phosphorylation of the
JNK/STAT3 pathway in OPLL cells. Therefore, the role of circSKIL in
the osteogenic differentiation of ligament cells may occur via the
regulation of the JNK/STAT3 pathway. Inhibition of this pathway may
serve as a strategy to alleviate the osteogenic differentiation of
ligament cells.
CircRNAs can act as miRNA sponges and regulate the
expression of target mRNAs as RNA-induced silencing complexes
(10). For example, a circRNA named
cerebellar degeneration-related protein 1 transcript (CDR1)
promoted the osteogenic differentiation of PDLSCs by acting as an
miR-7 inhibitor, which triggers growth differentiation factor 5
upregulation, followed by Smad1/5/8 and p38 MAPK phosphorylation
(32). CircPOMT1 and circMCM3AP are
known to influence the osteogenic differentiation of human
adipose-derived stem cells by targeting hsa-miR-6881-3p via the BMP
signaling pathway (33). In
addition, several miRNAs, such as miR-204 and miR-211, are able to
directly target RUNX2, suppressing its transcriptional activity,
thereby inhibiting osteogenic differentiation (34). To date, our knowledge of the role of
miRNAs in OPLL is relatively limited compared to other bone
disorders, but miRNAs, such as miR-10a (35), miR-1(36), and miR-17-5p (37), have been reported to inhibit target
genes involved in the osteogenesis of OPLL ligament cells.
Therefore, several miRNAs might be sponged via circSKIL, thus
regulating the osteogenic differentiation of ligament cells, a
hypothesis that requires further studies.
A limitation of the present study is that the
control samples were not collected from healthy subjects, but from
patients with CSM. To exclude the role of CSM on the OPLL mechanism
as much as possible, only patients with moderate symptoms were
considered for the present study, while those with severe symptoms
were excluded.
In conclusion, the present results demonstrated that
circSKIL was involved in osteogenic differentiation by regulating
the JNK/STAT3 pathway in OPLL ligament cells. The present study
provides a novel mechanism for the progression of OPLL.
Acknowledgements
Not applicable.
Funding
Funding: This work was supported by Xiamen Health and Family
Planning Commission (grant no. WKJ2016-2-20).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
GR designed the study. NS, YL and BH performed the
experiments and analyzed the data. JF, GL and XC collected and
assembled data. GR and NS confirmed the authenticity of the raw
data. All authors contributed to the preparation of manuscript and
approved the final version.
Ethics approval and consent to
participate
The present study was approved by the Ethics
Committee of the First Affiliated Hospital, Xiamen University
(Xiamen, China) and written informed consent was obtained from all
patients.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Ramos-Remus C, Russell AS, Gomez-Vargas A,
Hernandez-Chavez A, Maksymowych WP, Gamez-Nava JI, Gonzalez-Lopez
L, Garcia-Hernandez A, Meono-Morales E, Burgos-Vargas R and
Suarez-Almazor ME: Ossification of the posterior longitudinal
ligament in three geographically and genetically different
populations of ankylosing spondylitis and other
spondyloarthropathies. Ann Rheum Dis. 57:429–433. 1998.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Xu N, Yu M, Liu X, Sun C, Chen Z and Liu
Z: A systematic review of complications in thoracic spine surgery
for ossification of the posterior longitudinal ligament. Eur Spine
J. 26:1803–1809. 2017.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Guo Q, Ni B and Yang J, Zhu Z and Yang J:
Simultaneous ossification of the posterior longitudinal ligament
and ossification of the ligamentum flavum causing upper thoracic
myelopathy in DISH: Case report and literature review. Eur Spine J.
20 (Suppl 2):S195–S201. 2011.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Nam DC, Lee HJ, Lee CJ and Hwang SC:
Molecular pathophysiology of ossification of the posterior
longitudinal ligament (OPLL). Biomol Ther (Seoul). 27:342–348.
2019.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Yu F, Cui Y, Zhou X, Zhang X and Han J:
Osteogenic differentiation of human ligament fibroblasts induced by
conditioned medium of osteoclast-like cells. Biosci Trends.
5:46–51. 2011.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Wang Y, Niu H, Liu Y, Yang H, Zhang M and
Wang L: Promoting effect of long non-coding RNA SNHG1 on osteogenic
differentiation of fibroblastic cells from the posterior
longitudinal ligament by the microRNA-320b/IFNGR1 network. Cell
Cycle. 19:2836–2850. 2020.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Dong J, Xu X, Zhang Q, Yuan Z and Tan B:
Dkk1 acts as a negative regulator in the osteogenic differentiation
of the posterior longitudinal ligament cells. Cell Biol Int.
44:2450–2458. 2020.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Zhang H, Xu C, Liu Y and Yuan W:
MicroRNA-563 promotes the osteogenic differentiation of posterior
longitudinal ligament cells by inhibiting SMURF1. Zhonghua Wai Ke
Za Zhi. 55:203–207. 2017.PubMed/NCBI View Article : Google Scholar : (In Chinese).
|
|
9
|
Santosh B, Varshney A and Yadava PK:
Non-coding RNAs: Biological functions and applications. Cell
Biochem Funct. 33:14–22. 2015.PubMed/NCBI View
Article : Google Scholar
|
|
10
|
Xie L, Huang W, Fang Z, Ding F, Zou F, Ma
X, Tao J, Guo J, Xia X, Wang H, et al: CircERCC2 ameliorated
intervertebral disc degeneration by regulating mitophagy and
apoptosis through miR-182-5p/SIRT1 axis. Cell Death Dis.
10(751)2019.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Patil S, Dang K, Zhao X, Gao Y and Qian A:
Role of LncRNAs and CircRNAs in bone metabolism and osteoporosis.
Front Genet. 11(584118)2020.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Qian DY, Yan GB, Bai B, Chen Y, Zhang SJ,
Yao YC and Xia H: Differential circRNA expression profiles during
the BMP2-induced osteogenic differentiation of MC3T3-E1 cells.
Biomed Pharmacother. 90:492–499. 2017.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Chen G, Wang Q, Yang Q, Li Z, Du Z, Ren M,
Zhao H, Song Y and Zhang G: Circular RNAs hsa_circ_0032462,
hsa_circ_0028173, hsa_circ_0005909 are predicted to promote CADM1
expression by functioning as miRNAs sponge in human osteosarcoma.
PLoS One. 13(e0202896)2018.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Zhang M, Jia L and Zheng Y: circRNA
expression profiles in human bone marrow stem cells undergoing
osteoblast differentiation. Stem Cell Rev Rep. 15:126–138.
2019.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Gu X, Li M, Jin Y, Liu D and Wei F:
Identification and integrated analysis of differentially expressed
lncRNAs and circRNAs reveal the potential ceRNA networks during
PDLSC osteogenic differentiation. BMC Genet. 18(100)2017.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Fehlings MG, Tetreault LA, Riew KD,
Middleton JW, Aarabi B, Arnold PM, Brodke DS, Burns AS, Carette S,
Chen R, et al: A clinical practice guideline for the management of
patients with degenerative cervical myelopathy: Recommendations for
patients with mild, moderate, and severe disease and nonmyelopathic
patients with evidence of cord compression. Global Spine J. 7
(Suppl 3):70S–83S. 2017.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Tetreault L, Nakashima H, Kato S,
Kryshtalskyj M, Nagoshi N, Nouri A, Singh A and Fehlings MG: A
systematic review of classification systems for cervical
ossification of the posterior longitudinal ligament. Global Spine
J. 9:85–103. 2019.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Li Y, Wang Y, Li Y, Luo W, Jiang J, Zhao J
and Liu C: Controllable synthesis of biomimetic hydroxyapatite
nanorods with high osteogenic bioactivity. ACS Biomater Sci Eng.
6:320–328. 2020.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408.
2001.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Miao J, Sun J, Shi J, Chen Y and Chen D: A
novel anterior revision surgery for the treatment of cervical
ossification of posterior longitudinal ligament: Case report and
review of the literature. World Neurosurg. 113:212–216.
2018.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Franceschi RT, Ge C, Xiao G, Roca H and
Jiang D: Transcriptional regulation of osteoblasts. Ann N Y Acad
Sci. 1116:196–207. 2007.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Liu Y, Zhao Y, Chen Y, Shi G and Yuan W:
RUNX2 polymorphisms associated with OPLL and OLF in the Han
population. Clin Orthop Relat Res. 468:3333–3341. 2010.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Jafary F, Hanachi P and Gorjipour K:
Osteoblast differentiation on collagen scaffold with immobilized
alkaline phosphatase. Int J Organ Transplant Med. 8:195–202.
2017.PubMed/NCBI
|
|
24
|
Wei QS, Huang L, Tan X, Chen ZQ, Chen SM
and Deng WM: Serum osteopontin levels in relation to bone mineral
density and bone turnover markers in postmenopausal women. Scand J
Clin Lab Invest. 76:33–39. 2016.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Tsao YT, Huang YJ, Wu HH, Liu YA, Liu YS
and Lee OK: Osteocalcin mediates biomineralization during
osteogenic maturation in human mesenchymal stromal cells. Int J Mol
Sci. 18(159)2017.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Asari T, Furukawa K, Tanaka S, Kudo H,
Mizukami H, Ono A, Numasawa T, Kumagai G, Motomura S, Yagihashi S
and Toh S: Mesenchymal stem cell isolation and characterization
from human spinal ligaments. Biochem Biophys Res Commun.
417:1193–1199. 2012.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Ishihara K and Hirano T: Molecular basis
of the cell specificity of cytokine action. Biochim Biophys Acta.
1592:281–296. 2002.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Lin J, Cai R, Sun B, Dong J, Zhao Y, Miao
Q and Chen C: Gd@C82(OH)22 harnesses inflammatory regeneration for
osteogenesis of mesenchymal stem cells through JNK/STAT3 signaling
pathway. J Mater Chem B. 6:5802–5811. 2018.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Yu X, Wan Q, Cheng G, Cheng X, Zhang J,
Pathak JL and Li Z: CoCl2, a mimic of hypoxia, enhances bone marrow
mesenchymal stem cells migration and osteogenic differentiation via
STAT3 signaling pathway. Cell Biol Int. 42:1321–1329.
2018.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Matsuguchi T, Chiba N, Bandow K, Kakimoto
K, Masuda A and Ohnishi T: JNK activity is essential for Atf4
expression and late-stage osteoblast differentiation. J Bone Miner
Res. 24:398–410. 2009.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Yang Q, Han Y, Liu P, Huang Y, Li X, Jia
L, Zheng Y and Li W: Long noncoding RNA GAS5 promotes osteogenic
differentiation of human periodontal ligament stem cells by
regulating GDF5 and p38/JNK signaling pathway. Front Pharmacol.
11(701)2020.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Li X, Zheng Y, Zheng Y, Huang Y, Zhang Y,
Jia L and Li W: Circular RNA CDR1as regulates osteoblastic
differentiation of periodontal ligament stem cells via the
miR-7/GDF5/SMAD and p38 MAPK signaling pathway. Stem Cell Res Ther.
9(232)2018.PubMed/NCBI View Article : Google Scholar
|
|
33
|
Huang XQ, Cen X, Sun WT, Xia K, Yu LY, Liu
J and Zhao ZH: CircPOMT1 and circMCM3AP inhibit osteogenic
differentiation of human adipose-derived stem cells by targeting
miR-6881-3p. Am J Transl Res. 11:4776–4788. 2019.PubMed/NCBI
|
|
34
|
Huang J, Zhao L, Xing L and Chen D:
MicroRNA-204 regulates Runx2 protein expression and mesenchymal
progenitor cell differentiation. Stem Cells. 28:357–364.
2010.PubMed/NCBI View
Article : Google Scholar
|
|
35
|
Xu C, Zhang H, Gu W, Wu H, Chen Y, Zhou W,
Sun B, Shen X, Zhang Z, Wang Y, et al: The microRNA-10a/ID3/RUNX2
axis modulates the development of ossification of posterior
longitudinal ligament. Sci Rep. 8(9225)2018.PubMed/NCBI View Article : Google Scholar
|
|
36
|
Yuan X, Guo Y, Chen D, Luo Y, Chen D, Miao
J and Chen Y: Long non-coding RNA MALAT1 functions as miR-1 sponge
to regulate Connexin 43-mediated ossification of the posterior
longitudinal ligament. Bone. 127:305–314. 2019.PubMed/NCBI View Article : Google Scholar
|
|
37
|
Liao X, Tang D, Yang H, Chen Y, Chen D,
Jia L, Yang L and Chen X: Long non-coding RNA XIST may influence
cervical ossification of the posterior longitudinal ligament
through regulation of miR-17-5P/AHNAK/BMP2 signaling pathway.
Calcif Tissue Int. 105:670–680. 2019.PubMed/NCBI View Article : Google Scholar
|