Introduction
Prostate cancer (PC) has become one of the leading
causes of mortality in males across the world (1-5).
The risk factors of PC are complicated, including genetic factors,
age, external exposure and chronic urinary tract infections
(6). The incidence of PC has
increased markedly over recent years. For example, the incidence of
PC increased by nearly one-third between 2015 and 2018(7). Furthermore, numerous patients with PC
exhibit no symptoms and are therefore frequently misdiagnosed
(8). It is urgent to elucidate the
intrinsic mechanism underlying PC and to find efficient diagnosis
methods and treatments for PC.
miRNA is one of the non-protein coding RNAs, which
is capable of regulating gene expression at the post-translational
level. miRNA functions via binding its target gene, resulting in
inhibition of gene expression (9).
miRNAs were confirmed as vital functional molecules and are
involved in various types of cancer (10-13).
The interplay between miRNA and gene expression is complicated. One
miRNA often has more than one target gene and one gene is also
regulated by different miRNAs (14). As reported previously, miRNAs also
serve vital roles in PC (15,16).
miR-28-5p was found to exert tumor suppressor effects by mediating
SREBF2, which had been confirmed to be involved in PC (15). miR-636 also served a role in bone
metastasis of PC via targeting STAB1, MBNL2 and TNS1(16). Therefore, it may be a promising way
to identify the miRNAs that are associated with PC.
Phosphatase and tensin homolog (PTEN) is a common
tumor suppressor gene which has been found in many tumor types
(17,18). As reported, PTEN was also found to
be involved in PC (19).
Additionally, loss of PTEN expression indicated an unfavorable
prognosis in PC (20,21). The anomaly of DNA methylome and
transcriptome was caused by PTEN loss at different stages of PC
(22). A previous study indicated
that miRNA-1297 promoted cell proliferation via targeting PTEN in
testicular germ cell tumors (23).
Furthermore, concurrent activation of the ERK/AKT pathway is
implicated in PC progression (24).
Additionally, PTEN served as a negative regulator of these pathways
in PC, and PTEN loss may activate the ERK/AKT pathway and induce PC
progression (24). However, the
molecular basis of the association between miRNA-1297/PTEN and
ERK/AKT is unknown. To further study the intrinsic mechanism of PC,
the potential role of miRNA-1297/PTEN axis in PC was investigated
in the present study.
Materials and methods
Cell culture and transfection
RWPE-2 (CRL-11610), DU145 (HTB-81), VCaP (CRL-2876),
PC-3 (CRL-1435) and LNCaP (CRL-1740) cells were purchased from
American Type Culture Collection (ATCC). DU145, PC-3 and LNCaP
cells were cultured in RPMI-1640 medium (ATCC®30-2001™).
RWPE-2 cells were cultured in K-SFM (ATCC®17005-042™)
and VCaP cells were cultured in Dulbecco's modified Eagle's medium
(Gibco; Thermo Fisher Scientific, Inc.). All cells were cultured in
medium with fetal bovine serum (FBS; ATCC®30-2020™; 10%)
and gentamicin (Gibco; Thermo Fisher Scientific, Inc.; 50 mg/ml) at
a density of 2x104/ml in an incubator in an atmosphere
with 5% CO2 at 37˚C. After reaching 70% confluence, the
cells were transfected with 10 nM miRNA-1297 inhibitor (Shanghai
GenePharma, Inc.; 5'-CACCTGAATTACTTGAA-3'), NC inhibitor (Shanghai
GenePharma, Inc.; 5'-ATACTCAAGCTTCTGAC-3') or PTEN-overexpression
plasmid (Shanghai GenePharma, Inc.) using Lipofectamine 2000
(Invitrogen; Thermo Fisher Scientific, Inc.), according to the
manufacturer's protocols. In brief, the mixture of Lipofectamine
2000 and miRNA-1297 inhibitor or PTEN plasmid was incubated for 10
min at room temperature. Next, the mixture was added into medium
and incubated at 37˚C with cells for 48 h. Subsequently, cells were
used for the follow-up experiments.
Collection of clinical samples
The PC tissues and matched para-cancerous normal
tissues were collected at the North China University of Science and
Technology Affiliated Hospital between January 2016 and December
2017. The distance between the healthy and the cancer tissue was
within 5 cm and was confirmed by HE staining. A total of 90
patients who were diagnosed by histopathological examination were
enrolled in this study and all patients provided written informed
consent. All patients were male and the mean age was 57.2±7.1
(range, 50-79) years. The inclusion criteria were as follows:
Primary prostate cancer confirmed by the pathology, without
surgery, chemotherapy, radiotherapy or other treatment (25). The exclusion criteria were as
follows: Recurrent prostate cancer, treated by surgery,
radiotherapy or chemotherapy previously, combined with infectious
diseases, malignant tumors, severe liver and kidney disease,
pulmonary fibrosis, bone metabolic diseases, secondary renal
hypertension, systemic immune diseases and malignant tumor
complications (25). The present
study was approved by the Ethics Committee of North China
University of Science and Technology affiliated Hospital (Approval
number 2016-0311).
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR) assay
MirVana™ miRNA isolation kit (Ambion) was used for
the total RNA extraction from tissues and cells. DNA synthesis was
performed using TaqMan™ MicroRNA Reverse Transcription kit (kit no.
4366596; Thermo Fisher Scientific, Inc.) or 1st Strand cDNA
Synthesis kit (kit no. RR036A; Takara Bio, Inc.). Reverse
transcription was performed at 37˚C for 15 min and 85˚C for 5 sec.
ABI SYBR® Premix Ex Taq™ II (RR037B; Takara Bio, Inc.)
was used for quantification of the gene expression levels of PTEN
and miRNA-1297. The 7500 system (Applied Biosystems) was applied
for the RT-PCR assay. β-actin served as the internal control. The
miRNA-1297 primer (MBS825976) was designed and purchased from
MyBioSource company. The primer sequences for PTEN were as follows:
Forward, 5'-CGGCAGCATCAAATGTTTCAG-3' and reverse,
5'-AACTGGCAGGTAGAAGGCAACTC-3'. U6 was used as the control and the
primer sequences were as follows: Forward, 5'-CTCGCTTCGGCAGCACA-3'
and reverse, 5'-AACGCTTCACGAATTTGCGT-3'. The following
thermocycling conditions were used: 95˚C for 5 min, followed by 40
cycles of 95˚C for 5 sec and 60˚C for 34 sec. The gene expression
level were calculated using the 2-ΔΔCt method (26).
Cell viability assay
RWPE-2 cells were seeded into a 96-well plate
following transfection with miRNA-1297 inhibitor for 24 or 48 h.
Next, 200 µl medium was added to each well, with 10 µl freshly
prepared MTT (5 mg/ml) and 100 µl dissolved formazan liquid
(dissolved in PBS). Subsequently, the optical density (OD) at 490
nm of each well was measured. The ratio of viable cells was
calculated by dividing OD readings of the transfected groups into
the control group.
Transwell assay
The cell migration and invasion were investigated
using a Transwell assay. A total of 200 µl PC cells were suspended
in serum-free RPMI-1640 medium at a density of 1x106
cells/ml, prior to being added into the upper chambers.
Subsequently, 500 µl RPMI-1640 medium containing 10% FBS was added
into the lower chambers. Following incubation for 24 h at 37˚C,
cells that had migrated to the bottom surface of the insert were
fixed with 4% paraformaldehyde for 10 min and stained with 0.1%
crystal violet for 30 min both at room temperature. The cells
numbers in five randomly selected fields were counted under an
inverted microscope (magnification, x40; Olympus Corporation). For
the cell invasion assay, the upper chamber was coated with Matrigel
(BD Biosciences) and the other procedures were the same with cell
migration assay.
Luciferase reporter assay
The luciferase reporter assay was performed using
the Dual-Luciferase Reporter assay system kit (Promega Corporation;
E1910), according to the manufacturer's protocols. 293T cells were
used as the tool cell to detect the binding of miRNA-1297 and PTEN.
As predicted in the targetscan website (http://www.targetscan.org/), there are mainly four
binding sites between miRNA-1297 and PTEN. The wild-type (WT) or
mutant (MUT) gene located at positions 41-47, 1261-1268, 2619-2626
and 3800-3807 of the PTEN 3'-UTR was cloned into the reporter
vector (Beyotime Institute of Biotechnology; D2112). Next, 10 nM
the negative control (NC; BBI Life Sciences;
5'-UCACAACCUCCUAGAAAGAGUAGA-3') or miRNA-1297 mimic (BBI Life
Sciences; 5'-UUCAAGUAAUUCAGGUG-3') and 100 ng reporter plasmid were
co-transfected into the 293T cells. After 48 h, the dual-luciferase
reporter assay system (Promega Corporation) was applied for the
measurement of the relative luciferase activity. Firefly luciferase
activity was normalized to that of Renilla luciferase
activity.
Western blotting
Total protein was extracted using RIPA lysis buffer
(Boster Biological Technology; cat. no. AR0105). Next, the proteins
were quantified using a BCA kit (Beyotime Institute of
Biotechnology). A total of 20 µg protein (per lane) was loaded and
separated by 10% SDS-PAGE. Next, the PVDF membranes were used for
blotting the separated proteins and were then blocked with 5%
skimmed milk for 1 h at room temperature. Subsequently, the
membranes were incubated with the following primary antibodies:
Anti-p-Akt antibody (dilution, 1:1,000; cat. no. ab38449; Abcam),
anti-Akt antibody (dilution, 1:1,000; cat. no. ab32505; Abcam),
anti-Erk antibody (dilution, 1:1,000; cat. no. 9102; Cell Signaling
Technology, Inc.), anti-p-Erk (dilution, 1:1,000; cat. no. 4370;
Cell Signaling Technology, Inc.), anti-E-cadherin antibody
(dilution, 1:1,000; cat. no. 7559; Cell Signaling Technology,
Inc.), anti-N-cadherin antibody (dilution, 1:1,000; cat. no. 3195;
Cell Signaling Technology, Inc.), anti-MMP-2 antibody (dilution,
1:1,000; cat. no. 40994; Cell Signaling Technology, Inc.),
anti-MMP-9 antibody (dilution, 1:1,000; cat. no. 13667; Cell
Signaling Technology, Inc.) and anti-beta actin antibody (dilution,
1:2,000; cat. no. ab8226; Abcam) at 4˚C overnight. Next, the
membranes were incubated with HRP-conjugated anti-rabbit IgG
secondary antibody for 1 h at room temperature (dilution, 1:5,000;
Sigma-Aldrich; cat. no. 12-348). β-actin served as the control. An
enhanced chemiluminescence kit (Millipore) was used for
visualization of the positive bands. ImageJ software (National
Institutes of Health) was applied for quantification of band
density.
Statistical analysis
All data in the present study were processed by SPSS
software (version 19.0; IBM Corp.). All experiments were repeated
at least 3 times. The data are presented as the mean ± standard
deviation. The difference between groups was analyzed via student's
t test or one-way analysis of variance, followed by Duncan's post
hoc test. Paired Student's t-test was used for the comparison of
miRNA-1297 expression in tumor tissues and adjacent tissues.
Correlation analysis was performed using Pearson's correlation
analysis. A survival curve was constructed using the Kaplan-Meier
method, followed by the Log rank test. To construct the survival
curve, patients were divided into 2 groups according to the median
of miRNA-1297 expression (3.285). The patients with a relative
miRNA-1297 expression of >3.285 were included in the high
expression group and the remaining patients were included in the
low expression group. P<0.05 was considered to indicate a
statistically significant difference.
Results
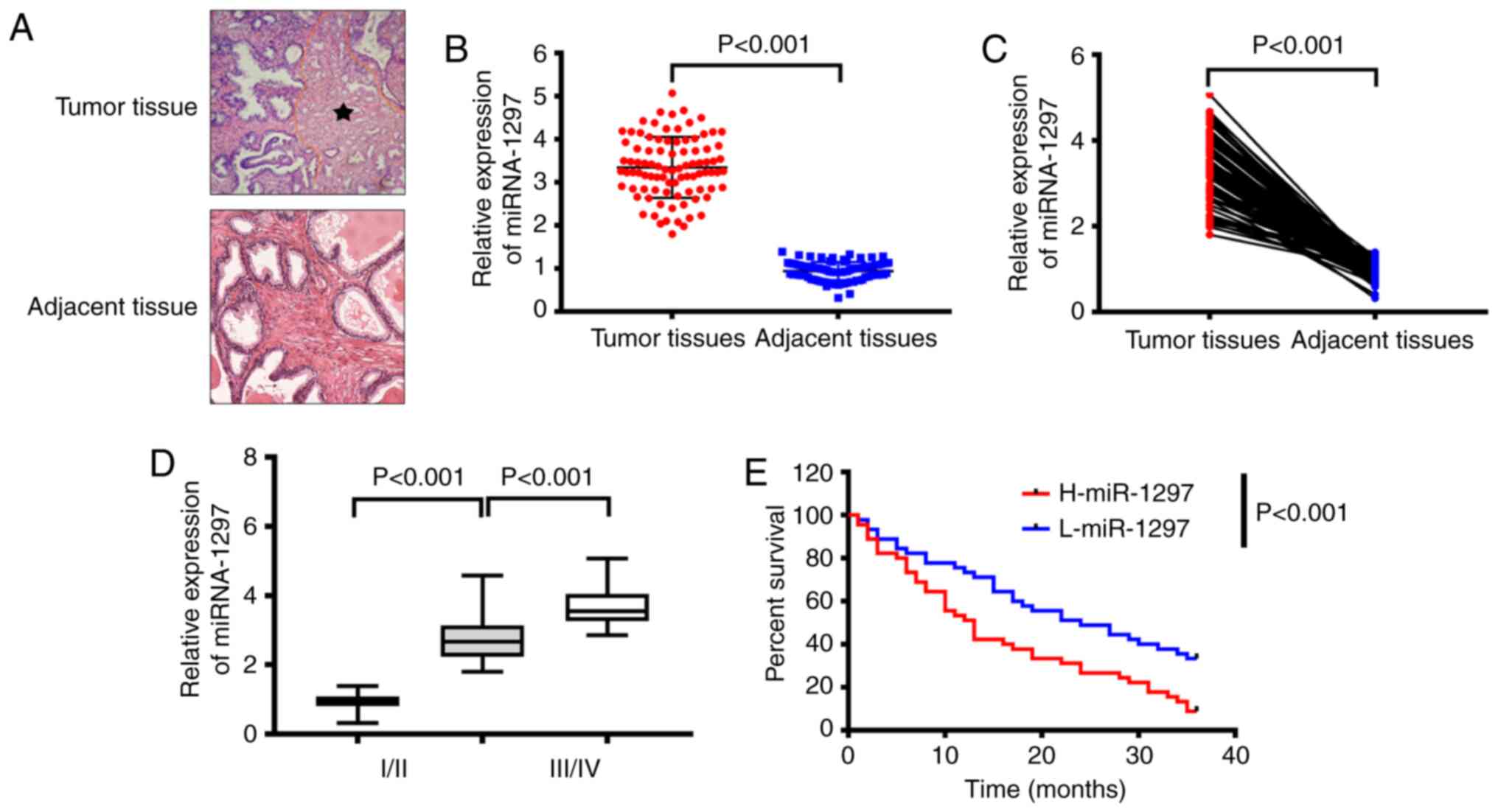
miRNA-1297 is highly expressed in PC
tissue
The present study first investigated the miRNA-1297
level in tumor tissues and para-carcinoma tissues. The results
demonstrated that the miRNA-1297 level was higher in tumor tissues
than that in para-carcinoma tissues (Fig. 1A and B). Furthermore, the miRNA-1297 level in
tumor tissues from patients with stage I/II PC was higher than that
in the non-cancerous tissues but lower than that in the tumor
tissues at stage III/IV (Fig. 1C).
Subsequently, the effects of miRNA-1297 on overall survival rates
were investigated. As shown in Fig.
1D, the overall survival rates in patients with high miRNA-1297
expression were lower than that in patients with low miRNA-1297
expression.
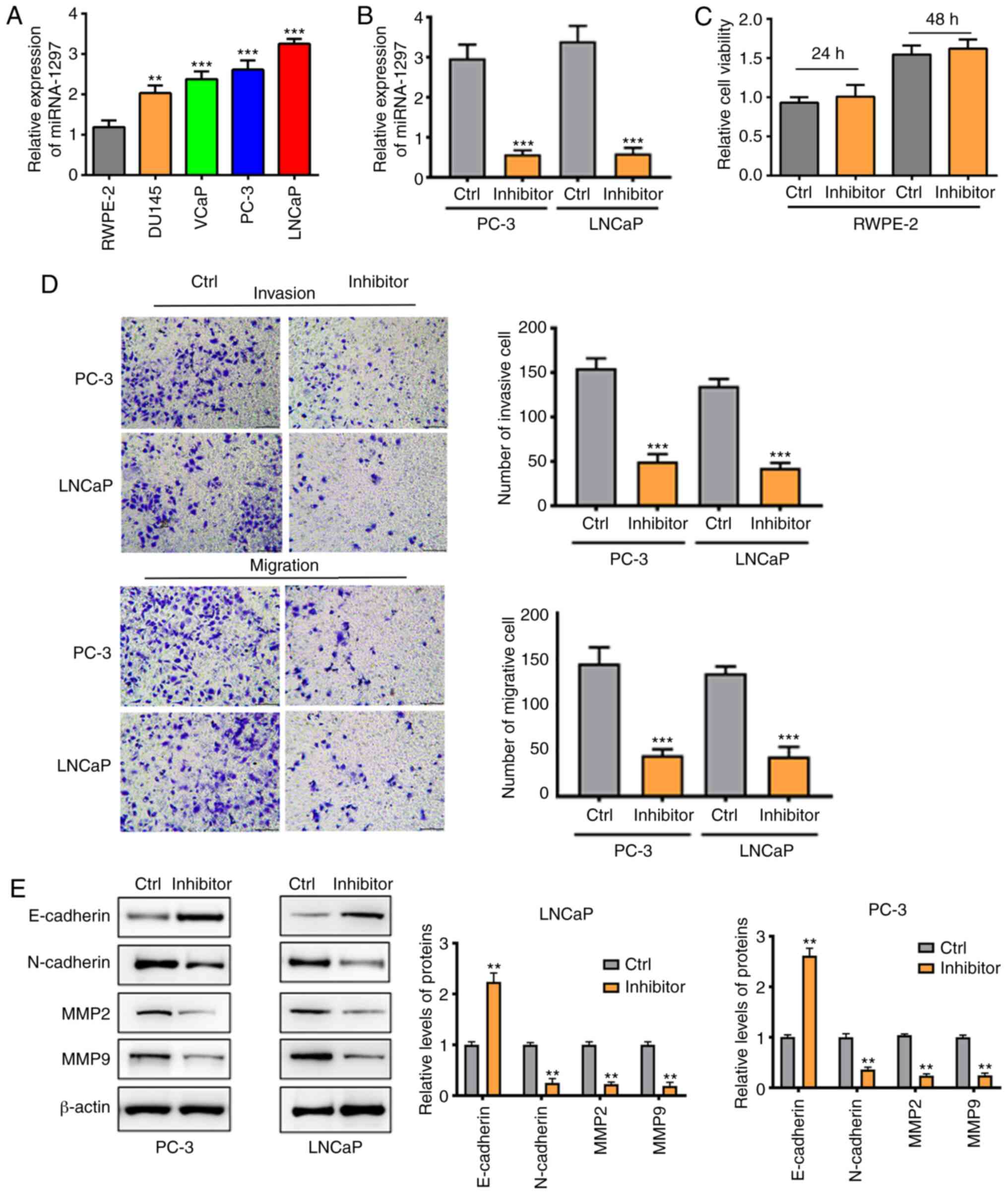
miRNA-1297 is overexpressed in PC
cells and silencing miRNA-1297 inhibits cell invasion and migration
in PC-3 cells and LNCaP cells
To begin with, the miRNA-1297 level was measured in
different PC cell lines. The results indicated that the miRNA-1297
level was notably increased in DU145, VCaP, PC-3 and LNCaP cancer
cells, compared with normal RWPE-2 cells (Fig. 2A). As cell invasion and migration
are important for cancer cells, the role of miRNA-1297 in cell
invasion and migration was further investigated. An inhibitor was
used to silence miRNA-1297 expression in cancer cells. As shown in
Fig. 2B, the miRNA-1297 level was
downregulated in the inhibitor groups, suggesting that miRNA-1297
was silenced successfully. The safety of the miRNA-1297 inhibitor
was assessed using RWPE-2 cells. As shown in Fig. 2C, transfection with the miRNA-1297
inhibitor did not influence the proliferation of RWPE-2 cells.
Next, the cell migration and invasion were investigated. Results
demonstrated that the number of migrated and invasive cells in the
inhibitor group was significantly less than that in the control
group (Fig. 2D). Furthermore, the
protein expression of MMP-2, MMP-9, E-cadherin and N-cadherin was
investigated. The Western blotting results indicated that silencing
miRNA-1297 may inhibit PC metastasis by decreasing MMP-2, MMP-9 and
N-cadherin expression (Fig.
2E).
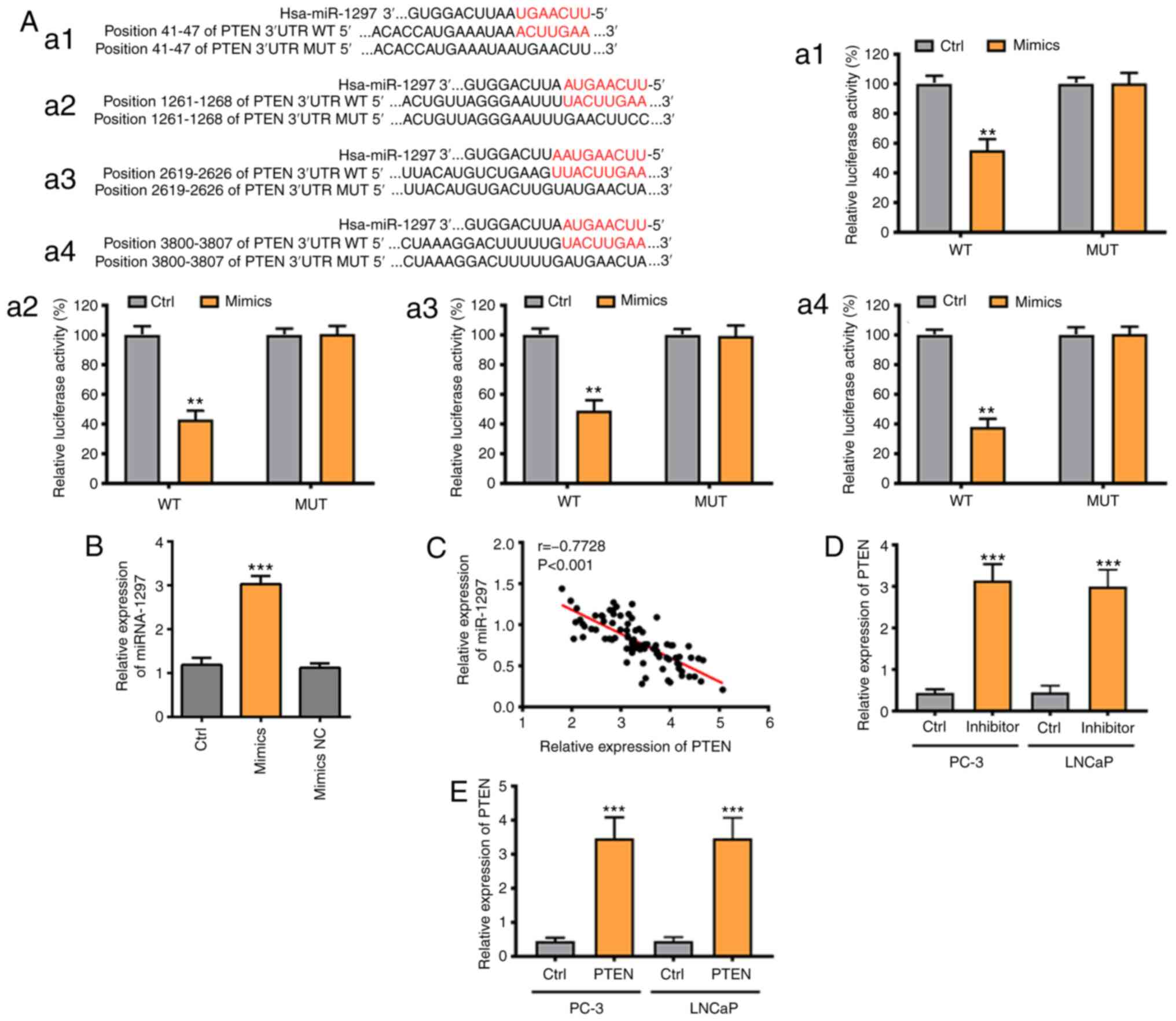
miRNA-1297 is capable of targeting
PTEN
As shown in Targetscan, PTEN was the predicted
target of miRNA-1297 with four binding sites (Fig. 3A). The binding sites were all
verified via a Luciferase Reporter Assay. The relative luciferase
activity in the wild-type of PTEN and in the mimics group was the
lowest among all the study groups and the luciferase activity
remained unchanged in the remaining groups (Fig. 3A). These results confirmed that
miRNA-1297 was able to bind to PTEN via four binding sites. PCR
results also indicated that the PTEN level was negatively
associated with the miRNA-1297 level in patients with PC (Fig. 3B). Furthermore, the PTEN level in
the inhibitor group and the PTEN-overexpression group was markedly
increased, compared with the control group (Fig. 3C and D). In conclusion, PTEN was the directive
target of miRNA-1297.
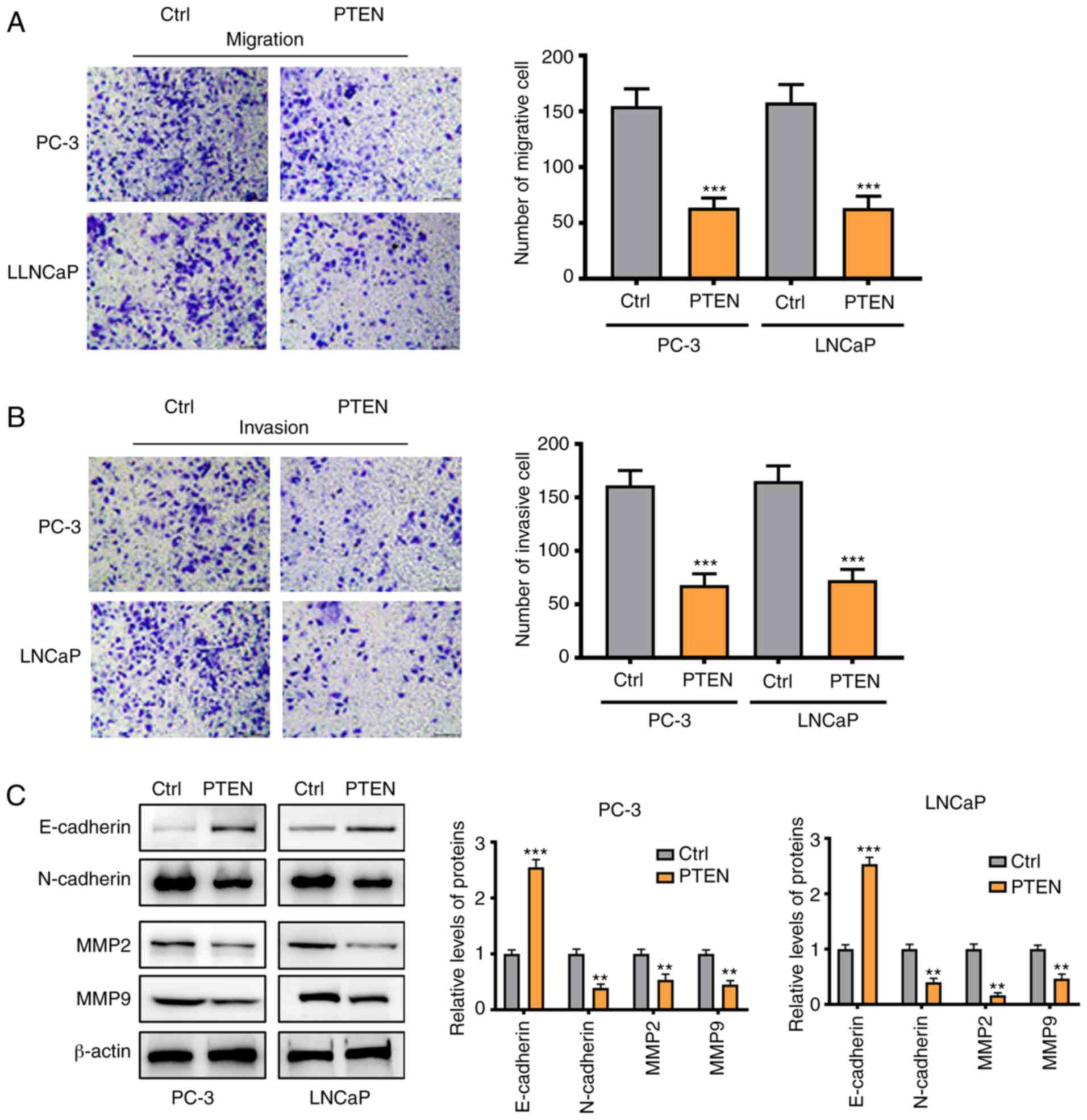
Silencing miRNA-1297 inhibits cell
invasion and migration of PC via upregulating PTEN and inhibiting
the AKT/ERK pathway
As shown in Fig. 4A
and B, cell migration and invasion
were suppressed in the PTEN-overexpression group, which was in
accordance with the results in the miRNA-1297 inhibitor group
(Fig. 2C and D). The Western blotting also yielded
similar results. Following overexpression of PTEN, the MMP-2, MMP-9
and N-cadherin protein expression was significantly decreased
(Fig. 4C). These results further
indicated that PTEN was the downstream target of miRNA-1297. To
investigate the signaling pathways influenced by miRNA-1297 and
PTEN, the levels of AKT, ERK and their corresponding phosphorylated
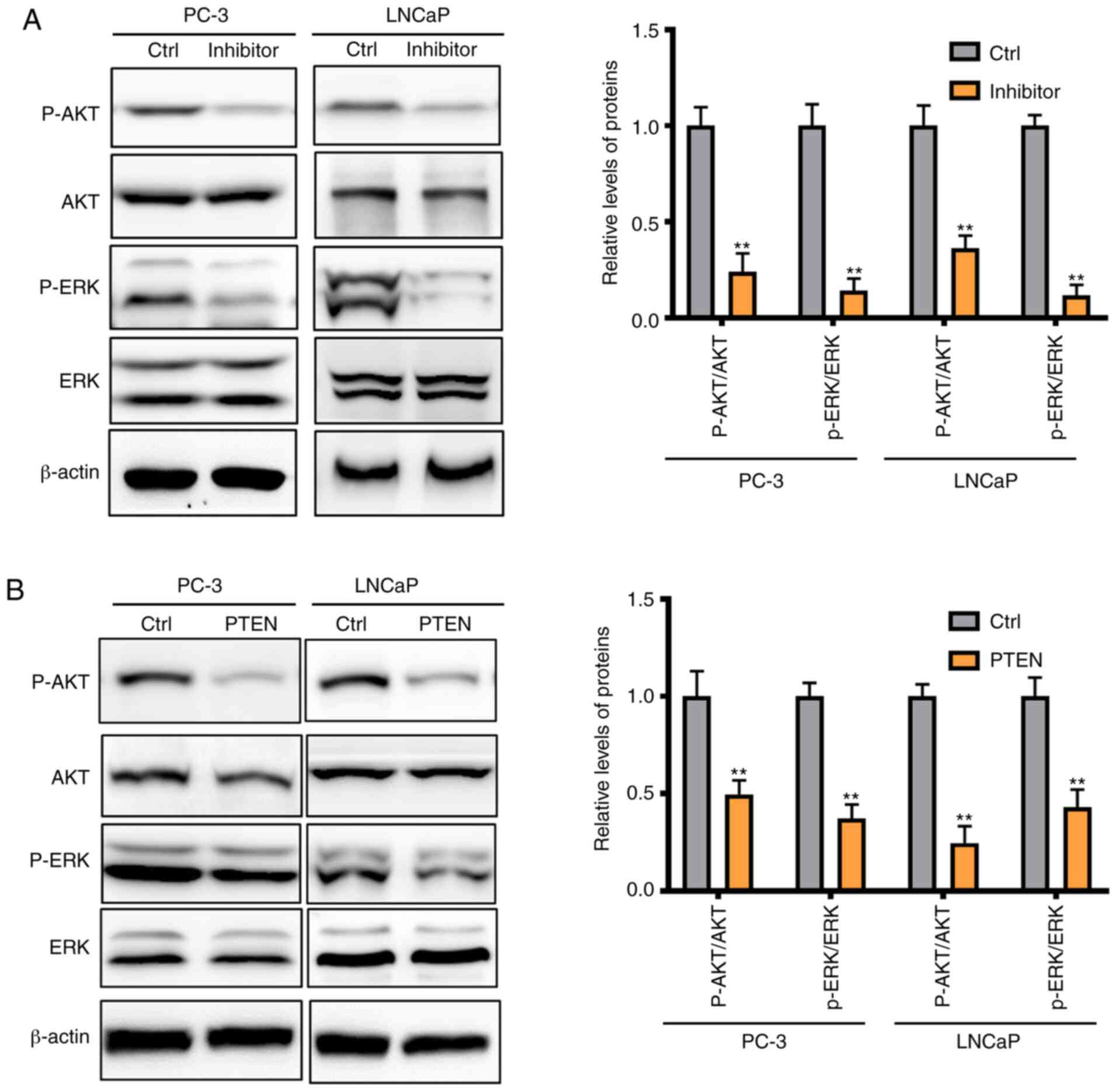
forms were also investigated in the present study. The expression
of p-AKT and p-ERK were significantly inhibited by silencing
miRNA-1297 while the total AKT and ERK levels remained unchanged
(Fig. 5A). Similar results were
found following overexpression of PTEN, and p-AKT and p-ERK
expression was decreased in the PTEN-overexpression group (Fig. 5B). All these findings suggested that
silencing miRNA-1297 inhibited cell invasion and migration via
upregulation of PTEN and suppression of the AKT/ERK pathway.
Discussion
PC is one of the major causes of fatality in males
and severely threatens the health of males (27). PTEN loss has been confirmed as a
crucial contributor for PC (26-28).
PTEN is associated with a poor prognosis and recurrence of PC
(28-30).
Luminal cells of the prostate, which have been confirmed to be the
prime origin of PC, generated tumors with stronger aggressivity
under the condition of PTEN loss (31,32).
Furthermore, PTEN was also reported to induce abnormal methylome
and transcriptome of DNA (22).
miRNA is confirmed as a crucial regulator in various
cancer types via binding to its target genes. Regulation of the
gene expression resulting in alteration of cell behaviors,
including cell apoptosis, proliferation, invasion and migration.
Therefore, screening the miRNA, which serves as the regulator of
PTEN, is of great significance for the treatment of PC. In the
present study, the results of Targetscan revealed that miRNA-1297
may bind to PTEN via four binding sites. miRNA-1297 was reported to
serve a role in various cancer types. For instance, miRNA-1297
promoted cell proliferation via targeting PTEN in germ cell tumors
(23). miRNA-1297 also exerted a
promotive effect on cervical carcinoma progression via targeting
PTEN (33). Furthermore, miRNA-1297
was confirmed as a contributor toward oral squamous cell carcinoma
progression via regulating PTEN (34). The present study revealed that
miRNA-1297 was overexpressed in PC tissues. Furthermore, miRNA-1297
was upregulated in patients with stage III/IV PC, compared with
those with stage I/II PC. The patients with higher miRNA-1297 level
also had lower survival rates. These findings demonstrated that
miRNA-1297 also acted as an oncogene that is involved in the
progression of PC. The present study further investigated the
miRNA-1297 level in different PC cell lines. It was found that
miRNA-1297 level was significantly upregulated in all the cancer
cells, particularly in PC-3 cells and LNCaP cells. The results
indicated that miRNA-1297 was overexpressed in the
hormone-sensitive PC LNCaP cell line and the hormone-insensitive PC
cell lines, PC-3, VCaP and DU145. Subsequently, the experiments
were performed using LNCaP (left supraclavicular lymph node
derived, early stage of PC) and PC-3 (bone derived, poorly
differentiated) cell lines, for the representation of different
types and stages of PC. The miRNA-1297 expression in
hormone-insensitive VCaP and DU145 cells was also upregulated,
indicating that miRNA-1297 expression may serve a role in bone and
brain metastases of PC. However, further research may be required.
Taken together, the results of the present study revealed that
miRNA-1297 served a broader role in different prostate cancer cells
and may be a promising therapeutic target.
Cell proliferation and invasion are of great
importance for cancer progression and numerous previous studied
have reported that they are regulated by miRNA (35-38).
The results of the present study indicated that silencing
miRNA-1297 inhibited cell migration and invasion in PC-3 cells and
LNCaP cells. This suggested that miRNA-1297 was involved in PC cell
migration and invasion.
PTEN was reported to be the directive target of
miRNA-1297 in numerous cancer types (23,33,34).
In the present study, the four binding sites predicted by
Targetscan were confirmed via Luciferase Reporter Assay. The
miRNA-1297 level was negatively associated with the PTEN level in
patients with PC and silencing miRNA-1297 may upregulate PTEN in PC
cells. In addition, overexpression of PTEN in PC cells
significantly suppressed the cell migration and invasion. Based on
these results, we hypothesized that miRNA-1297 was involved in the
progression of PC via targeting PTEN. MMP-2, MMP-9, E-cadherin and
N-cadherin were the metastasis-associated proteins which served
important roles in the metastasis of PC (39). The results of the present indicated
that miRNA-1297-silencing and PTEN-overexpression inhibited PC cell
migration via suppressing MMP-2, MMP-9 and N-cadherin expression.
These results were also consistent with those of previous reports
(39,40).
The AKT/ERK pathway was highly expressed in PC and
associated with an unfavorable prognosis (41,42).
Additionally, previous studies confirmed that the AKT/ERK pathway
was suppressed by PTEN (43-45).
In the present study, the phosphorylation level of AKT and ERK were
inhibited following silencing of miRNA-1297. These changes to the
AKT and ERK pathway may be attributed to the upregulation of PTEN.
These findings suggested that the AKT/ERK pathway was the
downstream pathway of miRNA-1297/PTEN axis. Taken together, these
results suggested that miRNA-1297-silencing restrained cell
migration and invasion via blockage of the AKT/ERK pathway by
targeting PTEN.
There are certain limitations to the present study.
To begin with, only the miRNA-1297-silenced model was used and the
overexpression of miRNA-1297 was not tested in the experiments.
Furthermore, the role of miRNA-1297 was not further investigated in
DU145 and VCaP cell lines. More cell models and more cell lines
should be used in future studies.
In conclusion, miRNA-1297 was found to be
overexpressed in PC tissues and associated with the progression of
PC. Silencing miRNA-1297 was confirmed to inhibit cell migration
and invasion via blocking of the AKT/ERK pathway by targeting PTEN.
These findings provided evidence of developing a novel strategy for
the treatment of PC in the future.
Acknowledgements
Not applicable.
Funding
Funding: No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
LW conceived and designed the experiments, performed
the experiments, analyzed the data, contributed
reagents/materials/analytical tools, wrote the manuscript, prepared
figures and tables, and reviewed drafts of the paper. JG performed
the experiments, analyzed the data and prepared figures and tables.
YZ performed the experiments. SK conceived and designed the
experiments, and reviewed drafts of the paper. LW and SK confirm
the authenticity of all the raw data. All authors read and approved
the final manuscript.
Ethics approval and consent to
participate
The present study was approved by the Ethics
Committee of North China University of Science and Technology
affiliated Hospital (approval no. 2016-0311).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2018. CA Cancer J Clin. 68:7–30. 2018.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Rani A, Dasgupta P and Murphy JJ: Prostate
cancer: The role of inflammation and chemokines. Am J Pathol.
189:2119–2137. 2019.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Mulholland EJ, Green WP, Buckley NE and
McCarthy HO: Exploring the potential of microRNA Let-7c as a
therapeutic for prostate cancer. Mol Ther Nucleic Acids.
18:927–937. 2019.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Rawla P: Epidemiology of prostate cancer.
World J Oncol. 10:63–89. 2019.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Bray F, Ferlay J, Soerjomataram I, Siegel
RL, Torre LA and Jemal A: Global cancer statistics 2018: GLOBOCAN
estimates of incidence and mortality worldwide for 36 cancers in
185 countries. CA Cancer J Clin. 68:394–424. 2018.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Cuzick J, Thorat MA, Andriole G, Brawley
OW, Brown PH, Culig Z, Eeles RA, Ford LG, Hamdy FC, Holmberg L, et
al: Prevention and early detection of prostate cancer. Lancet
Oncol. 15:e484–e492. 2014.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Pernar CH, Ebot EM, Wilson KM and Mucci
LA: The epidemiology of prostate cancer. Cold Spring Harb Perspect
Med. 8(a030361)2018.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Grozescu T and Popa F: Prostate cancer
between prognosis and adequate/proper therapy. J Med Life. 10:5–12.
2017.PubMed/NCBI
|
|
9
|
Bertoli G, Cava C and Castiglioni I:
MicroRNAs: New biomarkers for diagnosis, prognosis, therapy
prediction and therapeutic tools for breast cancer. Theranostics.
5:1122–1143. 2015.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Qadir MI and Faheem A: miRNA: A diagnostic
and therapeutic tool for pancreatic cancer. Crit Rev Eukaryot Gene
Expr. 27:197–204. 2017.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Tutar L, Özgür A and Tutar Y: Involvement
of miRNAs and pseudogenes in cancer. Methods Mol Biol. 1699:45–66.
2018.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Tutar Y: miRNA and cancer; computational
and experimental approaches. Curr Pharm Biotechnol.
15(429)2014.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Van Roosbroeck K and Calin GA: Cancer
hallmarks and MicroRNAs: The therapeutic connection. Adv Cancer
Res. 135:119–149. 2017.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Acunzo M and Croce CM: MicroRNA in cancer
and cachexia-a mini-review. The J Infect Dis. 212 (Suppl
1):S74–S77. 2015.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Fazio S, Berti G, Russo F, Evangelista M,
D'Aurizio R, Mercatanti A, Pellegrini M and Rizzo M: The miR-28-5p
targetome discovery identified SREBF2 as one of the mediators of
the miR-28-5p tumor suppressor activity in prostate cancer cells.
Cells. 9(354)2020.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Zhu Z, Wen Y, Xuan C, Chen Q, Xiang Q,
Wang J, Liu Y, Luo L, Zhao S, Deng Y and Zhao Z: Identifying the
key genes and microRNAs in prostate cancer bone metastasis by
bioinformatics analysis. FEBS Open Bio. 10:674–688. 2020.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Tsai YS, Jou YC, Tsai HT, Shiau AL, Wu CL
and Tzai TS: Prothymosin-α enhances phosphatase and tensin homolog
expression and binds with tripartite motif-containing protein 21 to
regulate Kelch-like ECH-associated protein 1/nuclear factor
erythroid 2-related factor 2 signaling in human bladder cancer.
Cancer Sci. 110:1208–1219. 2019.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Ullman D, Dorn D, Rais-Bahrami S and
Gordetsky J: Clinical utility and biologic implications of
phosphatase and tensin homolog (PTEN) and ETS-related gene (ERG) in
prostate cancer. Urology. 113:59–70. 2018.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Morais CE, Gurgel DC, Teixeira AC, Mattos
TVA, Silva AVAd and Tavora F: Prevalence of ERG expression and PTEN
loss in a Brazilian prostate cancer cohort. Braz J Med Biol Res.
52:e8483. 2019.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Chaux A, Peskoe SB, Gonzalez-Roibon N,
Schultz L, Albadine R, Hicks J, De Marzo AM, Platz EA and Netto GJ:
Loss of PTEN expression is associated with increased risk of
recurrence after prostatectomy for clinically localized prostate
cancer. Mod Pathol. 25:1543–1549. 2012.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Lotan TL, Gurel B, Sutcliffe S, Esopi D,
Liu W, Xu J, Hicks JL, Park BH, Humphreys E, Partin AW, et al: PTEN
protein loss by immunostaining: Analytic validation and prognostic
indicator for a high risk surgical cohort of prostate cancer
patients. Clin Cancer Res. 17:6563–6573. 2011.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Wang C, Feng Y, Zhang C, Cheng D, Wu R,
Yang Y, Sargsyan D, Kumar D and Kong AN: PTEN deletion drives
aberrations of DNA methylome and transcriptome in different stages
of prostate cancer. FASEB J. 34:1304–1318. 2020.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Yang NQ, Zhang J, Tang QY, Guo JM and Wang
GM: miRNA-1297 induces cell proliferation by targeting phosphatase
and tensin homolog in testicular germ cell tumor cells. Asian Pac J
Cancer Prev. 15:6243–6246. 2014.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Patel R, Gao M, Ahmad I, Fleming J, Singh
LB, Rai TS, McKie AB, Seywright M, Barnetson RJ, Edwards J, et al:
Sprouty2, PTEN, and PP2A interact to regulate prostate cancer
progression. J Clin Invest. 123:1157–1175. 2013.PubMed/NCBI View
Article : Google Scholar
|
|
25
|
Liauw SL, Kropp LM, Dess RT and Oto A:
Endorectal MRI for risk classification of localized prostate
cancer: Radiographic findings and influence on treatment decisions.
Urol Oncol. 34:416. e415–421. 2016.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408.
2001.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2019. CA Cancer J Clin. 69:7–34. 2019.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Sun J, Li S, Wang F, Fan C and Wang J:
Identification of key pathways and genes in PTEN mutation prostate
cancer by bioinformatics analysis. BMC Med Genet.
20(191)2019.PubMed/NCBI View Article : Google Scholar
|
|
29
|
de Bono JS, De Giorgi U, Rodrigues DN,
Massard C, Bracarda S, Font A, Arranz Arija JA, Shih KC, Radavoi
GD, Xu N, et al: Randomized phase II study evaluating Akt blockade
with ipatasertib, in combination with abiraterone, in patients with
metastatic prostate cancer with and without PTEN loss. Clin Cancer
Res. 25:928–936. 2019.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Wise HM, Hermida MA and Leslie NR:
Prostate cancer, PI3K, PTEN and prognosis. Clin Sci (Lond).
131:197–210. 2017.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Wang ZA, Toivanen R, Bergren SK, Chambon P
and Shen MM: Luminal cells are favored as the cell of origin for
prostate cancer. Cell Rep. 8:1339–1346. 2014.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Pietrzak K, Kuzyakiv R, Simon R, Bolis M,
Bär D, Aprigliano R, Theurillat JP, Sauter G and Santoro R: TIP5
primes prostate luminal cells for the oncogenic transformation
mediated by PTEN-loss. Proc Natl Acad Sci USA. 117:3637–3647.
2020.PubMed/NCBI View Article : Google Scholar
|
|
33
|
Chen Z, Zhang M, Qiao Y, Yang J and Yin Q:
MicroRNA-1297 contributes to the progression of human cervical
carcinoma through PTEN. Artif Cells Nanomed Biotechnol. 46 (Supp
2):S1120–S1126. 2018.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Liang L, Feng L and Wei B: microRNA-1297
involves in the progression of oral squamous cell carcinoma through
PTEN. Saudi J Biol Sci. 25:923–927. 2018.PubMed/NCBI View Article : Google Scholar
|
|
35
|
Chen X, Zhang L, Song Q and Chen Z:
MicroRNA-216b regulates cell proliferation, invasion and cycle
progression via interaction with cyclin T2 in gastric cancer.
Anticancer Drugs. 31:623–631. 2020.PubMed/NCBI View Article : Google Scholar
|
|
36
|
Hu X, Tan S, Yin H, Khoso PA, Xu Z and Li
S: Selenium-mediated gga-miR-29a-3p regulates LMH cell
proliferation, invasion, and migration by targeting COL4A2.
Metallomics. 12:449–459. 2020.PubMed/NCBI View Article : Google Scholar
|
|
37
|
Huang R, Li J, Pan F, Zhang B and Yao Y:
The activation of GPER inhibits cells proliferation, invasion and
EMT of triple-negative breast cancer via CD151/miR-199a-3p
bio-axis. Am J Transl Res. 12:32–44. 2020.PubMed/NCBI
|
|
38
|
Wan P, Bai X, Yang C, He T, Luo L, Wang Y,
Fan M, Wang Z, Lu L, Yin Y, et al: miR-129-5p inhibits
proliferation, migration, and invasion in rectal adenocarcinoma
cells through targeting E2F7. J Cell Physiol. 235:5689–5701.
2020.PubMed/NCBI View Article : Google Scholar
|
|
39
|
Jin W, Chen F, Wang K, Song Y, Fei X and
Wu B: miR-15a/miR-16 cluster inhibits invasion of prostate cancer
cells by suppressing TGF-beta signaling pathway. Biomed
Pharmacother. 104:637–644. 2018.PubMed/NCBI View Article : Google Scholar
|
|
40
|
Yang F, Yu N, Wang H, Zhang C, Zhang Z, Li
Y, Li D, Yan L, Liu H and Xu Z: Downregulated expression of
hepatoma-derived growth factor inhibits migration and invasion of
prostate cancer cells by suppressing epithelial-mesenchymal
transition and MMP2, MMP9. PLoS One. 13(e0190725)2018.PubMed/NCBI View Article : Google Scholar
|
|
41
|
Mukherjee R, Bartlett JM, Krishna NS,
Underwood MA and Edwards J: Raf-1 expression may influence
progression to androgen insensitive prostate cancer. Prostate.
64:101–107. 2005.PubMed/NCBI View Article : Google Scholar
|
|
42
|
Steelman LS, Chappell WH, Abrams SL, Kempf
RC, Long J, Laidler P, Mijatovic S, Maksimovic-Ivanic D, Stivala F,
Mazzarino MC, et al: Roles of the Raf/MEK/ERK and
PI3K/PTEN/Akt/mTOR pathways in controlling growth and sensitivity
to therapy-implications for cancer and aging. Aging (Albany NY).
3:192–222. 2011.PubMed/NCBI View Article : Google Scholar
|
|
43
|
Bao L, Yan Y, Xu C, Ji W, Shen S, Xu G,
Zeng Y, Sun B, Qian H, Chen L, et al: MicroRNA-21 suppresses PTEN
and hSulf-1 expression and promotes hepatocellular carcinoma
progression through AKT/ERK pathways. Cancer Lett. 337:226–236.
2013.PubMed/NCBI View Article : Google Scholar
|
|
44
|
Zheng S, Wang S, Zhang Q, Zhang Z and Xu
S: Avermectin inhibits neutrophil extracellular traps release by
activating PTEN demethylation to negatively regulate the PI3K-ERK
pathway and reducing respiratory burst in carp. J Hazard Mater.
389(121885)2020.PubMed/NCBI View Article : Google Scholar
|
|
45
|
Lu X, Xue B, Zhang T, Zhou X and Zhang Y:
Down-regulation of microRNA-10a mediates the anti-tumor effect of
icaritin in A549 cells via the PTEN/AKT and ERK pathway. Gen
Physiol Biophys. 38:525–533. 2019.PubMed/NCBI View Article : Google Scholar
|