Introduction
According to World Health Organization estimates,
alcohol consumption resulted in 3 million alcohol-attributable
deaths globally (5.3% of all deaths worldwide in 2016) and 5.1% of
the global disease burden in 2016(1). Alcohol abuse represents the most
common inducer of liver disease and is a considerable contributor
to the global burden of disease (2). More seriously, harmful alcohol
consumption is well recognized to be the major cause of alcoholic
liver disease (ALD) and liver cirrhosis worldwide (3). A previous study confirmed that
drinking pattern, alcohol quantity and nutritional status directly
influence the progression of alcoholic liver disease (4). In addition, it is intriguing to note
that intestinal homeostasis has been recently shown to serve a
pivotal role in modulating the development of alcohol-induced liver
injury (5-7).
Alcohol consumption may cause an imbalance in
intestinal flora by notable altering bacterial abundance and
diversity (8), and the profiles of
disordered gut microbiota involved in alcohol-related liver disease
have been well characterized with an increase in
Proteobacteria and Fusobacteria and a decrease in
Bacteroidetes and Lactobacillus species (9,10).
Another predominant feature of ALD pathogenesis is leakiness of the
gut barrier, resulting in the translocation of enterobacteria and
bacterial products (particularly endotoxemia, LPS) from the
intestinal lumen into the liver (11). These pathogen-associated molecular
patterns (PAMPs; e.g., LPS) potently activate toll-like receptors
(TLRs, particularly TLR4) and lead to nuclear factor-κB (NF-κB) and
inflammatory cytokine production (8,12,13).
These cytokines consequently attract leukocytes in liver tissues,
which further promotes hepatic injury (14,15).
Therefore, it can be reasonably inferred that improving
alcohol-induced intestinal dysfunction may represent a key for
effective therapy.
Notably, liver steatosis is the common response to
excessive alcohol exposure and is characterized by the deposition
of fat and abnormal lipid metabolism in hepatocytes (16,17),
of which fatty-acid transport and oxidation are disturbed. Alcohol
enhances upregulation of hepatic fatty acid transporters (18), including CD36/fatty acid translocase
and fatty acid transport protein. In addition, previous studies
have revealed that alcohol or its metabolic products inactivate the
peroxisome proliferator activated receptor alpha (PPAR-α) and
stimulate the peroxisome proliferator activated receptor gamma
(PPAR-γ), which acts directly with the retinoid X receptor and
participates in regulating fatty-acid transport and oxidation
(19,20). Meanwhile, the inflammation responses
stimulated by gut-derived LPS exaggerate the alcohol-induced liver
function disorders (21), likely
worsen the lipid metabolism disturbance in the liver.
Previous studies have discovered an efficient
nutraceutical intervention via immunomodulatory probiotics and
prebiotics to prevent gut barrier disruption and modulate
intestinal microbiome balance (22-25).
To date, Bacillus subtilis (B. subtilis) and its
metabolic products, including nattokinase, carbohydrase and
protease, are widely used as food ingredients and food additives.
B. subtilis-based probiotics are increasingly being used to
maintain intestinal health, and the beneficial attributes of B.
subtilis are credited to its ability to produce antimicrobial
peptides and small extracellular effector molecules and to interact
with a host with the aid of adhesion and attachment features
(26-28).
Additionally, several important clinical trials have demonstrated
that B. subtilis-based probiotic supplementation may improve
perturbed gut microbiota and even relieve the adverse effects of
antibiotic-associated diarrhea (29,30).
In conclusion, dietary B. subtilis supplementation have
great potential to restore disordered gut microbiota; however, the
effects of dietary B. subtilis supplementation on
alcohol-induced liver injury have been seldom reported. The present
study characterized the efficacy of B. subtilis strains as
potential beneficial probiotics in alcohol-induced liver injury and
characterized the underlying mechanisms.
Materials and methods
Bacterial strains and cultures
The Bacillus strain used in the present study
was B. subtilis (CMCC 1.3358), which was commercially
available from China General Microbiological Culture Collection
Center. For experimental purposes, single colonies of the
Bacillus strain were grown in LB broth at 37˚C overnight
with shaking (200 rpm). Next, overnight cultures were diluted
(1:100) in LB broth and left to grow at 37˚C with shaking (200 rpm)
until OD600 was reached 0.5-0.6 (~2-3 h). Bacillus strains
were then pelleted by centrifugation (6,000 x g) at room
temperature for 2 min, diluted in Fresh PBS solution and mixed to
obtain the appropriate bacterial density (B. subtilis:
5x108 CFU/ml).
Animal study
The animal experiments in the present study were
approved by the Ethics and Clinical Research Committee of Nankai
University (Project IRM-DWLL-2016121). A total of 40 male C57BL/6J
mice (20-22 g; 4-6 weeks old) used in the present study were
purchased from the Beijing Hfk Bioscience Co., Ltd. Animals had a
sanitary status of SPF and were housed in regulation cages (22±1˚C,
relative humidity of 50±10% and 12-h light/dark cycle) with ad
libitum access to food and water. All the mice were subdivided
into groups (10 mice in each group): Group 1, Control (Ctrl); group
2, ethyl alcohol (EtOH) group; group 3, ethyl alcohol and B.
subtilis supplementation (EtOH+BS), group 4, ethyl alcohol and
PBS (EtOH+PBS). The alcohol-induced liver injury model was
constructed in mice using the Lieber-DeCarli diet as previously
described (31). In brief, all mice
were allowed to acclimatize to the laboratory conditions for 2 days
and to a liquid diet for 5 days, and two mice were housed in each
cage. During the acclimatization to liquid diet, prophylactic
supplementation with B. subtilis or PBS were performed
daily. Following the acclimatization, the alcohol-fed mice were fed
with a diet containing 3% (vol/vol) ethanol for 10 days, while the
Ctrl mice received an isocaloric amount of maltodextrin as pair-fed
control. The mice in the EtOH + BS group were administered
bacterial suspension in PBS (200 µl) by gavage every day. Body
weights were measured every other day. On day 16, the mice received
a single dose of ethanol via oral gavage (5 g/kg body weight),
followed by a 9 h-fast, and were sacrificed by anesthesia with
isoflurane (4%) prior to excision of tissue samples.
Intestinal permeability assays
To determine the intestinal permeability,
fluorescein isothiocyanate (FITC)-dextran (4 kDa; Sigma-Aldrich;
Merck KGaA) was orally administered (600 mg/kg body weight) to mice
4 h before sacrifice. Blood samples were collected quickly and
subsequently centrifuged (1,500 x g, 4˚C, 15 min) to prepare serum.
Fluorescence was recorded using a spectrophotometer (Tecan Group,
Ltd.) at an excitation wavelength of 485 nm and emission wavelength
of 528 nm.
Biochemical analysis
Serum aspartate transaminase (AST) and alanine
transaminase (ALT) were measured using the Infinity reagent (cat.
nos. TR71121 and TR70121; Thermo Fisher Scientific). Hepatic
triglyceride (TG) levels were measured using the Triglyceride
Liquid Reagents kit (cat. no. BC0625, Beijing Solarbio Science
& Technology Co., Ltd.), according to the manufacturer's
protocols. Hepatic lipid peroxidation was quantified by measuring
malondialdehyde (MDA) using Micro MDA assay kit (cat. no. BC0025,
Beijing Solarbio Science & Technology Co., Ltd.), and the
oxidative stress of liver tissues was characterized by testing the
decreased glutathione content using a reduced glutathione (GSH)
assay kit (cat. no. BC1175, Beijing Solarbio Science &
Technology Co., Ltd.). Serum LPS was determined using the Mouse
Lipopolysaccharides (LPS) ELISA kit (cat. no. CSB-E13066m,
Cusabio). The fecal IgA levels were quantified using a Mouse sIgA
ELISA kit (cat. no. CSB-E08413m, Cusabio). The expression level of
IL-6 in serum was determined using a Mouse IL-6 ELISA kit (cat. no.
SEKM-0007, Beijing Solarbio Science & Technology Co., Ltd.).
Serum levels of total cholesterol (TCH), low-density lipoprotein
cholesterol (LDL-C), and high-density lipoprotein cholesterol
(HDL-C) were detected colorimetrically by mindray 2000 M
autoanalyzer according to the manufacturer's protocols.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Total liver and colon RNA were extracted using
TRIzol reagent (Invitrogen; Thermo Fisher Scientific, Inc.), and
the total RNA concentration was quantified using the NanoPhotometer
N50 (Implen NanoPhotometers), and cDNA was synthesized at 42˚C for
2 min on an Mastercycler nexus PCR using PrimeScript RT reagent kit
with gDNA Eraser (Takara Bio, Inc.). The synthesized cDNA was
stored at -20˚C, and RT-qPCR was conducted on a
LightCycler® 96 system (Roche Diagnostics) using a TB
Green Premix Ex Taq II (Tli RNaseH Plus; Takara Bio, Inc.). The
thermocycling conditions were as follows: Initial denaturation for
10 min at 95˚C, then 40 cycles of 10 sec at 95˚C, 10 sec at 62˚C
and 10 sec at 72˚C, followed by 95˚C for 60 sec and dissociation
curve analysis. The primer sequences are listed in Table I. Relative gene expression was
normalized to the 18S and calculated by the 2-ΔΔCt
method (32).
 | Table IPrimers for reverse
transcription-quantitative polymerase chain reaction. |
Table I
Primers for reverse
transcription-quantitative polymerase chain reaction.
| Genes | Forward
primers | Reverse
primers |
|---|
| IL-6 |
CCACTTCACAAGTCGGAGGCTTA |
CCAGTTTGGTAGCATCCATCATTTC |
| MCP-1 |
CCAGCCTACTCATTGGGATCA |
CTTCTGGGCCTGCTGTTCA |
| TNF-α |
ACCCTCACACTCAGATCATCTTC |
TGGTGGTTTGCTACGACGT |
| TLR4 |
GGCATGGCATGGCTTAAACC |
CATCGGTTGATCTTGGGAGAATT |
| NF-κB |
GGGACTATGACTTGAATGCG |
ATACGCTGACCCTAGCCTG |
| Zo-1 |
GACCTTGATTTGCATGACGA |
AGGACCGTGTAATGGCAGAC |
|
Occludin |
GAAAGTCCACCTCCTTACAGA |
CGGATAAAAAGAGTACGCTGG |
| Muc2 |
GCTCGGAACTCCAGAAAGAAG |
GCCAGGGAATCGGTAGACAT |
| Reg3b |
GGAATTCGATGTCCAAAA |
CGGTCGACGTGAACTTTGCAGACATA |
| Reg3g |
TTCCTGTCCTCCATGATCAAA |
CATCCACCTCTGTTGGGTTC |
| 18s |
AGGGGAGAGCGGGTAAGAGA |
GGACAGGACTAGGCGGAACA |
Western blotting
Total protein was extracted from livers using a RIPA
buffer containing 1% protease inhibitor and 1% phenylmethylsulfonyl
fluoride (cat. no. R0010; Beijing Solarbio Science & Technology
Co., Ltd.) and was measured by the BCA assay. Total protein (50 µg
per lane) was separated by 10% SDS-polyacrylamide gel, transferred
onto a polyvinylidene difluoride membrane (Whatman plc), and
blocked with 5% skimmed milk at room temperature for 1 h. Membranes
were immunostained with primary antibodies against TLR4 (cat. no.
14358; dilution, 1:1,000; CST) at 4˚C overnight, as well as
antibody against reference protein β-actin (cat. no. K200058M;
dilution, 1:1,000; Beijing Solarbio Science & Technology Co.,
Ltd.). Following incubation with a goat anti-rabbit horseradish
peroxidase-conjugated secondary antibody (cat. no. SE134; dilution,
1:5,000; Beijing Solarbio Science & Technology Co., Ltd.) was
carried out at room temperature for 1 h. Immunoreactive proteins
were stained with ECL Western Blotting Substrate (Beijing Solarbio
Science & Technology Co., Ltd.). Images were digitalized using
the ChemiDoc™ XRS system (Bio-Rad Laboratories, Inc.). β-actin
(Beijing Solarbio Science & Technology Co., Ltd.) was used as
an internal standard.
Histopathological observation
For the histological analysis, the liver and colonic
tissues were stained with hematoxylin and eosin (H&E). In
brief, tissues were fixed in 10% formalin at room temperature for
24 h, and paraffin-embedded sections (5 µm) were stained with
hematoxylin (10 min) and eosin (30 sec) at room temperature.
DNA extraction and 16S rRNA
amplification sequencing
Total genomic DNA was extracted from 150-200 mg of
stool samples using the QIAamp PowerFecal DNA kit (Qiagen GmbH).
The hypervariable V3-V4 region (341F and 805R) of the the
prokaryotic 16S rRNA gene was amplified. Following purification,
these amplicons were equally combined and then subjected to a
sequencing library preparation according to the manufacturer's
protocols. DNA concentration and quality were determined by the
NanoPhotometer N50 (Implen NanoPhotometers) and Qubit Fluorometer
(Thermo Fisher Scientific, Inc.). Deep DNA pyrosequencing
procedures were performed on a paired-end Illumina MiSeq PE300
(2x300 bp) platform at the Novogene Corporation, according to the
manufacturer's protocols. These raw sequences were processed
following the QIIME (v1.9.1) pipeline (33), and the analysis of gut microbiota
diversity and composition of fecal samples was determined, of which
Shannon indexes and Simpson indexes were calculated with the
operational taxonomic unit (OTU) table (34). The β diversity of fecal microbiota
was assessed by principal coordinate analysis based on Bray-Curtis
distance.
Statistical analysis
All experimental results were obtained from at least
three independent times, and representative data are presented as
the mean ± standard deviation. One-way analysis of variance was
used to determine whether the groups were statistically different
(P<0.05). GraphPad Prism 7.0 (GraphPad Software, Inc.) was
applied to all statistical analysis.
Results
Dietary B. subtilis supplementation
alleviates alcohol-induced injury
To investigate the effect of the prophylactic
dietary B. subtilis supplementation on alcohol-induced
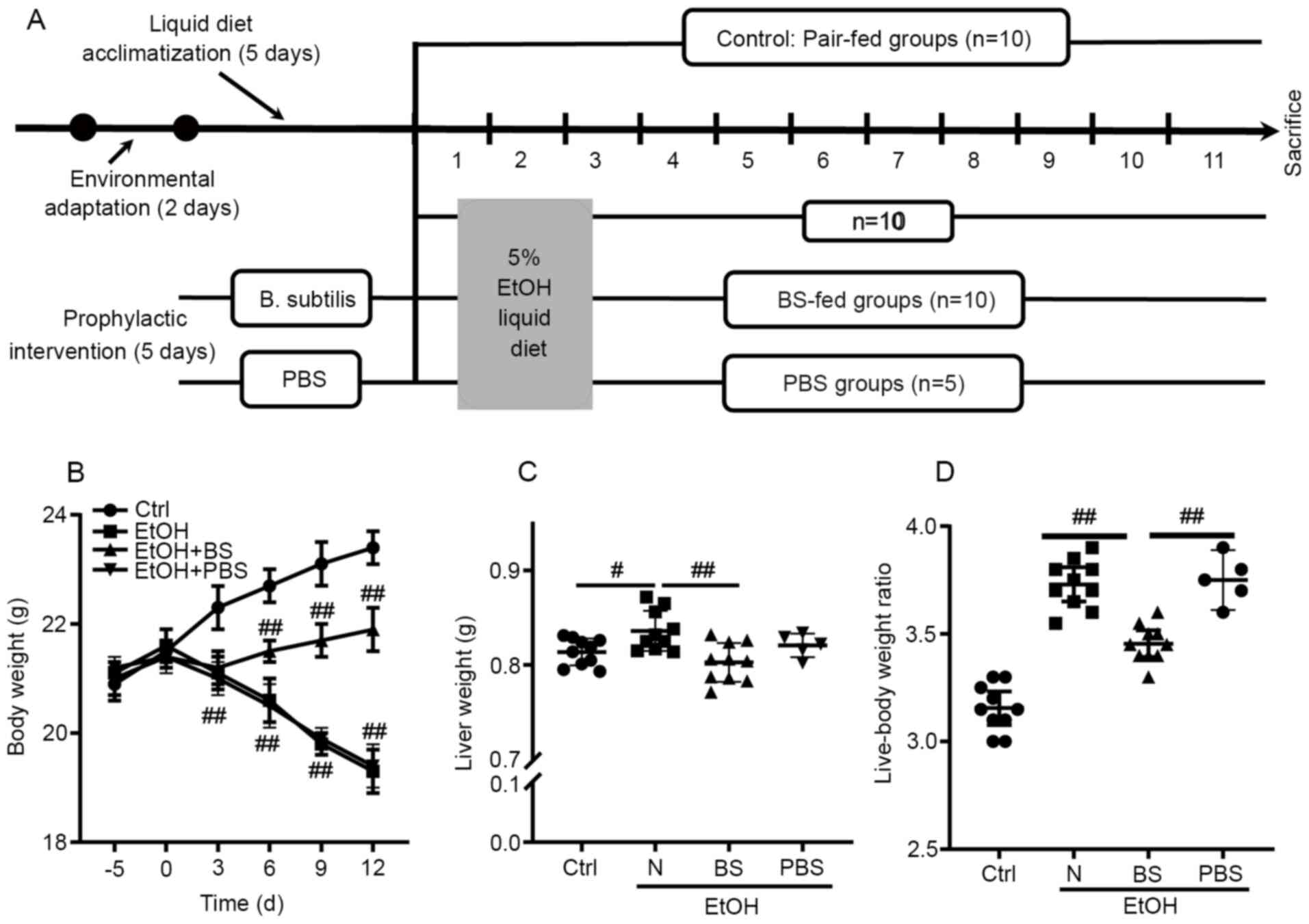
injury, a murine model was constructed as shown in Fig. 1A. Notably, alcohol feeding caused
continuous body weight loss in the EtOH group compared with the
Ctrl group; however, dietary B. subtilis supplementation
notably prevented alcohol-induced body weight loss (Fig. 1B). Following these mice being
sacrificed, it was observed that ethanol feeding significantly
increased the murine liver weight and the liver-body weight ratio
compared with Ctrl mice, and dietary B. subtilis
supplementation significantly decreased the liver weight (Fig. 1B and C). Notably, it was found that
supplementation with PBS-bacterial vehicle did not cause notable
effects in EtOH-fed mice (Fig.
1B-D); therefore, the effects of PBS supplementation in
EtOH-fed mice were excluded from further experiments. In
conclusion, dietary B. subtilis supplementation likely
alleviated alcohol-induced injury.
Dietary B. subtilis supplementation
alleviates alcohol-induced liver injury
The development of alcohol-induced liver injury
(EtOH group, Fig. 2A) was
confirmed, as indicated by notably increased levels of ALT, AST and
hepatic triglycerides compared with the Ctrl group (Fig. 2B-E). In addition, it was
investigated whether dietary B. subtilis supplementation may
relieve alcohol-induced liver injury in mice. Microscopic
morphological examination confirmed the protective effect of
dietary B. subtilis supplementation with a decrease in clear
vacuoles and neutrophil infiltration in H&E-stained images
(Fig. 2A). Dietary B.
subtilis supplementation significantly decreased hepatic
triglyceride levels in treated mice compared with EtOH-fed mice
(Fig. 2C). Additionally, the
hepatic dysfunction induced by alcohol was improved as demonstrated
by decreased serum ALT and AST levels (Fig. 2B and E), as well as the partial decrease in
serum TCH and LDL-C level (Fig.
S1). Furthermore, hepatic oxidative stress was induced by
alcohol consumption with increased levels of hepatic thiobarbituric
acid-reactive substances (TBARS) in the EtOH mice (1.53±0.07 µmol/g
liver) compared with the Ctrl mice (1.03±0.09 µmol/g liver), and
dietary B. subtilis supplementation significantly decreased
hepatic MDA levels to 1.17±0.06 µmol/g liver (Fig. 2D). GSH depletion was another trait
of ethanol-induced liver injury, and the administration of B.
subtilis notably increased the content of reduced GSH (Fig. S2). Consistent with these protective
effects, dietary B. subtilis supplementation significantly
decreased the expression of peroxisome proliferator-activated
receptor-γ (PPAR-γ) and fatty acid translocase (CD36; Fig. 2F and G), which likely ameliorated the hepatic
lipid dysfunction induced by alcohol.
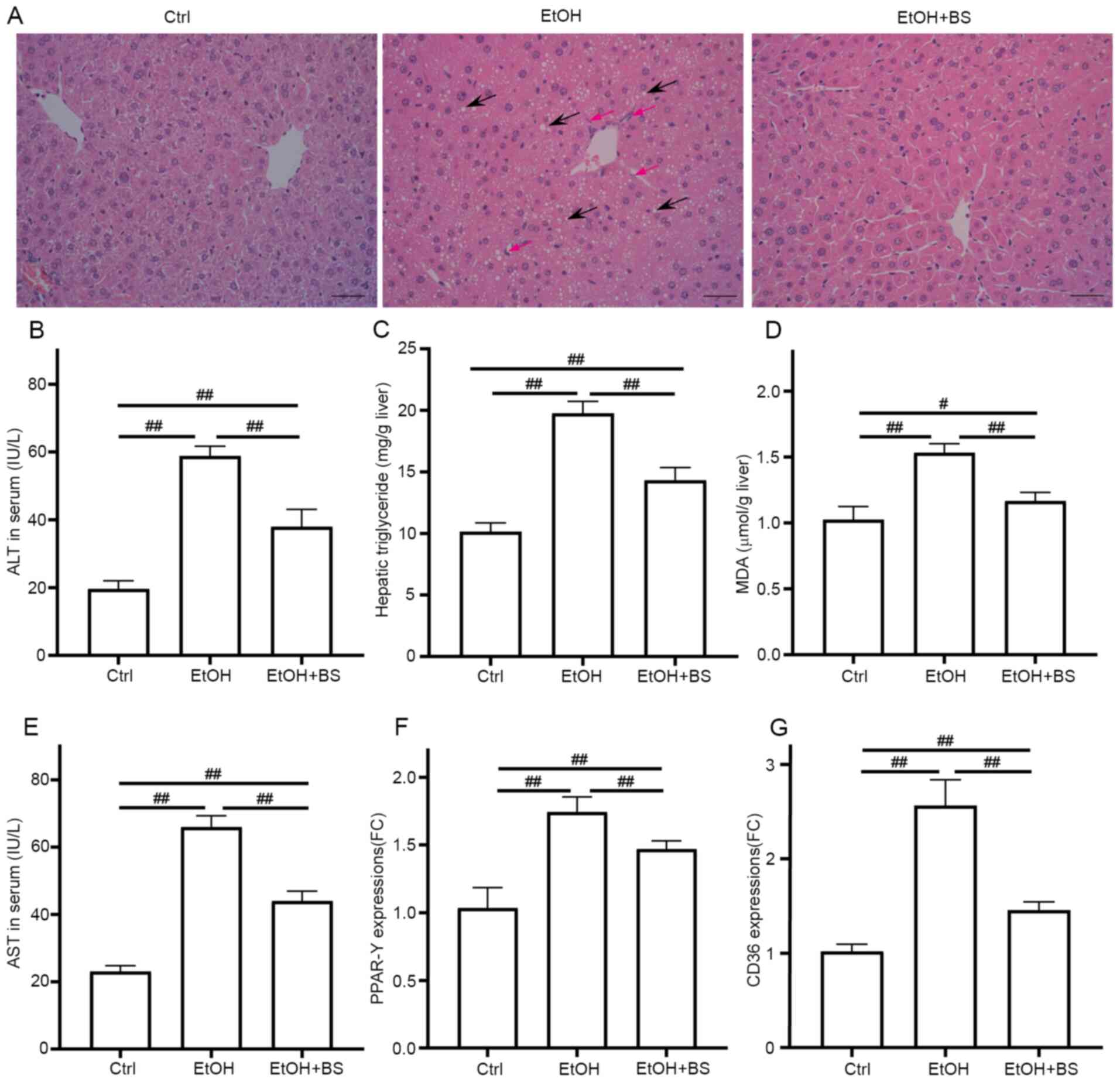 | Figure 2BS supplementation alleviates
alcohol-induced liver injury. (A) Representative photomicrographs
of hematoxylin and eosin-stained liver sections, black arrows show
the steatosis and red arrows show neutrophil infiltration
(magnification, x200). (B) Serum ALT levels. (C) Hepatic
triglyceride levels. (D) Hepatic TBARS levels. (E) Serum AST
levels. (F and G) Hepatic expression of gene PPAR-γ and
CD36, respectively. Data are presented as the mean ±
standard deviation of at least three independent experiments.
#P<0.05; ##P<0.01. BS, B.
subtilis supplementation; ALT, alanine transaminase; TBARS,
thiobarbituric acid-reactive substances; AST, aspartate
transaminase; PPAR-γ, peroxisome proliferator activated
receptor gamma; EtOH, ethyl alcohol; Ctrl, control; MDA,
malondialdehyde. |
Dietary B. subtilis supplementation
preserves the intestinal barrier
Previous studies have characterized gut barrier
dysfunction in alcohol-induced endotoxemia and liver damage
(35-37),
and disordered intestinal integrity may be due to the direct injury
induced by alcohol and indirect inflammatory reactions (38,39).
In the present study, alcohol consumption resulted in a significant
increase in intestinal permeability. Specifically, serum FITC
concentrations increased to 1,183±76.18 ng/ml from 439.2±23.53
ng/ml in Ctrl mice (Fig. 3A), and
evidence of colon damage was noted through microscopic
morphological examination of H&E-stained samples (Fig. 3B). However, dietary B.
subtilis supplementation significantly restored intestinal
permeability to a certain degree, decreasing FITC concentrations to
786.8±45.8 ng/ml (Fig. 3A). In
addition, the damaged gut barrier resulted in the translocation of
LPS from lumen to blood, and dietary B. subtilis
supplementation significantly decreased LPS levels to 57.43±3.35
ng/ml from 88.04±7.6 ng/ml in EtOH mice (Fig. 3C). Consistent with amelioration of
intestinal barrier dysfunction, dietary B. subtilis
supplementation significantly restored Muc2, Zo-1 and
occludin expression levels in the colon (Fig. 3D). Additionally, alcohol consumption
notably damaged immunoglobulin A (IgA) production and secretion
with fecal IgA levels decreased by 24% in the EtOH group compared
with the Ctrl group, and dietary B. subtilis supplementation
significantly restored the level of IgA secretion (Fig. 3E). Furthermore, dietary B.
subtilis supplementation notably reversed the alcohol-induced
decrease in Reg3g and Reg3b mRNA expression to levels
equivalent to those noted in the Ctrl group (Fig. 3D), thereby improving intestinal
barrier function.
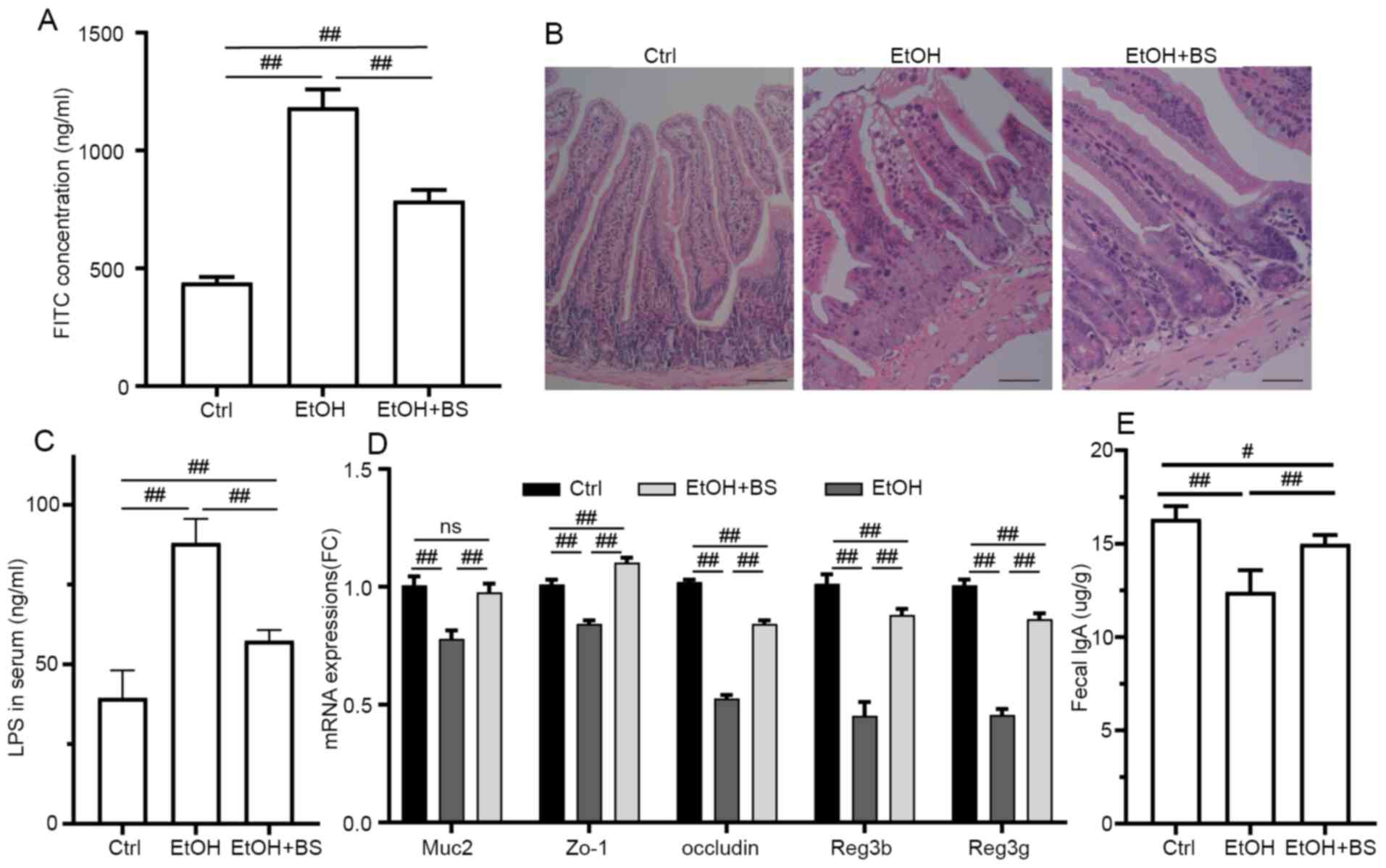 | Figure 3BS supplementation improves
alcohol-induced colonic injury. (A) Plasma concentration of
FITC-dextran (administered via oral gavage 4 h prior to
euthanasia). (B) Representative photomicrographs of hematoxylin and
eosin-stained colonic sections (magnification, x200). (C) Serum LPS
levels. (D) Colonic expression of Muc2, Zo-1,
occludin, Reg3b and Reg3g genes, respectively.
(E) Fecal IgA levels. Data are presented as the mean ± standard
deviation of at least three independent experiments.
#P<0.05; ##P<0.01. BS, B.
subtilis supplementation; LPS, lipopolysaccharide; IgA,
immunoglobulin A; EtOH, ethyl alcohol; Ctrl, control. |
Dietary B. subtilis supplementation
ameliorates alcohol-induced liver inflammation via the TLR4
pathway
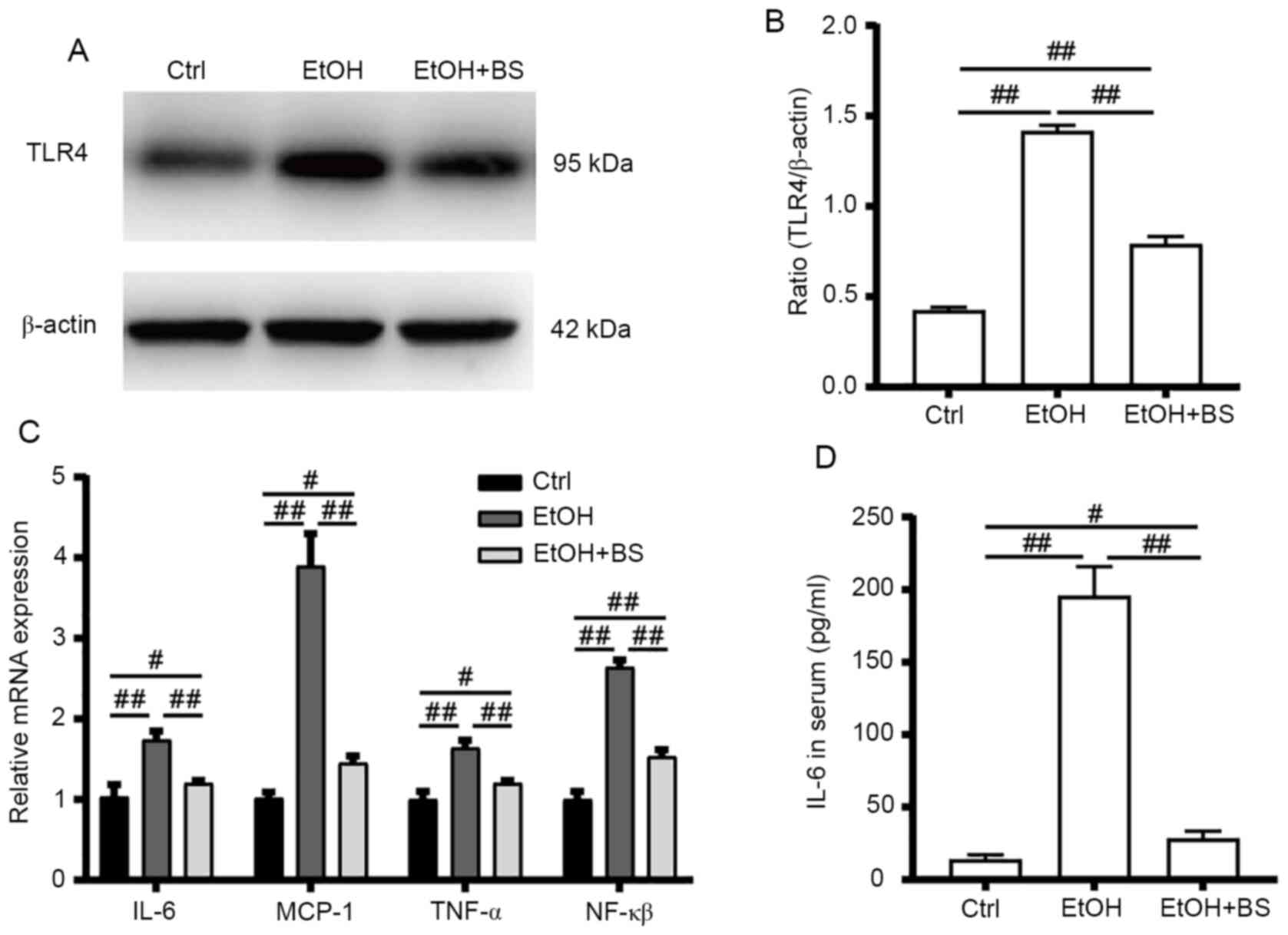
Alcohol consumption increases gut bacteria-derived
PAMPs (e.g., LPS) in the bloodstream, and PAMPs trigger hepatic
inflammation, which contributes toward the development of
alcohol-induced liver diseases (21). In the present study, alcohol
consumption increased LPS levels in the serum, which stimulated
TLR4 to induce the release of critical proinflammatory cytokines
that are required to activate potent immune responses.
Alcohol-induced increases in TLR4 protein levels were accompanied
by significant upregulation of the mRNA levels of TLR4 target genes
(Fig. 4A-C), including
NF-κB, tumor necrosis factor alpha (TNF-α),
IL-6 and monocyte chemoattractant protein-1 (MCP-1),
indicating activation of the alcohol-induced hepatic TLR4 pathway.
However, dietary B. subtilis supplementation significantly
decreased hepatic TLR4 protein levels by 53% (P<0.01) with
concomitant significant decreases in TLR4-regulated mRNA levels of
NF-κB, TNF-α, IL-6 and MCP-1
(P<0.01) in the treated group, compared with the EtOH group
(Fig. 4C).
Dietary B. subtilis supplementation
improves alcohol-induced gut microbial dysbiosis
It has been reported that dysbiosis of the gut
microbiome and metabolome is associated with the progression of
alcohol-induced liver injury (5,40,41).
The present study characterized the protective effects of dietary
B. subtilis supplementation on alcohol-induced perturbation
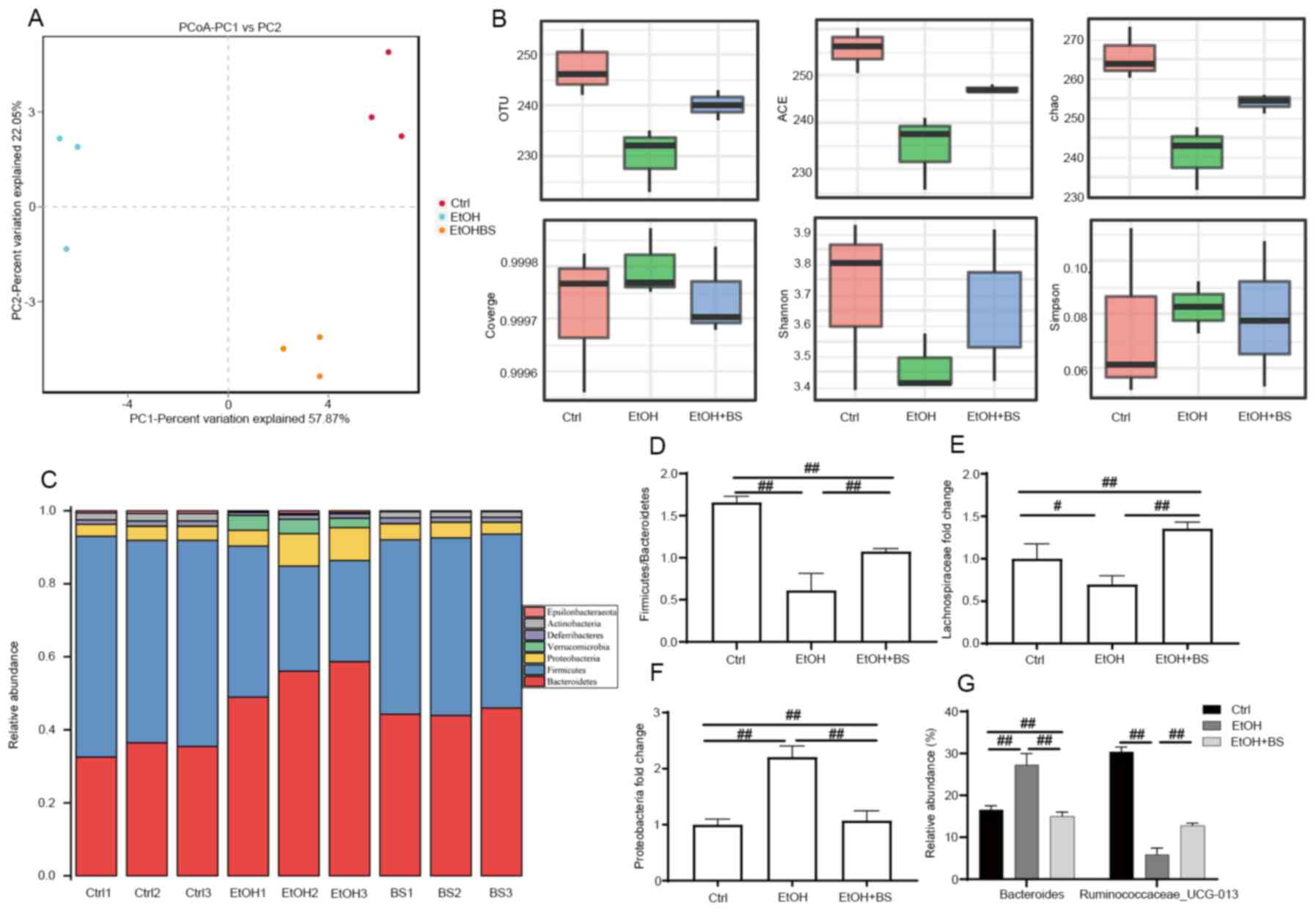
of intestinal microbiota composition. OTU-based principal
coordinate analysis revealed that the gut microbiota structures of
each tested group exhibited individual variations and that the
experimental groups significantly differed from the Ctrl group
(Fig. 5A). The diversity of gut
microbiota in EtOH-fed mice compared with Ctrl mice was
significantly decreased as evidenced by decreased Shannon indexes
and increased Simpson indexes; however, dietary B. subtilis
supplementation reversibly increased microbiota diversity (Fig. 5B). Analysis of the gut microbiota at
phylum levels revealed a significantly altered composition of gut
microbiota in Ctrl, EtOH and EtOH + BS groups (Fig. 5C). The phyla Firmicutes,
Bacteroidetes and Proteobacteria dominated the fecal microbial
communities (Fig. 5C), and alcohol
consumption significantly increased the abundance of Proteobacteria
and decreased that of Firmicutes (Fig.
5D-F); however, dietary B. subtilis supplementation
markedly decreased the level of Proteobacteria to that noted in the
Ctrl group (Fig. 5F). Notably,
further analysis at the family level revealed a marked decrease in
Lachnospiraceae in EtOH-fed mice (Fig.
5E), while dietary B. subtilis supplementation decreased
this trend. Members of the Lachnospiraceae family are important
butyrate producers that reside in the intestinal microbiota
(42). Furthermore, in the EtOH
groups, the relative increase in Bacteroide abundance and
decrease in Ruminococcaceae_UCG-013 abundance at the species
level were reversed by dietary B. subtilis supplementation
(Fig. 5G). In summary, dietary
B. subtilis supplementation partly alleviated
alcohol-induced intestinal microbiota dysbiosis.
Discussion
Changes in genetic, environmental and dietary
factors, as well as alcohol consumption, contribute toward
intestinal dysbiosis and even severe liver diseases via the
gut-liver axis (43,44). Furthermore, the gut-liver axis
facilitates close functional and bidirectional communication
between the intestine and the liver, which serves an important role
in hepatic pathogenesis and therapeutic targets (45,46).
Although B. subtilis-based probiotic supplementation may
improve perturbed gut microbiota and disordered intestinal
barriers, the protective effects of dietary B. subtilis
supplementation on hepatic pathogenesis remain unclear.
The robust intestinal barrier serves a vital role by
spatially compartmentalizing bacteria in the lumen through the
secretion of mucus and is fortified by the production of SIgA and
AMPs (47). Within the gut lumen,
SIgA interacts with various intestinal antigens, including
self-antigens and the intestinal microbiota and limits the access
of intestinal antigens to blood circulation, thereby influencing
the composition of intestinal microbiota (48,49).
Alcohol consumption notably inhibits the excretion of SIgA, thereby
resulting in the overgrowth of opportunistic pathogenic enteric
bacteria. Dietary B. subtilis supplementation quickly
depletes free oxygen present in the intestine and produces
antibacterial substances, which constrains pathogen overgrowth and
restores gut microbiota homeostasis. Additionally, multiple AMPs
(defensins, lysozymes and C-type lectins) are produced by
epithelial cells of intestinal tissues and contribute toward the
host defence against microorganisms via diverse innate immune
responses (50). Among these AMPs,
REG3 lectins (particularly REG3b and REG3g) are abundantly
expressed in enterocytes and exert bactericidal activity as a host
defence mechanism in the gut (51).
The expression of REG3 lectins is inhibited by various factors,
including antibiotics, alcohol and a high-fat diet (52,53).
By contrast, supplementation with specific probiotics enhances the
expression of Reg3g, and this finding is confirmed in the
present study by the partial restoration of Reg3b and
Reg3g expression following dietary B. subtilis
supplementation.
Efforts investigating potential therapies for
alcohol-induced liver injury have been ongoing for decades, and
dietary probiotic or prebiotic supplementation is a major strategy.
As stated above, alcohol-induced liver injury is associated with
gut barrier dysfunction. Furthermore, animal studies have
demonstrated that dietary B. subtilis supplementation
decreased the LPS translocation and abrogated the endotoxin
signalling cascade, thereby attenuating alcohol-induced hepatic
cytokine production, inflammatory responses and liver damage.
Probiotics prevent and ameliorate the course of digestive disorders
and may be of interest as co-adjuvants in the treatment of
metabolic disorders, including obesity, ALD and non-alcoholic fatty
liver disease (54,55). However, the mechanisms of action of
probiotics are diverse and strain specific. In general,
supplementation with dietary probiotics normalizes perturbed
intestinal microbial communities, competitively excludes enteric
pathogens and bacteriocin production, and suppresses intestinal
inflammation via the downregulation of TLR expression in
enterocytes (56). Probiotics are
safely tolerated, and several probiotics, including
Lactobacillus GG, L. acidophilus, L.
bulgaricus, B. bifidum, B. longum and
Streptococcus thermophilus, have been demonstrated to be
effective in the context of alcohol-induced liver injury (57-59).
It is well recognized that probiotics supplementation may primarily
ameliorate alcohol-induced intestinal dysbiosis.
Clinical and mouse model studies have revealed that
alcohol causes hepatocellular damage through oxidative stress and
lipids metabolism disorder (16,60).
Specifically, alcohol or its metabolites directly disturbs the
redox homeostasis in liver, resulting in overproduction of lipid
peroxidation end-products, including MDA. Therefore, the
restoration of liver function is partially due to the decrease in
oxidative stress (MDA) and the increases in antioxidant responses
(GSH) in livers, which may be mediated by the nuclear factor
erythroid 2-related factor 2 (NRF2) (61,62).
Additionally, the administration of B. subtilis
significantly reduces the gene expression of CD36 and
PPAR-γ, which likely restores the efficient transport of
triglycerides in liver. In this context, the suppression of fatty
acid oxidation is likely alleviated via the transcription factor
SREBP-1c and PPAR-α, which may be able to decrease the accumulation
of triglycerides in the liver and participate in preventing
mitochondrial dysfunction (63,64).
The results of the present study clearly
demonstrated that the prophylactic intervention of B.
subtilis partially restored the gut microbiota homeostasis and
protected the intestinal tract against alcohol abuse, and the
intact intestinal barrier decreased the translocation of bacterial
endotoxin (LPS), which serve a crucial role in alleviating the
hepatic inflammation via TLR4 signaling pathways. The present study
provides the basis for future evaluations of the nutritional
application of B. subtilis in the prevention and management
of clinical alcohol-induced liver injury and pave the way for
developing B. subtilis-based probiotics as the food
additives.
Supplementary Material
BS supplementation alleviates
alcohol-induced lipid disorders in mice. (A) The serum TCH level.
(B) The serum HDL-C level. (C) The serum LDL-C level.
##P<0.01. Ctrl, control; EtOH, ethyl alcohol; BS, B.
subtilis supplementation; TCH, total cholesterol; HDL-C,
high density lipoprotein cholesterol; LDL-C, low density
lipoprotein cholesterol.
BS supplementation improves the
reduced GSH level in liver tissues. The reduced GSH levels in liver
tissues. ##P<0.01. Ctrl, control; EtOH, ethyl
alcohol; BS, B. subtilis supplementation; GSH,
glutathione.
Acknowledgements
Not applicable.
Funding
Funding: The present study was supported by the Affiliated
Hospital of Weifang Medical University, the Weifang Medical
University PhD Startup Fund (grant no. 2017 BSQD39), the National
Natural Science Foundation of China (grant no. 41907362) and the
Project Funded by China Postdoctoral Science Foundation (grant no.
2019M651016).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
PZ and XZ designed the research; PZ, MeZ, ZY, MoZ,
WL, and NC performed research; PZ, YD and CC analyzed the data; PZ
and XZ wrote the manuscript and confirmed the authenticity of all
the raw data. All authors reviewed and approved the final
manuscript.
Ethics approval and consent to
participate
The animal experiments in the present study were
approved by the Ethics and Clinical Research Committee of Nankai
University (Project IRM-DWLL-2016121).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
World Health Organization (WHO): Global
status report on alcohol and health 2018. WHO, Geneva, Switzerland,
2018.
|
|
2
|
Vassallo G, Mirijello A, Ferrulli A,
Antonelli M, Landolfi R, Gasbarrini A and Addolorato G: Review
article: Alcohol and gut microbiota-the possible role of gut
microbiota modulation in the treatment of alcoholic liver disease.
Aliment Pharmacol Ther. 41:917–927. 2015.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Avila MA, Dufour JF, Gerbes AL, Zoulim F,
Bataller R, Burra P, Cortez-Pinto H, Gao B, Gilmore I, Mathurin P,
et al: Recent advances in alcohol-related liver disease (ALD):
Summary of a gut round table meeting. Gut. 69:764–780.
2020.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Bajaj JS: Alcohol, liver disease and the
gut microbiota. Nat Rev Gastroenterol Hepatol. 16:235–246.
2019.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Zhong W and Zhou Z: Alterations of the gut
microbiome and metabolome in alcoholic liver disease. World J
Gastrointest Pathophysiol. 5:514–522. 2014.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Cai X, Bao L, Wang N, Ren J, Chen Q, Xu M,
Li D, Mao R and Li Y: Dietary nucleotides protect against alcoholic
liver injury by attenuating inflammation and regulating gut
microbiota in rats. Food Funct. 7:2898–2908. 2016.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Chen J, Xuan YH, Luo MX, Ni XG, Ling LQ,
Hu SJ, Chen JQ, Xu JY, Jiang LY, Si WZ, et al: Kaempferol
alleviates acute alcoholic liver injury in mice by regulating
intestinal tight junction proteins and butyrate receptors and
transporters. Toxicology. 429(152338)2020.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Zhou Z and Zhong W: Targeting the gut
barrier for the treatment of alcoholic liver disease. Liver Res.
1:197–207. 2017.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Jiang Y, Chen B, Duan C, Sun B, Yang J and
Yang S: Multigene editing in the escherichia coli genome via the
CRISPR-Cas9 system. Appl Environ Microbiol. 81:2506–2514.
2015.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Addolorato G, Ponziani FR, Dionisi T,
Mosoni C, Vassallo GA, Sestito L, Petito V, Picca A, Marzetti E,
Tarli C, et al: Gut microbiota compositional and functional
fingerprint in patients with alcohol use disorder and
alcohol-associated liver disease. Liver Int. 40:878–888.
2020.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Parlesak A, Schafer C, Schutz T, Bode JC
and Bode C: Increased intestinal permeability to macromolecules and
endotoxemia in patients with chronic alcohol abuse in different
stages of alcohol-induced liver disease. J Hepatol. 32:742–747.
2000.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Hoque R, Farooq A, Ghani A, Gorelick F and
Mehal WZ: Lactate reduces liver and pancreatic injury in Toll-like
receptor- and inflammasome-mediated inflammation via GPR81-mediated
suppression of innate immunity. Gastroenterology. 146:1763–1774.
2014.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Kageyama S, Nakamura K, Fujii T, Ke B,
Sosa RA, Reed EF, Datta N, Zarrinpar A, Busuttil RW and
Kupiec-Weglinski JW: Recombinant relaxin protects liver transplants
from ischemia damage by hepatocyte glucocorticoid receptor: From
bench-to-bedside. Hepatology. 68:258–273. 2018.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Cui K, Yan G, Xu C, Chen Y, Wang J, Zhou
R, Bai L, Lian Z, Wei H, Sun R and Tian Z: Invariant NKT cells
promote alcohol-induced steatohepatitis through interleukin-1β in
mice. J Hepatol. 62:1311–1318. 2015.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Wieser V, Tymoszuk P, Adolph TE, Grander
C, Grabherr F, Enrich B, Pfister A, Lichtmanegger L, Gerner R,
Drach M, et al: Lipocalin 2 drives neutrophilic inflammation in
alcoholic liver disease. J Hepatol. 64:872–880. 2016.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Osna NA, Donohue TM Jr and Kharbanda KK:
Alcoholic liver disease: Pathogenesis and current management.
Alcohol Res. 38:147–161. 2017.PubMed/NCBI
|
|
17
|
You Y, Li WZ, Zhang S, Hu B, Li YX, Li HD,
Tang HH, Li QW, Guan YY, Liu LX, et al: SNX10 mediates
alcohol-induced liver injury and steatosis by regulating the
activation of chaperone-mediated autophagy. J Hepatol. 69:129–141.
2018.PubMed/NCBI View Article : Google Scholar
|
|
18
|
You M and Arteel GE: Effect of ethanol on
lipid metabolism. J Hepatol. 70:237–248. 2019.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Valenzuela R and Videla LA: The importance
of the long-chain polyunsaturated fatty acid n-6/n-3 ratio in
development of non-alcoholic fatty liver associated with obesity.
Food Funct. 2:644–648. 2011.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Zhang W, Sun Q, Zhong W, Sun X and Zhou Z:
Hepatic peroxisome proliferator-activated receptor gamma signaling
contributes to alcohol-induced hepatic steatosis and inflammation
in mice. Alcohol Clin Exp Res. 40:988–999. 2016.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Mandrekar P and Szabo G: Signalling
pathways in alcohol-induced liver inflammation. J Hepatol.
50:1258–1266. 2009.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Shimizu H, Masujima Y, Ushiroda C,
Mizushima R, Taira S, Ohue-Kitano R and Kimura I: Dietary
short-chain fatty acid intake improves the hepatic metabolic
condition via FFAR3. Sci Rep. 9:16574. 2019.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Makki K, Deehan EC, Walter J and Bäckhed
F: The impact of dietary fiber on gut microbiota in host health and
disease. Cell Host Microbe. 23:705–715. 2018.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Tsai YL, Lin TL, Chang CJ, Wu TR, Lai WF,
Lu CC and Lai HC: Probiotics, prebiotics and amelioration of
diseases. J Biomed Sci. 26(3)2019.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Vétizou M, Pitt JM, Daillère R, Lepage P,
Waldschmitt N, Flament C, Rusakiewicz S, Routy B, Roberti MP, Duong
CP, et al: Anticancer immunotherapy by CTLA-4 blockade relies on
the gut microbiota. Science. 350:1079–1084. 2015.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Khochamit N, Siripornadulsil S, Sukon P
and Siripornadulsil W: Antibacterial activity and
genotypic-phenotypic characteristics of bacteriocin-producing
Bacillus subtilis KKU213: Potential as a probiotic strain.
Microbiol Res. 170:36–50. 2015.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Compaoré CS, Nielsen DS, Ouoba LII, Berner
TS, Nielsen KF, Sawadogo-Lingani H, Diawara B, Ouédraogo GA,
Jakobsen M and Thorsen L: Co-production of surfactin and a novel
bacteriocin by Bacillus subtilis subsp. subtilis H4
isolated from Bikalga, an African alkaline Hibiscus
sabdariffa seed fermented condiment. Int J Food Microbiol.
162:297–307. 2013.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Elshaghabee FMF, Rokana N, Gulhane RD,
Sharma C and Panwar H: Bacillus as potential probiotics: Status,
concerns, and future perspectives. Front Microbiol.
8(1490)2017.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Horosheva TV, Vodyanoy V and Sorokulova I:
Efficacy of Bacillus probiotics in prevention of
antibiotic-associated diarrhoea: A randomized, double-blind,
placebo-controlled clinical trial. JMM Case Rep. 1:2014.
|
|
30
|
Hickson M: Probiotics in the prevention of
antibiotic-associated diarrhoea and Clostridium difficile
infection. Therap Adv Gastroenterol. 4:185–197. 2011.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Bertola A, Mathews S, Ki SH, Wang H and
Gao B: Mouse model of chronic and binge ethanol feeding (the NIAAA
model). Nat Protoc. 8:627–637. 2013.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408.
2001.PubMed/NCBI View Article : Google Scholar
|
|
33
|
Caporaso JG, Kuczynski J, Stombaugh J,
Bittinger K, Bushman FD, Costello EK, Fierer N, Peña AG, Goodrich
JK, Gordon JI, et al: QIIME allows analysis of high-throughput
community sequencing data. Nat Methods. 7:335–336. 2010.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Hill TC, Walsh KA, Harris JA and Moffett
BF: Using ecological diversity measures with bacterial communities.
FEMS Microbiol Ecol. 43:1–11. 2003.PubMed/NCBI View Article : Google Scholar
|
|
35
|
Cho YE and Song BJ: Pomegranate prevents
binge alcohol-induced gut leakiness and hepatic inflammation by
suppressing oxidative and nitrative stress. Redox Biol. 18:266–278.
2018.PubMed/NCBI View Article : Google Scholar
|
|
36
|
Shao T, Zhao C, Li F, Gu Z, Liu L, Zhang
L, Wang Y, He L, Liu Y, Liu Q, et al: Intestinal HIF-1α deletion
exacerbates alcoholic liver disease by inducing intestinal
dysbiosis and barrier dysfunction. J Hepatol. 69:886–895.
2018.PubMed/NCBI View Article : Google Scholar
|
|
37
|
Antón M, Rodríguez-González A, Ballesta A,
González N, Del Pozo A, de Fonseca FR, Gómez-Lus ML, Leza JC,
García-Bueno B, Caso JR and Orio L: Alcohol binge disrupts the rat
intestinal barrier: The partial protective role of
oleoylethanolamide. Br J Pharmacol. 175:4464–4479. 2018.PubMed/NCBI View Article : Google Scholar
|
|
38
|
Ambade A, Lowe P, Kodys K, Catalano D,
Gyongyosi B, Cho Y, Iracheta-Vellve A, Adejumo A, Saha B, Calenda
C, et al: Pharmacological inhibition of CCR2/5 signaling prevents
and reverses alcohol-induced liver damage, steatosis, and
inflammation in mice. Hepatology. 69:1105–1121. 2019.PubMed/NCBI View Article : Google Scholar
|
|
39
|
Ferrere G, Wrzosek L, Cailleux F, Turpin
W, Puchois V, Spatz M, Ciocan D, Rainteau D, Humbert L, Hugot C, et
al: Fecal microbiota manipulation prevents dysbiosis and
alcohol-induced liver injury in mice. J Hepatol. 66:806–815.
2017.PubMed/NCBI View Article : Google Scholar
|
|
40
|
Arab JP, Arrese M and Shah VH: Gut
microbiota in non-alcoholic fatty liver disease and alcohol-related
liver disease: Current concepts and perspectives. Hepatol Res.
50:407–418. 2020.PubMed/NCBI View Article : Google Scholar
|
|
41
|
Dubinkina VB, Tyakht AV, Odintsova VY,
Yarygin KS, Kovarsky BA, Pavlenko AV, Ischenko DS, Popenko AS,
Alexeev DG, Taraskina AY, et al: Links of gut microbiota
composition with alcohol dependence syndrome and alcoholic liver
disease. Microbiome. 5(141)2017.PubMed/NCBI View Article : Google Scholar
|
|
42
|
Vital M, Penton CR, Wang Q, Young VB,
Antonopoulos DA, Sogin ML, Morrison HG, Raffals L, Chang EB,
Huffnagle GB, et al: A gene-targeted approach to investigate the
intestinal butyrate-producing bacterialcommunity. Microbiome.
1(8)2013.PubMed/NCBI View Article : Google Scholar
|
|
43
|
Yan AW, Fouts DE, Brandl J, Stärkel P,
Torralba M, Schott E, Tsukamoto H, Nelson KE, Brenner DA and
Schnabl B: Enteric dysbiosis associated with a mouse model of
alcoholic liver disease. Hepatology. 53:96–105. 2011.PubMed/NCBI View Article : Google Scholar
|
|
44
|
Mutlu EA, Gillevet PM, Rangwala H,
Sikaroodi M, Naqvi A, Engen PA, Kwasny M, Lau CK and Keshavarzian
A: Colonic microbiome is altered in alcoholism. Am J Physiol
Gastrointest Liver Physiol. 302:G966–G978. 2012.PubMed/NCBI View Article : Google Scholar
|
|
45
|
Wang L, Fouts DE, Stärkel P, Hartmann P,
Chen P, Llorente C, DePew J, Moncera K, Ho SB, Brenner DA, et al:
Intestinal REG3 lectins protect against alcoholic steatohepatitis
by reducing mucosa-associated microbiota and preventing bacterial
translocation. Cell Host Microbe. 19:227–239. 2016.PubMed/NCBI View Article : Google Scholar
|
|
46
|
Tilg H, Cani PD and Mayer EA: Gut
microbiome and liver diseases. Gut. 65:2035–2044. 2016.PubMed/NCBI View Article : Google Scholar
|
|
47
|
Wells JM, Brummer RJ, Derrien M, MacDonald
TT, Troost F, Cani PD, Theodorou V, Dekker J, Méheust A, de Vos WM,
et al: Homeostasis of the gut barrier and potential biomarkers. Am
J Physiol Gastrointest Liver Physiol. 312:G171–G193.
2017.PubMed/NCBI View Article : Google Scholar
|
|
48
|
Lindner C, Thomsen I, Wahl B, Ugur M,
Sethi MK, Friedrichsen M, Smoczek A, Ott S, Baumann U, Suerbaum S,
et al: Diversification of memory B cells drives the continuous
adaptation of secretory antibodies to gut microbiota. Nat Immunol.
16:880–888. 2015.PubMed/NCBI View Article : Google Scholar
|
|
49
|
Rogier EW, Frantz AL, Bruno ME, Wedlund L,
Cohen DA, Stromberg AJ and Kaetzel CS: Secretory antibodies in
breast milk promote long-term intestinal homeostasis by regulating
the gut microbiota and host gene expression. Proc Natl Acad Sci
USA. 111:3074–3079. 2014.PubMed/NCBI View Article : Google Scholar
|
|
50
|
Brown GD, Willment JA and Whitehead L:
C-type lectins in immunity and homeostasis. Nat Rev Immunol.
18:374–389. 2018.PubMed/NCBI View Article : Google Scholar
|
|
51
|
Seo GY, Giles DA and Kronenberg M: The
role of innate lymphoid cells in response to microbes at mucosal
surfaces. Mucosal Immunol. 13:399–412. 2020.PubMed/NCBI View Article : Google Scholar
|
|
52
|
Guerville M, Leroy A, Sinquin A,
Laugerette F, Michalski MC and Boudry G: Western-diet consumption
induces alteration of barrier function mechanisms in the ileum that
correlates with metabolic endotoxemia in rats. Am J Physiol
Endocrinol Metab. 313:E107–E120. 2017.PubMed/NCBI View Article : Google Scholar
|
|
53
|
van Ampting MT, Loonen LM, Schonewille AJ,
Konings I, Vink C, Iovanna J, Chamaillard M, Dekker J, van der Meer
R, Wells JM and Bovee-Oudenhoven IMJ: Intestinally secreted C-type
lectin Reg3b attenuates salmonellosis but not listeriosis in mice.
Infect Immun. 80:1115–1120. 2012.PubMed/NCBI View Article : Google Scholar
|
|
54
|
Eslamparast T, Eghtesad S, Hekmatdoost A
and Poustchi H: Probiotics and nonalcoholic fatty liver disease.
Middle East J Dig Dis. 5:129–136. 2013.PubMed/NCBI
|
|
55
|
Koutnikova H, Genser B, Monteiro-Sepulveda
M, Faurie MJ, Rizkalla S, Schrezenmeir J and Clément K: Impact of
bacterial probiotics on obesity, diabetes and non-alcoholic fatty
liver disease related variables: A systematic review and
meta-analysis of randomised controlled trials. BMJ Open.
9(e017995)2019.PubMed/NCBI View Article : Google Scholar
|
|
56
|
Hong M, Han DH, Hong J, Kim DJ and Suk KT:
Are probiotics effective in targeting alcoholic liver diseases?
Probiotics Antimicrob Proteins. 11:335–347. 2019.PubMed/NCBI View Article : Google Scholar
|
|
57
|
Wang J, Dong X, Shao Y, Guo H, Pan L, Hui
W, Kwok LY, Zhang H and Zhang W: Genome adaptive evolution of
Lactobacillus casei under long-term antibiotic selection
pressures. BMC Genomics. 18(320)2017.PubMed/NCBI View Article : Google Scholar
|
|
58
|
Koga H, Tamiya Y, Mitsuyama K, Ishibashi
M, Matsumoto S, Imaoka A, Hara T, Nakano M, Ooeda K, Umezaki Y and
Sata M: Probiotics promote rapid-turnover protein production by
restoring gut flora in patients with alcoholic liver cirrhosis.
Hepatol Int. 7:767–774. 2013.PubMed/NCBI View Article : Google Scholar
|
|
59
|
Forsyth CB, Farhadi A, Jakate SM, Tang Y,
Shaikh M and Keshavarzian A: Lactobacillus GG treatment ameliorates
alcohol-induced intestinal oxidative stress, gut leakiness, and
liver injury in a rat model of alcoholic steatohepatitis. Alcohol.
43:163–172. 2009.PubMed/NCBI View Article : Google Scholar
|
|
60
|
Gao B, Xu MJ, Bertola A, Wang H, Zhou Z
and Liangpunsakul S: Animal models of alcoholic liver disease:
Pathogenesis and clinical relevance. Gene Expr. 17:173–186.
2017.PubMed/NCBI View Article : Google Scholar
|
|
61
|
Barrera C, Valenzuela R, Rincón M,
Espinosa A, Echeverria F, Romero N, Gonzalez-Mañan D and Videla LA:
Molecular mechanisms related to the hepatoprotective effects of
antioxidant-rich extra virgin olive oil supplementation in rats
subjected to short-term iron administration. Free Radic Biol Med.
126:313–321. 2018.PubMed/NCBI View Article : Google Scholar
|
|
62
|
Valenzuela R, Illesca P, Echeverría F,
Espinosa A, Rincón-Cervera MA, Ortiz M, Hernandez-Rodas MC,
Valenzuela A and Videla LA: Molecular adaptations underlying the
beneficial effects of hydroxytyrosol in the pathogenic alterations
induced by a high-fat diet in mouse liver: PPAR-α and Nrf2
activation, and NF-κB down-regulation. Food Funct. 8:1526–1537.
2017.PubMed/NCBI View Article : Google Scholar
|
|
63
|
Ortiz M, Soto-Alarcón SA, Orellana P,
Espinosa A, Campos C, López-Arana S, Rincón MA, Illesca P,
Valenzuela R and Videla LA: Suppression of high-fat diet-induced
obesity-associated liver mitochondrial dysfunction by
docosahexaenoic acid and hydroxytyrosol co-administration. Dig
Liver Dis. 52:895–904. 2020.PubMed/NCBI View Article : Google Scholar
|
|
64
|
Soto-Alarcón SA, Ortiz M, Orellana P,
Echeverría F, Bustamante A, Espinosa A, Illesca P, Gonzalez-Mañán
D, Valenzuela R and Videla LA: Docosahexaenoic acid and
hydroxytyrosol co-administration fully prevents liver steatosis and
related parameters in mice subjected to high-fat diet: A molecular
approach. Biofactors. 45:930–943. 2019.PubMed/NCBI View Article : Google Scholar
|