Introduction
The coronavirus disease 2012 (COVID-19) pandemic is
becoming an increasingly serious threat to global public health.
The causative coronavirus, severe acute respiratory syndrome
coronavirus 2 (SARS-CoV-2), primarily infects a subpopulation of
airway epithelial cells that co-express the viral entry molecules
angiotensin-converting enzyme 2 (ACE2) and transmembrane serine
protease 2 (TMPRSS2) (1-3).
Notably, several lines of evidence suggest that SARS-CoV-2 can also
infect human intestinal epithelial cells, as ACE2 and TMPRSS2 are
co-expressed in the lower gastrointestinal tract, particularly
enterocytes and progenitor cells of the ileum and colon (2-5).
Targeted infection and active replication of SARS-CoV-2 in
ACE2-expressing enterocytes have been demonstrated using human
intestinal organoids (5-8).
SARS-CoV-2 RNA has been detected in stool specimens and anal/rectal
swabs of patients with COVID-19 (9-11),
and infectious viruses have been isolated from feces of patients
(8). A considerable percentage of
patients with COVID-19 present with concurrent gastrointestinal
symptoms, such as diarrhea and abdominal pain (12-14).
These findings raise the possibility that there are adverse effects
between the enteric infection of SARS-CoV-2 and the intestinal
microbiome.
Gut microbiota serves key roles in the crosstalk
between the intestinal and respiratory tracts, which is called the
gut-lung axis, via which gut microbe-derived molecules (including
structural components, metabolites and toxins, among others)
modulate airway physiology and immunity (15-17).
Intestinal dysbiosis leads to aberrant immune tone in the airway
mucosa, which can trigger dysregulated immune responses to
respiratory viral infection (15-17).
Recent microbiome studies have revealed that patients with COVID-19
have compositional changes in the specific taxa of enteric bacteria
(18-21).
Notably, a subset of the changes correlate with the serum levels of
proinflammatory cytokines, symptom severity and fecal SARS-CoV-2
virus load, suggesting that COVID-19-related intestinal dysbiosis
is closely associated with the disease pathophysiology (19-21).
Numerous animal and clinical studies have
demonstrated that the oral intake of probiotic strains of various
lactic acid bacteria (LAB) species exhibited prophylactic and
therapeutic efficacy against infection by respiratory RNA viruses
(22-26).
Life-threatening symptoms and complications of COVID-19 are caused
by hyperinflammation owing to complex immune dysregulation
involving neutrophilia, lymphocytopenia, reduced T-cell immunity
and excessive production of inflammatory mediators (27-29).
Specific probiotic LAB strains, such as L. plantarum strain
DR7 and L. paracasei strain 8700:2, have superior
immunomodulatory and anti-inflammatory abilities against
respiratory viral infection and may therefore be suitable for
therapeutic use (30,31). On the other hand, certain
proinflammatory LAB strains are known to induce innate cytokine
changes that can trigger early antiviral immune responses and may
therefore be employed prophylactically (32,33).
Notably, previous preclinical studies and randomized controlled
trials have demonstrated that among probiotic LAB, Lactobacillus
plantarum (L. plantarum) (33-39),
Bifidobacterium longum (B. longum) (40,41)
and Lactococcus lactis ssp. Lactis (L. lactis
ssp. lactis) (42) exhibit
robust protective effects against influenza virus infection through
enhancing host innate immunity.
The genera Lactobacillus,
Bifidobacterium and Lactococcus are the most
representative LAB and have been recognized as having probiotic
properties beneficial for the human health (43,44).
L. plantarum is a Gram-positive, facultatively anaerobic,
rod-shaped bacterium with a plant origin and is distributed in the
human intestinal tract and oral cavity. It is a heterofermentative
LAB closely associated with various fermented plant foods, such as
pickles, sauerkraut and kimchi. B. longum is a
Gram-positive, obligatory anaerobic, heterofermentative bacterium
with V- or Y-shaped morphology and inhabits the human intestine
predominantly from newborns to elderly people. L. lactis
ssp. lactis is a Gram-positive, facultatively anaerobic,
spherical bacterium commonly present in raw milk and fermented
dairy products.
Qingfei Paidu decoction (QFPD), the Chinese word for
‘lung cleansing and detoxifying decoction’, is a Chinese herbal
medicine newly formulated and specifically optimized against
COVID-19, and its therapeutic use has been encouraged in the
Chinese official management guidelines (45). Clinical trials in China have
demonstrated that QFPD accelerated recovery and prevented disease
progression in mild to critical cases (46-49).
A retrospective clinical study has also indicated that QFPD
decreased the blood levels of COVID-19 biomarkers, such as
C-reactive protein, creatine kinase and lactate dehydrogenase
(50). Our previous study has
demonstrated that the pharmacological action of QFPD was associated
with the upregulation of the plasma levels of TNF-α, IL-1β, IL-18
and IL-8, which are key cytokines that mediate early innate immune
responses to viral infection (51).
Therefore, rebalancing the gut microbiome using
probiotics may be effective for the control of COVID-19. However,
to the best of our knowledge, no studies have focused on the
efficacy of probiotics in patients with COVID-19, and the rationale
for using probiotics against COVID-19 remains unclear. In addition,
there is a requirement for investigating diverse prophylactic
options owing to the frequent emergence and rapid spread of novel
SARS-CoV-2 variants carrying immune escape mutations. To explore
the immunological efficacy of probiotics for preventing COVID-19, a
single-arm, double-blind, prospective trial combined with an in
vitro cytokine response assay was conducted using L.
plantarum, B. longum and L. lactis ssp.
lactis. The innate cytokine changes induced by QFPD were
used as an indicator of the anti-COVID-19 immunomodulatory
potential of the LAB. Furthermore, the effects of LAB ingestion on
the activity of innate immune cells were examined.
Materials and methods
Subjects
Participants were recruited through the University
Hospital Medical Information Network-Clinical Trials Registry
website, Takanawa Clinic (Tokyo, Japan) website, announcements in
an e-mail newsletter and personal contacts. Individuals who met all
the following inclusion criteria were enrolled: i) Healthy adults
between the ages of 20 and 70; and ii) having negative PCR and
IgM/IgG antibodies tests for SARS-CoV-2 at study entry (no previous
and current SARS-CoV-2 infection). Chest imaging tests were not
used in the present study. Individuals were excluded from this
trial if they met any of the following exclusion criteria: i)
Pregnant; ii) breastfeeding; iii) duplicate enrollment in other
clinical trials; iv) history of infectious disease within 6 months
before the enrollment; v) current or past history of chronic
inflammatory, immune-related or neoplastic diseases; vi) history of
medicinal drug use within 6 months before the enrollment; and vii)
underlying conditions associated with higher risk of COVID-19,
including hypertension, cardiovascular disease, cerebrovascular
disease, diabetes, obesity (body mass index ≥30) (52,53),
chronic obstructive pulmonary disease and chronic kidney disease.
Therefore, the enrolled subjects had no recorded and reported
comorbidities.
Subject recruitment
In our previous study using QFPD on 18 healthy
subjects (51), the effect sizes r
obtained were 0.816 (TNF-α), 0.881 (IL-1β), 0.724 (IL-18) and 0.796
(IL-8). The average value of 0.804 was employed as an estimated
effect size for the present trial. A priori two-tailed power
analysis was conducted with a level of significance α of 0.05, a
desired power 1-β of 0.8 and the estimated effect size of 0.804,
which suggested a required total sample size of 15 individuals.
Participant recruitment took place between 27 May
2020 and 2 June 2020 at Takanawa Clinic (Tokyo, Japan). A total of
20 volunteers were screened for eligibility, indicated to be
eligible and enrolled in the present trial (Fig. 1). In vitro cytokine response
assay was performed between 7 and 9 June 2020. LAB were
administered to all the enrolled participants between 25 June and
17 August 2020, 2 of whom were excluded from the main analysis due
to no visit to Takanawa Clinic following the LAB prescription.
Consequently, 18 subjects (1 male and 17 females; age, 28-66 years;
mean age ± SD, 44.2±10.1 years) completed the intervention, and the
data were subjected to statistical analysis.
Study design
The present study comprised two sequential
experimental procedures: An in vitro cytokine response assay
and a single-arm, double-blind, prospective trial.
The optimal LAB in each subject were determined
using co-culture of the peripheral blood with each LAB.
QFPD-induced innate cytokine changes were used as an indicator to
evaluate the anti-COVID-19 immunomodulatory potential of LAB. The
QFPD-induced innate cytokine index (QICI) was defined as follows:
QICI=(TNF-α) x (IL-1β) x (IL-18) x (IL-8)/(IL-6), where brackets
represent the plasma level of the cytokine in pg/ml. IL-6 is a
critical driver of complex immune dysregulation in patients with
COVID-19 and was thus adopted as the denominator (54-56).
The LAB with the highest and lowest QICI were used
in a subsequent clinical trial to examine whether the ingested LAB
could reproduce the in vitro cytokine responses (in
vitro QICI). The trial consisted of three consecutive sessions:
i) Validation (intervention using the LAB with the highest QICI);
ii) washout; and iii) control (intervention using the LAB with the
lowest QICI) sessions. The primary outcome measure was the changes
in the plasma levels of TNF-α, IL-1β, IL-18, IL-8 and IL-6 and the
QICI after each 7-day LAB session compared with those at baseline.
The secondary outcome measure was the changes in hematological
parameters after each 7-day LAB session compared with those at
baseline.
In vitro cytokine response assay
L. plantarum SNK12 [2x1012
colony-forming unit/g (cfu/g); Bio-Lab Co., Ltd.] is a probiotic
strain with potent immunomodulatory, anti-inflammatory and
antiviral abilities (57,58). A probiotic B. longum BB536
strain (1.5x1011 cfu/g; Morinaga Milk Industry Co.,
Ltd.) has been reported to exhibit various clinical benefits, such
as anti-allergic effects (59,60),
protection against viral infection (41,61)
and modulation of gut microbiota (62-64).
L. lactis ssp. lactis demonstrates immunomodulatory
and antiviral effects through activating plasmacytoid dendritic
cells and increasing their ability to produce interferons (IFNs)
(42). The commercially available
probiotic strain LLL970 (1x1011 cfu/g; Synbio Tech,
Inc.) was used in the present study.
Live LAB cells were disrupted by vibrating them
twice at 4,600 rpm (the number of the figure-8-shaped movement of
sample tubes per minute) for 3 min at room temperature with 0.5 g
of 3.0-mm zirconia beads in a commercial bead vibrator (PS-2000;
Kurabo Industries Ltd.) followed by heating at 60˚C for 20 min.
Dead LAB were suspended at a 100 µg/ml concentration in Dulbecco's
PBS (without Ca2+ and Mg2+).
The ability of the LAB to stimulate QFPD-like
cytokine production by peripheral blood immune cells was evaluated
as described in previous studies (65-67).
Briefly, heparinized peripheral blood (0.4 ml) from each subject
was co-cultured with each killed LAB suspension (4 µg) in 1 ml
RPMI-1640 medium (DS Pharma Biomedical Co., Ltd.) supplemented with
10% fetal bovine serum (FBS; Sigma-Aldrich; Merck KGaA) in a
humidified incubator with 5% CO2 at 37˚C for 48 h.
Concentrations of TNF-α, IL-1β, IL-8 and IL-6 in culture
supernatants were measured using the V-PLEX Proinflammatory Panel 1
Human kit (cat. no. K15049D-1; Meso Scale Diagnostics, LLC) and the
concentration of IL-18 was measured using the Human IL-18 ELISA kit
(cat. no. ab215539; Abcam) according to the manufacturers'
protocols. The data were used to calculate the QICI value for the
characterization of the species with the highest or lowest
QICI.
Clinical trial
The present trial was a single-arm, double-blind,
prospective trial. Each subject was instructed to orally ingest the
live LAB with the highest QICI (1x1011 cfu/day) in the
in vitro cytokine response assay twice daily in the morning
and evening between meals for 7 days (days 1-7). After a 7-day
washout period (days 8-14), a negative control trial was conducted,
in which the LAB with the lowest QICI in the in vitro
cytokine response assay (1x1011 cfu/day) was orally
administered twice daily in the morning and evening between meals
for 7 days (days 15-21). Peripheral blood samples were obtained
from each subject on days 0, 8 and 22. Neither the subjects nor
physicians in charge were aware of the results of the in
vitro LAB assessment, prescribed LAB or their QICI properties
until the final blood sampling was completed. Concentrations of
plasma TNF-α, IL-1β, IL-18, IL-8 and IL-6 were quantified as
aforementioned. Hematological and blood biochemical tests
(parameters as listed in Table
SII) were outsourced to SRL, Inc.
Innate immune cell activity
assays
A total of ~9 months after the completion of the
trial, 10 healthy subjects were randomly selected from the 18 trial
participants and randomly assigned to either the L.
plantarum group (n=5) or the B. longum group (n=5)
through simple randomization. Ingestion of L. plantarum and
B. longum and blood sampling were conducted with the same
protocol as that of the trial (1x1011 cfu/day; twice
daily for 7 days). After a 7-day ingestion, innate immune cell
activity was measured using standard methods as described below in
detail.
Neutrophil activity
Measurement of the phagocytic activity of
neutrophils was outsourced to BML, Inc. Briefly, heparinized
peripheral blood (0.1 ml) from each subject was mixed with 40 µl
fluorescent microbeads (Fluoresbrite® YG Carboxylate
Microspheres 1.75 µm; Polysciences, Inc.) diluted 4-fold with
Dulbecco's PBS [-(without Ca2+ and Mg2+)] and
incubated with gentle agitation at 37˚C for 30 min. The samples
were treated with 2 ml 10X FACS lysing solution (BD Biosciences) at
4˚C for 15 min to lyse erythrocytes under gentle hypotonic
conditions, followed by flow cytometric analysis using FACSCalibur™
flow cytometer (BD Biosciences). Granulocytes were characterized as
medium-sized cells with high granularity and separated by setting a
medium forward scatter (FSC)/high side scatter (SSC) gating. The
percentage of fluorescence-positive granulocytes (granulocytes that
phagocytosed the fluorescent microbeads) to the total count of
granulocytes was calculated using the BD CellQuest™ Pro software
version 6.0 (BD Biosciences).
Natural killer (NK) cell activity
Analysis of NK cell activity using chromium-51
(51Cr) release assay was outsourced to SRL, Inc.
Lymphocytes were isolated from 5 ml peripheral blood using density
gradient centrifugation (Lymphosepar I; Immuno-Biological
Laboratories Co., Ltd.) according to the manufacturer's
instructions. The lymphocytes were washed twice with Dulbecco's PBS
(-) and resuspended at 1x106 cells/ml in RPMI-1640
supplemented with 10% FBS. A total of 200-µl aliquots (effector
cells; 2x105) were mixed with human chronic myelogenous
leukemia K562 cells (target cells; 1x104 cells/10 µl;
cat. no. CCL-243; American Type Culture Collection) radiolabeled
with 51Cr (PerkinElmer, Inc.) and incubated at 37˚C for
3.5 h in a 5% CO2 incubator. The cells were collected by
centrifugation, and the remaining 51Cr radioactivity was
measured using WIZARD® Automatic Gamma Counter
(PerkinElmer, Inc.). 51Cr-loaded K562 cells treated with
effector-free culture medium (RPMI-1640; 10% FBS) were used for the
quantification of spontaneously released 51Cr.
Macrophage activity
The serum level of neopterin, an activation marker
produced primarily by IFN-γ-stimulated monocytes and macrophages
(68,69), was assessed for macrophage
activity. Determination of serum neopterin was outsourced to SRL,
Inc. Serum (0.3 ml) was analyzed by a reverse-phase
high-performance liquid chromatography column-switching method
(LC-2000Plus; JASCO Corporation) (70) using Wakosil GP-N6 4.6x150 mm as a
pretreatment column and Wakosil-II 5C18 HG 4.6x250 mm as an
analysis column (FUJIFILM Wako Pure Chemical Corporation). The
neopterin level was determined by detecting its native fluorescence
(excitation, 353 nm; emission, 438 nm) with a fluorescence detector
(FP-2025; JASCO Corporation).
Statistical analysis
For the in vitro cytokine response assay,
when IL-1β was undetectable in negative control samples (14 of 20
enrolled subjects; Table SI),
0.5x lower limit of detection (0.05 pg/ml) was used to calculate
the QICI value.
In the analysis of the trial data, the interquartile
range (IQR) method was used to identify outliers; any values that
fell below Q1-1.5x IQR or above Q3 + 1.5x IQR (Q1, first quartile;
Q3, third quartile) were considered outliers and removed from the
statistical analysis (71,72). In order to perform statistical
tests of matched pairs, the paired values of the outliers were
removed, even if they fell into the non-outlier range. When
calculating QICI scores, outliers and zero values were handled as
follows: i) The outliers were replaced with mean values that were
calculated from the non-outliers to avoid unreasonable reduction of
QICI scores by simple removal of the outliers; and ii) zero values
in the measurement of IL-1β and IL-18 were replaced with the values
of 0.5x lower limit of detection (IL-1β, 0.05 pg/ml; IL-18, 8.30
pg/ml) (73,74).
The normality of the data was firstly examined using
the normal quantile-quantile plots and the Shapiro-Wilk test. On
the basis of the results from these normality tests, the Friedman
test was used, followed by the Nemenyi post hoc test for the data
from the clinical trial; a two-tailed paired Student's t-test was
used for the data from the assays of innate immune cell
activity.
All statistical analyses were performed with EZR
v1.53 (Saitama Medical Center, Jichi Medical University), which is
a graphical user interface for R (R Foundation for Statistical
Computing; https://www.R-project.org/) (75). A priori sample size
calculation and post hoc power analysis were performed using
G*Power v3.1.9.2 (Department of Experimental Psychology, Heinrich
Heine University Düsseldorf; https://www.psychologie.hhu.de/arbeitsgruppen/allgemeine-psychologie-und-arbeitspsychologie/gpower)
(76). Spearman's rank correlation
coefficients for the post hoc power analysis were calculated with
EZR v1.53(75). P<0.05 was
considered to indicate a statistically significant difference.
Results
Selection of LAB with the highest and
lowest QICI
Firstly, the species with the highest and lowest
QICI among the three probiotic LAB (L. plantarum, B.
longum and L. lactis ssp. lactis) were determined
in each of the 20 study subjects, using in vitro cytokine
response assay. L. plantarum demonstrated the highest QICI
in all subjects, whereas B. longum demonstrated the lowest
QICI (Table SI). L.
plantarum had a 52-7,210-fold (mean ± SD, 1,350±1,870; 95% CI,
458-2,250) higher QICI than B. longum and a 3-188-fold (mean
± SD, 33±44; 95% CI, 12-54) higher QICI than L. lactis ssp.
lactis. The present results indicated that L.
plantarum had a superior QFPD-like ability to stimulate innate
cytokine production by blood immune cells. Therefore, L.
plantarum and B. longum were selected in all subjects
for the subsequent clinical trial.
L. plantarum ingestion induces
QFPD-like innate cytokine responses in vivo
To investigate whether L. plantarum could
reproduce in vivo the QFPD-like immunomodulatory activity
observed in vitro, a single-arm, double-blind, prospective
trial that included three consecutive sessions was conducted: i) A
validation session using L. plantarum in the first 7 days;
ii) a 7-day washout period; and iii) a control session using B.
longum in the last 7 days. The peripheral blood samples that
were obtained before (day 0) and after (day 8) L. plantarum
ingestion and after B. longum ingestion (day 22) were
evaluated for plasma TNF-α, IL-1β, IL-18, IL-8 and IL-6, as well as
the QICI.
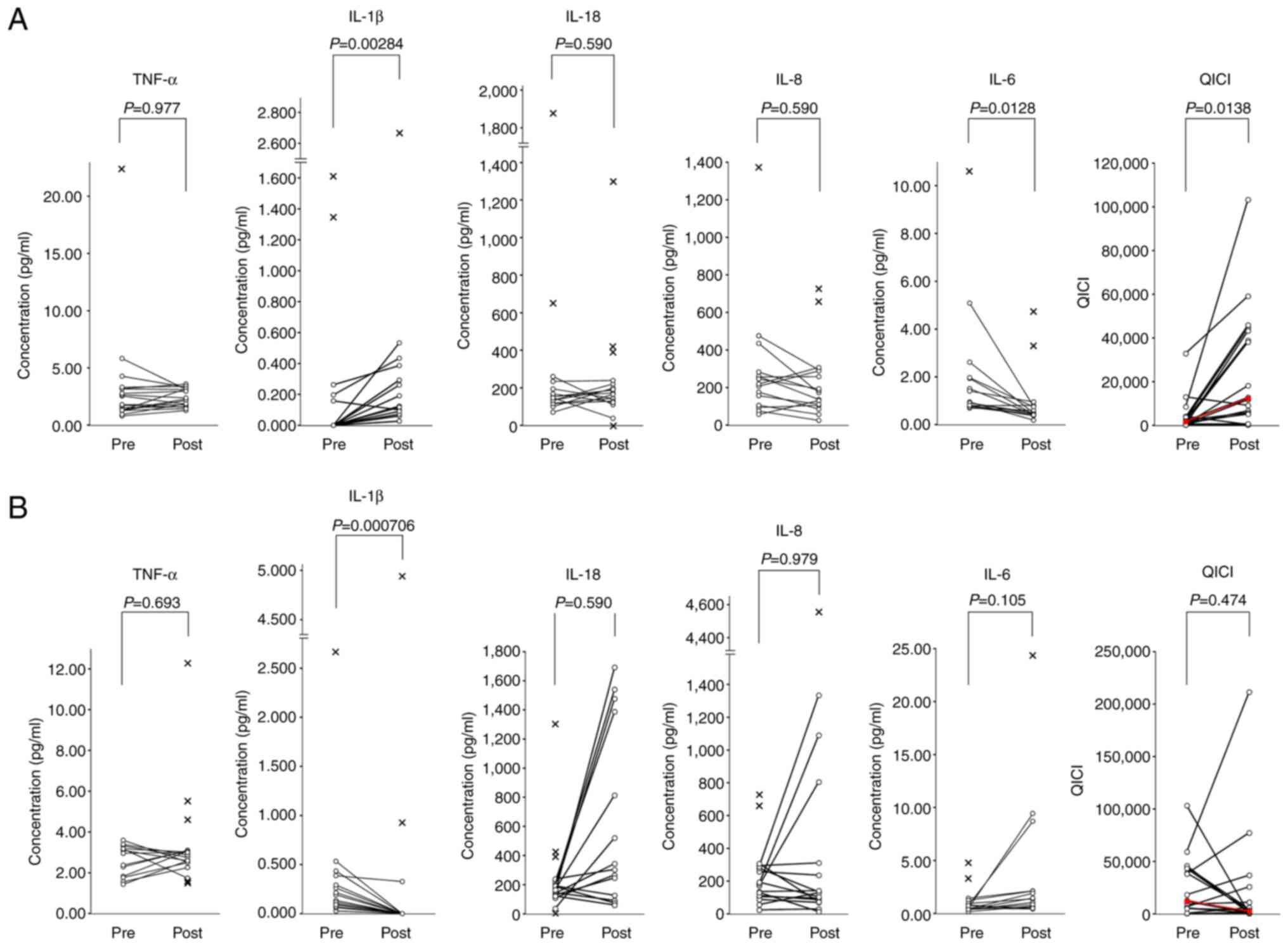
As indicated in Table
I, oral intake of L. plantarum significantly increased
plasma IL-1β [median (IQR), 0.000 (0.000-0.000) vs. 0.134
(0.092-0.292) pg/ml; P=0.0000310 after Friedman test; P=0.00284
after Nemenyi post hoc test] and decreased plasma IL-6 [median
(IQR), 1.180 (0.812-2.130) vs. 0.495 (0.425-0.775) pg/ml; P=0.0131
after Friedman test; P=0.0128 after Nemenyi post hoc test]. There
were no significant differences in the plasma levels of TNF-α,
IL-18 and IL-8. The QICI value was significantly increased [mean
fold change, 17-fold; median (IQR), 1,760 (680-3,550) vs. 12,300
(5,440-42,200); P=0.0173 after Friedman test; P=0.0138 after
Nemenyi post hoc test]. The QICI values were increased in 16 of 18
subjects (88.9%) with considerable individual differences in the
fold change (1-128-fold; mean fold change, 19-fold; Fig. 2A), suggesting that there are large
variations in responsiveness to L. plantarum among
individuals.
 | Table ICytokine changes in Lactobacillus
plantarum- and Bifidobacterium longum-administered
subjects. |
Table I
Cytokine changes in Lactobacillus
plantarum- and Bifidobacterium longum-administered
subjects.
| | Nemenyi post hoc
test |
|---|
| | Day 0 | Day 8 | Day 22 | Friedman test | Lactobacillus
plantarum (Day 0 vs. Day 8) | Bifidobacterium
longum (Day 8 vs. Day 22) |
|---|
| Cytokines | Median | (IQR) | Median | (IQR) | Median | (IQR) | P-value | P-value | P-value |
|---|
| TNF-α, pg/ml | 1.790 | (1.300-3.160) | 2.330 | (1.780-3.150) | 2.760 | (2.530-3.020) | 0.558 | | |
| IL-1β, pg/ml | 0.000 | (0.000-0.000) | 0.134 | (0.092-0.292) | 0.000 | (0.000-0.000) |
0.0000310a |
0.00284a |
0.000706a |
| IL-18, pg/ml | 154 | (126-214) | 155 | (132-191) | 324 | (219-991) | 0.146 | | |
| IL-8, pg/ml | 232 | (144-315) | 155 | (101-263) | 134 | (87-311) | 0.584 | | |
| IL-6, pg/ml | 1.180 | (0.812-2.130) | 0.495 | (0.425-0.775) | 1.420 | (0.692-2.220) | 0.0131a | 0.0128a | 0.105 |
| QFPD-induced innate
cytokine index, pg3/ml3 | 1,760 | (680-3,550) | 12,300 | (5,440-42,200) | 2,780 | (940-9,820) | 0.0173a | 0.0138a | 0.474 |
By contrast, oral intake of B. longum induced
a significant decrease in plasma IL-1β [median (IQR), 0.134
(0.092-0.292) vs. 0.000 (0.000-0.000) pg/ml; P=0.0000310 after
Friedman test; P=0.000706 after Nemenyi post hoc test]; however,
the QICI value did not change significantly [mean fold change,
2-fold; median (IQR), 12,300 (5,440-42,200) vs. 2,780 (940-9,820);
P=0.0173 after Friedman test; P=0.474 after Nemenyi post hoc test].
The QICI values increased in 8 of 18 subjects (44.4%), but the fold
changes were markedly lower (1-9-fold; mean fold change, 3-fold;
Fig. 2B) than those obtained
during the L. plantarum session. The present results
suggested that orally administered L. plantarum induced an
in vivo cytokine change similar to that induced by oral QFPD
in a previous experiment (51).
L. plantarum ingestion also caused a minor
but significant change the mean corpuscular hemoglobin
concentration [median (IQR), 33.3 (32.4-33.9) vs. 32.6
(32.1-33.1)%; P=0.00456 after Friedman test; P=0.00836 after
Nemenyi post hoc test] (Table
SII). No significant changes in the hematological parameters
were observed during the B. longum session (Table SII).
The post hoc two-tailed power analysis revealed that
satisfactory effect sizes were obtained [0.845 (QICI in the L.
plantarum session)-1.20 (IL-1β in the B. longum
session)], as well as statistical powers [0.908 (QICI in the L.
plantarum session)-0.996 (IL-1β in the B. longum
session)] after completion of the trial (Table SIII).
Effects of L. plantarum ingestion on
the innate immune cell activity
Subsequently, the present study examined whether the
L. plantarum-induced cytokine changes (increase in the QICI
score) led to the increased activity of innate immune cells. L.
plantarum ingestion significantly enhanced the activity of NK
cells, which are key effectors of antiviral innate immunity that
directly attack virus-infected host cells (Table II; Fig. 3A) (77,78).
By contrast, B. longum ingestion significantly promoted the
phagocytic activity of neutrophils (Table II; Fig. 3B). Neither L. plantarum nor
B. longum ingestion activated macrophages, as assessed by
the serum neopterin levels (Table
II; Fig. 3C). The present
results further supported the immunological benefits of L.
plantarum and B. longum against viral infection.
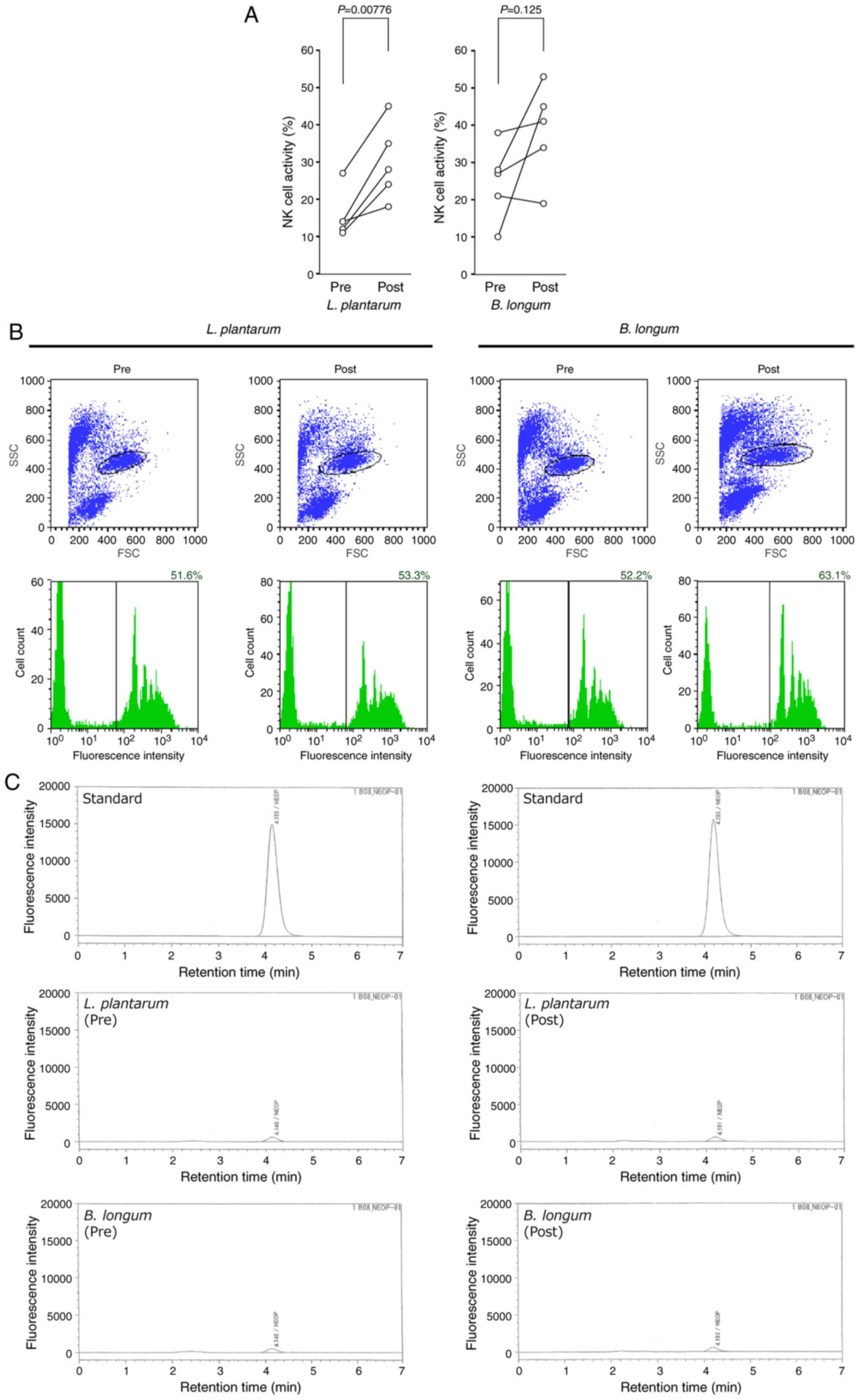 | Figure 3Effects of L. plantarum or
B. longum ingestion on the innate immune cell activity. (A)
NK cell activity before (pre) and after (post) the L.
plantarum or B. longum ingestion. (B) Phagocytic
activity of neutrophils. Representative flow cytometry plots are
presented in the upper panels. The vertical and horizontal axes are
SSC and FSC, respectively. Areas surrounded by black lines
represent granulocyte populations characterized as medium FSC/high
SSC (granulocyte gating). Histograms of fluorescence intensities of
the granulocytes separated by the granulocyte gating are presented
in the lower panels. The vertical and horizontal axes demonstrate
cell count and fluorescence intensity, respectively. A black
vertical line in each histogram indicates the threshold of
fluorescence-positive granulocytes (granulocytes that phagocytosed
the fluorescent microbeads). The phagocytic activity was calculated
as the ratio of fluorescence-positive granulocytes to the total
count of granulocytes and presented in the upper right corner of
each histogram. (C) Macrophage activity. The serum levels of
neopterin, an activation marker of monocytes and macrophages, were
determined using reverse-phase high-performance liquid
chromatography. Representative chromatograms are presented. The
vertical and horizontal axes show the intensity of native
fluorescence of neopterin and retention time, respectively. NK,
natural killer; L. plantarum, Lactobacillus
plantarum; B. longum, Bifidobacterium longum;
FSC, forward scatter; SSC, side scatter. |
| Table IIEffects of Lactobacillus
plantarum and Bifidobacterium longum ingestion on the
activity of innate immune cells. |
Table II
Effects of Lactobacillus
plantarum and Bifidobacterium longum ingestion on the
activity of innate immune cells.
| | Lactobacillus
plantarum | Bifidobacterium
longum |
|---|
| Activities of
innate immune cells | Baseline, mean ±
SEM | Post-ingestion,
mean ± SEM | P-value | 95% CI | Baseline, mean ±
SEM | Post-ingestion,
mean ± SEM | P-value | 95% CI |
|---|
| Phagocytic activity
of neutrophils, % | 54.50±4.39 | 57.80±4.47 | 0.512 | -9.28-15.80 | 50.90±4.80 | 57.00±5.22 | 0.0154a | 1.90-10.20 |
| Natural killer cell
activity, % | 15.60±2.60 | 30.00±4.17 |
0.00776b | 6.32-22.50 | 24.80±4.11 | 38.40±5.13 | 0.125 | -5.91-33.10 |
| Macrophage
activity, pmol/ml | 3.80±0.34 | 3.00±0.28 | 0.178 | -2.16-0.56 | 4.20±0.34 | 4.00±0.28 | 0.374 | -0.76-0.36 |
Discussion
A large body of evidence has suggested that L.
plantarum strengthens several aspects of the host defense
mechanism against the infection by respiratory viruses,
particularly seasonal and highly pathogenic influenza viruses. For
example, oral administration of L. plantarum in mice
significantly suppressed viral replication in the lungs and reduced
airway inflammation, thereby increasing survival rates (33-39).
The underlying immunological effects are known to be diverse,
including stimulation of type I IFN production (37,39),
enhancement of NK cell activity (36,39),
promotion of T helper type 1 cell-mediated immune responses
(33,34,39)
and activation of IgA-dependent mucosal immunity in the small
intestine and lung (34,35). Similarly, randomized controlled
trials have demonstrated that oral intake of L. plantarum
reduced the risk of upper/lower respiratory tract infection and
alleviated the respiratory symptoms of infected patients (30,79).
The protective efficacy is associated with enhancement of the
phagocytic activity of granulocytes, reduction of the plasma
proinflammatory cytokines IFN-γ and TNF-α, elevation of the
anti-inflammatory cytokines IL-4 and IL-10, activation of
CD8+ T cells and induction of the specific secretory IgA
neutralizing antibodies in the bronchoalveolar lavage fluid (BALF)
and sera (30,31).
In line with the wide variety of immunomodulatory
abilities of L. plantarum, the SNK12 strain was indicated to
exhibit protective effects against influenza A virus subtype H1N1
(57,58). The SNK12 strain has been isolated
from traditional non-salted pickles of autochthonous red turnip
that have been produced in the Kiso area of Nagano prefecture in
Japan for >400 years (80).
Recent animal studies demonstrated that oral administration of
L. plantarum strain SNK12 to mice before influenza viral
challenge suppressed the viral load in the BALF and lung, induced a
higher titer of neutralizing antibodies in the BALF and sera and
higher levels of specific IgA in BALF and feces compared with
control mice and mice treated with the anti-influenza drug
oseltamivir (57,58).
The present study revealed that L. plantarum
SNK12 could also upregulate a subset of proinflammatory cytokines,
as assessed both in vitro and in humans. This
immunomodulatory effect is likely contradictory to its potential
for clinical benefits against the influenza virus, since the
patients present with a broad range of inflammatory symptoms.
However, it is notable that acute, low-grade inflammation is a
physiological basis for early stages of host defense mechanisms and
has been demonstrated to serve a protective role against viral
infection. Kechaou et al (32) demonstrated that a proinflammatory
L. plantarum strain with superior ability to stimulate the
production of IL-8 and IL-12 markedly inhibited viral proliferation
in the lung and alleviated clinical symptoms in mice when orally
administered before or after influenza virus challenge.
Furthermore, Park et al (33) revealed that ingestion by mice of a
probiotic L. plantarum strain conferred protection against
influenza virus by elevating both IL-12 and IFN-γ levels in the
BALF and inducing low-grade inflammation.
QFPD consists of 21 traditional Chinese herbs
optimized specifically against the symptoms of COVID-19(43). QFPD has demonstrated satisfactory
therapeutic benefits in patients with mild-to-severe disease in
clinical trials in China (46-50)
and has been recommended officially for the treatment of
COVID-19(43). Our recent clinical
study indicated that oral QFPD upregulated the blood levels of
TNF-α, IL-1β, IL-18 and IL-8, which are key mediators of innate
immune responses to viral infection (51). TNF-α, IL-1β and IL-18 are induced
directly by toll like receptor (TLR)7/TLR8 and NLR family pyrin
domain containing 3 (NLRP3), foreign single-stranded RNA (ssRNA)
sensors in dendritic cells and macrophages (81-83).
These ‘immediate-early’ cytokines initiate and coordinate a broad
spectrum of downstream antiviral immune cascades (81-83).
Notably, recent metagenomics studies demonstrated that the TLR7/8-
and NLRP3/inflammasome-mediated inflammatory pathways are strongly
suppressed in the upper airway of patients with COVID-19 and those
non-responsive to SARS-CoV-2 infection early in the course of the
disease (84,85). These findings highlight the
importance of the active TLR7/8- and NLRP3-driven inflammatory
pathways in the early stages of anti-SARS-CoV-2 immunity. We
hypothesized that QFPD, which mimics the blood cytokine environment
produced by TLR7/8- and NLRP3-driven early innate immune responses
to ssRNA viruses, may be effective in preventing SARS-CoV-2
infection (51). The present study
suggested that L. plantarum, which can stimulate innate
cytokine changes similar to those induced by oral QFPD, may also
potentially provide a protective benefit against COVID-19.
IL-6 serves key roles in complex immune
dysregulation and systemic hyperinflammation, which are hallmarks
of severe COVID-19, and IL-6 blood level has been associated with
COVID-19 severity and mortality (54-56).
The production of IL-6 and TNF-α is stimulated directly by common
TLR7/8-driven intracellular signaling in response to ssRNA viruses
(81-83).
However, L. plantarum ingestion significantly increased the
plasma level of TNF-α, whereas it downregulated IL-6 plasma level
in the present trial. Although the mechanism of this opposite
effect remains unknown, the ability of L. plantarum to
reduce blood IL-6 may be indicative of its prophylactic
administration to uninfected individuals.
Recent transcriptomic studies have demonstrated
that exhausted NK cell responses determine severity and fatality of
COVID-19. Liu et al (86)
have identified IL-15-mediated attenuated inflammation in NK cells
as being primarily associated with COVID-19 severity. A study by
Sahoo et al (87) has also
highlighted that IL15-mediated NK cell exhaustion, senescence and
apoptosis are important determinants for severe/fatal COVID-19.
L. plantarum-induced activation of NK cells may therefore be
efficacious as an adjunctive therapy to improve NK cell exhaustion
and dysfunction in severe or fatal COVID-19.
The main limitations of the current study are the
small number of participants, the selection of uninfected
individuals as subjects and a female-biased gender ratio in the
subjects. Further clinical studies with larger cohorts are required
to confirm the conclusions and determine generalizability. The
uninfected subjects employed were healthy, in order to examine the
feasibility of prophylactic, daily use of L. plantarum. As a
result, no chest CT images of the subjects were obtained.
Additional studies are essential to clarify whether L.
plantarum can induce the similar cytokine changes and improve
chest CT findings in patients with COVID-19. Further studies are
also required to identify probiotic strains with the ability to
strengthen innate immunity by inducing moderate physiological
inflammation. In addition, in the control session using B.
longum (day 15-21), blood samples were obtained at day 8 as
baseline, not at day 14. Since the plasma IL-1β and IL-6 levels had
been up- or down-regulated by L. plantarum by day 8, it is
not possible to exclude the possibility that the IL-1β and IL-6
levels were spontaneously restored to the day 0 levels during days
8-22 without the effects of B. longum. Similarly, there was
a significant increase in the QICI in the L. plantarum
session (Table I, comparison
between day 0 and day 8), whereas no significant change in the QICI
was observed in the B. longum session (comparison between
day 8 and day 22). B. longum sustained the QICI score that
had been upregulated by L. plantarum. Therefore, it cannot
be excluded either that B. longum also had positive effects
on the QICI score in vivo.
Considering the recent emergence of novel
SARS-CoV-2 variants with relevant mutations that potentially affect
the efficacy of vaccines and therapeutic antibodies, there is an
increasing need to prepare diverse prophylactic and therapeutic
options against COVID-19. The present study indicated that L.
plantarum exhibited a superior ability to mimic inflammatory
innate cytokine responses essential for early stages of host
defense mechanism against viral infection. Daily consumption of
probiotic L. plantarum strains may be a reasonably safe,
cost-effective and viable option to protect uninfected individuals
against SARS-CoV-2 infection during the pandemic.
Supplementary Material
In vitro cytokine response
assay.
Hematological changes in
Lactobacillus plantarum- and Bifidobacterium
longum-administered subjects.
Two-tailed post hoc power analysis
(α=0.05; n=18).
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the
current study are available from the corresponding author on
reasonable request.
Authors' contributions
YK, YN, TE, TA and TN conceived the study. YK, YN,
TE, TA and TN developed the methodology. TN performed formal
analysis. YK, KA, KY and TE performed/interpreted the experiments.
YK, KA, KY and TE provided resources. YK and TE curated data. TN
wrote the original draft. YK, YN, KA, KY, TE and TA reviewed and
edited the manuscript. TN was involved in visualization. YK, YN, TA
and TN supervised the study. YK, KA and TE were involved in project
administration. YK, TE and TN confirm the authenticity of all the
raw data. All authors have read and approved the final
manuscript.
Ethics approval and consent to
participate
The present study was carried out in accordance
with The Code of Ethics of the World Medical Association (The
Declaration of Helsinki). All procedures were reviewed and approved
by the Ethics Committees of Takanawa Clinic (approval nos. 2016-3,
20 December 2016 and 2020-2, 7 May 2020; Tokyo, Japan). A signed
informed consent form was obtained from each participant prior to
inclusion in the present study.
Patient consent for publication
Not applicable.
Competing interests
YK, KA, KY and TE are employees of Takanawa Clinic.
TA and TN serve as research advisors to Takanawa Clinic and receive
advisory fees.
References
|
1
|
Lukassen S, Chua RL, Trefzer T, Kahn NC,
Schneider MA, Muley T, Winter H, Meister M, Veith C, Boots AW, et
al: SARS-CoV-2 receptor ACE2 and TMPRSS2 are primarily expressed in
bronchial transient secretory cells. EMBO J.
39(e105114)2020.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Sungnak W, Huang N, Bécavin C, Berg M,
Queen R, Litvinukova M, Talavera-López C, Maatz H, Reichart D,
Sampaziotis F, et al: SARS-CoV-2 entry factors are highly expressed
in nasal epithelial cells together with innate immune genes. Nat
Med. 26:681–687. 2020.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Ziegler CGK, Allon SJ, Nyquist SK, Mbano
IM, Miao VN, Tzouanas CN, Cao Y, Yousif AS, Bals J, Hauser BM, et
al: SARS-CoV-2 receptor ACE2 is an interferon-stimulated gene in
human airway epithelial cells and is detected in specific cell
subsets across tissues. Cell. 181:1016–1035.e19. 2020.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Lee JJ, Kopetz S, Vilar E, Shen JP, Chen K
and Maitra A: Relative abundance of SARS-CoV-2 entry genes in the
enterocytes of the lower gastrointestinal tract. Genes (Basel).
11(645)2020.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Zang R, Gomez Castro MF, McCune BT, Zeng
Q, Rothlauf PW, Sonnek NM, Liu Z, Brulois KF, Wang X, Greenberg HB,
et al: TMPRSS2 and TMPRSS4 promote SARS-CoV-2 infection of human
small intestinal enterocytes. Sci Immunol.
5(eabc3582)2020.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Lamers MM, Beumer J, van der Vaart J,
Knoops K, Puschhof J, Breugem TI, Ravelli RBG, Paul van Schayck J,
Mykytyn AZ, Duimel HQ, et al: SARS-CoV-2 productively infects human
gut enterocytes. Science. 369:50–54. 2020.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Xiao F, Tang M, Zheng X, Liu Y, Li X and
Shan H: Evidence for gastrointestinal infection of SARS-CoV-2.
Gastroenterology. 158:1831–1833.e3. 2020.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Zhou J, Li C, Liu X, Chiu MC, Zhao X, Wang
D, Wei Y, Lee A, Zhang AJ, Chu H, et al: Infection of bat and human
intestinal organoids by SARS-CoV-2. Nat Med. 26:1077–1083.
2020.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Chen Y, Chen L, Deng Q, Zhang G, Wu K, Ni
L, Yang Y, Liu B, Wang W, Wei C, et al: The presence of SARS-CoV-2
RNA in the feces of COVID-19 patients. J Med Virol. 92:833–840.
2020.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Kipkorir V, Cheruiyot I, Ngure B, Misiani
M and Munguti J: Prolonged SARS-CoV-2 RNA detection in anal/rectal
swabs and stool specimens in COVID-19 patients after negative
conversion in nasopharyngeal RT-PCR test. J Med Virol.
92:2328–2331. 2020.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Wu Y, Guo C, Tang L, Hong Z, Zhou J, Dong
X, Yin H, Xiao Q, Tang Y, Qu X, et al: Prolonged presence of
SARS-CoV-2 viral RNA in faecal samples. Lancet Gastroenterol
Hepatol. 5:434–435. 2020.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Gu J, Han B and Wang J: COVID-19:
Gastrointestinal manifestations and potential fecal-oral
transmission. Gastroenterology. 158:1518–1519. 2020.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Wong SH, Lui RN and Sung JJ: Covid-19 and
the digestive system. J Gastroenterol Hepatol. 35:744–748.
2020.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Yang L and Tu L: Implications of
gastrointestinal manifestations of COVID-19. Lancet Gastroenterol
Hepatol. 5:629–630. 2020.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Budden KF, Gellatly SL, Wood DL, Cooper
MA, Morrison M, Hugenholtz P and Hansbro PM: Emerging pathogenic
links between microbiota and the gut-lung axis. Nat Rev Microbiol.
15:55–63. 2017.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Dumas A, Bernard L, Poquet Y,
Lugo-Villarino G and Neyrolles O: The role of the lung microbiota
and the gut-lung axis in respiratory infectious diseases. Cell
Microbiol. 20(e12966)2018.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Zhang D, Li S, Wang N, Tan HY, Zhang Z and
Feng Y: The cross-talk between gut microbiota and lungs in common
lung diseases. Front Microbiol. 11(301)2020.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Gu S, Chen Y, Wu Z, Chen Y, Gao H, Lv L,
Guo F, Zhang X, Luo R, Huang C, et al: Alterations of the gut
microbiota in patients with coronavirus disease 2019 or H1N1
influenza. Clin Infect Dis. 71:2669–2678. 2020.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Gou W, Fu Y, Yue L, Chen GD, Cai X, Shuai
M, Xu F, Yi X, Chen H, Zhu Y, et al: Gut microbiota may underlie
the predisposition of healthy individuals to COVID-19. J Genet
Genomics. 48:792–802. 2021.
|
|
20
|
Zuo T, Liu Q, Zhang F, Lui GC, Tso EY,
Yeoh YK, Chen Z, Boon SS, Chan FK, Chan PK, et al: Depicting
SARS-CoV-2 faecal viral activity in association with gut microbiota
composition in patients with COVID-19. Gut. 70:276–284.
2021.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Zuo T, Zhang F, Lui GCY, Yeoh YK, Li AYL,
Zhan H, Wan Y, Chung ACK, Cheung CP, Chen N, et al: Alterations in
gut microbiota of patients with COVID-19 during time of
hospitalization. Gastroenterology. 159:944–955.e8. 2020.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Lehtoranta L, Pitkaranta A and Korpela R:
Probiotics in respiratory virus infections. Eur J Clin Microbiol
Infect Dis. 33:1289–1302. 2014.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Lei WT, Shih PC, Liu SJ, Lin CY and Yeh
TL: Effect of probiotics and prebiotics on immune response to
influenza vaccination in adults: A systematic review and
meta-analysis of randomized controlled trials. Nutrients.
9(1175)2017.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Luoto R, Ruuskanen O, Waris M, Kalliomaki
M, Salminen S and Isolauri E: Prebiotic and probiotic
supplementation prevents rhinovirus infections in preterm infants:
A randomized, placebo-controlled trial. J Allergy Clin Immunol.
133:405–413. 2014.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Turner RB, Woodfolk JA, Borish L, Steinke
JW, Patrie JT, Muehling LM, Lahtinen S and Lehtinen MJ: Effect of
probiotic on innate inflammatory response and viral shedding in
experimental rhinovirus infection-a randomised controlled trial.
Benef Microbes. 8:207–215. 2017.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Yeh TL, Shih PC, Liu SJ, Lin CH, Liu JM,
Lei WT and Lin CY: The influence of prebiotic or probiotic
supplementation on antibody titers after influenza vaccination: A
systematic review and meta-analysis of randomized controlled
trials. Drug Des Devel Ther. 12:217–230. 2018.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Blanco-Melo D, Nilsson-Payant BE, Liu WC,
Uhl S, Hoagland D, Møller R, Jordan TX, Oishi K, Panis M, Sachs D,
et al: Imbalanced host response to SARS-CoV-2 drives development of
COVID-19. Cell. 181:1036–1045.e9. 2020.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Cao X: COVID-19: Immunopathology and its
implications for therapy. Nat Rev Immunol. 20:269–270.
2020.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Giamarellos-Bourboulis EJ, Netea MG,
Rovina N, Akinosoglou K, Antoniadou A, Antonakos N, Damoraki G,
Gkavogianni T, Adami ME, Katsaounou P, et al: Complex immune
dysregulation in COVID-19 patients with severe respiratory failure.
Cell Host Microbe. 27:992–1000.e3. 2020.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Chong HX, Yusoff NAA, Hor YY, Lew LC,
Jaafar MH, Choi SB, Yusoff MSB, Wahid N, Abdullah MFIL, Zakaria N,
et al: Lactobacillus plantarum DR7 improved upper
respiratory tract infections via enhancing immune and inflammatory
parameters: A randomized, double-blind, placebo-controlled study. J
Dairy Sci. 102:4783–4797. 2019.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Rask C, Adlerberth I, Berggren A, Ahren IL
and Wold AE: Differential effect on cell-mediated immunity in human
volunteers after intake of different lactobacilli. Clin Exp
Immunol. 172:321–332. 2013.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Kechaou N, Chain F, Gratadoux JJ, Blugeon
S, Bertho N, Chevalier C, Le Goffic R, Courau S, Molimard P, Chatel
JM, et al: Identification of one novel candidate probiotic
Lactobacillus plantarum strain active against influenza
virus infection in mice by a large-scale screening. Appl Environ
Microbiol. 79:1491–1499. 2013.PubMed/NCBI View Article : Google Scholar
|
|
33
|
Park MK, Ngo V, Kwon YM, Lee YT, Yoo S,
Cho YH, Hong SM, Hwang HS, Ko EJ, Jung YJ, et al: Lactobacillus
plantarum DK119 as a probiotic confers protection against
influenza virus by modulating innate immunity. PLoS One.
8(e75368)2013.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Kawashima T, Hayashi K, Kosaka A,
Kawashima M, Igarashi T, Tsutsui H, Tsuji NM, Nishimura I, Hayashi
T and Obata A: Lactobacillus plantarum strain YU from
fermented foods activates Th1 and protective immune responses. Int
Immunopharmacol. 11:2017–2024. 2011.PubMed/NCBI View Article : Google Scholar
|
|
35
|
Kikuchi Y, Kunitoh-Asari A, Hayakawa K,
Imai S, Kasuya K, Abe K, Adachi Y, Fukudome S, Takahashi Y and
Hachimura S: Oral administration of Lactobacillus plantarum
strain AYA enhances IgA secretion and provides survival protection
against influenza virus infection in mice. PLoS One.
9(e86416)2014.PubMed/NCBI View Article : Google Scholar
|
|
36
|
Kim DH, Chung WC, Chun SH, Han JH, Song MJ
and Lee KW: Enhancing the natural killer cell activity and
anti-influenza effect of heat-treated Lactobacillus
plantarum nF1-fortified yogurt in mice. J Dairy Sci.
101:10675–10684. 2018.PubMed/NCBI View Article : Google Scholar
|
|
37
|
Maeda N, Nakamura R, Hirose Y, Murosaki S,
Yamamoto Y, Kase T and Yoshikai Y: Oral administration of
heat-killed Lactobacillus plantarum L-137 enhances
protection against influenza virus infection by stimulation of type
I interferon production in mice. Int Immunopharmacol. 9:1122–1125.
2009.PubMed/NCBI View Article : Google Scholar
|
|
38
|
Park S, Kim JI, Bae JY, Yoo K, Kim H, Kim
IH, Park MS and Lee I: Effects of heat-killed Lactobacillus
plantarum against influenza viruses in mice. J Microbiol.
56:145–149. 2018.PubMed/NCBI View Article : Google Scholar
|
|
39
|
Takeda S, Takeshita M, Kikuchi Y, Dashnyam
B, Kawahara S, Yoshida H, Watanabe W, Muguruma M and Kurokawa M:
Efficacy of oral administration of heat-killed probiotics from
Mongolian dairy products against influenza infection in mice:
Alleviation of influenza infection by its immunomodulatory activity
through intestinal immunity. Int Immunopharmacol. 11:1976–1983.
2011.PubMed/NCBI View Article : Google Scholar
|
|
40
|
Kawahara T, Takahashi T, Oishi K, Tanaka
H, Masuda M, Takahashi S, Takano M, Kawakami T, Fukushima K,
Kanazawa H and Suzuki T: Consecutive oral administration of
Bifidobacterium longum MM-2 improves the defense system
against influenza virus infection by enhancing natural killer cell
activity in a murine model. Microbiol Immunol. 59:1–12.
2015.PubMed/NCBI View Article : Google Scholar
|
|
41
|
Namba K, Hatano M, Yaeshima T, Takase M
and Suzuki K: Effects of Bifidobacterium longum BB536
administration on influenza infection, influenza vaccine antibody
titer, and cell-mediated immunity in the elderly. Biosci Biotechnol
Biochem. 74:939–945. 2010.PubMed/NCBI View Article : Google Scholar
|
|
42
|
Sugimura T, Takahashi H, Jounai K, Ohshio
K, Kanayama M, Tazumi K, Tanihata Y, Miura Y, Fujiwara D and
Yamamoto N: Effects of oral intake of plasmacytoid dendritic
cells-stimulative lactic acid bacterial strain on pathogenesis of
influenza-like illness and immunological response to influenza
virus. Br J Nutr. 114:727–733. 2015.PubMed/NCBI View Article : Google Scholar
|
|
43
|
Shi HY, Zhu X, Li WL, Mak JWY, Wong SH,
Zhu ST, Guo SL, Chan FKL, Zhang ST and Ng SC: Modulation of gut
microbiota protects against viral respiratory tract infections: A
systematic review of animal and clinical studies. Eur J Nutr: Apr
14, 2021 (Epub ahead of print).
|
|
44
|
Wang F, Pan B, Xu S, Xu Z, Zhang T, Zhang
Q, Bao Y, Wang Y, Zhang J, Xu C and Xue X: A meta-analysis reveals
the effectiveness of probiotics and prebiotics against respiratory
viral infection. Biosci Rep. 41(BSR20203638)2021.PubMed/NCBI View Article : Google Scholar
|
|
45
|
Wei PF (ed): Diagnosis and Treatment
Protocol for Novel Coronavirus Pneumonia (Trial Version 7). Chin
Med J (Engl). 133:1087–1095. 2020.PubMed/NCBI View Article : Google Scholar
|
|
46
|
Cao P, Wu S, Wu T, Deng Y, Zhang Q, Wang K
and Zhang Y: The important role of polysaccharides from a
traditional Chinese medicine-Lung cleansing and detoxifying
decoction against the COVID-19 pandemic. Carbohydr Polym.
240(116346)2020.PubMed/NCBI View Article : Google Scholar
|
|
47
|
Shi N, Liu B, Liang N, Ma Y, Ge Y, Yi H,
Wo H, Gu H, Kuang Y, Tang S, et al: Association between early
treatment with Qingfei Paidu decoction and favorable clinical
outcomes in patients with COVID-19: A retrospective multicenter
cohort study. Pharmacol Res. 161(105290)2020.PubMed/NCBI View Article : Google Scholar
|
|
48
|
Zhao ZH, Zhou Y, Li WH, Huang QS, Tang ZH
and Li H: Analysis of traditional Chinese medicine diagnosis and
treatment strategies for COVID-19 based on ‘The Diagnosis and
Treatment Program for Coronavirus Disease-2019’ from Chinese
Authority. Am J Chin Med. 48:1035–1049. 2020.PubMed/NCBI View Article : Google Scholar
|
|
49
|
Zhong LLD, Lam WC, Yang W, Chan KW, Sze
SCW, Miao J, Yung KKL, Bian Z and Wong VT: Potential targets for
treatment of coronavirus disease 2019 (COVID-19): A review of
Qing-Fei-Pai-Du-Tang and its major herbs. Am J Chin Med.
48:1051–1071. 2020.PubMed/NCBI View Article : Google Scholar
|
|
50
|
Xin S, Cheng X, Zhu B, Liao X, Yang F,
Song L, Shi Y, Guan X, Su R, Wang J, et al: Clinical retrospective
study on the efficacy of Qingfei Paidu decoction combined with
Western medicine for COVID-19 treatment. Biomed Pharmacother.
129(110500)2020.PubMed/NCBI View Article : Google Scholar
|
|
51
|
Kageyama Y, Aida K, Kawauchi K, Morimoto
M, Ebisui T, Akiyama T and Nakamura T: Qingfei Paidu decoction, a
Chinese herbal medicine against COVID-19, elevates the blood levels
of pro-inflammatory cytokines: An open-label, single-arm pilot
study. World Acad Sci J. 3(25)2021.
|
|
52
|
Popkin BM, Du S, Green WD, Beck MA,
Algaith T, Herbst CH, Alsukait RF, Alluhidan M, Alazemi N and
Shekar M: Individuals with obesity and COVID-19: A global
perspective on the epidemiology and biological relationships. Obes
Rev. 21(e13128)2020.PubMed/NCBI View Article : Google Scholar
|
|
53
|
Kompaniyets L, Goodman AB, Belay B,
Freedman DS, Sucosky MS, Lange SJ, Gundlapalli AV, Boehmer TK and
Blanck HM: Body mass index and risk for COVID-19-related
hospitalization, intensive care unit admission, invasive mechanical
ventilation, and Death-United States, march-december 2020. MMWR
Morb Mortal Wkly Rep. 70:355–361. 2021.PubMed/NCBI View Article : Google Scholar
|
|
54
|
Elshazli RM, Toraih EA, Elgaml A,
El-Mowafy M, El-Mesery M, Amin MN, Hussein MH, Killackey MT, Fawzy
MS and Kandil E: Diagnostic and prognostic value of hematological
and immunological markers in COVID-19 infection: A meta-analysis of
6320 patients. PLoS One. 15(e0238160)2020.PubMed/NCBI View Article : Google Scholar
|
|
55
|
Mesas AE, Cavero-Redondo I, Álvarez-Bueno
C, Sarriá Cabrera MA, Maffei de Andrade S, Sequí-Dominguez I and
Martínez-Vizcaíno V: Predictors of in-hospital COVID-19 mortality:
A comprehensive systematic review and meta-analysis exploring
differences by age, sex and health conditions. PLoS One.
15(e0241742)2020.PubMed/NCBI View Article : Google Scholar
|
|
56
|
Zhang X, Tan Y, Ling Y, Lu G, Liu F, Yi Z,
Jia X, Wu M, Shi B, Xu S, et al: Viral and host factors related to
the clinical outcome of COVID-19. Nature. 583:437–440.
2020.PubMed/NCBI View Article : Google Scholar
|
|
57
|
Okumura A, Watanabe T, Hayashi K and
Yoshida H: Oral administration of distribution-processed
Lactobacillus plantarum strain SNK12 protects against
influenza virus infection. Jpn Pharmacol Ther. 47:1607–1612.
2019.
|
|
58
|
Watanabe T, Hayashi K, Kan T, Ohwaki M and
Kawahara T: Anti-influenza virus effects of Enterococcus faecalis
KH2 and Lactobacillus plantarum SNK12 RNA. Biosci Microbiota
Food Health. 40:43–49. 2021.PubMed/NCBI View Article : Google Scholar
|
|
59
|
Xiao JZ, Kondo S, Yanagisawa N, Miyaji K,
Enomoto K, Sakoda T, Iwatsuki K and Enomoto T: Clinical efficacy of
probiotic Bifidobacterium longum for the treatment of
symptoms of Japanese cedar pollen allergy in subjects evaluated in
an environmental exposure unit. Allergol Int. 56:67–75.
2007.PubMed/NCBI View Article : Google Scholar
|
|
60
|
Xiao JZ, Kondo S, Yanagisawa N, Takahashi
N, Odamaki T, Iwabuchi N, Iwatsuki K, Kokubo S, Togashi H, Enomoto
K and Enomoto T: Effect of probiotic Bifidobacterium longum
BB536 [corrected] in relieving clinical symptoms and modulating
plasma cytokine levels of Japanese cedar pollinosis during the
pollen season. A randomized double-blind, placebo-controlled trial.
J Investig Allergol Clin Immunol. 16:86–93. 2006.PubMed/NCBI
|
|
61
|
Iwabuchi N, Xiao JZ, Yaeshima T and
Iwatsuki K: Oral administration of Bifidobacterium longum
ameliorates influenza virus infection in mice. Biol Pharm Bull.
34:1352–1355. 2011.PubMed/NCBI View Article : Google Scholar
|
|
62
|
Lau AS, Yanagisawa N, Hor YY, Lew LC, Ong
JS, Chuah LO, Lee YY, Choi SB, Rashid F, Wahid N, et al:
Bifidobacterium longum BB536 alleviated upper respiratory
illnesses and modulated gut microbiota profiles in Malaysian
pre-school children. Benef Microbes. 9:61–70. 2018.PubMed/NCBI View Article : Google Scholar
|
|
63
|
Odamaki T, Sugahara H, Yonezawa S,
Yaeshima T, Iwatsuki K, Tanabe S, Tominaga T, Togashi H, Benno Y
and Xiao JZ: Effect of the oral intake of yogurt containing
Bifidobacterium longum BB536 on the cell numbers of
enterotoxigenic Bacteroides fragilis in microbiota. Anaerobe.
18:14–18. 2012.PubMed/NCBI View Article : Google Scholar
|
|
64
|
Odamaki T, Xiao JZ, Iwabuchi N, Sakamoto
M, Takahashi N, Kondo S, Miyaji K, Iwatsuki K, Togashi H, Enomoto T
and Benno Y: Influence of Bifidobacterium longum BB536
intake on faecal microbiota in individuals with Japanese cedar
pollinosis during the pollen season. J Med Microbiol. 56:1301–1308.
2007.PubMed/NCBI View Article : Google Scholar
|
|
65
|
Holvoet S, Zuercher AW, Julien-Javaux F,
Perrot M and Mercenier A: Characterization of candidate
anti-allergic probiotic strains in a model of th2-skewed human
peripheral blood mononuclear cells. Int Arch Allergy Immunol.
161:142–154. 2013.PubMed/NCBI View Article : Google Scholar
|
|
66
|
Medina M, Izquierdo E, Ennahar S and Sanz
Y: Differential immunomodulatory properties of Bifidobacterium
logum strains: Relevance to probiotic selection and clinical
applications. Clin Exp Immunol. 150:531–538. 2007.PubMed/NCBI View Article : Google Scholar
|
|
67
|
Niers LE, Timmerman HM, Rijkers GT, van
Bleek GM, van Uden NO, Knol EF, Kapsenberg ML, Kimpen JL and
Hoekstra MO: Identification of strong interleukin-10 inducing
lactic acid bacteria which down-regulate T helper type 2 cytokines.
Clin Exp Allergy. 35:1481–1489. 2005.PubMed/NCBI View Article : Google Scholar
|
|
68
|
Hoffmann G, Wirleitner B and Fuchs D:
Potential role of immune system activation-associated production of
neopterin derivatives in humans. Inflamm Res. 52:313–321.
2003.PubMed/NCBI View Article : Google Scholar
|
|
69
|
Gieseg SP, Baxter-Parker G and Lindsay A:
Neopterin, inflammation, and Oxidative Stress: What could we be
missing? Antioxidants (Basel). 7(80)2018.PubMed/NCBI View Article : Google Scholar
|
|
70
|
Hausen A, Fuchs D, König K and Wachter H:
Determination of neopterine in human urine by reversed-phase
high-performance liquid chromatography. J Chromatogr. 227:61–70.
1982.PubMed/NCBI View Article : Google Scholar
|
|
71
|
Gleiss A, Sanchez-Cabo F, Perco P, Tong D
and Heinze G: Adaptive trimmed t-statistics for identifying
predominantly high expression in a microarray experiment. Stat Med.
30:52–61. 2011.PubMed/NCBI View Article : Google Scholar
|
|
72
|
Krzywinski M and Altman N: Visualizing
samples with box plots. Nat Methods. 11:119–120. 2014.PubMed/NCBI View Article : Google Scholar
|
|
73
|
Domthong U, Parikh CR, Kimmel PL and
Chinchilli VM: Assessment, Serial Evaluation, Subsequent Sequelae
of Acute Kidney Injury Consortium. Assessing the agreement of
biomarker data in the presence of left-censoring. BMC Nephrol.
15(144)2014.PubMed/NCBI View Article : Google Scholar
|
|
74
|
Yu X, Lakerveld AJ, Imholz S, Hendriks M,
Ten Brink SCA, Mulder HL, de Haan K, Schepp RM, Luytjes W, de Jong
MD, et al: Antibody and local cytokine response to respiratory
syncytial virus infection in community-dwelling older adults.
mSphere. 5:e00577–20. 2020.PubMed/NCBI View Article : Google Scholar
|
|
75
|
Kanda Y: Investigation of the freely
available easy-to-use software ‘EZR’ for medical statistics. Bone
Marrow Transplant. 48:452–458. 2013.PubMed/NCBI View Article : Google Scholar
|
|
76
|
Faul F, Erdfelder E, Lang AG and Buchner
A: G*Power 3: A flexible statistical power analysis program for the
social, behavioral, and biomedical sciences. Behav Res Methods.
39:175–191. 2007.PubMed/NCBI View Article : Google Scholar
|
|
77
|
Waggoner SN, Reighard SD, Gyurova IE,
Cranert SA, Mahl SE, Karmele EP, McNally JP, Moran MT, Brooks TR,
Yaqoob F, et al: Roles of natural killer cells in antiviral
immunity. Curr Opin Virol. 16:15–23. 2016.PubMed/NCBI View Article : Google Scholar
|
|
78
|
Björkström NK, Strunz B and Ljunggren HG:
Natural killer cells in antiviral immunity. Nat Rev Immunol: Jun
11, 2021 (Epub ahead of print).
|
|
79
|
Berggren A, Lazou Ahren I, Larsson N and
Onning G: Randomised, double-blind and placebo-controlled study
using new probiotic lactobacilli for strengthening the body immune
defence against viral infections. Eur J Nutr. 50:203–210.
2011.PubMed/NCBI View Article : Google Scholar
|
|
80
|
International Home Medical (IHM):
Plant-origin nano-particled (Pro)-Biogenics immunophilus SNK.
Plant-origin nano-sized lactic acid bacterium SNK®. IHM
Inc., Tolyo, 2021. https://www.ihmg.jp/nyusan_snk_en/. Accessed October
14, 2021.
|
|
81
|
Chen N, Xia P, Li S, Zhang T, Wang TT and
Zhu J: RNA sensors of the innate immune system and their detection
of pathogens. IUBMB Life. 69:297–304. 2017.PubMed/NCBI View Article : Google Scholar
|
|
82
|
Moreno-Eutimio MA, Lopez-Macias C and
Pastelin-Palacios R: Bioinformatic analysis and identification of
single-stranded RNA sequences recognized by TLR7/8 in the
SARS-CoV-2, SARS-CoV, and MERS-CoV genomes. Microbes Infect.
22:226–229. 2020.PubMed/NCBI View Article : Google Scholar
|
|
83
|
Slaats J, Ten Oever J, van de Veerdonk FL
and Netea MG: IL-1β/IL-6/CRP and IL-18/ferritin: Distinct
inflammatory programs in infections. PLoS Pathog.
12(e1005973)2016.PubMed/NCBI View Article : Google Scholar
|
|
84
|
Mick E, Kamm J, Pisco AO, Ratnasiri K,
Babik JM, Castañeda G, DeRisi JL, Detweiler AM, Hao SL, Kangelaris
KN, et al: Upper airway gene expression reveals suppressed immune
responses to SARS-CoV-2 compared with other respiratory viruses.
Nat Commun. 11(5854)2020.PubMed/NCBI View Article : Google Scholar
|
|
85
|
van der Made CI, Simons A,
Schuurs-Hoeijmakers J, van den Heuvel G, Mantere T, Kersten S, van
Deuren RC, Steehouwer M, van Reijmersdal SV, Jaeger M, et al:
Presence of genetic variants among young men with severe COVID-19.
JAMA. 324:663–673. 2020.PubMed/NCBI View Article : Google Scholar
|
|
86
|
Liu C, Martins AJ, Lau WW, Rachmaninoff N,
Chen J, Imberti L, Mostaghimi D, Fink DL, Burbelo PD, Dobbs K, et
al: Time-resolved systems immunology reveals a late juncture linked
to fatal COVID-19. Cell. 184:1836–1857.e22. 2021.PubMed/NCBI View Article : Google Scholar
|
|
87
|
Sahoo D, Katkar GD, Khandelwal S,
Behroozikhah M, Claire A, Castillo V, Tindle C, Fuller M, Taheri S,
Rogers TF, et al: AI-guided discovery of the invariant host
response to viral pandemics. EBioMedicine.
68(103390)2021.PubMed/NCBI View Article : Google Scholar
|