Introduction
Neural diseases and injuries, such as intracerebral
hemorrhage, spinal cord injuries and peripheral nerve injuries, may
be life-threatening, and often have detrimental impacts on
patients' daily life and work activities (1-3).
Various treatment strategies, including pharmacotherapy and
surgery, have been investigated; however, effective strategies are
still lacking due to inconsistent clinical benefits (1-3).
Stem cell-based transplantation therapy has notable therapeutic
potential, but acquisition of an adequate, ready-to-use stem cells
for transplantation remains a major challenge. Neural progenitor
(NPCs)/stem cells derived from aborted human fetuses and cadavers
may be ideal candidates, but it is impossible to obtain a
sufficient number of transplantable cells for clinical
applications. Furthermore, although human embryonic stem cells
possess a high tendency for neural differentiation, the ethical
issues limit their clinical application (4). Neural-lineage cells can easily
differentiate from induced pluripotent stem cells (iPSCs), but the
use of retroviruses to generate iPSCs causes safety concerns on
retroviral DNA integration into the host cell genome (5). Mesenchymal stem cells (MSCs) isolated
from bone marrow and adipose tissues also exhibit some neurogenic
differentiation potential (6-9),
but procuring MSCs from patients requires invasive surgical
procedures, and the neurogenic potential of these cells is
limited.
Dental tissue-derived stem cells have high
neurogenic potential because of their embryonic neural crest
origin, and may represent an alternative cell source for
neuroregeneration (10).
Furthermore, some dental stem cells were shown to express
pluripotency markers like NANOG and OCT4 that are not usually
expressed in other MSCs (11,12),
and their neuroregenerative potential has been corroborated by
in vivo studies on stroke (13) and neurodegenerative diseases
(14). Dental stem cells can be
easily collected from extracted teeth, cryopreserved and thawed
when required. Stem cells from apical papilla (SCAP) with strong
survival in harsh environments and high proliferative capacity
represent excellent seed cells for tissue engineering (15). In previous studies, SCAP
transplanted into the injured spinal cord or sciatic nerve
significantly improved nerve regeneration and exerted
neuroprotective effects (16,17).
It was reported that small molecules could drive the
neural reprogramming of somatic cells (18). A previous study applied the same
small molecule cocktail used by Hu et al (18) and successfully enhanced the
differentiation of dental pulp stem cells, SCAP and gingiva-derived
MSCs into neural-lineage cells within 14 days, and a longer
induction period leads to lower proliferation of the cells
(19). Therefore, the present study
aimed to optimize this induction period. Secondly, due to neural
crest origin, SCAP are considered to be more amenable to
reprogramming into neural-lineage cells compared with human
fibroblasts (10,20). To the best of our knowledge, the
present study was the first to investigate the early phase of SACP
differentiation into neurons. Based on the aforementioned studies,
7 days was chosen as the induction period. It was hypothesized that
small molecules, including valproic acid (VPA), CHIR99021, Repsox,
forskolin, SP600125, GO6983 and Y-27632, may exert rapid effects on
the neurogenic differentiation of SCAP, and that this process may
differ from that observed in human fibroblasts (18). The present study investigated the
differentiation of SCAP into neural-lineage cells within 7 days of
treatment with small molecules.
Materials and methods
Culture of SCAP
Human SCAP were gifted by Dr Anibal Diogenes
(Department of Endodontics, University of Texas Health Science
Center at San Antonio, TX, USA) (21). Cells were seeded in α-minimum
essential medium (α-MEM) containing 10% (v/v) fetal bovine serum
and 1% (v/v) penicillin-streptomycin antibiotic solution (all
Thermo Fisher Scientific, Inc.). Cultures were maintained at 37˚C
in a 5% CO2 incubator. The culture medium was changed
every 2-3 days, and cells were subcultured when they reached 80%
confluence.
Neural induction of SCAP
SCAP were seeded onto 6-well culture plates at a
density of 20,000 cells/cm2. The neural-induction
protocol used was based on a previous study (18) with some modifications, including
shortening the culture duration from 8 to 7 days. When the SCAP
reached 60% confluence, the medium was changed to neural-induction
medium (NIM) comprising of DMEM/F12/Neurobasal A at 1:1
supplemented with 0.5% (v/v) N2, 1% (v/v) B27 (all Thermo Fisher
Scientific, Inc.), 100 mM cAMP (Sigma-Aldrich; Merck KGaA) and 20
ng/ml basic fibroblast growth factor (Thermo Fisher Scientific,
Inc.), in the absence or presence of the chemical cocktail VCRFSGY
(0.5 mM VPA, 3 µM CHIR99021, 1 µM Repsox, 10 µM forskolin, 10 µM
SP600125, 5 µM GO6983 and 5 µM Y-27632) (NIMS). All of these small
molecules were obtained from Sigma-Aldrich; Merck KGaA. The culture
medium was refreshed on day 3. Cells were examined using light
microscopy at a magnification of x20 or harvested for western
blotting, immunocytofluorescence, reverse
transcription-quantitative (RT-q)PCR and electrophysiological
assays.
Western blotting
Cells were lysed by adding M-PER solution (Thermo
Fisher Scientific, Inc.) containing protease inhibitor cocktail 4˚C
for 20 min. The resulting protein samples were quantified using a
BCA kit (Thermo Fisher Scientific, Inc.). Equal amounts of protein
(25 µg) were loaded per lane onto a 7.5% gel, resolved using
SDS-PAGE and subsequently transferred onto Immun-Blot PVDF
membranes (Cyvita). The membranes were blocked with 5% (w/v)
skimmed milk for 1 h and incubated overnight at 4˚C with primary
antibodies against Nestin (1:100; cat. no. ab105389; Abcam),
paired-box gene 6 (Pax6; 1:1,000; cat. no. 60433; Cell Signaling
Technology, Inc.), Sry-related HMG box 2 (Sox2; 1:1,000; cat. no.
3579S; Cell Signaling Technology, Inc.), neurofilament protein
(NFM; 1:1,000; cat. no. RMO-270; Invitrogen; Thermo Fisher
Scientific, Inc.), neuron-specific nuclear protein (NeuN; 1:1,000;
cat. no. 702022; Invitrogen; Thermo Fisher Scientific, Inc.) and
microtubule-associated protein 2 (MAP2; 1:500; cat. no. 13-1500;
Invitrogen; Thermo Fisher Scientific, Inc.). After washing with
Tris-buffered saline/0.1% Tween-20, the membranes were incubated
with horseradish peroxidase-conjugated anti-mouse IgG or
horseradish peroxidase-conjugated anti-rabbit IgG (both Cell
Signaling Technology, Inc.) secondary antibodies for 2 h at room
temperature. The antibody-antigen complexes were visualized using
Pierce ECL Western Blotting Substrate (Thermo Fisher Scientific,
Inc.). Relative densities quantified using ImageJ software (v1.6.0;
National Institute of Health).
Immunocytofluorescence
Cells were fixed with 4% (v/v) paraformaldehyde at
room temperature for 20 min, washed three times with
phosphate-buffered saline (PBS), permeabilized with 0.5% (w/v)
Triton X-100 for 5 min at 4˚C, and blocked in PBS containing 5%
(w/v) bovine serum albumin (Sigma-Aldrich; Merck KGaA) at room
temperature for 1 h. The fixed samples were incubated with primary
antibodies against Nestin (1:100; cat. no. ab105389; Abcam), Pax6
(1:200; cat. no. 60433; Cell Signaling Technology, Inc.) and Sox2
(1:400; cat. no. 3579S; Cell Signaling Technology, Inc.) overnight
at 4˚C. After removal of excess primary antibodies by
washing with PBS, the samples were incubated with Alexa Fluor
488-conjugated goat anti-mouse or tetramethylrhodamine-conjugated
goat anti-rabbit (both Abcam) secondary antibodies for 1 h at room
temperature in the dark, followed by washing with PBS. The cell
nuclei were stained with DAPI for 5 min at room temperature and the
samples were imaged by laser scanning microscopy (LSM710; Carl
Zeiss AG) at x20 magnification at specific excitation/emission
wavelengths for TRITC (540/570 nm) and Alexa Fluor 488 (490/520
nm).
RT-qPCR
Total RNA was extracted from cultured SCAP using an
RNeasy Plus Mini kit (Qiagen, Inc.). The extracted RNA was reverse
transcribed to cDNA using the SuperScript®
VILO® Master Mix (Thermo Fisher Scientific, Inc.)
according to the manufacturer's instructions. The reaction was
incubated at 25˚C for 10 min, followed by
42˚C for 60 min, and terminated at 85˚C for 5
min. RT-qPCR was carried out using a StepOne Real-Time PCR system
(Applied Biosystems; Thermo Fisher Scientific, Inc.) and
SYBR™ Green Master Mix (Thermo Fisher Scientific, Inc.).
The primer sequences for the neural markers are shown in Table I. The 2-ΔΔCq method
(22) was used to calculate the
Cq-value for each gene and target gene expression levels were
normalized to the housekeeping gene GAPDH. Analysis of all gene
markers was repeated three times. RT-qPCR amplifications were
carried out using the following parameters: 2 min At 50˚C, 20 sec
at 95˚C, 40 cycles of 3 sec at 95˚C, 30 sec at 60˚C, 15 sec at 95˚C
and 1 min at 60˚C.
 | Table IPrimer sequences utilized for
RT-qPCR. |
Table I
Primer sequences utilized for
RT-qPCR.
| Gene | Primer sequence,
5'-3' |
|---|
| NFM | |
|
Forward |
GTCAAGATGGCTCTGGATATAGAAATC |
|
Reverse |
TACAGTGGCCCAGTGATGCTT |
| MAP2 | |
|
Forward |
TTGGTGCCGAGTGAGAAGAA |
|
Reverse |
GGTCTGGCAGTGGTTGGTTAA |
| GAPDH | |
|
Forward |
TGCACCACCAACTGCTTAGC |
|
Reverse |
GGCATGGACTGTGGTCATGAG |
Cell proliferation assay
SCAP were seeded into 96-well plates at a density of
10,000 cells/well and preincubated at 37˚C under 5%
CO2 for 24 h. The original culture medium was discarded
and replaced with α-MEM, neural-induction medium or
neural-induction medium with small molecules. The samples were
analyzed on days 1, 3, 5 and 7 using a Cell Counting Kit (CCK)-8
cell proliferation assay kit (Dojindo Molecular Technologies, Inc.)
according to the manufacturer's instructions. Briefly, CCK-8
solution (10 µl) was added to each well and incubated for 3 h at
37˚C under 5% CO2. Subsequently, the absorbance was
measured at 450 nm using a SpectraMax® M2 microplate
reader (Molecular Devices, LLC).
Electrophysiological assay
Whole-cell patch-clamp recordings were conducted on
the basis of a previously published method by Li et al
(23). The cells were bathed with a
solution containing (in mM): 145 NaCl, 1.5 KCl, 2 CaCl2,
2 MgCl2, 10 HEPES And 10 glucose (pH adjusted to 7.4
with NaOH). The patch-clamp pipettes had a resistance of 6-8 MΩ
with an internal solution containing (in mM): 125 KCl, 1
CaCl2, 2 MgCl2, 2 Mg-ATP, 2
Na2ATP, 10 HEPES And 10 EGTA (pH adjusted to 7.2 with
KOH). The recordings were acquired using a A&M amplifier (Model
2400; A-M Systems, Inc.). Data were filtered at 4 kHz and analyzed
with pCLAMP version 10.4 software (Axon Instruments). All
experiments were performed at room temperature (22-25˚C). For
voltage-clamp experiments, the cells were clamped at -80 mV for 400
ms and depolarized for 40 ms with voltage pulses from -80 to 200 mV
at 20 mV intervals. Na+ and K+ current
amplitudes were measured at the peak outward and inward values,
respectively, and reported as current densities (pA/pF) for
comparison. The Na+ and K+ currents were
selectively blocked by 1 µM tetrodotoxin (TTX) and 35 mM tetraethyl
ammonium (TEA), respectively. For current-clamp experiments, the
cells were set at -80 mV holding potential, and the action
potentials were elicited by step current injection of 100-300 pA
for 1,000 ms.
Statistical analysis
All experiments were conducted in triplicate.
Differences between SCAP were cultured in neural-induction medium
(NIM-SCAP) and cultured in neural-induction medium with small
molecules (NIMS-SCAP) were analyzed using an unpaired Student's
t-tests, while differences among multiple cell groups were examined
using a Tukeys post-hoc with a One-way ANOVA. The data were
expressed as mean ± standard error of the mean. P<0.05 was
considered to indicate a statistically significant difference. All
statistical analyses were carried out using SPSS version 19.0
software (IBM Corp.).
Results
Morphological changes during neural
induction
SCAP (Fig. 1A) were
subjected to different neural-induction culture protocols over a
period of 7 days. The cells were observed using phase-contrast
light microscopy every 15-30 min during the first 12 h, and then on
days 1, 3, 5 and 7. Morphological changes were discernible by 30
min in NIMS-SCAP (Fig. 1C), but
only after 24 h in NIM-SCAP (Fig.
1D). The major initial changes included the appearance of an
elongated morphology and bipolar neurites (Fig. 1C-K). NIMS-SCAP exhibited marked
differences in cellular morphology compared with NIM-SCAP from days
1-7, including adoption of a rounded shape with a greater number of
and longer neurite outgrowths after treatment with the small
molecule cocktail. In contrast, NIM-SCAP mostly retained the
morphological characteristics of MSCs, except for the appearance of
neurites at the poles, with the entire cells being spindle-shaped.
In addition, NIMS-SCAP neurites tended to become elongated until
they connected with the neurites of adjacent cells.
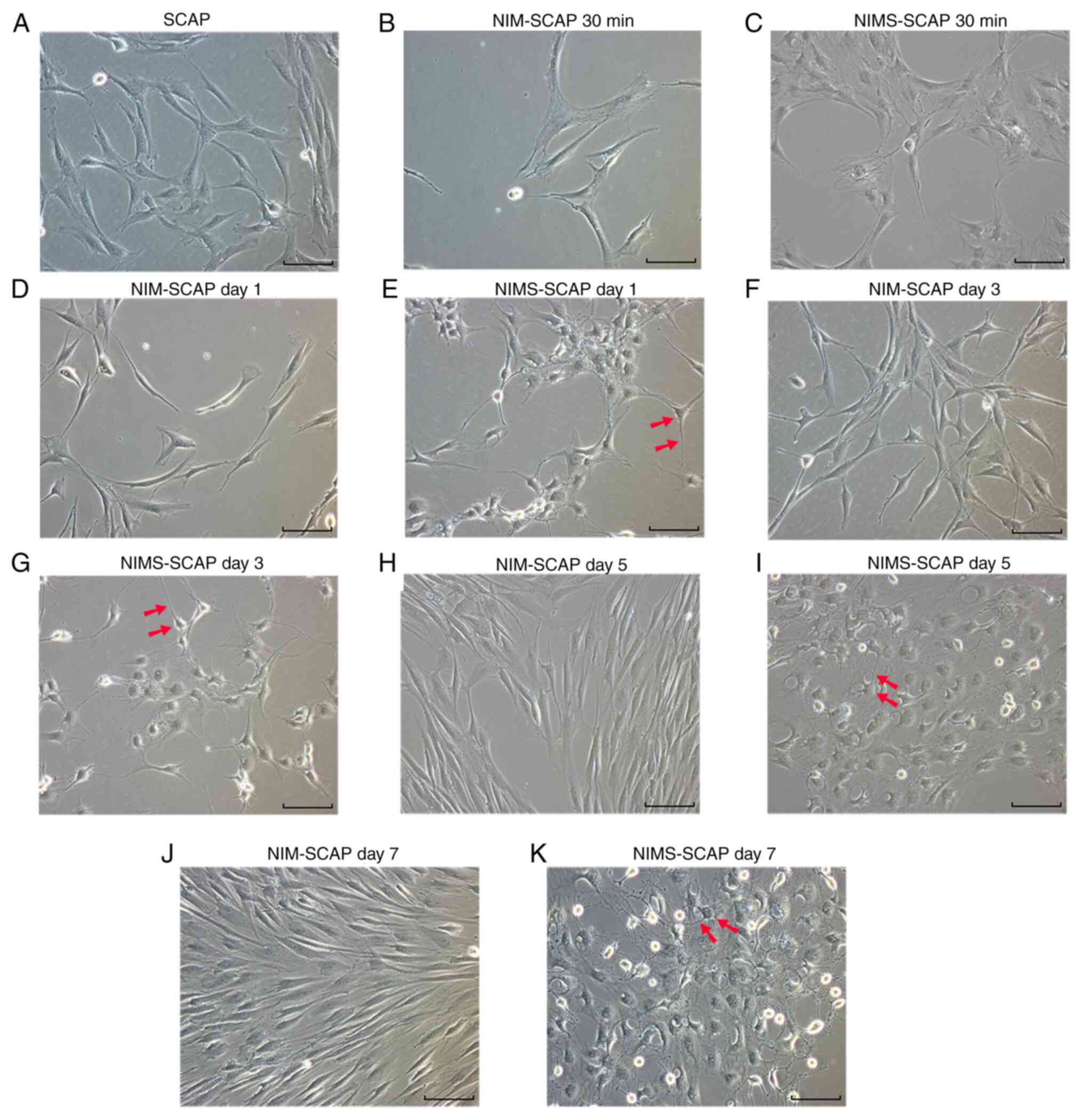 | Figure 1Morphological changes during neural
induction. (A) SCAP morphology. NIM-SCAP morphology at (B) 30 min,
days (D) 1, (F) 3, (H) 5 and (J) 7. NIMS-SCAP morphology at (C) 30
min, days (E) 1, (G) 3, (I) 5 and (K) 7. Scale bar, 100 µm;
magnification, x20. Arrows, cell body and neurite. SCAP, apical
papilla; NIM, neural-induction medium; NIMS, neural-induction
medium with small molecules. |
Variation in the expression of
NPC-related protein markers
The protein expression levels of NPC-related markers
in NIM-SCAP and NIMS-SCAP were analyzed using western blotting on
days 1, 3, 5 and 7 (Fig. 2A and
B). Nestin, Pax6 and Sox2 showed
trends toward initial upregulation, followed by subsequent
decreases, regardless of the presence of small molecules. In
addition, all three NPC-related proteins peaked on day 3 under both
neural-induction culture conditions. These results suggested that
SCAP were induced into a NPC-like cell state at day 3, but that
this state was unstable and short-lived, with the expression levels
of NPC-related proteins decreasing over the following 4 days.
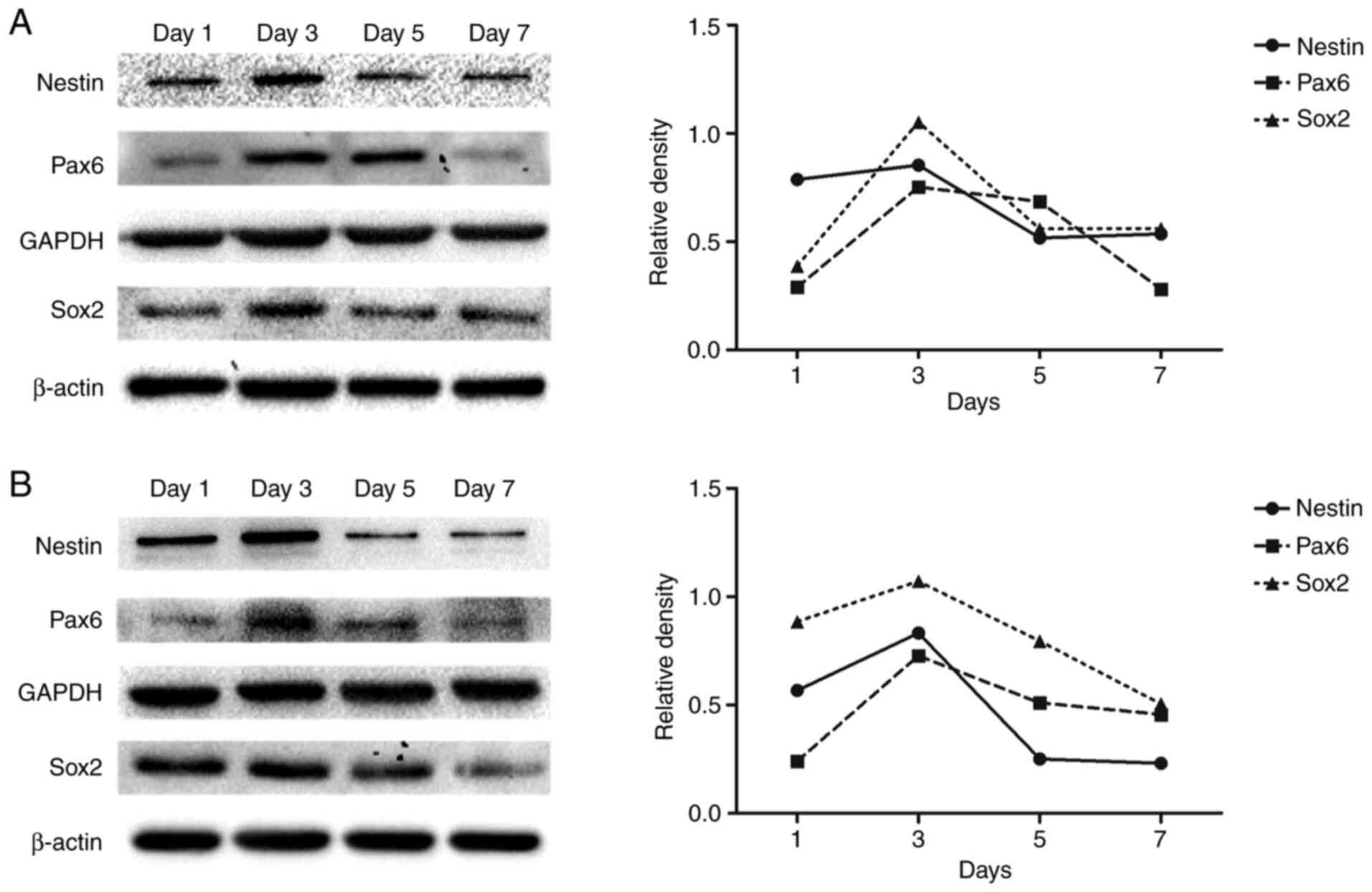 | Figure 2Variation in the expression of
NPC-related protein markers. (A) Detection of Nestin, Pax6 and Sox2
protein expression levels in NIM-SCAP by western blotting on days
1, 3, 5 and 7. Corresponding line plots show the relative densities
normalized to GAPDH (Nestin and Pax6) or β-actin (Sox2) as
endogenous controls. (B) Detection of Nestin, Pax6 and Sox2 protein
expression levels in NIMS-SCAP by western blotting on days 1, 3, 5
and 7. Corresponding line plots show the relative densities
normalized to GAPDH (Nestin and Pax6) or β-actin (Sox2) as
endogenous controls. SCAP, apical papilla; NIM, neural-induction
medium; NIMS, neural-induction medium with small molecules; NPC,
neural progenitor cell; Pax6, paired-box gene 6; Sox2, Sry-related
HMG box 2. |
NPC-related protein expression during
the NPC-like cell state
Both NIM-SCAP and NIMS-SCAP exhibited increased
Nestin (Fig. 3A-C), Pax6 (Fig. 3D-F) and Sox2 (Fig. 3G-I) expression on day 3 compared
with undifferentiated SCAP, according to qualitative
immunocytofluorescence analysis. Nestin was expressed relatively
strongly in SCAP, and thus there were minimal differences in the
staining intensities compared with NIM-SCAP and NIMS-SCAP.
Meanwhile, immunofluorescence staining of Pax6 and Sox2 was
strongly enhanced in NIM-SCAP and NIMS-SCAP after induction, most
notably Sox2 expression in NIMS-SCAP.
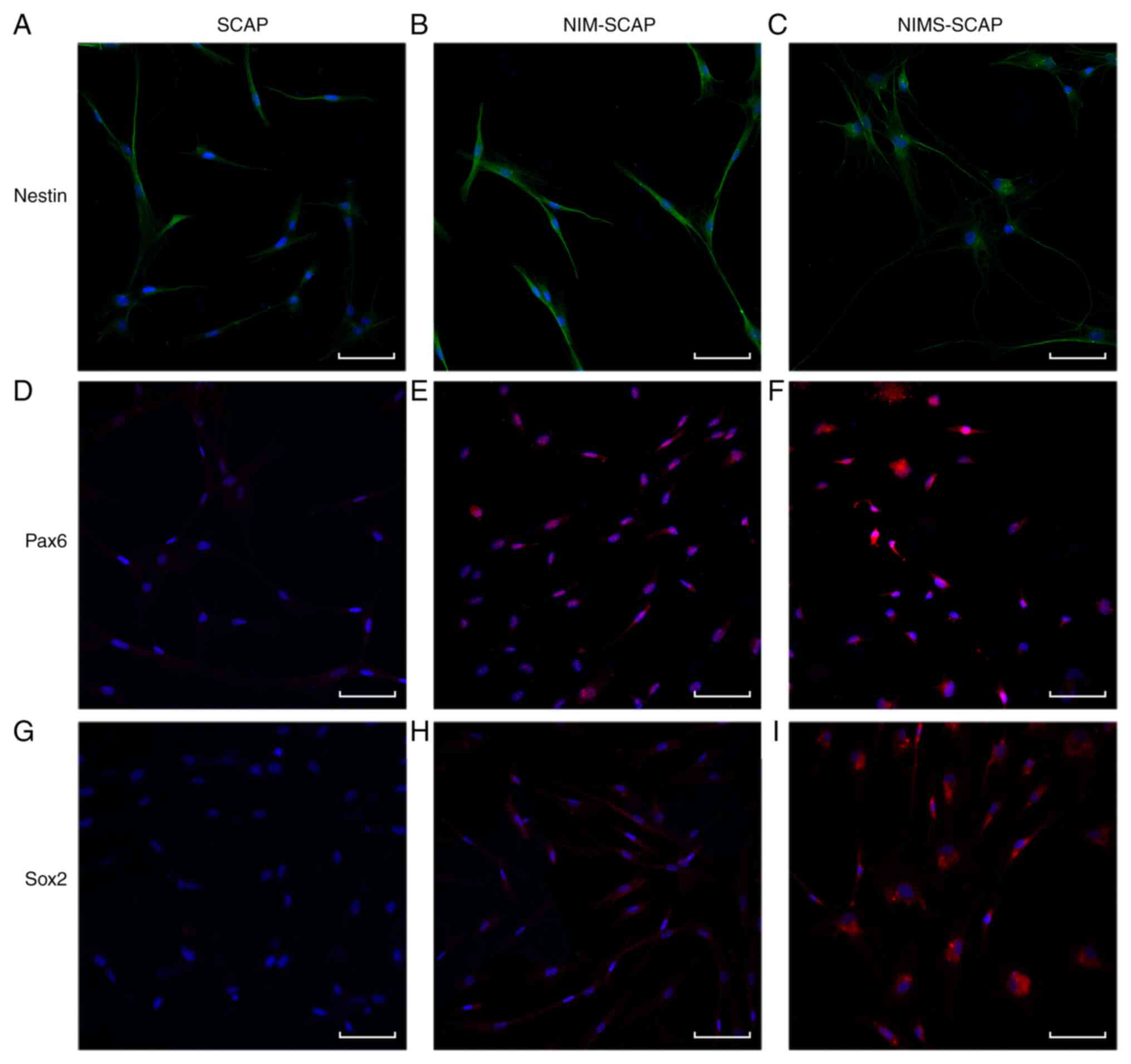 | Figure 3NPC-related protein expression during
the NPC-like cell state. (A-C) Immunocytofluorescence for detection
of Nestin expression in SCAP, NIM-SCAP and NIMS-SCAP at day 3.
(D-F) Immunocytofluorescence for detection of Pax6 expression in
SCAP, NIM-SCAP and NIMS-SCAP at day 3. (G-I) Immunocytofluorescence
for detection of Sox2 expression in SCAP, NIM-SCAP and NIMS-SCAP at
day 3. Nuclei were counterstained with DAPI (blue). Scale bar, 100
µm; magnification, x200. SCAP, apical papilla; NIM,
neural-induction medium; NIMS, neural-induction medium with small
molecules; NPC, neural progenitor cell; Pax6, paired-box gene 6;
Sox2, Sry-related HMG box 2. |
Expression of differentiated
neuron-related markers
NFM, NeuN and MAP2 expression levels were
significantly upregulated in NIMS-SCAP compared with NIM-SCAP
(Fig. 4B-D, respectively), while
these differentiated neuron markers were hardly detectable in
undifferentiated SCAP (Fig. 4A).
The gene expression levels of the neuron markers in NIM-SCAP and
NIMS-SCAP were analyzed on days 1, 3, 5 and 7 using RT-qPCR
(Fig. 4E and F). NFM and MAP2 gene expression levels
were signifincantly increased on days 1-5 in both NIM-SCAP and
NIMS-SCAP. NFM expression continued to increase until day 7 in
NIMS-SCAP, while MAP2 expression peaked on day 5 and was decreased
on day 7, but remained higher compared with that on day 3.
Meanwhile, NFM and MAP2 gene expression levels were only slightly
increased in NIM-SCAP. NFM and MAP2 gene expression levels were
significantly higher in NIMS-SCAP compared with NIM-SCAP on both
days 5 and 7.
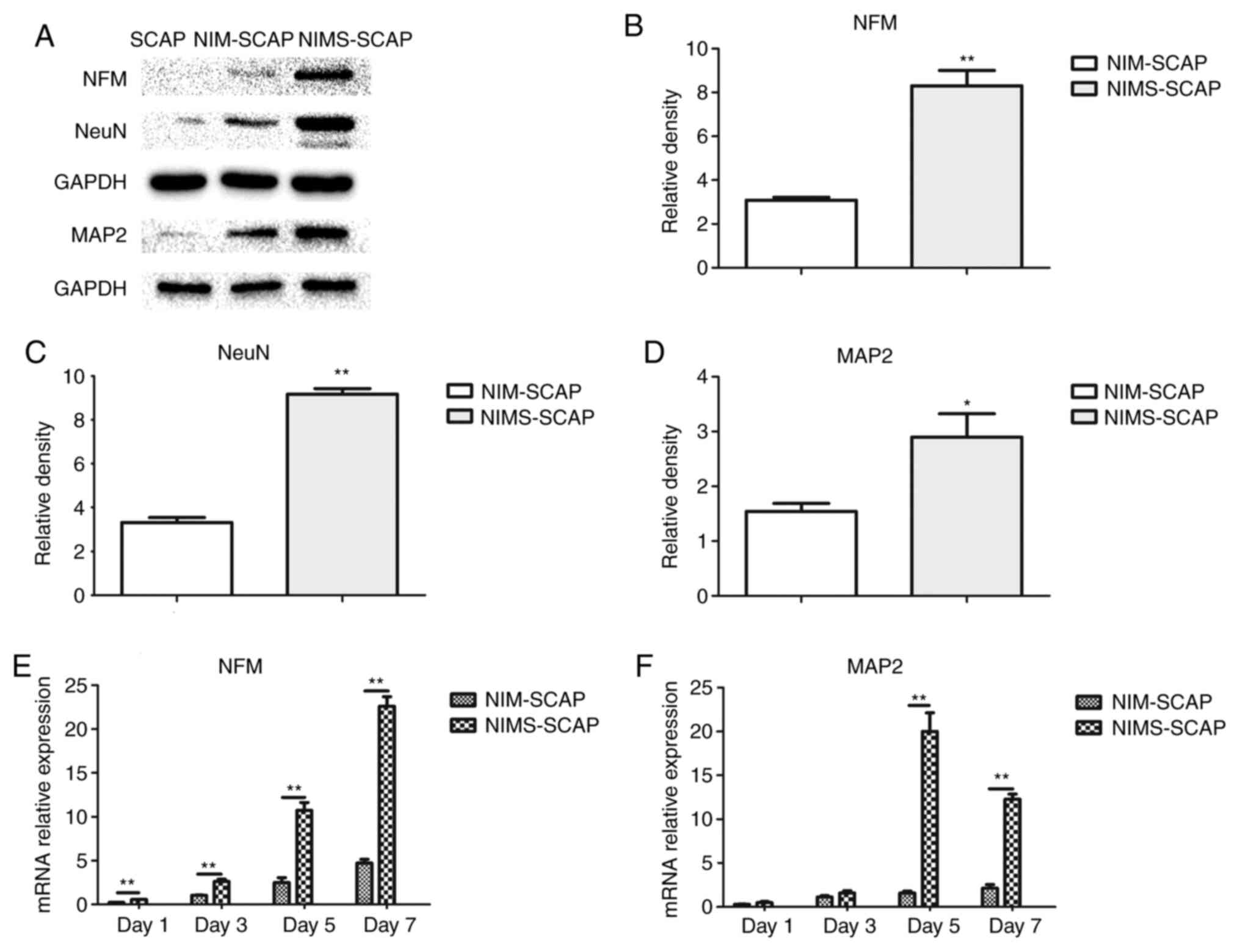 | Figure 4Expression of differentiated
neuron-related markers. (A) Detection of NFM, NeuN and MAP2 protein
expression levels in SCAP, NIM-SCAP and NIMS-SCAP on day 7 using
western blotting. Relative densities quantified using ImageJ for
(B) NFM (n=3), (C) NeuN (n=3) and (D) MAP2 (n=3). (E) NFM gene
expression levels in NIM-SCAP and NIMS-SCAP on days 1, 3, 5 and 7
determined using RT-qPCR (n=6). (F) MAP2 gene expression levels in
NIM-SCAP and NIMS-SCAP on days 1, 3, 5, and 7 determined by RT-qPCR
(n=6). All data normalized to SCAP, which were assigned an
arbitrary value of 1. *P<0.05,
**P<0.01. NFM, neurofilament protein; NeuN,
neuron-specific nuclear protein; MAP2, microtubule-associated
protein 2; SCAP, apical papilla; NIM, neural-induction medium;
NIMS, neural-induction medium with small molecules; RT-qPCR,
reverse transcription-quantitative PCR. |
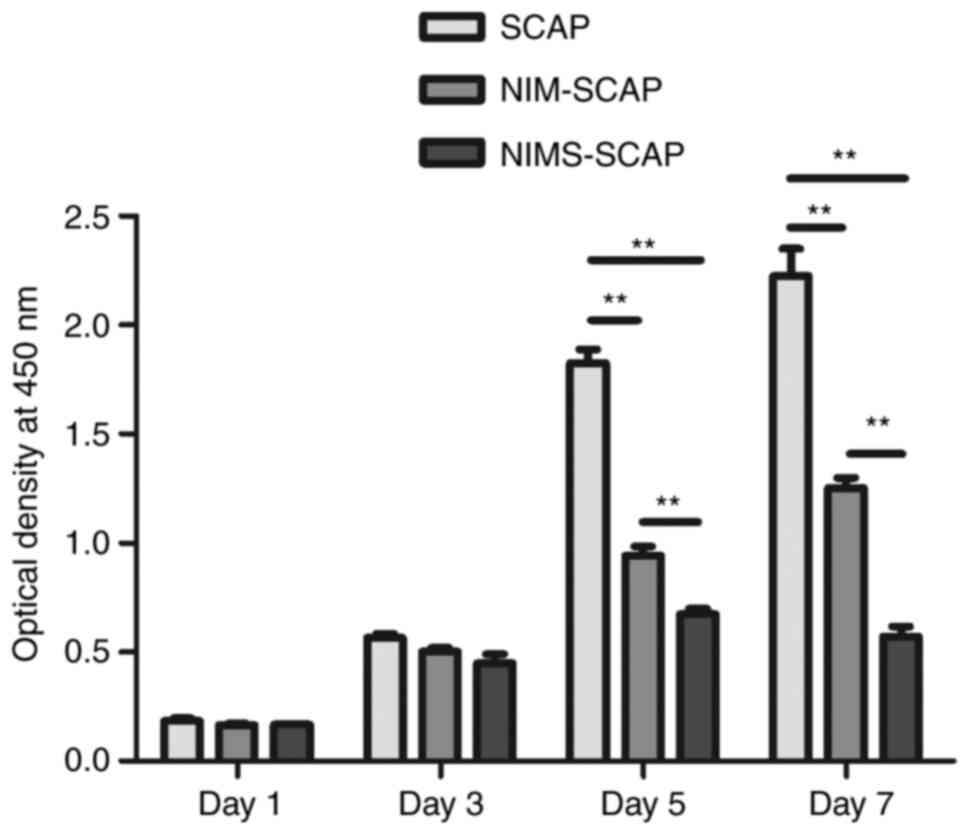
Cell proliferation assay
NIMS-SCAP proliferated more slowly compared with
NIM-SCAP and SCAP, as evaluated using CCK-8 assays, but there were
no significant differences in proliferation of SCAP, NIM-SCAP and
NIMS-SCAP on days 1 and 3 (Fig. 5).
Undifferentiated SCAP proliferated rapidly from days 3-5, while the
proliferation rates of NIM-SCAP and NIMS-SCAP slowed down. The
proliferation rate was slowest in NIMS-SCAP and the proliferation
had ceased by day 7. Some cells failed to survive until day 7.
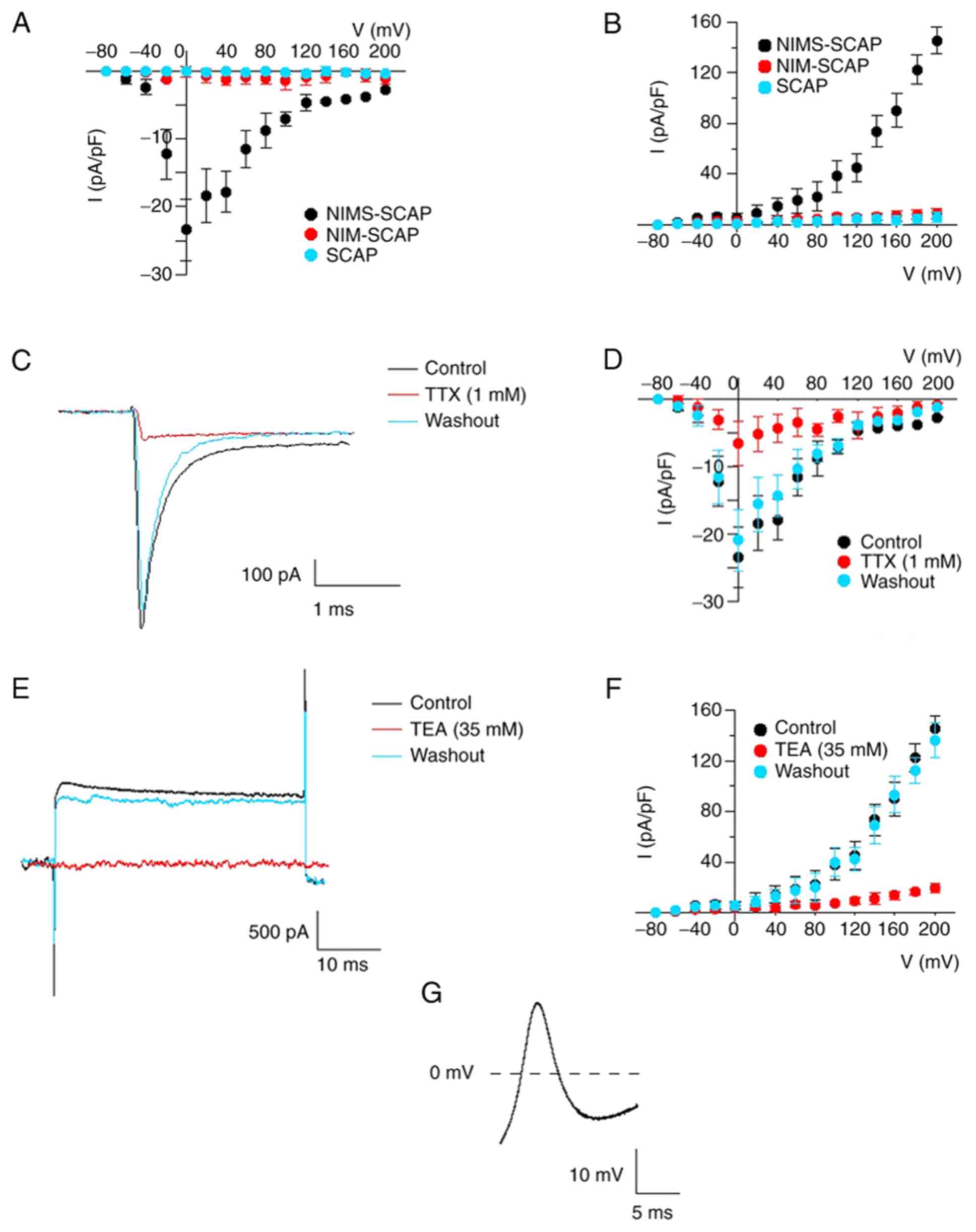
Electrophysiological functions
The cells that had a neuron-like morphology were
selected. Whole-cell patch-clamp studies were performed to measure
the voltage-dependent Na+ and K+ currents in
the cells. The results indicated that there were voltage-dependent
Na+ and K+ currents in NIMS-SCAP
(Na+ currents 18/45, K+ currents 16/45), and
not in NIM-SCAP and SCAP (Fig. 6A
and B). The inward Na+
currents were reversibly blocked by 1 µM TTX and rapidly recovered
with washing off TTX (Fig. 6C and
D). Similarly, the outward
K+ current was reversibly blocked by 35 mM TEA and
rapidly recovered with washing off TEA (Fig. 6E and F). A single action potential (AP) was
observed in a subset of NIMS-SCAP (n=3; AP 5/45; Fig. 6G), without any observed spontaneous
AP activity during the current clamp experiments.
Discussion
A select combination of small molecules has been
shown to reprogram human dermal fibroblasts into MSCs (24), and to reprogram mouse fibroblasts
into NPCs (25), functional neurons
(26), cardiomyocytes (27) and endothelial cells (28). Chemical-based strategies, which
avoid the use of viral vectors, transgenic manipulation or gene
modification, represent safe methods for the generation of
clinically relevant cell lineages for regenerative transplantation
and disease treatment (29-31).
Furthermore, small molecules can exert transient, reversible and
dose-dependent effects on the targeted cells, thereby allowing the
timing and dosage to be precisely controlled and fine-tuned
(29). Chemical-based approaches
therefore represent a new paradigm and viable alternative to
gene-based approaches for cell reprogramming.
A diverse variety of small-molecule chemicals have
previously been utilized for cellular reprogramming (30-34).
These chemicals can be divided into several categories based on
their effects on cell physiology and signaling pathways: Histone
deacetylase inhibitors, such as VPA; signaling pathway inhibitors,
such as inhibitors of transforming growth factor (TGF)-β signaling
(Repsox), glycogen synthase kinase (GSK)-3β (CHIR99021), protein
kinase C (GO6983) and ROCK (Y-27632) and adenylate cyclase
activators, such as forskolin, and SP600125, which was reported to
improve the reprogramming efficiency of overexpressed exogenous
transcription factors (35).
Therefore, the present study examined the effect of the chemical
cocktail VCRFSGY (0.5 mM VPA, 3 µM CHIR99021, 1 µM Repsox, 10 µM
forskolin, 10 µM SP600125, 5 µM GO6983, 5 µM Y-27632) on SCAP
reprogramming.
Changes in cell morphology were observed as early as
30 min after application of the small molecules, compared with no
changes until 24 h in the NIM-SCAP. The morphology of NIM-SCAP
remained similar to that of SCAP for the first 7 days of
neural-induction culture, with some residual adult MSCs memory. In
contrast to this, NIMS-SCAP were rapidly transformed into a
neural-like phenotype with a larger number of and longer neurite
outgrowths that became connected to one another to form a web-like
network. Y-27632 was reported to promote neurite outgrowth in
olfactory ensheathing cells (OECs) by inhibiting the Rho-associated
coiled-coil-containing protein kinase (ROCK)/F-actin pathway
Rho-associated protein kinase signaling pathway, and TGF-β1
shortened the length of the neurites and decreased cell elongation
(36). In addition, TGF-β1
treatment reprogrammed OECs into a flattened morphology (37). In the present study, the effects of
Repsox on the TGF-β signaling specifically were not demonstrated.
Repsox is a known inhibitor of the TGF-β1 signaling pathway,
therefore it was hypothesized that this inhibitor contributed to
the enhanced neurite outgrowth and elongation of NIMS-SCAP in the
present study.
NPCs, as a nascent neural-lineage cell type, can
usually be identified by the expression of upstream neural markers,
such as Nestin (38), Pax6(39) and Sox2(40). The present study demonstrated
time-dependent changes in the expression of these NPC-related
proteins in NIM-SCAP and NIMS-SCAP, with increases during the early
phase of neural induction followed by subsequent downregulation.
These results suggested that SCAP may be induced by small molecules
into an NPC-like cell state by day 3, followed by further
differentiation into neuron-like cells. The NPC self-renewal marker
Nestin (38) was expressed at a
lower level in NIMS-SCAP compared with NIM-SCAP, suggesting that
NIMS-SCAP progressed to a more mature neural stage more rapidly.
The immunocytofluorescence results revealed that Sox2 expression
was more prominent in NIMS-SCAP compared with in NIM-SCAP or SCAP.
Both VPA and GO6983 have previously been shown to induce cells into
a more plastic state (30,32), while Sox2 expression levels in iPSCs
treated with VPA were similar to those in embryonic stem cells
(30). Moreover, VPA, Repsox and
CHIR99021 inhibited histone deacetylases, while TGF-β and GSK-3
activated the expression of endogenous Sox2 (25,30,41-43).
In contrast to the present results, the immunostaining analysis
reported Hu et al (18), who
formulated the small molecule cocktail-based induction protocol
used in the present study, showed no expression of neural
progenitor markers (Nestin, Pax6 or Sox2) during the induction
procedure. These differences may be due to the cell types used in
both studies, for example SCAP have higher neurogenic potential
compared with human fibroblasts (10). Even without any neural induction,
SCAP can also express Nestin due to its neural crest origin
(10).
In the present study, the expression levels of
neural progenitor markers continued to decline on days 5-7,
therefore it was hypothesized that the induced SCAP were
progressing further along the neural differentiation pathway. The
expression levels of differentiated neural markers in the cells
were compared on day 7 using western blotting. NFM, NeuN and MAP2,
as markers of differentiated neurons (44-46),
were significantly upregulated, demonstrating that the small
molecules dramatically accelerated neural-lineage differentiation.
There was a consistent increase in gene expression in NIMS-SCAP on
days 1-5, and NFM expression was detected up to day 7. MAP2
expression declined on day 7 compared with day 5; however, it
remained at a much higher level compared with that in NIM-SCAP.
Previously, the combination of GSK-3β inhibition (CHIR99021) and
adenylate cyclase signaling activation (forskolin) has been shown
to improve neuronal reprogramming efficiency (47), and CHIR99021 and Repsox increases
the neuronal conversion of human fibroblasts via overexpression of
Achaete-scute homolog 1 and neurogenin 2(31). In the present study, small molecules
dramatically decreased the duration required for differentiation
into neural-lineage cells.
The proliferation of NIMS-SCAP slowed significantly
on day 5 compared with that of NIM-SCAP, as shown by the CCK-8
assays. Moreover, some cells failed to survive until day 7. Taken
together with the presence of differentiated neuron markers, these
results suggested that stemness was decreased after 5 days of
induction with small molecules, thus preventing stem/progenitor
cells from undergoing self-renewal and proliferation, given that
neurons are highly differentiated and mitotically inactive cells
(19). In contrast to the present
results, Hu et al (18)
found that the majority of induced neuronal cells survived for
10-12 days, which could suggest that differentiated neuron-like
cells are in a mitotically inactive state after small-molecule
induction.
Besides morphological changes and upregulation of
protein and gene expression, the most important characteristics of
neuron is its electrophysiological activities (23). The result of whole-cell patch-clamp
studies revealed that NIMS-SCAP resulted in voltage-dependent
Na+ and K+ currents, which were blocked by
TTX and TEA and restored after the blocker was washed away.
Moreover, a single action potential was observed. These results
suggested that some of the cells have been transformed into mature
neurons. These results were consistent with previous studies
reporting that dental-derived stem cells differentiated into
neuron-like cells (23,48). In previous studies, neurosphere
culture was used to promote the neurogenic differentiation of
cells, which is also widely utilized as a classical method to
induce and amplify neural lineage cells (25,49).
By using the method of neurosphere culture, neuron maturation takes
~1 month (23), while induction by
small molecules significantly reduces the time for neuron
differentiation to have electrophysiological functions. The
combination of small molecules and neurosphere culture can identify
spontaneous cell postsynaptic currents (23).
In summary, the present study demonstrated that
small molecules can rapidly induce SCAP into an NPC-like state,
which are then easily differentiated into neuron-like cells. Cell
proliferation decreased in line with the downregulation of
NPC-related marker expression and ceased by day 5, indicating that
differentiated neuron-like cells had been generated. This was the
first study to efficiently induce SCAP into NPC. The translational
application of NPC derived from SCAP for nerve regeneration and
repair needs to be further studied by animal models.
Acknowledgements
Not applicable.
Funding
Funding: This work was funded by The Health and Medical Research
Fund (Hong Kong) - Full Grant (grant no. 06172396; to CZ).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
PW and CZ designed the experiments. QC, SJ, TZ and
ZS performed the experiments. QC, CY and PW analyzed the data. QC
and BCH analyzed the data and wrote the manuscript. CY, PW, and CZ
revised the manuscript. All authors read and approved the final
manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Wilkinson DA, Pandey AS, Thompson BG, Keep
RF, Hua Y and Xi G: Injury mechanisms in acute intracerebral
hemorrhage. Neuropharmacology. 134:240–248. 2018.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Hurlbert RJ, Hadley MN, Walters BC, Aarabi
B, Dhall SS, Gelb DE, Rozzelle CJ, Ryken TC and Theodore N:
Pharmacological therapy for acute spinal cord injury. Neurosurgery.
72 (Suppl 2):S93–S105. 2013.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Panagopoulos GN, Megaloikonomos PD and
Mavrogenis AF: The present and future for peripheral nerve
regeneration. Orthopedics. 40:e141–e156. 2017.PubMed/NCBI View Article : Google Scholar
|
|
4
|
McComish SF and Caldwell MA: Generation of
defined neural populations from pluripotent stem cells. Philos
Trans R Soc Lond B Biol Sci. 373(20170214)2018.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Chambers SM, Fasano CA, Papapetrou EP,
Tomishima M, Sadelain M and Studer L: Highly efficient neural
conversion of human ES and iPS cells by dual inhibition of SMAD
signaling. Nat Biotechnol. 27:275–280. 2009.PubMed/NCBI View
Article : Google Scholar
|
|
6
|
Wang Y, Li ZW, Luo M, Li YJ and Zhang KQ:
Biological conduits combining bone marrow mesenchymal stem cells
and extracellular matrix to treat long-segment sciatic nerve
defects. Neural Regen Res. 10:965–971. 2015.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Zhao H, Cheng L, Du X, Hou Y, Liu Y, Cui Z
and Nie L: Transplantation of cerebral dopamine neurotrophic factor
transducted BMSCs in contusion spinal cord injury of rats:
Promotion of nerve regeneration by alleviating neuroinflammation.
Mol Neurobiol. 53:187–199. 2016.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Georgiou M, Golding JP, Loughlin AJ,
Kingham PJ and Phillips JB: Engineered neural tissue with aligned,
differentiated adipose-derived stem cells promotes peripheral nerve
regeneration across a critical sized defect in rat sciatic nerve.
Biomaterials. 3:242–251. 2015.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Hsueh YY, Chang YJ, Huang TC, Fan SC, Wang
DH, Chen JJ, Wu CC and Lin SC: Functional recoveries of sciatic
nerve regeneration by combining chitosan-coated conduit and
neurosphere cells induced from adipose-derived stem cells.
Biomaterials. 35:2234–2244. 2014.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Parisi L and Manfredi E: Applicability of
tooth derived stem cells in neural regeneration. Neural Regen Res.
11:1704–1707. 2016.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Lima RL, Holanda-Afonso RC, Moura-Neto V,
Bolognese AM, DosSantos MF and Souza MM: Human dental follicle
cells express embryonic, mesenchymal and neural stem cells markers.
Arch Oral Biol. 73:121–128. 2017.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Athanassiou-Papaefthymiou M, Papagerakis P
and Papagerakis S: Isolation and characterization of human adult
epithelial stem cells from the periodontal ligament. J Dent Res.
94:1591–1600. 2015.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Yamagata M, Yamamoto A, Kako E, Kaneko N,
Matsubara K, Sakai K, Sawamoto K and Ueda M: Human dental
pulp-derived stem cells protect against hypoxic-ischemic brain
injury in neonatal mice. Stroke. 44:551–554. 2013.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Shamir C, Venugopal C and Dhanushkodi A:
Dental pulp stem cells for treating neurodegenerative diseases.
Neural Regen Res. 10:1910–1911. 2015.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Huang GT, Sonoyama W, Liu Y, Liu H, Wang S
and Shi S: The hidden treasure in apical papilla: The potential
role in pulp/dentin regeneration and bioroot engineering. J Endod.
34:645–651. 2008.PubMed/NCBI View Article : Google Scholar
|
|
16
|
De Berdt P, Vanacker J, Ucakar B, Elens L,
Diogenes A, Leprince JG, Deumens R and des Rieux A: Dental apical
papilla as therapy for spinal cord injury. J Dent Res.
94:1575–1581. 2015.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Kolar MK, Itte VN, Kingham PJ, Novikov LN,
Wiberg M and Kelk P: The neurotrophic effects of different human
dental mesenchymal stem cells. Sci Rep. 7(12605)2017.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Hu W, Qiu B, Guan W, Wang Q, Wang M, Li W,
Gao L, Shen L, Huang Y, Xie G, et al: Direct conversion of normal
and Alzheimer's disease human fibroblasts into neuronal cells by
small molecules. Cell Stem Cell. 17:204–212. 2015.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Heng BC, Jiang S, Yi B, Gong T, Lim LW and
Zhang C: Small molecules enhance neurogenic differentiation of
dental-derived adult stem cells. Arch Oral Biol. 102:26–38.
2019.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Alt E, Yan Y, Gehmert S, Song YH, Altman
A, Gehmert S, Vykoukal D and Bai X: Fibroblasts share mesenchymal
phenotypes with stem cells, but lack their differentiation and
colony-forming potential. Biol cell. 103:197–208. 2011.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Ruparel NB, de Almeida JF, Henry MA and
Diogenes A: Characterization of a stem cell of apical papilla cell
line: Effect of passage on cellular phenotype. J Endod. 39:357–363.
2013.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408.
2001.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Li D, Zou XY, El-Ayachi I, Romero LO, Yu
Z, Iglesias-Linares A, Cordero-Morales JF and Huang GT: Human
dental pulp stem cells and gingival mesenchymal stem cells display
action potential capacity in vitro after neuronogenic
differentiation. Stem Cell Rev Rep. 15:67–81. 2019.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Lai PL, Lin H, Chen SF, Yang SC, Hung KH,
Chang CF, Chang HY, Lu FL, Lee YH, Liu YC, et al: Efficient
generation of chemically induced mesenchymal stem cells from human
dermal fibroblasts. Sci Rep. 7(44534)2017.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Cheng L, Hu W, Qiu B, Zhao J, Yu Y, Guan
W, Wang M, Yang W and Pei G: Generation of neural progenitor cells
by chemical cocktails and hypoxia. Cell Res. 24:665–679.
2014.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Cheng L, Gao L, Guan W, Mao J, Hu W, Qiu
B, Zhao J, Yu Y and Pei G: Direct conversion of astrocytes into
neuronal cells by drug cocktail. Cell Res. 25:1269–1272.
2015.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Fu Y, Huang C, Xu X, Gu H, Ye Y, Jiang C,
Qiu Z and Xie X: Direct reprogramming of mouse fibroblasts into
cardiomyocytes with chemical cocktails. Cell Res. 25:1013–1024.
2015.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Sayed N, Wong WT, Ospino F, Meng S, Lee J,
Jha A, Dexheimer P, Aronow BJ and Cooke JP: Transdifferentiation of
human fibroblasts to endothelial cells: Role of innate immunity.
Circulation. 131:300–309. 2015.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Xie M, Cao N and Ding S: Small molecules
for cell reprogramming and heart repair: Progress and perspective.
ACS Chem Biol. 9:34–44. 2014.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Huangfu D, Maehr R, Guo W, Eijkelenboom A,
Snitow M, Chen AE and Melton DA: Induction of pluripotent stem
cells by defined factors is greatly improved by small-molecule
compounds. Nat Biotechnol. 26:795–797. 2008.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Ladewig J, Mertens J, Kesavan J, Doerr J,
Poppe D, Glaue F, Herms S, Wernet P, Kögler G, Müller FJ, et al:
Small molecules enable highly efficient neuronal conversion of
human fibroblasts. Nat Methods. 9:575–578. 2012.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Gafni O, Weinberger L, Mansour AA, Manor
YS, Chomsky E, Ben-Yosef D, Kalma Y, Viukov S, Maza I, Zviran A, et
al: Derivation of novel human ground state naive pluripotent stem
cells. Nature. 504:282–286. 2013.PubMed/NCBI View Article : Google Scholar
|
|
33
|
PLOS ONE Staff. Correction: Neurotrophic
requirements of human motor neurons defined using amplified and
purified stem cell-derived cultures. PLoS One.
10(e0119195)2015.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Liu ML, Zang T, Zou Y, Chang JC, Gibson
JR, Huber KM and Zhang CL: Small molecules enable neurogenin 2 to
efficiently convert human fibroblasts into cholinergic neurons. Nat
Commun. 4(2183)2013.PubMed/NCBI View Article : Google Scholar
|
|
35
|
Zhu S, Ambasudhan R, Sun W, Kim HJ,
Talantova M, Wang X, Zhang M, Zhang Y, Laurent T, Parker J, et al:
Small molecules enable OCT4-mediated direct reprogramming into
expandable human neural stem cells. Cell Res. 24:126–129.
2014.PubMed/NCBI View Article : Google Scholar
|
|
36
|
Li Y, Huo S, Fang Y, Zou T, Gu X, Tao Q
and Xu H: ROCK inhibitor Y27632 induced morphological shift and
enhanced neurite outgrowth-promoting property of olfactory
ensheathing cells via YAP-dependent up-regulation of L1-CAM. Front
Cell Neurosci. 12(489)2018.PubMed/NCBI View Article : Google Scholar
|
|
37
|
Li Y, Zou T, Xue L, Yin ZQ, Huo S and Xu
H: TGF-β1 enhances phagocytic removal of neuron debris and neuronal
survival by olfactory ensheathing cells via integrin/MFG-E8
signaling pathway. Mol Cell Neurosci. 85:45–56. 2017.PubMed/NCBI View Article : Google Scholar
|
|
38
|
Bernal A and Arranz L: Nestin-expressing
progenitor cells: Function, identity and therapeutic implications.
Cell Mol Life Sci. 75:2177–2195. 2018.PubMed/NCBI View Article : Google Scholar
|
|
39
|
Sakayori N, Kikkawa T and Osumi N: Reduced
proliferation and excess astrogenesis of Pax6 heterozygous neural
stem/progenitor cells. Neurosci Res. 74:116–121. 2012.PubMed/NCBI View Article : Google Scholar
|
|
40
|
Han DW, Tapia N, Hermann A, Hemmer K,
Höing S, Araúzo-Bravo MJ, Zaehres H, Wu G, Frank S, Moritz S, et
al: Direct reprogramming of fibroblasts into neural stem cells by
defined factors. Cell Stem Cell. 10:465–472. 2012.PubMed/NCBI View Article : Google Scholar
|
|
41
|
Ichida JK, Blanchard J, Lam K, Son EY,
Chung JE, Egli D, Loh KM, Carter AC, Di Giorgio FP, Koszka K, et
al: A small-molecule inhibitor of TGF-beta signaling replaces sox2
in reprogramming by inducing nanog. Cell Stem Cell. 5:491–503.
2009.PubMed/NCBI View Article : Google Scholar
|
|
42
|
Li W, Zhou H, Abujarour R, Zhu S, Young
Joo J, Lin T, Hao E, Schöler HR, Hayek A and Ding S: Generation of
human-induced pluripotent stem cells in the absence of exogenous
Sox2. Stem Cells. 27:2992–3000. 2009.PubMed/NCBI View Article : Google Scholar
|
|
43
|
Maherali N and Hochedlinger K: Tgfbeta
signal inhibition cooperates in the induction of iPSCs and replaces
Sox2 and cMyc. Curr Biol. 19:1718–1723. 2009.PubMed/NCBI View Article : Google Scholar
|
|
44
|
Steinschneider R, Delmas P, Nedelec J,
Gola M, Bernard D and Boucraut J: Appearance of neurofilament
subunit epitopes correlates with electrophysiological maturation in
cortical embryonic neurons cocultured with mature astrocytes. Dev
Brain Res. 95:15–27. 1996.PubMed/NCBI View Article : Google Scholar
|
|
45
|
Weyer A and Schilling K: Developmental and
cell type-specific expression of the neuronal marker NeuN in the
murine cerebellum. J Neurosci Res. 73:400–409. 2003.PubMed/NCBI View Article : Google Scholar
|
|
46
|
Soltani MH, Pichardo R, Song Z, Sangha N,
Camacho F, Satyamoorthy K, Sangueza OP and Setaluri V:
Microtubule-associated protein 2, a marker of neuronal
differentiation, induces mitotic defects, inhibits growth of
melanoma cells, and predicts metastatic potential of cutaneous
melanoma. Am J Pathol. 166:1841–1850. 2005.PubMed/NCBI View Article : Google Scholar
|
|
47
|
Pfisterer U, Ek F, Lang S, Soneji S,
Olsson R and Parmar M: Small molecules increase direct neural
conversion of human fibroblasts. Sci Rep. 6(38290)2016.PubMed/NCBI View Article : Google Scholar
|
|
48
|
Gervois P, Struys T, Hilkens P, Bronckaers
A, Ratajczak J, Politis C, Brône B, Lambrichts I and Martens W:
Neurogenic maturation of human dental pulp stem cells following
neurosphere generation induces morphological and
electrophysiological characteristics of functional neurons. Stem
Cells Dev. 24:296–311. 2015.PubMed/NCBI View Article : Google Scholar
|
|
49
|
Zhang Q, Nguyen PD, Shi S, Burrell JC, Xu
Q, Cullen KD and Le AD: Neural crest stem-like cells
non-genetically induced from human gingiva-derived mesenchymal stem
cells promote facial nerve regeneration in rats. Mol Neurobiol.
55:6965–6983. 2018.PubMed/NCBI View Article : Google Scholar
|