Introduction
Obesity is a significant risk factor for many
metabolic diseases such as obstructive sleep apnea, cardiovascular
diseases, diabetes, cancers, and osteoarthritis (1,2).
Obesity can be considered a consequence of energy imbalance, which
causes an increase in differentiated adipocytes and excessive fat
accumulation (3,4). Adipogenesis, the process of maturation
of preadipocytes, is an integrated process involving the activation
of several signaling pathways (5).
The transcription factors such as the peroxisome
proliferation-activity receptor (PPAR) γ and CCAAT/enhancer-binding
protein (C/EBP) family are crucial for adipogenesis (6-8).
These major adipogenic-related factors control the expression of
the lipid metabolizing enzymes to form mature adipocytes. C/EBP
families are expressed at certain times in the process of
adipogenesis. C/EBP β and C/EBP δ promoters are activated in the
early stage of differentiation and act in the direct regulation of
adipogenesis (9). Activated C/EBP β
then regulate its neighboring promoter element, which subsequently
encodes the PPAR γ and C/EBP α genes during the later stage of
differentiation (10,11). The major transcription factors
upregulate adipocyte differentiation-related genes such as
adiponectin, leptin, and fatty acid binding protein (Fabp4) during
adipocyte differentiation (12,13).
Therefore, the control of adipogenesis involves suppressing this
transcriptional regulation. Many signaling pathways influence
adipocyte differentiation. The Akt pathway and the extracellular
signal-regulated kinase (ERK) 1/2 pathway are shown to be
responsible for adipogenesis (14,15).
The Akt is in a key position to promote insulin-like growth
factor-1 expression and adipocyte differentiation, while Akt
pathway inhibition is shown to reduce adipogenesis (16).
Many studies have focused on developing anti-obesity
drugs (17). Although several drugs
are currently available to treat obesity, their long-term use may
cause severe side effects (18).
Thus, multiple research studies are focused on new natural products
with potential for anti-obesity activity (19). The anti-obesity activity of various
plant extracts is reported to be mediated by the regulation of
adipogenesis (20). Garlic
(Allium sativum), one of the oldest medicinal plants, is
originally from Asia. Allium vegetables comprise one of the
natural sources of organosulfur compounds and show advantages in
therapeutic application, mostly owing to their cardiovascular
protective effects, lowering of cholesterol, and anti-cancer
properties (21). In addition,
Allium has possibly beneficial effects in the treatment of
obesity by adipogenesis inhibition (22), energy expenditure increase (23) and influence on expression of
inflammatory mediators from serum (24). The synthesis of alliin
(S-allyl-L-cysteine sulfoxide, SACSO), considered to be the major
component of garlic, was first reported by Stoll and Seebeck in
1951(25). Alliin, which has strong
antioxidant properties, has been used as a treatment remedy for
some diseases (26,27). Previous research found that alliin
can help in decreasing the serum levels of glucose and insulin
(28). Alliin also can regulate the
anti-inflammatory effects of preadipocytes by reducing cytokine
levels, such as IL-6 and TNF (29).
Alliin was shown to possess many biological effects. However, the
direct effect of alliin in adipogenesis has not yet been explored.
The aim of this study was to evaluate the effects of alliin on the
adipogenic differentiation of 3T3-L1 cells and its potential
association with the regulation of adipogenic transcription factors
PPAR γ, C/EBP, adiponectin and Fabp4, as well as possible
mechanism.
Materials and methods
Materials
Alliin, obtained from Abcam (ab141896), was
dissolved in dimethylsulfoxide (DMSO), and stored at -20˚C.
Cell culture
3T3-L1 fibroblasts were obtained from Japanese
Collection of Research Bioresources Cell Bank (Osaka, Japan). Cells
were cultured in DMEM (Sigma-Aldrich; Merck KGaA) supplemented with
10% fetal bovine serum (FBS; Gibco Life Technologies), 100 U/ml
penicillin, 100 µg/ml streptomycin, and 0.25 µg/ml amphotericin B
(Biological Industries) in a CO2 ncubator. The cells
were then trypsinized after reaching 80-90% confluence. Cells at
passages 2-15 were used in this study.
Adipocyte differentiation
The cells were differentiated following previously
reported method with some modifications (30). Briefly, adipocyte differentiation
begins after cell cultures reached 100% confluence. For adipogenic
induction, the medium used was DMEM containing 10% FBS, 1 µM
dexamethasone (Sigma-Aldrich; Merck KGaA), 0.5 mM
3-isobutyl-1-methylxanthine (Wako), and 10 µg/ml insulin (Funakoshi
Co. Ltd.) for 2 days. The cells were then transferred to DMEM
containing 10% FBS and 10 µg/ml insulin for 6 days. The medium was
changed every 2-3 days. Mature adipocytes were obtained on day 7 or
8. To evaluate the effects of alliin on adipogenesis, various
concentrations of alliin (10-40 µg/ml) were used during the
induction process. Cells that were not treated with alliin were
identified as the control.
Oil red O staining
To confirm the lipid accumulation in cultured cells,
Oil red O staining (Sigma-Aldrich; Merck KGaA) was performed. Eight
days after induction, mature adipocytes were fixed with 4%
paraformaldehyde phosphate buffer solution for 30 min at room
temperature and then stained with Oil red O working solution (6:4
of oil red stock solution: Distilled water) for 15 min. PBS was
washed three times to remove the excess Oil red O dye. The images
were observed under a parallel phase contrast microscope (Olympus
IX70 inverted microscope, Olympus Optical CO, Ltd.). For
quantitative analysis, the percentage of positively stained areas
were calculated using imageJ. Results are expressed as percentage
of Oil red O-stained area compared to control. For each sample, the
experiments were performed in triplicates.
Measurement of cell viability
Cell viability was determined using a Cell Counting
Kit-8 assay (CCK-8; DOJINDO Lab, Osaka, Japan). 3T3-L1 cells were
seeded in 96-well plates (5x103 cells/well) for 2 days.
Cells were then cultured with various concentrations of alliin
(0-40 µg/ml) for 7 days. Cell Counting Kit-8 (CCK-8; Dojindo
Laboratories) was used according the manufacturer's instruction.
Cells were incubated with 10 µl of CCK-8 reagent for 2 h at 37˚C.
The result was determined using a Synergy™ HTX Multi-Mode
Microplate Reader (BioTek Instruments) at 450 nm.
Reverse transcription-quantitative PCR
(RT-qPCR)
Total RNA was extracted from cells using
TRIzol® reagent (Ambion®; Gibco Life
Technologies) according to manufacturer's instructions. The
quantity and quality of isolated RNA were detected using a Nano
Drop spectrophotometer (Nano Drop® ND-1000, Thermo
Fisher Scientific, Inc.). Complementary DNA (cDNA) was synthesized
in a 20-µl reaction using 2 µg RNA, 20 pmol Oligo dT12-18
(Invitrogen; Thermo Fisher Scientific, Inc., 18418-012), 0.5 µl
RNase inhibitor (Promega Corporation, N211A), 0.2 µl ReverTra Ace
(Toyobo, TRT-101), 2 µl dNTP Mixture (Takara, 4030), 4 µl 5X RT
buffer (Toyobo, TRT-101), and 10 mM DEPC water. RT-qPCR was
performed in a 25-µl reaction, containing 1 µl cDNA, 10 pmol
forward and reverse primers, and 12.5 µl SYBR Premix Ex Taq II
(Takara, RR820A). PCR amplification was set as follows: 95˚C for 30
sec, 40 cycles of 95˚C for 5 sec, and 60˚C for 30 sec; then the
final dissociation was set at 95˚C for 15 sec, 60˚C for 30 sec, and
95˚C for 15 sec. The data were quantified using the 2-∆∆Cq method
and were normalized against the levels of β-actin (31). The primer sequences used for PCR are
provided in Table I.
 | Table IPrimer sequences for reverse
transcription-quantitative PCR. |
Table I
Primer sequences for reverse
transcription-quantitative PCR.
| Name of gene | Primers
(5'-3') | Melting temperature
(˚C) | Product length
(bp) | Genbank code |
|---|
| β-actin | F:
CATCCGTAAAGACCTCTATGCCAAC ATGGAGCCACCGATCCACA | 64.1 | 193 | NM_007393.5 |
| | R:
ATGGAGCCACCGATCCACA | 64.9 | | |
| PPAR γ | F:
GTGCCAGTTTCGATCCGTAGA | 66.2 | 167 | NM_001113418.1 |
| | R:
GGCCAGCATCGTGTAGATGA | 66.2 | | |
| C/EBP α | F:
GGACAAGAACAGCAACGAGTA | 61.8 | 237 | NM_001287514.1 |
| | R:
GCAGTTGCCATGGCCTTGA | 69.7 | | |
| C/EBP β | F:
TGGACAAGCTGAGCGACGAG | 69.1 | 192 | NM_001287738.1 |
| | R:
TGTGCTGCGTCTCCAGGTTG | 70.0 | | |
| Adiponectin | F:
GCACTGGCAAGTTCTACTGCAA | 66.4 | 156 | NM_009605.5 |
| | R:
GTAGGTGAAGAGAACGGCCTTGT | 66.4 | | |
| Leptin | F:
CCACACACAGCTGGAAACTC | 63.4 | 216 | NM_008493.3 |
| | R:
GCCTTGCTTCAGATCCATCC | 65.9 | | |
| Fabp4 | F:
CCAATGAGCAAGTGGCAAGA | 66.2 | 179 | NM_024406.3 |
| | R:
GATGCCAGGCTCCAGGATAG | 65.9 | | |
| PI3K | F:
TCCTGCTTCATACCGAGCTT | 63.8 | 212 | NM_001024955.2 |
| | R:
CATGACATCCTCCCTCTCGT | 64.2 | | |
| AKT | F:
CCCTTCTACAACCAGGACCA | 63.9 | 210 | NM_009652.3 |
| | R:
ATACACATCCTGCCACACGA | 64.1 | | |
| MAPK | F:
TGCCAGGCTGAACTACAGTG | 64.1 | 169 | NM_008927.4 |
| | R:
CACAAGGCTCCCTCTCAGAC | 64.1 | | |
| ERK | F:
TCAGAGGCAGGTGGATCTCT | 64.0 | 188 | NM_011949.3 |
| | R:
GGTGCCATCATCAACATCTG | 64.1 | | |
Statistical analysis
The results are presented as the mean ± standard
error of the mean (SEM) of three experiments. SPSS 13.0 and
GraphPad Prism 6 software were used for statistical analysis. The
statistical significance between each group was computed using
one-way analysis of variance followed by Dunnett's test. P<0.05
was considered statistically significant.
Results
Effect of alliin on intracellular
lipid accumulation during adipogenic differentiation
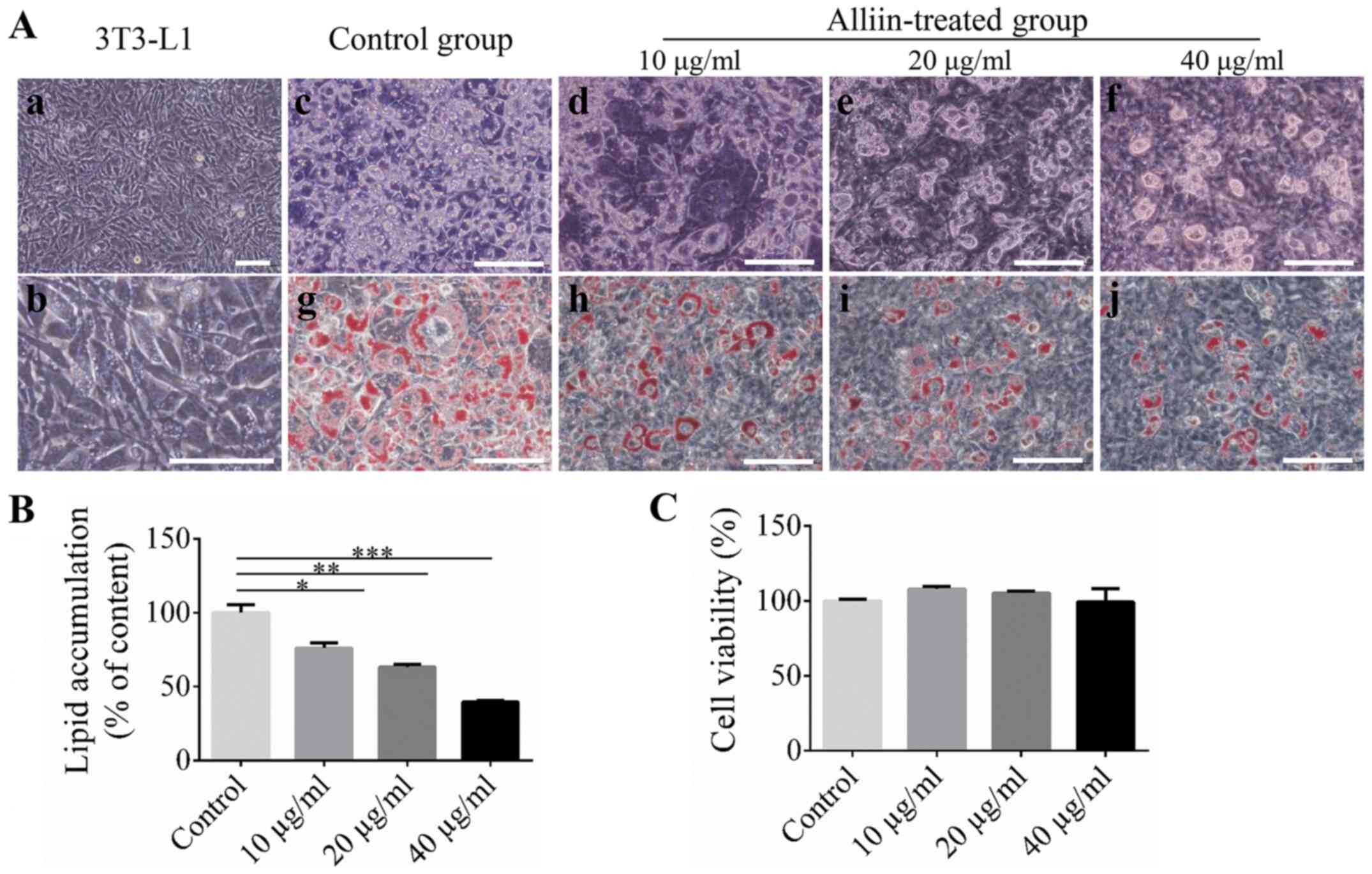
3T3-L1 cells showed fibroblastic-like morphology
before adipogenic induction (Fig.
1A-a and -b). Then, the
suppression effect of alliin on adipogenic differentiation was
examined. During adipogenic differentiation, the 3T3-L1 cells were
treated with different concentrations of alliin (10, 20, 40 µg/ml)
for 7 days, and the cells treated without alliin were used as
control. 3T3-L1 cells showed a morphological change from
fibroblastic-shape to round-shape, and then oil droplets were
formed and maintained after 7 days of induction (Fig. 1A-c-f). Oil red O staining was
performed to evaluate lipid accumulation levels. The Oil red
O-stained area decreased significantly in the alliin-treated groups
compared with the control group (Fig.
1A-g-j). Meanwhile, quantitative analysis showed that
alliin-treated group displayed dramatically lower lipid
accumulation in a dose-dependent manner compared to the control
group. 40 µg/ml alliin treatment showed the greatest effects on the
lipid accumulation which level dropped to 39.5% (Fig. 1B). The effect of alliin on cell
viability was examined. 3T3-L1 cells were treated with different
concentrations of alliin (10-40 µg/ml) for 7 days. Alliin has no
effect on cell viability even at a concentration of 40 µg/ml
(Fig. 1C). These data indicated
that alliin may contain anti-adipogenic potential.
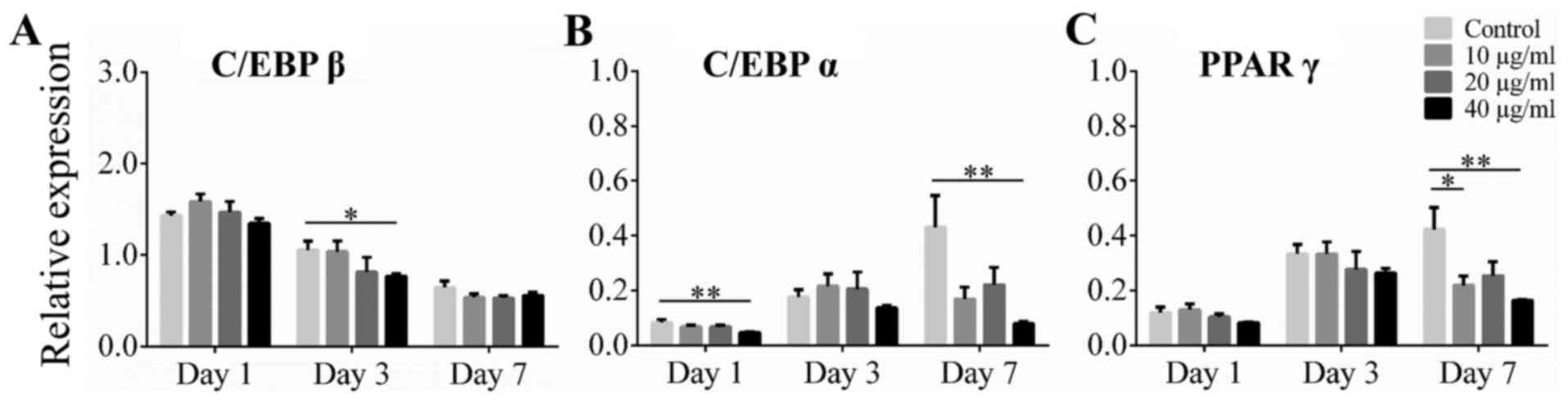
Effect of alliin on adipogenic
transcription markers C/EBP β, C/EBP α, and PPAR γ
To examine the effect of alliin on adipogenic
transcription markers during induction, cells were analyzed by
RT-qPCR. The expression levels of C/EBP β, C/EBP α and PPAR γ were
assessed on dayz 1, 3 and 7 of induction, respectively. The mRNA
level of C/EBP β, induced by the differentiation medium, reached
its peak on day 1, and then decreased gradually over time. Cells
treated with 40 µg/ml alliin on day 3 showed a significant decrease
in lipid accumulation compared to cells in the control group, but
there was no significant difference on day 7 (Fig. 2A). The expression of C/EBP α and
PPAR γ were gradually induce during adipogenic differentiation in
control group. Expression level of C/EBP α was significantly
reduced by treatment with 40 µg/ml alliin compared to without
treatment on day 1 and day 7 (Fig.
2B) Treatment with 40 µg/ml alliin also significantly inhibited
the expression of PPAR γ on day 7 (Fig.
2C). These results indicate that 40 µg/ml alliin suppress
adipogenesis by downregulating the expression of C/EBP α and PPAR
γ.
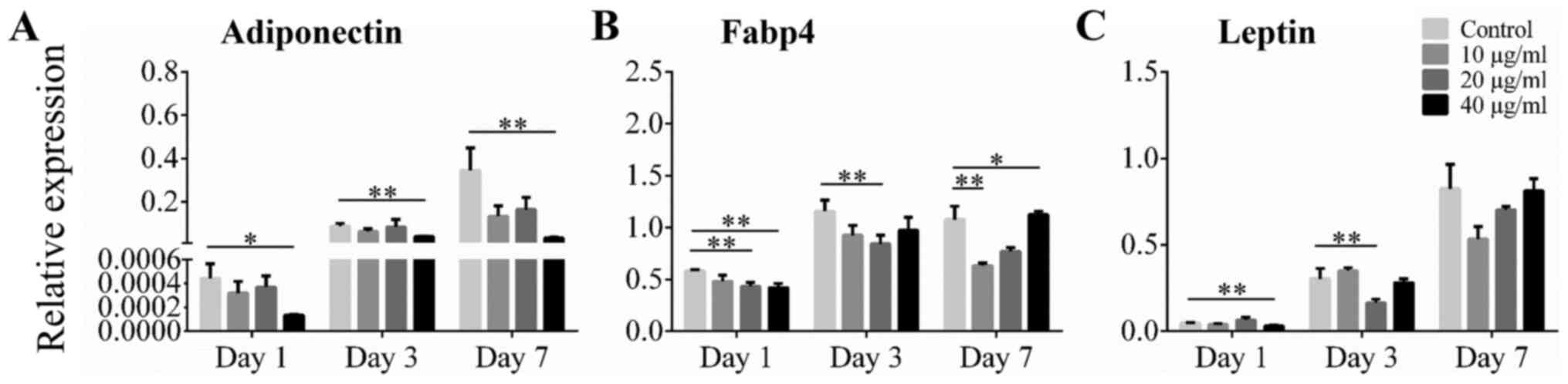
Effect of alliin on adipocyte-related
genes adiponectin, Fabp4, and leptin
Expression of adipocyte-related genes was also
analyzed using RT-qPCR. During adipogenic differentiation,
expression of adiponectin mRNA increased over time. The level of
adiponectin mRNA was significantly lower with treatment of 40 µg/ml
alliin than without treatment at all time points examined (Fig. 3A). The level of Fabp4 mRNA was
significantly decreased in cells cultured with 40 µg/ml alliin than
in those without treatment on day 1 and day 7 (Fig. 3B). Whereas, the level of leptin mRNA
was significantly lower in the 40 µg/ml alliin treatment group than
in the control group on day 1 (Fig.
3C). These results indicate that 40 µg/ml alliin inhibits
adipogenesis and leads to reduce expression of adiponectin and
Fabp4.
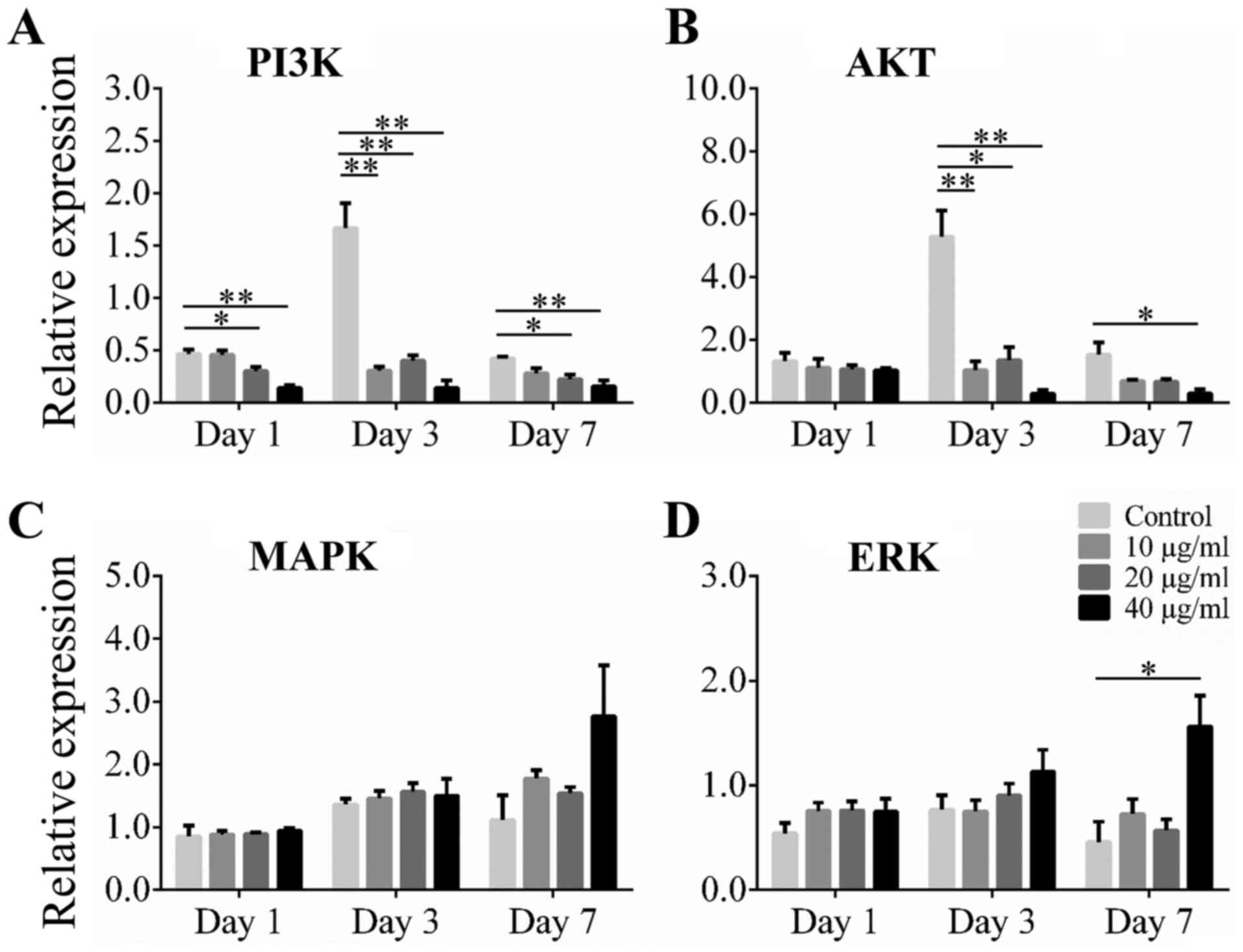
Effect of Alliin on the Akt and ERK
1/2 pathways related genes during adipogenic differentiation
Akt and ERK 1/2 are upstream regulators in the
adipocyte differentiation pathway, including C/EBP α and PPAR γ
pathways. To investigate the possible mechanism of alliin on
adipogenic differentiation, the Akt and ERK 1/2 pathways related
genes were examined. The 3T3-L1 cells were cultured with various
concentrations of alliin during the differentiation of adipocyte.
Results showed that expression of PI3K and Akt was significantly
increased at the early stage of differentiation. Expression of PI3K
was significantly inhibited in the 20 or 40 µg/ml alliin treatment
group compared to that in the control group at all time points
examined (Fig. 4A). The level of
AKT mRNA was significantly lower with treatment of 40 µg/ml alliin
than without treatment on days 3 and 7 (Fig. 4B). However, increased expression of
MAPK and ERK was observed in the treatment groups compared to that
in the control groups on day 7 (Fig.
4C). The level of ERK mRNA increased significantly with 40
µg/ml alliin treatment compared to that without treatment after 7
days (Fig. 4D). These results
suggest that 40 µg/ml alliin inhibits adipocyte differentiation by
reducing the expression of PI3K and Akt.
Discussion
In the current study, we indicated that alliin
treatment may contribute to decreased adipogenesis of 3T3-L1
adipocyte. This study revealed, for the first time, the direct
effect of alliin on adipogenesis. Moreover, we found an effect of
alliin on adipogenesis may be achieved by downregulating the Akt
and PI3K expression. Several recent reports indicate the benefit of
plant extracts as drugs (19,32).
Compared to conventional drugs, herbal medicines show less
potentially dangerous side effects. Many plant compounds, such as
ginsenoside, caffeic acid, berberine, anthocyanin, and capsaicin,
have been shown to inhibit adipogenesis (33-35).
Obesity is commonly caused by an excessive increase
of adipocytes. Adipogenesis is a complex regulated cellular
differentiation process involving many signaling pathways and
related molecules (36,37). The inhibition of adipogenesis
process can provide a target for the control and treatment of
obesity. During 3T3-L1 adipocyte differentiation, C/EBP β is
rapidly induced and is responsible for activating the adipogenic
regulators C/EBP α and PPAR γ (38). PPAR γ plays a crucial role in
adipogenesis of embryonic stem cells and fibroblasts. PPAR γ levels
are significantly induced when preadipocytes are converted into
adipocytes (39,40). The research suggested that white
adipose tissue under obesity condition a significant increment in
oxidative stress, pro-inflammatory status and depletion of n-3 long
chain polyunsaturated fatty acid (n-3 LCPUFA) (41). Whereas PPAR γ transcription factor
is regulated by n-3 LCPUFA that participate in the metabolism of
adipose tissue (42). So, the
regulation of adipogenesis occurs by controlling PPAR γ levels
(20). Moreover, PPAR γ has been
shown to induce and activate the transcriptional factor C/EBP α
promoter (43). C/EBP α is a
transcriptional factor of the C/EBP family and also plays an
important role in regulating adipogenesis (44). Thus, PPAR γ and C/EBP α genes are
upregulated and together activate the adipogenesis pathways
(45). In this study, a direct
effect of alliin on adipocyte differentiation was performed. Alliin
significantly decreased the lipid droplet accumulation. Expression
level of C/EBP β was significantly decreased in presence of 40
µg/ml alliin on day 3. Expression of C/EBP α and PPAR γ was lower
with 40 µg/ml alliin treatment on day 7, while the level of C/EBP β
was not affected. These result may correspond to C/EBP β induced in
the early stage of differentiation that subsequently activates the
adipogenesis markers, PPAR γ and C/EBP α, at the later stage of
differentiation. The results suggest that alliin may inhibit
adipogenesis by reducing C/EBP β, C/EBP α, and PPAR γ levels during
adipocyte differentiation.
In addition, PPAR γ and C/EBP α can be part of a
feedback loop and promote adipogenesis by leading the expression of
downstream genes, such as adiponectin, Fabp4, and leptin. These
downstream genes play major roles in inducing and maintaining
mature adipocytes (5). A recent
study indicated that the expression of adipocyte-related genes
during induction is PPAR γ dependent (46). In the alliin treatment group,
expression of leptin was lower during the early stage of
differentiation, while the level was not affected at day 7. This
may be due to the existence of different regulatory mechanisms of
leptin expression. Recent study noted that leptin can directly
induce the adipocyte differentiation in the early stage of
adipogenesis, while showing anti-adiposity effects after maturation
of adipocytes (47). Expression of
adiponection and Fabp4 was significantly inhibited with alliin
treatment after adipocyte induction. These results clearly imply
that alliin inhibits adipogenic differentiation by reducing the
expression of adipocyte-related genes, which may be through the
repression of PPAR γ and C/EBP α related pathways.
Insulin signaling pathway plays a critical role in
modulating adipogenesis. In the presence of insulin, the insulin
receptor is autophosphorylated, and subsequently proteins in the
insulin receptor substrate family are also phosphorylated, thereby
activating the two main signaling pathways, PI3K/Akt and MAPK/ERK
pathways (48). The function of
PI3K/Akt signaling is to activate adipogenic transcription marker
such as C/EBP α and promote adipocyte maturation during
adipogenesis (49). A study also
showed that expression of PPAR γ is significantly impeded in
Akt-deficient mice (50). Thus,
inhibition of PI3K/Akt signaling pathway may provide a therapeutic
target for obesity (51). The
MAPK/ERK signaling pathway plays a complicated role in the
regulation of adipogenesis. Activation of the MAPK/ERK pathway can
both inhibit and promote adipocyte differentiation. Tanabe et
al (52) reported that the
MAPK/ERK pathway works in a suppressive manner on adipocyte
differentiation when receiving a long-term or sustained
stimulation. In this study, the higher expression levels of ERK
with alliin treatment may correspond to the inhibition effects of
MAPK pathway on adipogenesis. In contrast, the expression level of
P13K and Akt mRNA expression were markedly decreased with alliin
treatment. Collectively, our study indicated that alliin resulted
in PI3K/Akt inhibition, thereby suppressing the expression of C/EBP
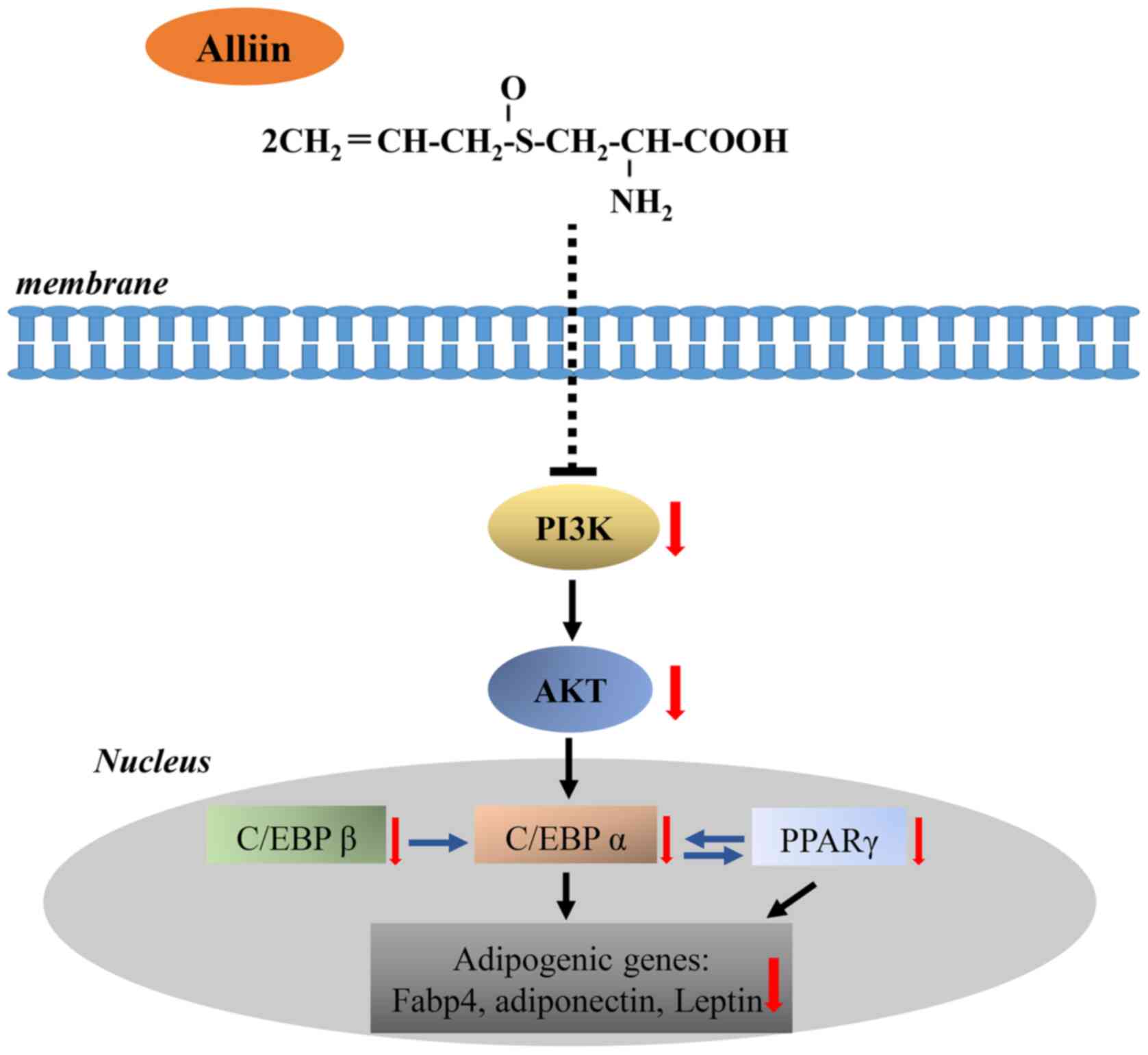
β, C/EBP α, and PPAR γ and adipocyte-related genes (Fig. 5). Although the results from this
study indicated that the Akt signaling pathway play an important
role in alliin inhibit adipocyte differentiation, there are still
limitations. First, the phosphorylation levels of Akt signaling
related proteins should be test. Second, whether the validity of
this theory still needs further experimental investigation.
In conclusion, the results demonstrated that alliin
in garlic can inhibit adipogenesis by reducing expression of major
transcriptional activators and their downstream genes, which may be
mediated by regulation of Akt signaling pathway. Alliin may provide
a possible naturally occurring therapeutic method for the
prevention and treatment of metabolic disease.
Acknowledgements
Not applicable.
Funding
Funding: The present study was supported in part by
Grants-in-Aid for Scientific Research (C) from Japan Society for
the Promotion of Science (JSPS; KAKENHI grant no. JP19K10192).
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
NL and KC performed all experiments, acquired data,
analyzed the data and drafted the manuscript. . KC and HD
contributed to the acquisition of data. JY, MY, HK and XL designed
the experiment. XL revised the manuscript. All authors read and
approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Wilson PW, Bozeman SR, Burton TM, Hoaglin
DC, Ben-Joseph R and Pashos CL: Prediction of first events of
coronary heart disease and stroke with consideration of adiposity.
Circulation. 118:124–130. 2008.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Jehan S, Myers AK, Zizi F, Pandi-Perumal
SR, Jean-Louis G and McFarlane SI: Obesity, obstructive sleep apnea
and type 2 diabetes mellitus: Epidemiology and pathophysiologic
insights. Sleep Med Disord. 2:52–58. 2018.PubMed/NCBI
|
|
3
|
Faria SL, Faria OP, Menezes CS, de Gouvêa
HR and de Almeida Cardeal M: Metabolic profile of clinically severe
obese patients. Obes Surg. 22:1257–1262. 2012.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Jéquier E and Tappy L: Regulation of body
weight in humans. Physiol Rev. 79:451–480. 1999.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Moseti D, Regassa A and Kim WK: Molecular
regulation of adipogenesis and potential anti-adipogenic bioactive
molecules. Int J Mol Sci. 17(124)2016.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Rosen ED, Hsu CH, Wang X, Sakai S, Freeman
MW, Gonzalez FJ and Spiegelman BM: C/EBPalpha induces adipogenesis
through PPARgamma: A unified pathway. Genes Dev. 16:22–26.
2002.PubMed/NCBI View Article : Google Scholar
|
|
7
|
El-Jack AK, Hamm JK, Pilch PF and Farmer
SR: Reconstitution of insulin-sensitive glucose transport in
fibroblasts requires expression of both PPARgamma and C/EBPalpha. J
Biol Chem. 274:7946–7951. 1999.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Lefterova MI and Lazar MA: New
developments in adipogenesis. Trends Endocrinol Metab. 20:107–114.
2009.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Tanaka T, Yoshida N, Kishimoto T and Akira
S: Defective adipocyte differentiation in mice lacking the
C/EBPbeta and/or C/EBPdelta gene. EMBO J. 16:7432–7443.
1997.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Clarke SL, Robinson CE and Gimble JM:
CAAT/enhancer binding proteins directly modulate transcription from
the peroxisome proliferator-activated receptor gamma2 promoter.
Biochem Biophys Res Commun. 240:99–103. 1997.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Farmer SR: Transcriptional control of
adipocyte formation. Cell Metab. 4:263–273. 2006.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Wei S, Zan LS, Wang HB, Cheng G, Du M,
Jiang Z, Hausman GJ, Mcfarland DC and Dondson MV:
Adenovirus-mediated interference of FABP4 regulates mRNA expression
of ADIPOQ, LEP and LEPR in bovine adipocytes. Genet Mol Res.
12:494–505. 2019.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Rosen ED, Walkey CJ, Puigserver P and
Spiegelman BM: Transcriptional regulation of adipogenesis. Genes
Dev. 14:1293–1307. 2000.PubMed/NCBI
|
|
14
|
Tseng YH, Butte AJ, Kokkotou E, Yechoor
VK, Taniguchi CM, Kriauciunas KM, Cypess AM, Niinobe M, Yoshikawa
K, Patti ME and Kahn CR: Prediction of preadipocyte differentiation
by gene expression reveals role of insulin receptor substrates and
necdin. Nat Cell Biol. 7:601–611. 2005.PubMed/NCBI View
Article : Google Scholar
|
|
15
|
Prusty D, Park BH, Davis KE and Farmer SR:
Activation of MEK/ERK signaling promotes adipogenesis by enhancing
peroxisome proliferator-activated receptor gamma (PPARgamma) and
C/EBPalpha gene expression during the differentiation of 3T3-L1
preadipocytes. J Biol Chem. 277:46226–46232. 2002.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Guo LX, Chen G, Yin ZY, Zhang YH and Zheng
XX: P-Synephrine exhibits anti-adipogenic activity by activating
the Akt/GSK3β signaling pathway in 3T3-L1 adipocytes. J Food
Biochem. 43(e13033)2019.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Powell AG, Apovian CM and Aronne LJ: New
drug targets for the treatment of obesity. Clin Pharmacol Ther.
90:40–51. 2011.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Yanovski SZ and Yanovski JA: Long-term
drug treatment for obesity: A systematic and clinical review. JAMA.
311:74–86. 2014.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Kumar P and Bhandari U: Common medicinal
plants with antiobesity potential: A special emphasis on fenugreek.
Anc Sci Life. 35:58–63. 2015.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Hatano T, Sameshima Y, Kawabata M, Yamada
S, Shinozuka K, Nakabayashi T and Mizuno H: St. John's wort
promotes adipocyte differentiation and modulates NF-κB activation
in 3T3-L1 cells. Biol Pharm Bull. 37:1132–1138. 2014.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Mikaili P, Maadirad S, Moloudizargari M,
Aghajanshakeri S and Sarahroodi S: Therapeutic uses and
pharmacological properties of garlic, shallot, and their
biologically active compounds. Iran J Basic Med Sci. 16:1031–1048.
2013.PubMed/NCBI
|
|
22
|
Kim HJ, Lee MJ, Jang JY and Lee SH:
Allium hookeri root extract inhibits adipogenesis by
promoting lipolysis in high fat diet-induced obese mice. Nutrients.
11(2262)2019.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Kagawa Y, Ozaki-Masuzawa Y, Hosono T and
Seki T: Garlic oil suppresses high-fat diet induced obesity in rats
through the upregulation of UCP-1 and the enhancement of energy
expenditure. Exp Ther Med. 19:1536–1540. 2020.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Xu C, Mathews AE, Rodrigues C, Eudy BJ,
Rowe CA, O'Donoughue A and Percival SS: Aged garlic extract
supplementation modifies inflammation and immunity of adults with
obesity: A randomized, double-blind, placebo-controlled clinical
trial. Clin Nutr ESPEN. 24:148–155. 2018.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Stoll A and Seeback E: Chemical
investigations on alliin, the specific principle of garlic. Adv
Enzymol Relat Subj Biochem. 11:377–400. 1951.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Slusarenko AJ, Patel A and Portz D:
Control of plant diseases by natural products: Allicin from garlic
as a case study. Eur J Plant Pathol. 121:313–322. 2008.
|
|
27
|
Borlinghaus J, Albrecht F, Gruhlke MC,
Nwachukwu ID and Slusarenko AJ: Allicin: Chemistry and biological
properties. Molecules. 19:12591–12618. 2014.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Upadhyay RK: Garlic: A potential source of
pharmaceuticals and pesticides: A review. Int J Green Phar.
10:1–28. 2016.
|
|
29
|
Quintero-Fabián S, Ortuño-Sahagún D,
Vázquez-Carrera M and López-Roa RI: Alliin, a Garlic (Allium
sativum) compound, prevents LPS-induced inflammation in 3T3-L1
adipocytes. Mediators Inflamm. 2013(381815)2013.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Zhang M, Sheng S, Zhang W, Zhang J, Zhang
Z, Zhang M, Hatch GM and Chen L: MiR27a promotes the development of
macrophage-like characteristics in 3T3-L1 preadipocytes. Int J Biol
Sci. 14:1599–1609. 2018.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408.
2001.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Ekor M: The growing use of herbal
medicines: Issues relating to adverse reactions and challenges in
monitoring safety. Front Pharmacol. 4(177)2014.PubMed/NCBI View Article : Google Scholar
|
|
33
|
Aoyagi R, Funakoshi-Tago M, Fujiwara Y and
Tamura H: Coffee inhibits adipocyte differentiation via
inactivation of PPARγ. Biol Pharm Bull. 37:1820–1825.
2014.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Liao CC, Ou TT, Huang HP and Wang CJ: The
inhibition of oleic acid induced hepatic lipogenesis and the
promotion of lipolysis by caffeic acid via up-regulation of
AMP-activated kinase. J Sci Food Agric. 94:1154–1162.
2014.PubMed/NCBI View Article : Google Scholar
|
|
35
|
Wu T, Jiang Z, Yin J, Long H and Zheng X:
Anti-obesity effects of artificial planting blueberry (Vaccinium
ashei) anthocyanin in high-fat diet-treated mice. Int J Food Sci
Nutr. 67:257–264. 2016.PubMed/NCBI View Article : Google Scholar
|
|
36
|
Prins JB and O'Rahilly S: Regulation of
adipose cell number in man. Clin Sci (Lond). 92:3–11.
1997.PubMed/NCBI View Article : Google Scholar
|
|
37
|
Guo L, Li X and Tang QQ: Transcriptional
regulation of adipocyte differentiation: A central role for
CCAAT/enhancer-binding protein (C/EBP) β. J Biol Chem. 290:755–761.
2015.PubMed/NCBI View Article : Google Scholar
|
|
38
|
Salma N, Xiao H and Imbalzano AN: Temporal
recruitment of CCAAT/enhancer-binding proteins to early and late
adipogenic promoters in vivo. J Mol Endocrinol. 36:139–151.
2006.PubMed/NCBI View Article : Google Scholar
|
|
39
|
Tontonoz P, Hu E and Spiegelman BM:
Stimulation of adipogenesis in fibroblasts by PPARgamma2, a
lipid-activated transcription factor. Cell. 79:1147–1156.
1994.PubMed/NCBI View Article : Google Scholar
|
|
40
|
Wang F, Mullican SE, DiSpirito JR, Peed LC
and Lazar MA: Lipoatrophy and severe metabolic disturbance in mice
with fat-specific deletion of PPARγ. Proc Natl Acad Sci USA.
110:18656–18661. 2013.PubMed/NCBI View Article : Google Scholar
|
|
41
|
Illesca P, Valenzuela R, Espinosa A,
Echeverría F, Soto-Alarcon S, Ortiz M and Videla LA: Hydroxytyrosol
supplementation ameliorates the metabolic disturbances in white
adipose tissue from mice fed a high-fat diet through recovery of
transcription factors Nrf2, SREBP-1c, PPAR-γ and NF-κB. Biomed
Pharmacother. 109:2472–2481. 2019.PubMed/NCBI View Article : Google Scholar
|
|
42
|
Echeverría F, Ortiz M, Valenzuela R and
Videla LA: Long-chain polyunsaturated fatty acids regulation of
PPARs, signaling: Relationship to tissue development and aging.
Prostaglandins Leukot Essent Fatty Acids. 114:28–34.
2016.PubMed/NCBI View Article : Google Scholar
|
|
43
|
Wu Z, Rosen ED, Brun R, Hauser S, Adelmant
G, Troy AE, McKeon C, Darlington GJ and Spiegelman BM:
Cross-regulation of C/EBPα and PPARγ controls the transcriptional
pathway of adipogenesis and insulin sensitivity. Mol Cell.
3:151–158. 1999.PubMed/NCBI View Article : Google Scholar
|
|
44
|
Tang QQ, Otto TC and Lane MD:
CCAAT/enhancer-binding protein beta is required for mitotic clonal
expansion during adipogenesis. Proc Natl Acad Sci USA. 100:850–855.
2003.PubMed/NCBI View Article : Google Scholar
|
|
45
|
Ghaben AL and Scherer PE: Adipogenesis and
metabolic health. Nat Rev Mol Cell Biol. 20:242–258.
2019.PubMed/NCBI View Article : Google Scholar
|
|
46
|
Gerhold DL, Liu F, Jiang G, Li Z, Xu J, Lu
M, Sachs JR, Bagchi A, Fridman A, Holder DJ, et al: Gene expression
profile of adipocyte differentiation and its regulation by
peroxisome proliferator-activated receptor-gamma agonists.
Endocrinology. 143:2106–2118. 2002.PubMed/NCBI View Article : Google Scholar
|
|
47
|
Lukaszewski MA, Eberlé D, Vieau D and
Breton C: Nutritional manipulations in the perinatal period program
adipose tissue in offspring. Am J Physiol Endocrinol Metab.
305:E1195–E1207. 2013.PubMed/NCBI View Article : Google Scholar
|
|
48
|
Son YH, Ka S, Kim AY and Kim JB:
Regulation of adipocyte differentiation via microRNAs. Endocrinol
Metab (Seoul). 29:122–135. 2014.PubMed/NCBI View Article : Google Scholar
|
|
49
|
Saltiel AR and Kahn CR: Insulin signalling
and the regulation of glucose and lipid metabolism. Nature.
414:799–806. 2001.PubMed/NCBI View Article : Google Scholar
|
|
50
|
Peng XD, Xu PZ, Chen ML, Hahn-Windgassen
A, Skeen J, Jacobs J, Sundararajan D, Chen WS, Crawford SE, Coleman
KG and Hay N: Dwarfism, impaired skin development, skeletal muscle
atrophy, delayed bone development, and impeded adipogenesis in mice
lacking Akt1 and Akt2. Genes Dev. 17:1352–1365. 2003.PubMed/NCBI View Article : Google Scholar
|
|
51
|
Tang QQ, Gronborg M, Huang H, Kim JW, Otto
TC, Pandey A and Lane MD: Sequential phosphorylation of CCAAT
enhancer-binding protein beta by MAPK and glycogen synthase kinase
3beta is required for adipogenesis. Proc Natl Acad Sci USA.
102:9766–9771. 2005.PubMed/NCBI View Article : Google Scholar
|
|
52
|
Tanabe Y, Koga M, Saito M, Matsunaga Y and
Nakayama K: Inhibition of adipocyte differentiation by mechanical
stretching through ERK-mediated downregulation of PPARgamma2. J
Cell Sci. 117:3605–3614. 2004.PubMed/NCBI View Article : Google Scholar
|