Introduction
Tuberculosis (TB) induced by Mycobacterium
tuberculosis, is one of the deadliest human infections
worldwide (1). In 2021, there were
5.8 million new TB cases globally, and nearly 1.5 million patients
died from TB-associated disease (2). Over the past 100 years, great
achievements have been made in TB treatment and control through the
Bacillus Calmette-Guerin vaccination and the application of anti-TB
drugs (3). However, there is still
a lack of understanding of the host-pathogen interactions that
occur to better control TB (4). In
view of the global TB infection rate of 1/3, early TB recognition
and treatment are urgently required (5). At present, clinical studies have
uncovered that early TB diagnosis is still quite difficult, and the
rates of morbidity and mortality are increased due to misdiagnosis
(6-8).
Cofilin (CFL), an actin decomposing factor, is
expressed in various cells and modulates the generation of actin
filaments by controlling polymer formation and degradation
(9). As the non-muscle isoform of
its gene product, CFL1 is required by the importin-9-regulated
nuclear migration of G-actin (10)
and is involved in transcription extension modulated by RNA
polymerase II (11). CFL1 was
previously recognized as a critical protein in the routine nuclear
structures and roles of remote cell types (12). CFL1 has an essential role in cell
movement and cytoplasmic changes and is directly associated with
the invasion, spread and drug resistance of various human
malignancies such as mammary tumors (13-16).
Of note, our previous proteomics studies on exosomes found that
CFL1 was significantly increased in the supernatant of
Mycobacterium avium infected macrophages (17) and was also upregulated in M.
avium-infected macrophages (18).
The present study aimed to explore the expression of
cofilin-1 in M. avium-infected macrophages and the blood of
patients with TB and investigate the possibility of CFL1 as a
marker for use in TB diagnosis.
Materials and methods
Patient sample preparation
Between May 2019 and July 2020, a total of 36
patients with active TB (16 female; age, 25-65 years; mean age,
46.6 years; 20 males, age, 24-63 years, mean age, 43.6 years) and
34 healthy subjects (17 female; age, 22-56 years; mean age, 40.2
years; 17 males, age, 20-55 years, mean age, 38.5 years) were
recruited for the present study at The First People's Hospital of
Kunshan Affiliated to Jiangsu University (Kunshan, China).
EDTA-treated peripheral blood (5 ml) and plasma were collected from
all subjects. Approval was obtained from the Ethics and Scientific
Committee of the Affiliated Kunshan Hospital of Jiangsu University,
and all participants signed an informed consent form (approval no.
IEC-C-007-A07-V3.0). The experiments were performed in accordance
with the Declaration of Helsinki Principles. TB patients were
diagnosed according to the clinical criteria from the WHO (19), and normal individuals included
those with the results of negative tuberculin skin tests.
Peripheral blood (5 ml) treated with EDTA was mixed
with PBS at a 1:1 ratio and added into 50 ml centrifuge tubes, to
which 20 ml Ficoll separation solution was added, and
centrifugation was performed at 1,200 x g for 30 min at room
temperature. Finally, peripheral blood mononuclear cells (PBMCs)
were transferred into new tubes and stored at -80˚C for the
subsequent experiments.
Cell-free plasma was isolated from all blood samples
using heparin anticoagulant tubes, and centrifuged at 4,000 x g for
30 min at room temperature. The plasma samples were then separated
into microcentrifuge tubes and stored at -80˚C.
Cell culture
THP-1 cells (The Cell Bank of the Type Culture
Collection of The Chinese Academy of Sciences) were cultured in
Dulbecco's modified Eagle's medium (DMEM) (Hyclone; GE Healthcare,
USA) with 10% fetal bovine serum (FBS) (Hyclone; GE Healthcare) and
1% penicillin-streptomycin, and cultured at 37˚C in a humid
condition with 5% CO2. THP-1 cells grown in 6-well
plates at 75% confluence were induced into macrophages with 0.1 nM
phorbol ester. Macrophages were infected with M. avium (Mol
1:10, 1:50, 1:100, 1:200) for 24 h.
Cell transfection
Small interfering RNA (siRNA) of CFL1 (50 nm)
(Shanghai GenePharma Co., Ltd.) were transfected into macrophages
grown in 6-well plates at 75% confluence using
Lipofectamine® 3000 reagent (Invitrogen; Thermo Fisher
Scientific, Inc.) at 37˚C for 24 h according to the manufacturer's
instructions. The sequences used were as follows: CFL1 siRNA sense,
5'-GAAGGUGCGUAAGUCUUCATT-3' and antisense,
5'-UGAAGACUUACGCACCUUCTT-3'; and scrambled siRNA sense,
5'-AUAUUCCUGCGAUAGCUCGTT-3' and antisense, 5'-CGAGCUAUCGCAGGAAUA
UTT-3'.
Reverse transcription-quantitative PCR
(RT-qPCR)
Total RNA was extracted using TRIzol®
reagent (Invitrogen; Thermo Fisher Scientific, Inc.) according to
the manufacturer's instructions. cDNA was synthesized from isolated
RNA with a PrimeScript™ RT reagent kit (Takara
Biotechnology Co., Ltd.). qPCR was performed with the Super SYBR
Green kit (Thermo Fisher Scientific, Inc.) on ABI7500 (Thermo
Fisher Scientific, Inc.) instrument. The qPCR primers were as
follows: CFL1-forward (F), 5'-TGCCCTCTCCTTTTCGTTTCC-3' and -reverse
(R), 5'-CTCCTCTGGCGTTGAAGACT-3'; and GAPDH-F,
5'-AGAAGGCTGGGGCTCATTTG-3' and -R, 5'-AGGGGCCATCCACAGTCTTC-3'. The
thermocycling conditions were: 95˚C for 10 min, followed by 36
cycles of 95˚C for 25 sec and 60˚C for 30 sec. The mRNA fold-change
was calculated by using the 2-∆∆Cq method (20).
Cytokine analysis
Macrophages cultured in 6-well plates were treated
with M. avium or transfected with CFL1-siRNA for 24 h. The
expression of CFL1 in the plasma and supernatant of transfected
cells of patients with TB was detected by ELISA (cat. no.
CSB-EL005280HU, Cusabio Technology LLC). The cytokine content was
determined by the standard curve method.
Western blotting
The PBMCs and transfected cells were lysed with RIPA
lysate (cat. no: P0013C, Beyotime Biotechnology Corp.) for BCA
protein quantification. The proteins (20 µg) were separated via
SDS-PAGE on 10% gel and subsequently transferred onto a PVDF
membrane (Bio-Rad Laboratories, Inc.). The membranes were first
incubated with primary anti-CFL1 antibodies (cat. no. CSB-PA001739;
1:1,200, Cusabio Technology LLC) at 37˚C for 60 min and then with
HRP-conjugated Affinipure goat anti-rabbit IgG (H+L) secondary
antibody at 37˚C for 120 min. After washing, immunoreactivity was
visualized using Chemiluminescent Substrate System (Thermo Fisher
Scientific, Inc.) according to the instructions of the
manufacturer. Band intensity was evaluated using ImageJ 1.45
software (National Institutes of Health).
Receiver operating characteristic
(ROC) curves
The diagnostic ability of CFL1 in patients infected
with TB was analyzed by ROC curves. The area under the ROC curve
(AUC) detects the capability of an index to differentiate two
groups. AUCs of 90-100%, 80-90%, 70-80%, 60-70%, 50-60% and <50%
generally imply excellent, good, fair, poor, bad (or failed) and
incapable (or random data), respectively. Therefore, the ROC curve
was used to evaluate the predictive power of CFL1 in discriminating
between healthy individuals and patients with TB.
Statistical analysis
All statistical tests were performed using GraphPad
Prism 5 (GraphPad Software, Inc.). One-way ANOVA followed by
Tukey's post hoc test were performed on this statistical software
and the statistical test used is mentioned in the figure legends.
All experimental results are expressed as the mean ± SEM from three
individual experiments. P<0.05 was considered to indicate a
statistically significant difference.
Results
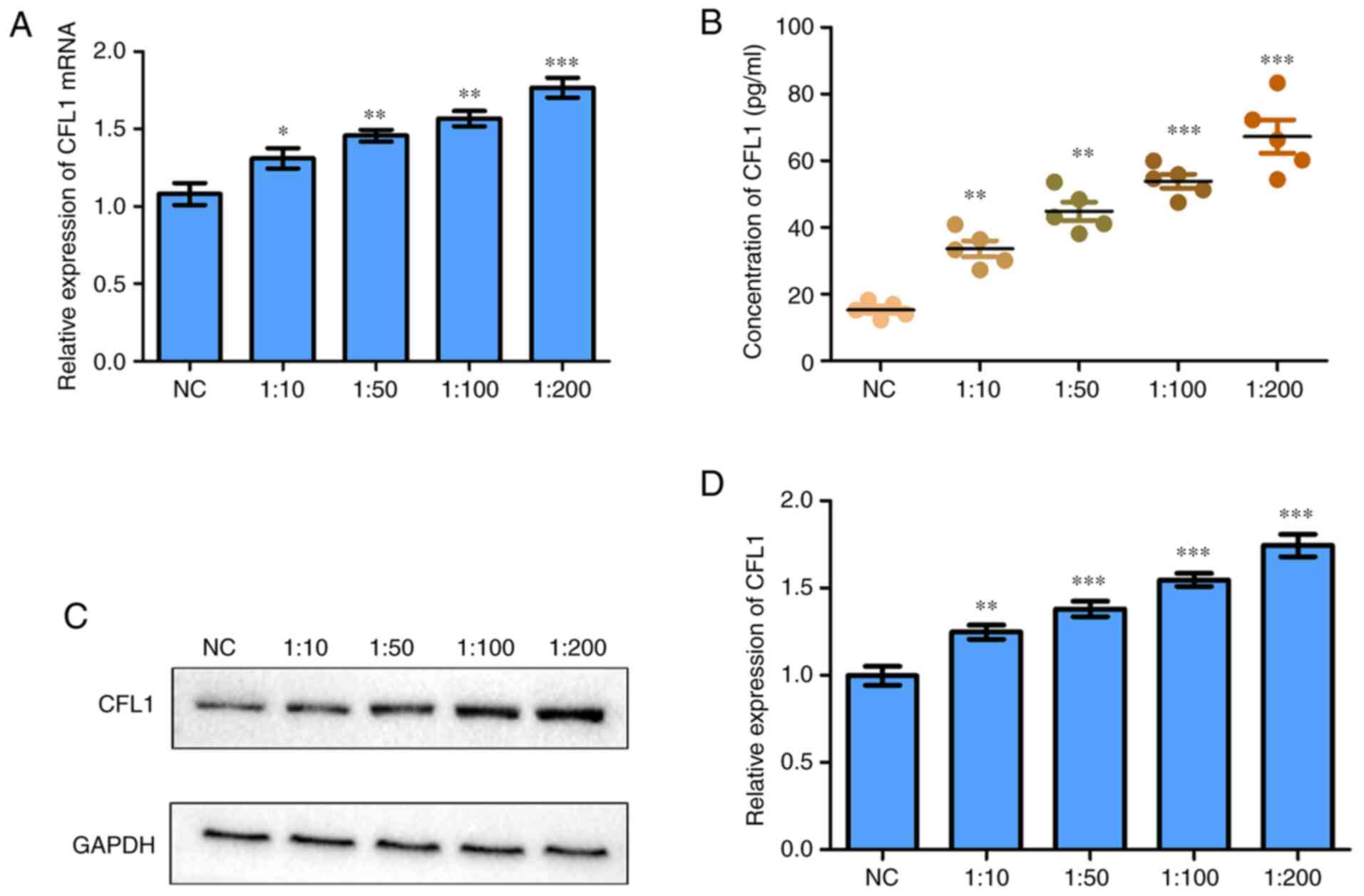
M. avium infection promotes the
expression of CFL1 in macrophages
ELISA, RT-qPCR and western blotting were performed
to detect the expression of CFL1 mRNAs and proteins in the
supernatant of M. avium infected macrophages or infected
cells. CFL1 mRNA was significantly elevated in the
macrophages as determined by RT-qPCR (Fig. 1A). Moreover, the results of ELISA
showed that CFL1 protein was significantly enhanced in M.
avium-infected macrophages (Fig.
1B). In addition, CFL1 protein were also significantly elevated
in M. avium-infected macrophages as determined by western
blotting (Fig. 1C and D).
M. avium promotes the expression of
CFL1 in macrophages
To explore the effect of M. avium on CFL1
expression in macrophages, CFL1 siRNA was transfected into
macrophages. It was observed that CFL1 mRNA expression
levels were significantly decreased in cells and M.
avium-infected macrophages transfected with siRNA compared with
the controls, whereas M. avium enhanced CFL1
expression in siRNA-transfected cells compared with the scrambled
siRNA or NC groups (Fig. 2A). In
addition, CFL1 protein expression was increased in the supernatant
and cells stimulated with M. avium compared with the
scrambled siRNA or NC groups (Fig.
2B-D). These findings revealed that M. avium could
promote CFL1 expression in macrophages.
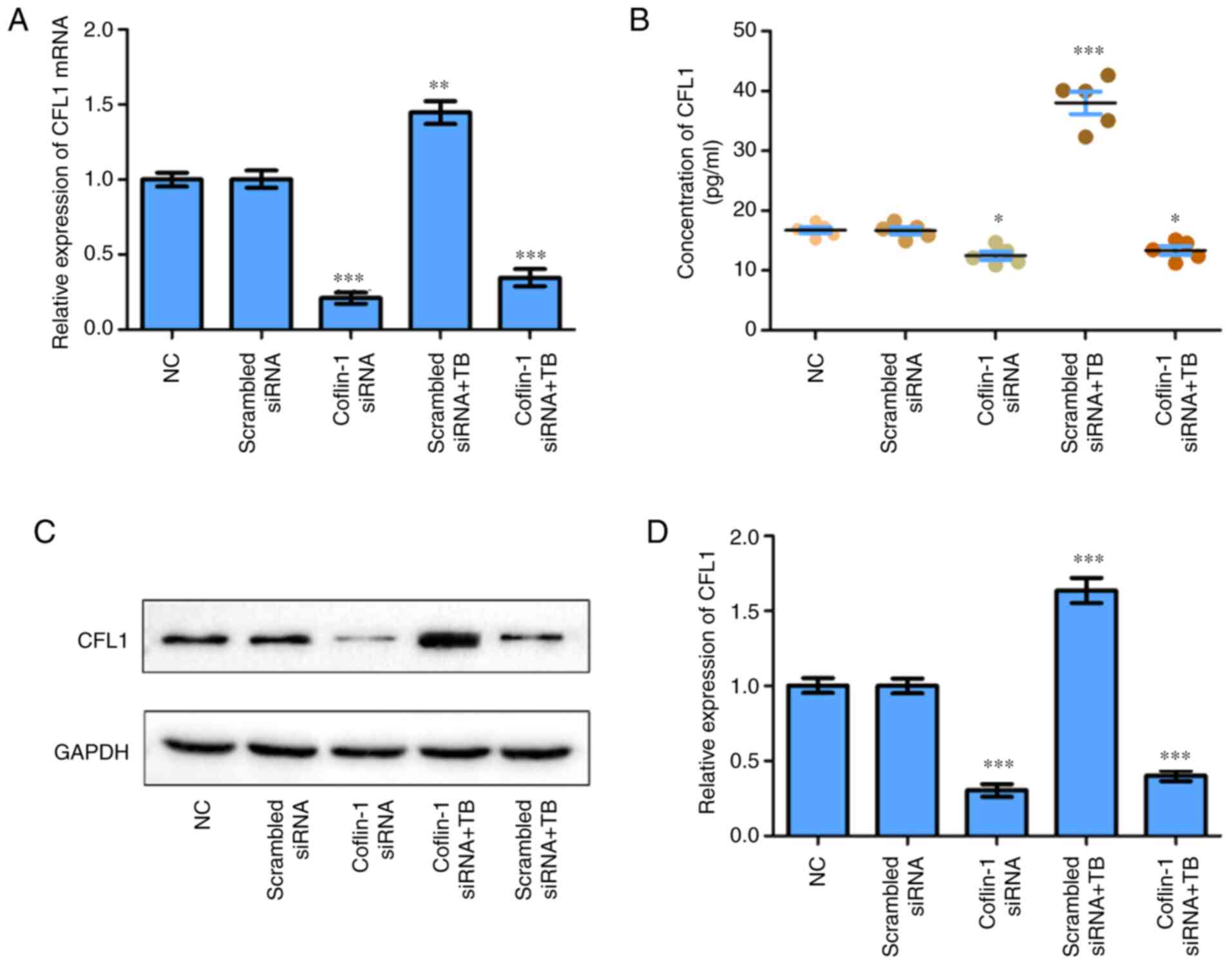 | Figure 2Expression of CFL1 in macrophages is
influenced by M. avium infection (TB). (A) Expression of
CFL1 mRNA in macrophages was regulated by CFL1 and M.
avium (TB) as determined via reverse transcription-quantitative
PCR. (B) CFL1 protein expression in the supernatant of M.
avium-infected macrophages as measured by ELISA. (C and D) CFL1
protein expression levels in CFL1-siRNA transfected macrophages or
M. avium-infected cells were detected via western blotting.
Scrambled siRNA, cofilin-1 siRNA, scrambled siRNA + TB, cofilin-1
siRNA vs. NC. *P<0.05, **P<0.005,
***P<0.001 vs. NC. NC, normal control; M.
avium, Mycobacterium avium; siRNA, small interfering
RNA; CFL1, cofilin-1; TB, tuberculosis. |
CFL1 is highly expressed in patients
infected with TB
In vitro experiments showed that infection
with M. avium could promote the expression of CFL1. RT-qPCR
results demonstrated that CFL1 mRNA expression was
significantly increased in patients with TB compared with normal
controls (Fig. 3A). In addition,
as determined by ELISA, CFL1 protein was markedly elevated in both
plasma and PBMCs from the TB patients compared with the normal
patients (Fig. 3B and C). Western blotting results showed that
the expression of CFL1 protein in the PBMCs of patients with TB was
significantly increased compared with the normal controls (Fig. 3D and E).
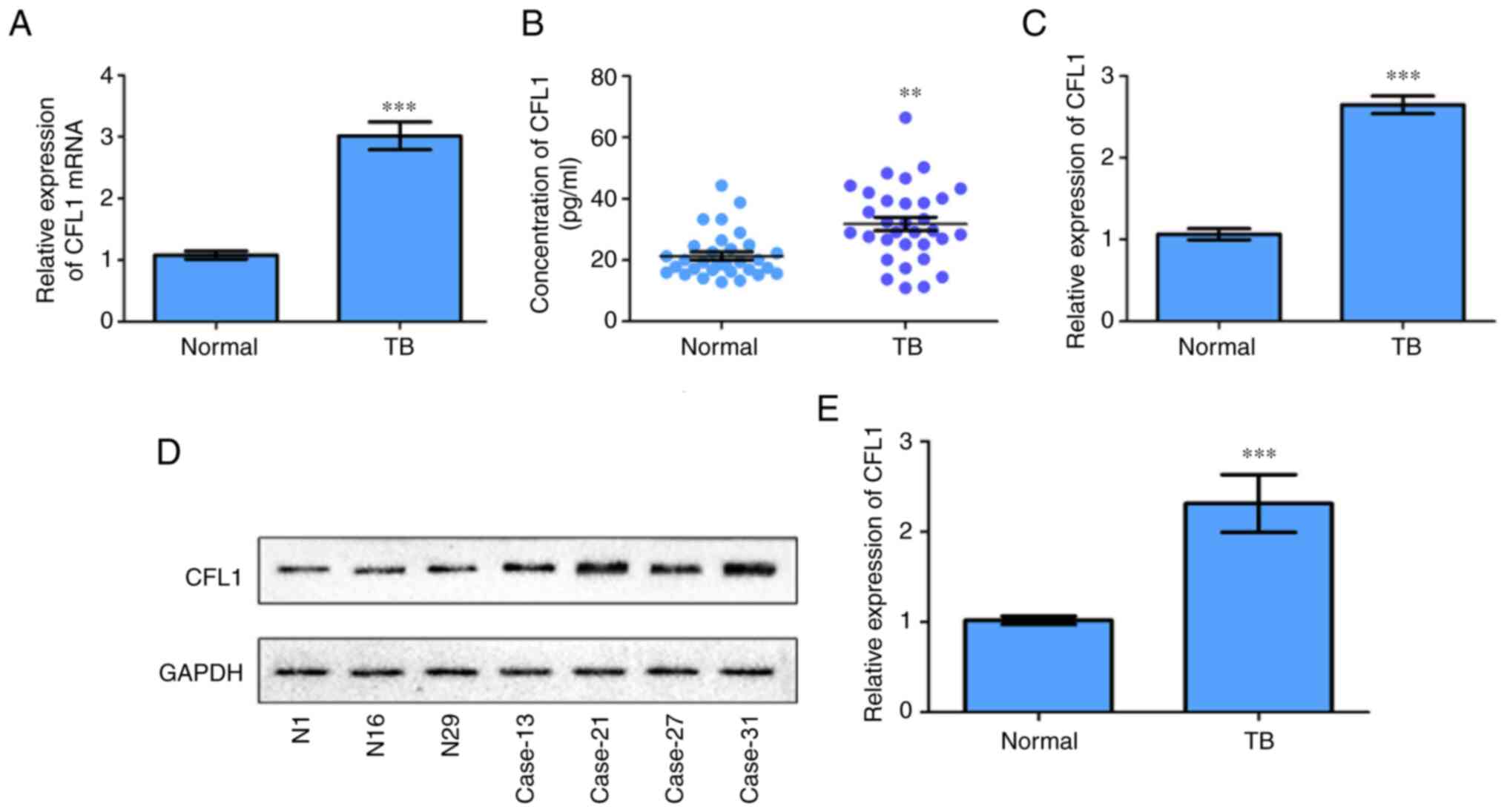 | Figure 3Expression levels of CFL1 in patients
infected with M. avium infection (TB) were quantified. (A)
CFL1 mRNA expression levels in the PBMCs of patients
infected with TB were detected via reverse
transcription-quantitative PCR. (B) The concentration of CFL1
protein in the plasma of patients infected with TB was measured by
ELISA. (C) The relative expression of CFL1 protein in PBMCs of
patients infected with TB was determined via western blotting.
**P<0.005, ***P<0.001. vs. Normal
cases. (D and E) Partial results of western blotting experiments of
CFL1 protein in PBMCs of patients infected with TB were performed
and the bands were statistically analyzed. TB cases (13#, 21#, 27#,
and 31#) vs. normal cases (N, 1#, 16#, 29#).
***P<0.001 vs. Normal. TB, tuberculosis; PBMCs,
peripheral blood mononuclear cells; CFL1, cofilin-1. |
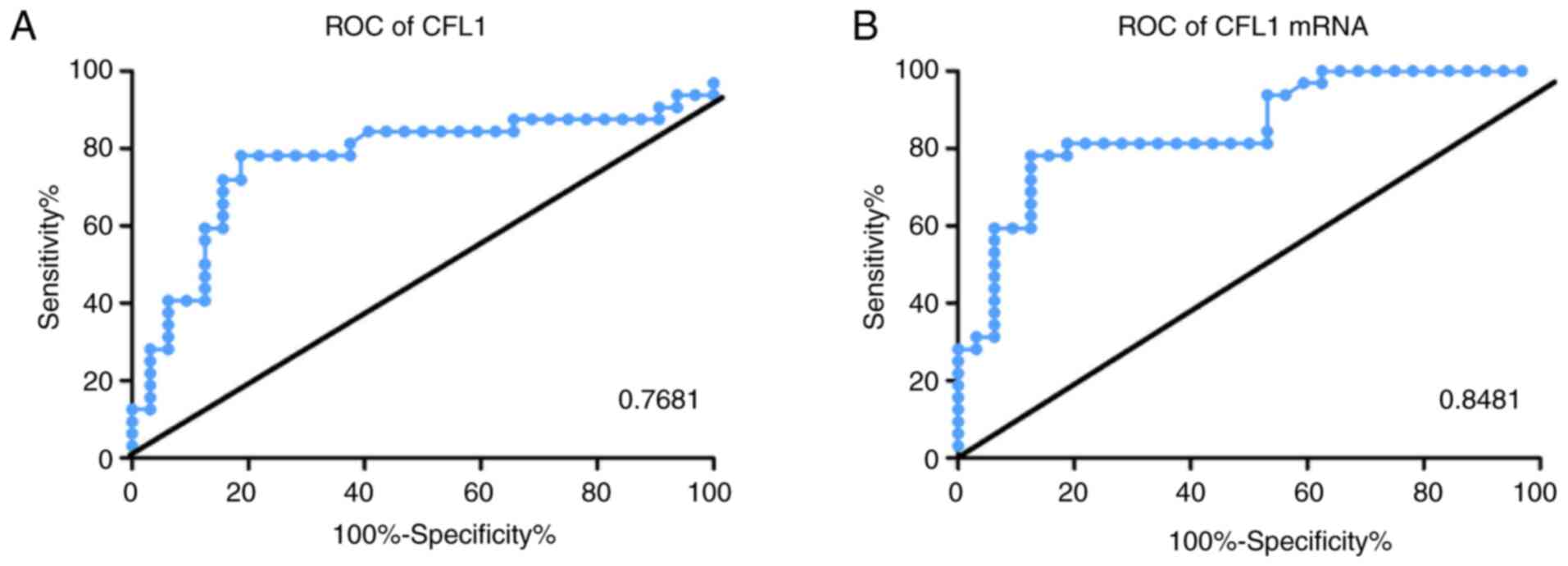
CFL1 may be an effective predictor of
patients with TB through ROC curve
ROC curve showed that CFL1 mRNA expression
was highly effective in distinguishing patients with TB from
healthy controls (AUC, 81.81%; Fig.
4A and B).
Discussion
CFL1, a primary actin-linking factor in platelets,
is required to sever actin filaments during reconstruction of the
actin cytoskeleton (21). CFL1 is
closely associated with tumor occurrence and can act as a
diagnostic marker for several types of tumors (22). For instance, excessive CFL1
expression was found to inhibit the progression and spread of
non-small cell lung cancer (23),
and CFL phosphorylation was found to be higher in bladder cancer
tissues compared with that observed in healthy bladder tissues
(24). An accumulating number of
reports have shown that CFL1 protein may mark the diagnosis and
prognosis of tumors (25-27).
For example CFL1 expression was found to be significantly increased
in the blood of patients with lung adenocarcinoma compared with
healthy individuals (28).
Moreover, previous literature suggests the
significant roles of CFL1 in microbial infection-related diseases.
For example, the activity of CFL1 following herpes simplex virus
(HSV)-1 infection may be modulated by a RhoGTPase-regulated
transduction pathway in the pathogenesis of HSV-1-induced
neurological disorders (29). CFL1
has been reported to be upregulated in exosomes derived from M.
avium-infected macrophages (17). In addition, infection by
Mycobacterium tuberculosis (M. tb) regulates
coronin1-mediated upregulation of intracellular cAMP, followed by
activation of CFL1 and the depolymerization of F-actin to suppress
phagosomal acidification and its maturation in macrophages
(30).
In the present study, upregulated CFL1 gene and
protein expression levels were found in M. avium-infected
macrophages. Meanwhile, the knockdown experiments revealed that
CFL1 expression was influenced by M. avium infection. The
expression of CFL1 in macrophages detected by immunohistochemistry
will be analyzed in future experiments.
In view of its numerous biological functions in
tumor cells and its potential as a diagnostic marker, we speculated
that CFL1 may also be used as a biomarker of TB infection. In the
in vitro experiments, CFL1 was notably upregulated in 36
patients with TB compared with healthy individuals. The CFL1
mRNA and protein expression levels of AUC in patients with TB and
controls were 0.84 and 0.76, respectively indicating that they have
good value as diagnostic markers. CFL1 expression showed diagnostic
value in detecting TB infection.
Due to the limited number of samples, this was a
preliminary study to explore the diagnostic value of CFL1 in the
peripheral blood of patients with TB. In future studies, we will
collect a large number of samples and add a sample group of
patients with latent TB. In summary, the results of the present
study indicated that CFL1 is a noteworthy target from the
perspective of TB peripheral blood diagnosis, and our research team
will continue to investigate it.
Acknowledgements
Not applicable.
Funding
Funding: This work was supported by grants from the Science and
Technology Project of Suzhou (grant no. KJXW2018068) and the
Medical Research Project of Jiangsu Health Commission (grant no.
Z2019001).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
HC and YX designed the present study. YX and ZZ
performed the experiments and wrote the manuscript, and MZ analyzed
the data and revised the manuscript. All authors read and approved
the final manuscript, and confirm the authenticity of all the raw
data.
Ethics approval and consent to
participate
Approval for the protocol of the present study was
obtained from the Ethics and Scientific Committee of the Affiliated
Kunshan Hospital of Jiangsu University, and all participants signed
an informed consent form (approval no. IEC-C-007-A07-V3.0).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Wang J, Wang Y, Tang L and Garcia RC:
Extracellular vesicles in mycobacterial infections: Their potential
as molecule transfer vectors. Front Immunol.
10(1929)2019.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Zhang M, Xie Y, Li S, Ye X, Jiang Y, Tang
L and Wang J: Proteomics analysis of exosomes from patients with
active tuberculosis reveals infection profiles and potential
biomarkers. Front Microbiol. 12(800807)2022.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Shimao T: Tuberculosis and its
control-lessons from the past and future prospect. Kekkaku.
80:481–489. 2015.PubMed/NCBI(In Japanese).
|
|
4
|
Dheda K, Barry CE III and Maartens G:
Tuberculosis. Lancet. 387:1211–1226. 2016.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Dye C, Floyd K and Uplekar M: In global
tuberculosis control: Surveillance, planning, financing. Vol. 1.
Ch., pp17-37, 2008.
|
|
6
|
Cheon SA, Cho HH, Kim J, Lee J, Kim HJ and
Park TJ: Recent tuberculosis diagnosis toward the end TB strategy.
J Microbiol Methods. 123:51–61. 2016.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Galimi R: Extrapulmonary tuberculosis:
Tuberculous meningitis new developments. Eur Rev Med Pharmacol Sci.
15:365–386. 2011.PubMed/NCBI
|
|
8
|
Furin J, Cox H and Pai M: Tuberculosis.
Lancet. 393:1642–1656. 2019.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Vartiainen MK, Mustonen T, Mattila PK,
Ojala PJ, Thesleff I, Partanen J and Lappalainen P: The three mouse
actin-depolymerizing factor/cofilins evolved to fulfill
cell-type-specific requirements for actin dynamics. Mol Biol Cell.
13:183–194. 2020.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Percipalle P: Co-transcriptional nuclear
actin dynamics. Nucleus. 4:43–52. 2013.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Obrdlik A and Percipalle P: The F-actin
severing protein cofilin-1 is required for RNA polymerase II
transcription elongation. Nucleus. 2:72–79. 2011.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Wiggan O, Schroder B, Krapf D, Bamburg JR
and DeLuca JG: Cofilin regulates nuclear architecture through a
myosin-II dependent mechanotransduction module. Sci Rep.
7(40953)2017.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Wang W, Mouneimne G, Sidani M, Wyckoff J,
Chen X, Makris A, Goswami S, Bresnick AR and Condeelis JS: The
activity status of cofilin is directly related to invasion,
intravasation, and metastasis of mammary tumors. J Cell Biol.
173:395–404. 2006.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Ono S: Mechanism of depolymerization and
severing of actin filaments and its significance in cytoskeletal
dynamics. Int Rev Cytol. 258:1–82. 2007.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Hotulainen P, Paunola E, Vartiainen MK and
Lappalainen P: Actin-depolymerizing factor and cofilin-1 play
overlapping roles in promoting rapid F-actin depolymerization in
mammalian nonmuscle cells. Mol Biol Cell. 16:649–664.
2005.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Kapoor S: Cofilin-1 overexpression and its
role in tumor growth and progression in systemic malignancies. Int
J Radiat Biol. 90(113)2014.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Wang JJ, Chen C, Xie PF, Pan Y, Tan YH and
Tang LJ: Proteomic analysis and immune properties of exosomes
released by macrophages infected with Mycobacterium avium.
Microbes Infect. 16:283–291. 2014.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Wang J, Yao Y, Wu J, Deng Z, Gu T, Tang X,
Cheng Y and Li G: The mechanism of cytoskeleton protein β-actin and
cofilin-1 of macrophages infected by Mycobacterium avium. Am
J Transl Res. 8:1055–1063. 2016.PubMed/NCBI
|
|
19
|
World Health Organization; International
Union Against Tuberculosis and Lung Disease; Royal Netherlands
Tuberculosis Association: Revised international definitions in
tuberculosis control. Int J Tuberc Lung Dis 5: 213: 215, 2001.
|
|
20
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408.
2001.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Dasgupta SK and Thiagarajan P:
Cofilin-1-induced actin reorganization in stored platelets.
Transfusion. 60:806–814. 2020.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Shishkin S, Eremina L, Pashintseva N,
Kovalev L and Kovaleva M: Cofilin-1 and other ADF/cofilin
superfamily members in human malignant cells. Int J Mol Sci.
18(10)2016.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Tsai CH, Lin LT, Wang CY, Chiu YW, Chou
YT, Chiu SJ, Wang HE, Liu RS, Wu CY, Chan PC, et al:
Over-expression of cofilin-1 suppressed growth and invasion of
cancer cells is associated with up-regulation of let-7 microRNA.
Biochim Biophys Acta. 1852:851–861. 2015.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Chung H, Kim B, Jung SH, Won KJ, Jiang X,
Lee CK, Lim SD, Yang SK, Song KH and Kim HS: Does phosphorylation
of cofilin affect the progression of human bladder cancer? BMC
Cancer. 13(45)2013.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Jiang N, Kham SK, Koh GS, Suang Lim JY,
Ariffin H, Chew FT and Yeoh AE: Identification of prognostic
protein biomarkers in childhood acute lymphoblastic leukemia (ALL).
J Proteomics. 74:843–857. 2011.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Peng XC, Gong FM, Zhao YW, Zhou LX, Xie
YW, Liao HL, Lin HJ, Li ZY, Tang MH and Tong AP: Comparative
proteomic approach identifies PKM2 and cofilin-1 as potential
diagnostic, prognostic and therapeutic targets for pulmonary
adenocarcinoma. PLoS One. 6(e27309)2011.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Guan M, Chen X, Ma Y, Tang L, Guan L, Ren
X, Yu B, Zhang W and Su B: MDA-9 and GRP78 as potential diagnostic
biomarkers for early detection of melanoma metastasis. Tumour Biol.
36:2973–2982. 2015.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Zheng Y, Fang Y, Li S and Zheng B:
Detection of plasma cofilin protein for diagnosis of lung cancer.
Nan Fang Yi Ke Da Xue Xue Bao. 33:1551–1553. 2013.PubMed/NCBI(In Chinese).
|
|
29
|
Xiang Y, Zheng K, Ju H, Wang S, Pei Y,
Ding W, Chen Z, Wang Q, Qiu X, Zhong M, et al: Cofilin 1-mediated
biphasic F-actin dynamics of neuronal cells affect herpes simplex
virus 1 infection and replication. J Virol. 86:8440–8451.
2012.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Saha S, Hazra A, Ghatak D, Singh AV, Roy S
and BoseDasgupta S: A bumpy ride of mycobacterial phagosome
maturation: Roleplay of coronin1 through Cofilin1 and cAMP. Front
Immunol. 12(687044)2021.PubMed/NCBI View Article : Google Scholar
|