Introduction
Parkinson's disease (PD) is a common age-related
neurodegenerative disease, with the age of onset ranging between 40
and 70 years (1,2). Currently, the prevalence of PD in
individuals aged ≥60 years in China is 1-2% (3). The seventh national census released
in May 2021 reported that ~18.7% of China's population is aged
>60 years, which suggests that the number of new cases of PD
will increase annually as a result of the aging population
(4). PD significantly impacts the
quality of life of those affected, whereby patients face losing
their independence and becoming reliant on nursing care in the
advanced stages of the disease (5). For this reason, research on the
pathogenesis and treatment of PD has major social and economic
implications. However, the lack of an in-depth understanding of PD
has led to a limited range of therapeutic strategies (6). Levodopa is currently the mainstay
symptomatic treatment for PD and can improve symptoms such as
muscular rigidity and postural instability; however, it cannot stop
disease progression (7). The side
effects of levodopa include nausea, agitation, psychological
symptoms and abnormal limb movement, with the efficacy of the drug
decreasing over a period of 3-5 years. However, even with
treatment, neuronal apoptosis continues to exacerbate the disease
(8). Therefore, the main focus of
PD research is to explore the etiology and pathogenesis of PD in
order to develop alternative therapeutic methods.
Oxidative stress is a major cause of neuronal
apoptosis in PD and, therefore, the discovery and development of
successful antioxidant treatments is an important focus of current
PD research (9). Traditional
Chinese herbal medicines are considered to be natural sources of
antioxidants, among which flavonoid compounds have been
demonstrated to exhibit pharmacological properties as a result of
their diverse bioactivity (10,11).
Tilianin is a natural polyphenolic flavonoid isolated from
Dracocephalum moldavica L. amiales and it has a variety of
pharmacological properties, including neuroprotective,
cardioprotective, antihypertensive, anti-atherosclerotic,
antioxidant, anti-inflammatory and antidiabetic effects (12). Previous studies have reported the
role of tilianin against oxidative stress in cerebral
ischemia/reperfusion injury via inhibition of the p38 MAPK
signaling pathway, as well as in tracheal epithelial cells via
inhibition of the ERK signaling pathway (13,14).
Based on these earlier studies, the aim of the present study was to
explore whether tilianin could protect against the inflammation and
oxidative stress that damages dopaminergic neurons in PD, and to
determine the role of the MAPK signaling pathway in this
mechanism.
Materials and methods
Cell culture and treatment
The dopaminergic neuron MES23.5 cell line (cat. no.
CVCL-J351; http://www.biovector.net/product/2277098.html) was
acquired from the BioVector National Type Culture Collection, Inc.
MES23.5 cells were maintained in DMEM (Thermo Fisher Scientific,
Inc.) supplemented with 10% FBS (Merck KGaA), 2% Sato's solution
containing 25 mg insulin, transferrin, selenium and sodium pyruvate
solution (ITS-A; cat. no. 51300044; Thermo Fisher Scientific,
Inc.), 0.315 mg/ml progesterone (cat. no. HY-N0437; MedChemExpress)
and 20 mg putrescine (cat. no. HY-N2407; MedChemExpress), in
addition to 100 U/ml penicillin/streptomycin in a humidified
atmosphere containing 5% CO2 at 37˚C. To construct the
PD model, MES23.5 cells were treated with 300 µmol/l
1-methyl-4-phenylpyridinium (MPP+; Sigma-Aldrich; Merck
KGaA) at 37˚C for 24 h, whereas untreated MES23.5 cells cultured
for 24 h at 37˚C in normal medium were used as a control. Tilianin
(Chengdu Pufei De Biotech Co., Ltd.) at concentrations of 0, 1, 3,
10 and 30 µM were selected for cell pretreatment in order to assess
its effects on cell viability (14).
Analysis of cell viability
Cell viability was analyzed using the Cell Counting
Kit-8 (CCK-8) assay (Beijing Solarbio Science & Technology Co.,
Ltd.). Briefly, following MES23.5 cell treatment with tilianin and
MPP+ (300 µmol/l) in a 96-well plate (2x104
cells/well) at 37˚C for 24, 48 and 72 h, the viability of the cells
was determined using 10 µl CCK-8 assay for 2 h according to the
manufacturer's protocol. The optical density at a wavelength of 450
nm was quantified using a microplate reader.
Immunofluorescence (IF) staining
Tyrosine hydroxylase (TH) expression was detected
via IF staining. MES23.5 cells (1x105 cells/well) were
fixed with 4% paraformaldehyde at room temperature for 15 min,
permeabilized with 0.5% Triton X-100 and then blocked with 5% goat
serum (Gibco; Thermo Fisher Scientific, Inc.) at 37˚C for 30 min.
Following incubation with mouse anti-TH primary antibody (1:250;
cat. no. ab137869; Abcam) at 4˚C overnight, cells were further
incubated with Alexa Fluor 350-conjugated goat anti-mouse secondary
antibody (1:250; cat. no. A0412; Beyotime Institute of
Biotechnology) for 15 min at 37˚C. Subsequently, cells were washed
three times with PBS and stained using 0.5 µg/ml DAPI (Beijing
Solarbio Science & Technology Co., Ltd.) at room temperature
for 15 min. TH-positive cells were counted in three randomly
selected visual fields using a fluorescence microscope
(magnification, x200).
Reverse transcription-quantitative PCR
(RT-qPCR)
Total RNA was extracted from MES23.5 cells using
TRIzol® reagent (Invitrogen; Thermo Fisher Scientific,
Inc.). The Hifair® III 1st Strand cDNA Synthesis
SuperMix kit (Shanghai Yeasen Biotechnology Co., Ltd.) was used to
reverse transcribe total RNA into cDNA using the reaction of at
25˚C for 5 min, 42˚C for 30 min, 85˚C for 5 min and 4˚C for 5 min.
qPCR was subsequently performed using a Hifair® III One
Step RT-qPCR SYBR Green Kit (Shanghai Yeasen Biotechnology Co.,
Ltd.) using an ABI 7500 thermocycler (Applied Biosystems; Thermo
Fisher Scientific, Inc.) The qPCR primers were as follows: TH
forward, 5'-GCCGTCTCAGAGCAGGATAC-3' and reverse,
5'-ACCTCGAAGCGCACAAAGTA-3'; IL-6 forward,
5'-AAAGAGGCACTGGCAGAAAA-3' and reverse, 5'-CAGGGGTGGTTATTGCATCT-3';
IL-1β forward, 5'-TACGAATCTCCGACCACCACTACAG-3' and reverse,
5'-TGGAGGTGGAGAGCTTTCAGTTCATATG-3'; TNF-α forward,
5'-AGGCACTCCCCCAAAAGATG-3' and reverse, 5'-ATAGCAAATCGGCTGACGGT-3'
and GAPDH forward, 5'-CTACCCCCAATGTGTCCGTC-3' and reverse,
5'-GGCCTCTCTTGCTCAGTGTC-3'. The following thermocycling conditions
were used for qPCR: Pre-denaturation was performed at 95˚C for 5
min, followed by 40 cycles of denaturation at 95˚C for 10 sec and
annealing at 60˚C for 30 sec, as well as elongation at 72˚C for 10
min. After normalization using GAPDH as an internal standard gene,
relative mRNA expression levels were quantified and analyzed using
the 2-ΔΔCq method (15).
Western blotting
Total protein was extracted from MES23.5 cells using
RIPA lysis buffer (Shanghai Yeasen Biotechnology Co., Ltd.).
Protein quantification was performed using a BCA kit (Shanghai
Yeasen Biotechnology Co., Ltd.). Total protein (20 µg protein/lane)
was separated by SDS-PAGE on a 12% gel. The separated proteins were
subsequently transferred onto a PVDF membrane and blocked with 5%
skimmed milk for 1 h at room temperature. The membranes were
incubated overnight at 4˚C with primary antibodies against TH (cat.
no. ab137869, 1:250; Abcam), manganese superoxide dismutase (MnSOD)
(cat. no. ab68155, 1:1,000; Abcam), catalase (cat. no. ab209211,
1:2,000; Abcam), Bcl-2 (cat. no. ab182858, 1:2,000; Abcam), Bax
(cat. no. ab32503, 1:1,000; Abcam), cleaved caspase-3 (cat. no.
ab32042; 1:500; Abcam), phosphorylated (p)-ERK1/2 (cat. no.
ab201015, 1:1,000; Abcam), ERK1/2 (cat. no. ab184699, 1:10,000;
Abcam), p-p38 (cat. no. 4511, 1:1,000; Cell Signaling Technology,
Inc.), p38 (cat. no. 8690, 1:1,000; Cell Signaling Technology,
Inc.), p-JNK (cat. no. 4668, 1:1,000; Cell Signaling Technology,
Inc.) and JNK (cat. no. 9252, 1:1,000; Cell Signaling Technology,
Inc.). Subsequently, the membranes were incubated with mouse
anti-rabbit IgG secondary antibodies conjugated to HRP (1:5,000;
cat. no. sc-2357, Santa Cruz Biotechnology, Inc.) at room
temperature for 2 h. Protein bands were visualized using Enhanced
Chemiluminescence Detection Reagent (MilliporeSigma). ImageJ
software Version 1.49 (National Institutes of Health) was used to
analyze the chemiluminescent signals. Membranes were probed with
anti-GAPDH antibody (Abcam, cat. no. ab9485, 1:2,500) as a loading
control.
Analysis of oxidative stress
A Reactive Oxygen Species (ROS) Assay Kit (cat. no.
C1300-1; Applygen Technologies, Inc.) was used to detect ROS
production using a fluorescence microscope (magnification, x40)
according to the manufacturer's protocol.
Cell apoptosis assay
The TUNEL Apoptosis Detection (FITC) Kit (Shanghai
Qcbio Science & Technologies Co., Ltd.) was used to observe the
apoptotic rate of MES23.5 cells. Briefly, the cells were fixed with
4% paraformaldehyde at room temperature for 30 min and
permeabilized with 0.1% Triton X-100 at room temperature for 3 min.
The permeabilized samples were then treated with DNase I at 37˚C
for 30 min to prepare the positive control slides. Following
incubation with 50 µl TUNEL working solution at 37˚C for 1 h in the
dark, the slides were immersed in DAPI solution (2 µg/ml, diluted
with PBS) for 5 min at room temperature. The samples were sealed
with VECTASHIELD® Antifade Mounting Medium (Vector
Laboratories, Inc.; Maravai LifeSciences) and then five regions of
apoptotic cells were randomly selected for viewing under a
fluorescence microscope (magnification, x20). The green
fluorescence at 520±20 nm was observed using a standard filter. The
blue fluorescence of DAPI was observed at 460 nm.
Statistical analysis
Data analysis was performed using GraphPad Prism 6
(GraphPad Software, Inc.). Data are presented as the mean ± SD.
Each experiment was conducted in triplicate. Differences among
multiple groups were analyzed using one-way ANOVA with a post hoc
Bonferroni multiple comparison test. P<0.05 was considered to
indicate a statistically significant difference.
Results
Effect of tilianin on the viability of
MES23.5 cells
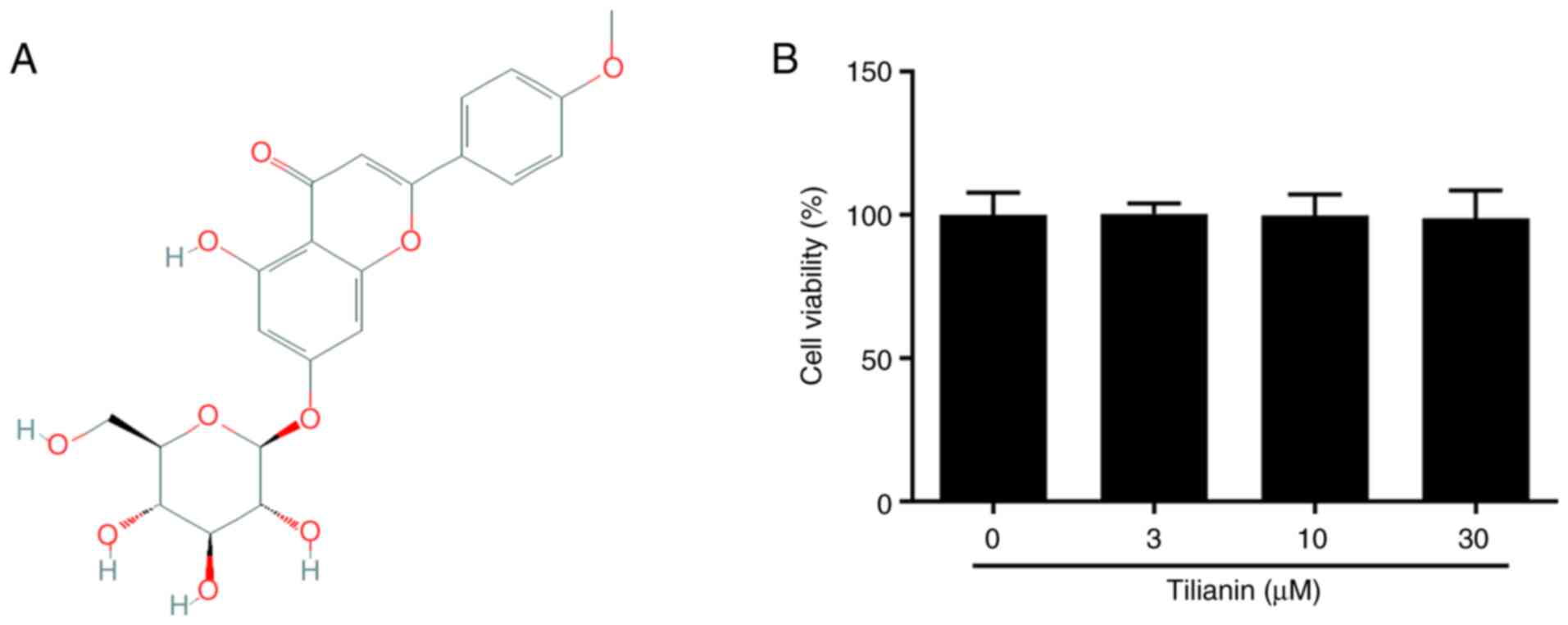
The chemical structure of tilianin is shown in
Fig. 1A. Cell viability analysis
demonstrated that different concentrations of tilianin did not
affect the viability of MES23.5 cells (Fig. 1B). Therefore, these results
suggested that tilianin exerted no cytotoxic effects on MES23.5
cells.
Effect of tilianin on
MPP+-induced loss of cell viability and TH
deficiency
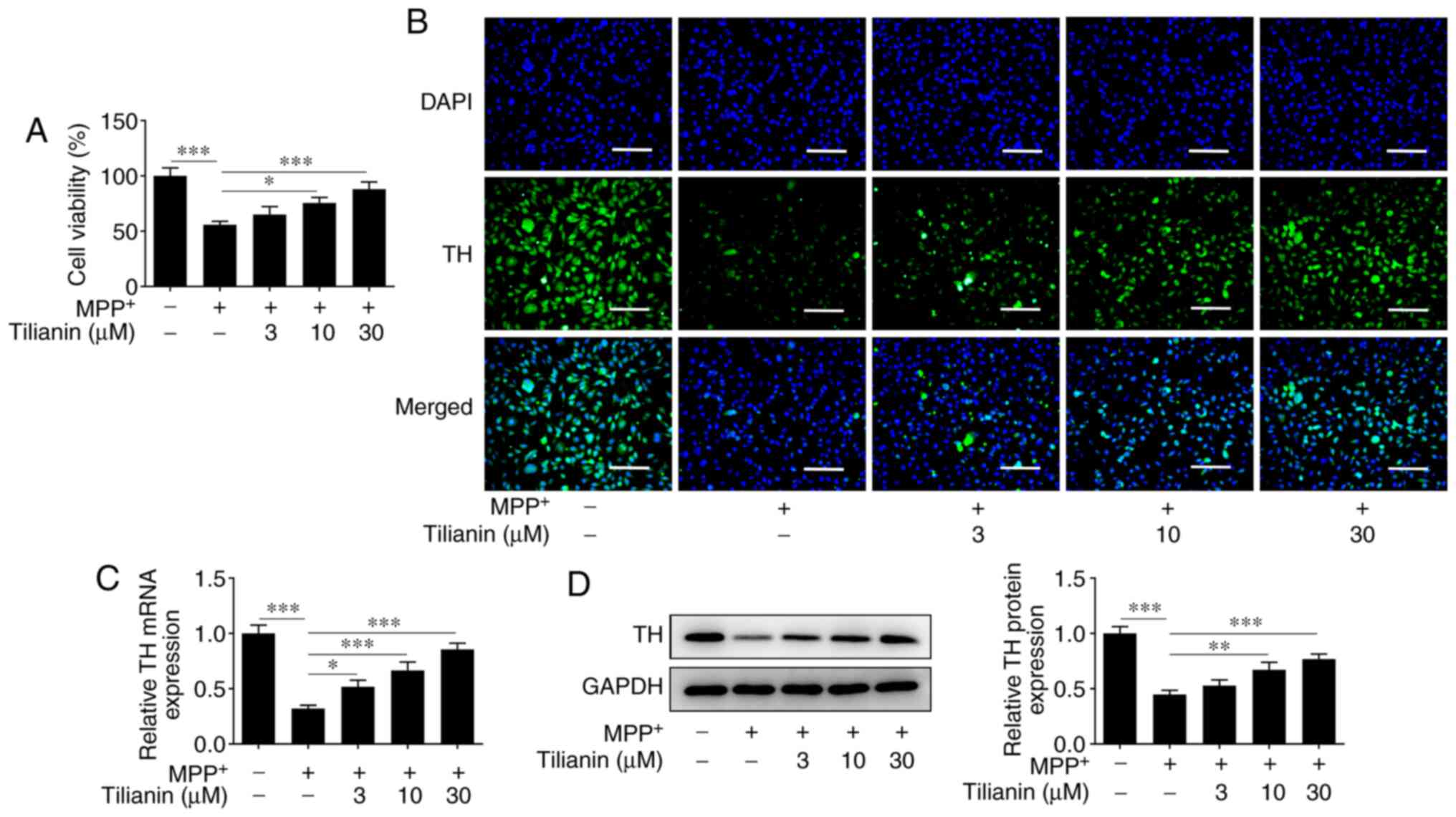
MPP+-stimulated MES23.5 cells exhibited
reduced viability. However, pretreatment with tilianin improved the
viability of MMP+-induced cells in a dose-dependent
manner (Fig. 2A). Moreover, the
results of the IF analysis demonstrated that MPP+
decreased the TH protein expression levels in MES23.5 cells
compared with those in the control group, whereas tilianin
pretreatment increased the TH levels in MMP+-induced
cells in a dose-dependent manner (Fig.
2B). Similar results were observed using RT-qPCR and western
blotting (Fig. 2C and D). These results indicated that tilianin
may effectively protect MES23.5 cells from MPP+-induced
reduction in viability and TH deficiency.
Effect of tilianin on the
MPP+-induced inflammatory response and oxidative
stress
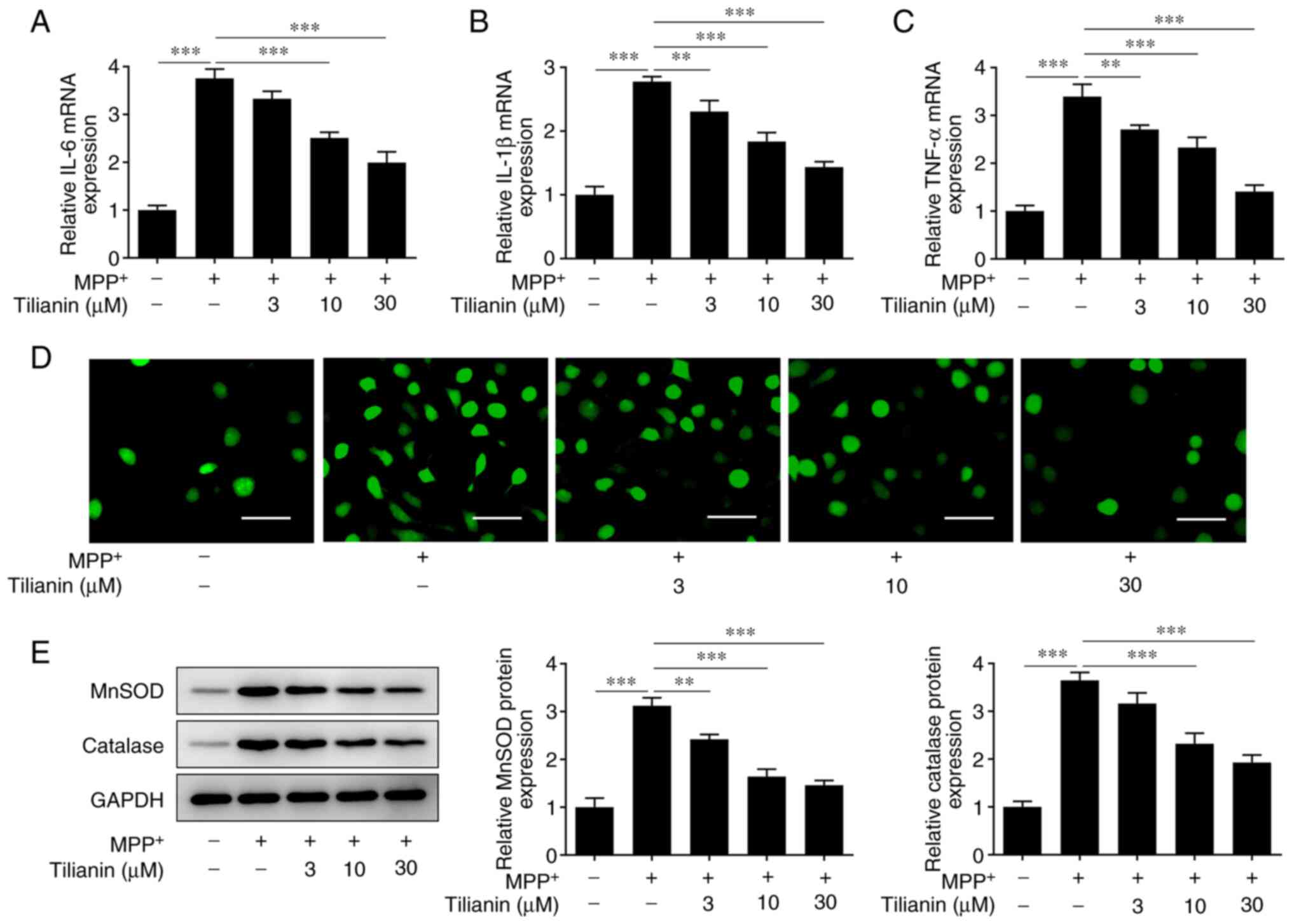
Further experiments revealed that MPP+
elevated the mRNA expression levels of the pro-inflammatory
cytokines IL-6, IL-1β and TNF-α in MES23.5 cells, whereas tilianin
pretreatment reduced their mRNA expression levels in a
dose-dependent manner (Fig. 3A-C).
It was also observed that MPP+-induced ROS production
decreased with increasing tilianin concentrations (Fig. 3D). Moreover, the protein expression
levels of MnSOD and catalase were found to be increased in
MPP+-stimulated cells, whereas these expression levels
were decreased in the tilianin pretreatment groups (Fig. 3E). These results demonstrated that
tilianin may ameliorate the MPP+-induced inflammatory
response and oxidative stress in MES23.5 cells.
Effect of tilianin on
MPP+-induced cell apoptosis
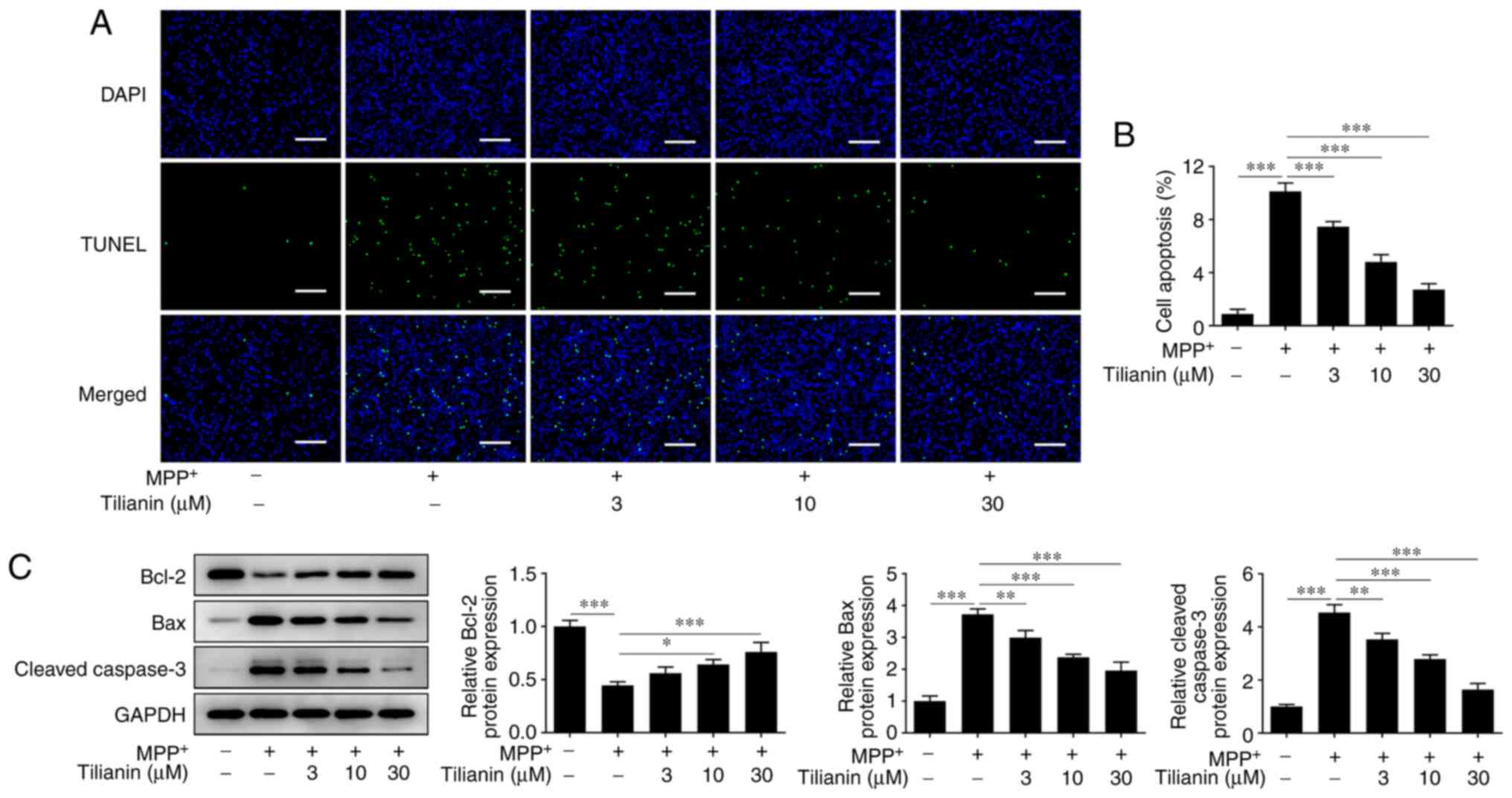
As shown in Fig. 4A
and B, an increase in the number
of TUNEL-positive (apoptotic) MES23.5 cells was observed in the
MPP+ group, whereas the number of apoptotic cells was
reduced by tilianin pretreatment in a dose-dependent manner.
Furthermore, the protein expression levels of the antiapoptotic
gene Bcl-2 were shown to be downregulated by MPP+
stimulation, whereas tilianin pretreatment upregulated Bcl-2
expression levels in MPP+-stimulated MES23.5 cells
(Fig. 4C). By contrast, the
protein expression levels of the apoptosis markers Bax and cleaved
caspase-3 were upregulated in MPP+-stimulated MES23.5
cells, but were downregulated following tilianin pretreatment.
Therefore, these results indicated that tilianin may prevent the
MPP+-induced apoptosis of MES23.5 cells.
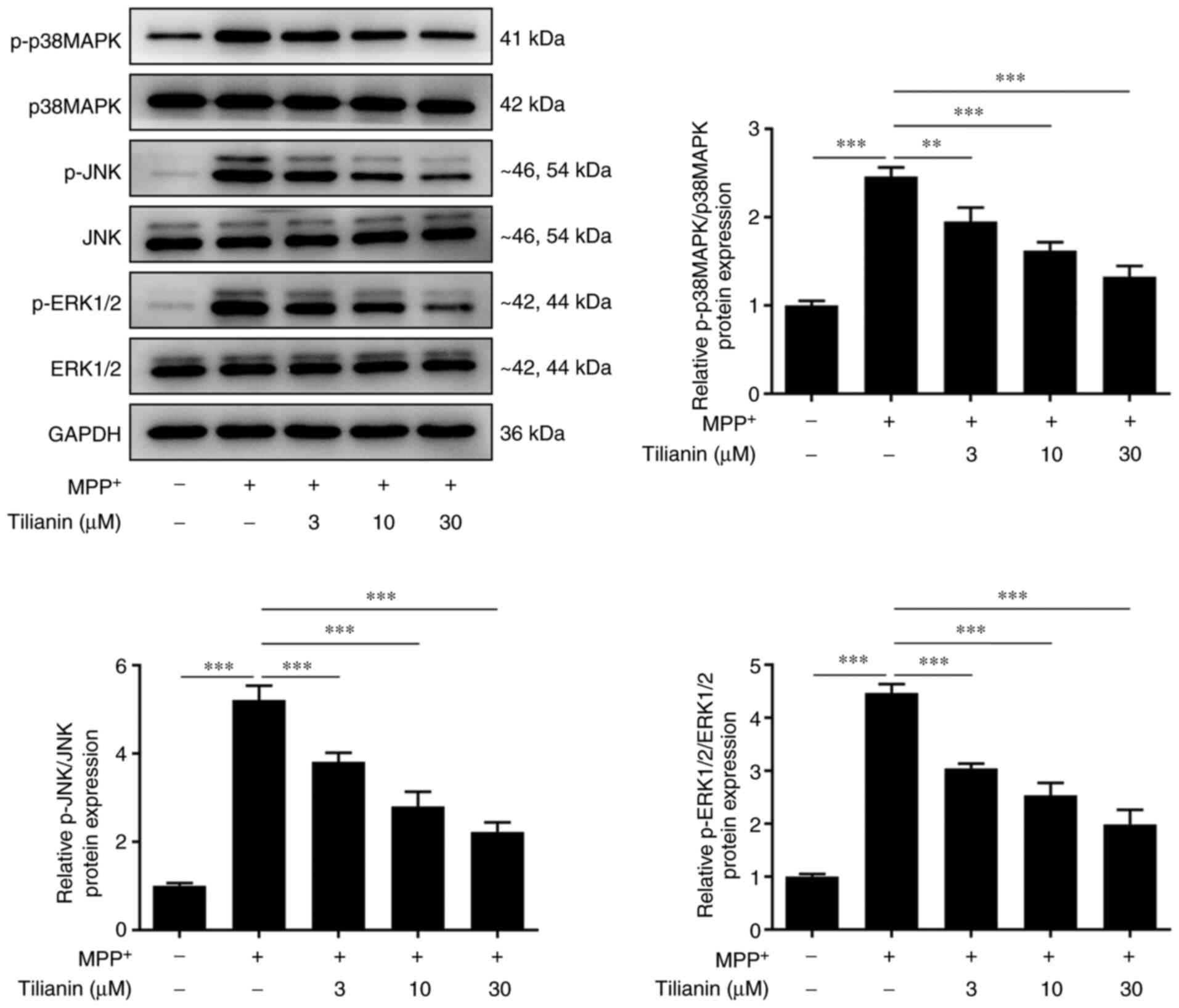
Effect of tilianin on the MAPK
signaling pathway in MPP+-stimulated MES23.5 cells
Western blotting demonstrated that the expression
levels of MAPK signaling pathway-related proteins (p-p38, p-ERK1/2
and p-JNK) were high in MPP+-stimulated MES23.5 cells,
and these protein expression levels were downregulated in a
dose-dependent manner when cells were pretreated with tilianin
(Fig. 5). These results suggested
that tilianin may inhibit MPP+-induced activation of the
p38, ERK1/2 and JNK signaling pathways in MES23.5 cells.
Discussion
The main pathological manifestations of PD are the
degeneration and loss of dopaminergic neurons in the substantia
nigra compacta in the midbrain; the formation of intraneuronal
inclusions, known as Lewy bodies, in the residual dopamine neurons;
and the appearance of dystrophic synapses (16-18).
The exact etiology of the degeneration and death of dopaminergic
neurons in PD remains unclear, but may involve numerous factors,
including genetics, environmental factors, aging and oxidative
stress (19,20). The exacerbation of oxidative stress
is considered to be an important contributor to dopaminergic neuron
injury in the pathogenesis of PD (21,22).
Oxidative stress signaling pathways can transmit
messages outside of cells and, therefore, cells rely on internal
signaling pathways to transduce signals intracellularly (23). As a downstream target of oxidative
stress, the MAPK signaling pathway transmits extracellular
oxidative stress signals inside the cell (24). The MAPK family is a group of
conserved serine/threonine protein kinases, which are major
signaling molecules in signal transduction and can form signaling
pathways that are crucial for signal transmission networks in
eukaryotic cells (25). Among MAPK
family members, the ERK1/2 signal transduction pathway regulates
cell proliferation and differentiation (26), whereas JNK and p38, collectively
named the MAPK stress signaling pathway, serve an important role in
stress responses, such as inflammation and apoptosis (27).
Tilianin, the main active component in D.
moldavica L., acts as a cardioprotective agent in myocardial
ischemia/reperfusion injury in rats via improving mitochondrial
dysfunction, inhibiting oxidative stress and, thereby, alleviating
cardiomyocyte apoptosis (28).
Previous studies have reported the role of tilianin against
oxidative stress in cerebral ischemia/reperfusion injury through
inhibiting p38 expression, whereas in tracheal epithelial cells
tilianin has been demonstrated to inhibit the ERK signaling
pathway, which also reduces oxidative stress (13,14).
In addition, Jiang et al (29) reported that tilianin ameliorated
memory impairment and neurodegeneration by inhibition of neuronal
apoptosis and inflammation in the hippocampus of rats with
permanent occlusion of the bilateral common carotid artery via
increasing p-CaMKII/ERK/CREB signal transduction. In addition, a
previous study has demonstrated that tilianin exhibits low
cytotoxicity (30). In the present
study, different concentrations of tilianin did not affect the
viability of MES23.5 cells, whereas tilianin pretreatment improved
the viability in MPP+-stimulated MES23.5 cells in a
dose-dependent manner. All tilianin pretreatment groups exhibited
lower pro-inflammatory cytokine mRNA expression levels, and
downregulated ROS levels and MnSOD and catalase protein expression
levels. MPP+-induced apoptosis of dopaminergic neurons
was also effectively alleviated by tilianin in a dose-dependent
manner. Furthermore, a dose-dependent decline in the expression of
MAPK signaling pathway-related proteins (p-p38, p-ERK1/2 and p-JNK)
was observed in MPP+-stimulated MES23.5 cells following
tilianin preconditioning.
Dopaminergic neuron injury leading to dysfunctional
dopamine synthesis is central to the onset of PD (31). TH, a rate-limiting enzyme in
dopamine biosynthesis that is mainly expressed in the brain and the
adrenal gland, is used by neurons to synthesize dopamine following
ingestion of tyrosine (32,33).
Overall, the function and expression of TH are important in
dopamine synthesis. In the present study, TH mRNA and protein
expression levels were decreased following MPP+
stimulation; however, tilianin preconditioning significantly
enhanced the mRNA and protein expression levels of TH in
MPP+-stimulated MES23.5 cells. This result suggested
that tilianin may prevent the degradation of TH in PD. The present
study mainly explored the effects of tilianin on PD in vitro
and the results revealed that tilianin attenuated
MPP+-induced oxidative stress and apoptosis of
dopaminergic neurons in a cellular model of PD. However, the
effects of tilianin on animals or humans with PD were not
investigated, and the protective role of tilianin in PD in
vivo will be further investigated and verified in future
studies.
In conclusion, the data in the present study
demonstrated that tilianin may serve an anti-inflammatory and
antioxidant role by inhibiting the MAPK signaling pathway, which
may suppress dopaminergic neuron injury in PD. Therefore, tilianin
may hold promise as a novel therapeutic agent in the treatment of
PD. However, data from further in vivo experiments and
clinical trials are needed to support the conclusions of the
present study.
Acknowledgements
Not applicable.
Funding
Funding: The present study was supported by Suzhou Vocational
Health College (grant no. SZWZY201703).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
JL and SX designed the study, performed the
experiments and drafted and revised the manuscript. JL analyzed the
data and SX performed the literature search. Both authors confirmed
the authenticity of the raw data and have read and approved the
final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Wickremaratchi MM, Ben-Shlomo Y and Morris
HR: The effect of onset age on the clinical features of Parkinson's
disease. Eur J Neurol. 16:450–456. 2009.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Rizek P, Kumar N and Jog MS: An update on
the diagnosis and treatment of Parkinson disease. CMAJ.
188:1157–1165. 2016.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Cui L, Hou NN, Wu HM, Zuo X, Lian YZ,
Zhang CN, Wang ZF, Zhang X and Zhu JH: Prevalence of Alzheimer's
disease and parkinson's disease in China: An updated systematical
analysis. Front Aging Neurosci. 12(603854)2020.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Tang Q, Wang C, Wu W, Cao Y, Chen G and Lu
J: China should emphasize key issues inherent in rational
medication management for the elderly. Biosci Trends. 15:262–265.
2021.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Ng JSC: Palliative care for Parkinson's
disease. Ann Palliat Med. 7:296–303. 2018.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Armstrong MJ and Okun MS: Diagnosis and
treatment of Parkinson disease: A review. JAMA. 323:548–560.
2020.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Pezzoli G and Zini M: Levodopa in
Parkinson's disease: From the past to the future. Expert Opin
Pharmacother. 11:627–635. 2010.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Vasta R, Nicoletti A, Mostile G, Dibilio
V, Sciacca G, Contrafatto D, Cicero CE, Raciti L, Luca A and Zappia
M: Side effects induced by the acute levodopa challenge in
Parkinson's disease and atypical parkinsonisms. PLoS One.
12(e0172145)2017.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Trist BG, Hare DJ and Double KL: Oxidative
stress in the aging substantia nigra and the etiology of
Parkinson's disease. Aging Cell. 18(e13031)2019.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Bai L, Li X, He L, Zheng Y, Lu H and Li J,
Zhong L, Tong R, Jiang Z, Shi J and Li J: Antidiabetic potential of
flavonoids from traditional Chinese medicine: A review. Am J Chin
Med. 47:933–957. 2019.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Wang ZY, Liu JG, Li H and Yang HM:
Pharmacological effects of active components of chinese herbal
medicine in the treatment of Alzheimer's disease: A review. Am J
Chin Med. 44:1525–1541. 2016.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Akanda MR, Uddin MN, Kim IS, Ahn D, Tae HJ
and Park BY: The biological and pharmacological roles of polyphenol
flavonoid tilianin. Eur J Pharmacol. 842:291–297. 2019.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Song WY, Song YS, Ryu HW, Oh SR, Hong J
and Yoon DY: Tilianin inhibits MUC5AC expression mediated via
down-regulation of EGFR-MEK-ERK-Sp1 signaling pathway in NCI-H292
human airway cells. J Microbiol Biotechnol. 27:49–56.
2017.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Jiang H, Fang J, Xing J, Wang L, Wang Q,
Wang Y, Li Z and Liu R: Tilianin mediates neuroprotection against
ischemic injury by attenuating CaMKII-dependent
mitochondrion-mediated apoptosis and MAPK/NF-κB signaling. Life
Sci. 216:233–245. 2019.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408.
2001.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Cacabelos R: Parkinson's disease: From
pathogenesis to pharmacogenomics. Int J Mol Sci.
18(551)2017.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Wakabayashi K, Tanji K, Odagiri S, Miki Y,
Mori F and Takahashi H: The Lewy body in Parkinson's disease and
related neurodegenerative disorders. Mol Neurobiol. 47:495–508.
2013.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Cao M, Wu Y, Ashrafi G, McCartney AJ,
Wheeler H, Bushong EA, Boassa D, Ellisman MH, Ryan TA and De
Camilli P: Parkinson Sac domain mutation in synaptojanin 1 impairs
clathrin uncoating at synapses and triggers dystrophic changes in
dopaminergic axons. Neuron. 93:882–896 e5. 2017.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Kim CY and Alcalay RN: Genetic forms of
Parkinson's disease. Semin Neurol. 37:135–146. 2017.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Bellou V, Belbasis L, Tzoulaki I,
Evangelou E and Ioannidis JP: Environmental risk factors and
Parkinson's disease: An umbrella review of meta-analyses.
Parkinsonism Relat Disord. 23:1–9. 2016.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Subramaniam SR and Chesselet MF:
Mitochondrial dysfunction and oxidative stress in Parkinson's
disease. Prog Neurobiol. 106-107:17–32. 2013.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Kamat PK, Kalani A, Rai S, Swarnkar S,
Tota S, Nath C and Tyagi N: Mechanism of oxidative stress and
synapse dysfunction in the pathogenesis of Alzheimer's disease:
Understanding the therapeutics strategies. Mol Neurobiol.
53:648–661. 2016.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Filomeni G, De Zio D and Cecconi F:
Oxidative stress and autophagy: The clash between damage and
metabolic needs. Cell Death Differ. 22:377–388. 2015.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Kim EK and Choi EJ: Pathological roles of
MAPK signaling pathways in human diseases. Biochim Biophys Acta.
1802:396–405. 2010.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Kyriakis JM and Avruch J: Mammalian MAPK
signal transduction pathways activated by stress and inflammation:
A 10-year update. Physiol Rev. 92:689–737. 2012.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Lu N and Malemud CJ: Extracellular
signal-regulated kinase: A regulator of cell growth, inflammation,
chondrocyte and bone cell receptor-mediated gene expression. Int J
Mol Sci. 20(3792)2019.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Schattauer SS, Bedini A, Summers F,
Reilly-Treat A, Andrews MM, Land BB and Chavkin C: Reactive oxygen
species (ROS) generation is stimulated by κ opioid receptor
activation through phosphorylated c-Jun N-terminal kinase and
inhibited by p38 mitogen-activated protein kinase (MAPK)
activation. J Biol Chem. 294:16884–16896. 2019.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Jiang H, Xing J, Fang J, Wang L, Wang Y,
Zeng L, Li Z and Liu R: Tilianin protects against
ischemia/reperfusion-induced myocardial injury through the
Inhibition of the Ca2+/calmodulin-dependent protein
kinase II-dependent apoptotic and inflammatory signaling pathways.
Biomed Res Int. 2020(5939715)2020.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Jiang H, Ashraf GM, Liu M, Zhao K, Wang Y,
Wang L, Xing J, Alghamdi BS, Li Z and Liu R: Tilianin ameliorates
cognitive dysfunction and neuronal damage in rats with vascular
dementia via p-CaMKII/ERK/CREB and ox-CaMKII-dependent MAPK/NF-κB
pathways. Oxid Med Cell Longev. 2021(6673967)2021.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Jiang H, Zeng L, Dong X, Guo S, Xing J, Li
Z and Liu R: Tilianin extracted from Dracocephalum moldavica
L. induces intrinsic apoptosis and drives inflammatory
microenvironment response on pharyngeal squamous carcinoma cells
via regulating TLR4 signaling pathways. Front Pharmacol.
11(205)2020.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Mullin S and Schapira AH: Pathogenic
mechanisms of neurodegeneration in Parkinson disease. Neurol Clin.
33:1–17. 2015.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Nagatsu T and Nagatsu I: Tyrosine
hydroxylase (TH), its cofactor tetrahydrobiopterin (BH4), other
catecholamine-related enzymes, and their human genes in relation to
the drug and gene therapies of Parkinson's disease (PD): Historical
overview and future prospects. J Neural Transm (Vienna).
123:1255–1278. 2016.PubMed/NCBI View Article : Google Scholar
|
|
33
|
Zhu Y, Zhang J and Zeng Y: Overview of
tyrosine hydroxylase in Parkinson's disease. CNS Neurol Disord Drug
Targets. 11:350–358. 2012.PubMed/NCBI View Article : Google Scholar
|