Introduction
Diabetic kidney disease (DKD) (also known as
diabetic nephropathy) has been reported to develop in ~40% of
patients with diabetes worldwide and is the primary cause of
end-stage renal disease (1,2). The
morphological characteristics of DKD include excessive deposition
of extracellular matrix (ECM) with mesangial expansion, thickening
of glomerular and tubular basement membranes, podocyte effacement
and tubulointerstitial fibrosis (3). Mesangial cells (MCs) are the main
cellular constituents of the glomerular mesangium, accounting for
~35% of the total cells in the glomerulus (4). The main function of glomerular MCs
includes supporting structural architecture of the glomerular
capillary, maintaining mesangial matrix homeostasis and regulating
the filtration surface area (5,6).
Under hyperglycemia, these cells may undergo overproliferation with
increasing synthesis of ECM proteins, which leads to
glomerulosclerosis in the occurrence and development of DKD
(7). Therefore, an improved
understanding of the role of MCs in the pathogenesis of DKD may
contribute to the development of novel therapeutic drugs for the
control of this disease.
Caveolae are small glycosphingolipid- and
cholesterol-enriched omega-shaped invaginations of the plasma
membrane (50-100 nm), which have a wide range of functions, such as
endocytosis, transcytosis, and regulation of cell differentiation,
proliferation and apoptosis (8,9).
Caveolin-1 (CAV-1) is an integral membrane protein required for
caveolae formation, and previous studies have suggested its
important role in the pathogenesis of DKD (10,11).
It has been confirmed that CAV-1 can promote the synthesis and
accumulation of the ECM in MCs under hyperglycemia by activating
several profibrotic pathways (12). In addition, CAV-1 has been shown to
be involved in the upregulation of NADPH oxidase (NOX) activity and
production of superoxide in response to high glucose (HG) in MCs
(13). Thus, increasing evidence
has indicated a pathogenic role of CAV-1/caveolae in DKD, which may
become a novel and potent therapeutic target for this disease.
Proanthocyanidin is widely distributed in plants and
is the active ingredient in grape seed, green and black tea,
wolfberry and fruit juices (14).
It has been reported that proanthocyanidin has biological
activities, including free radical-scavenging, antioxidant,
anti-inflammatory and antitumor properties (14,15).
Procyanidin B2 (PB2), the main component of proanthocyanidin, is
composed of two molecules of flavan-3-ol (-)-epicatechin linked by
a C4-C8 bond. PB2 has been reported to prevent reactive oxygen
species (ROS) generation and inhibit inflammation (16). In addition, previous studies have
indicated the protective effect of PB2 on DKD (17,18).
However, little is currently known about the molecular mechanism
underlying the protective effect of PB2 on DKD. The present study
aimed to investigate the cytoprotective effects of PB2 against cell
proliferation, oxidative stress, ECM accumulation and inflammation
in renal MCs, and explored the molecular mechanism associated with
such effects.
Materials and methods
Chemicals and reagents
PB2 (purity>99%), NSC23766, VAS2870 and LY2109761
were purchased from MedChemExpress. Chemicals, such as MTT and
DMSO, were obtained from MilliporeSigma. The following antibodies
were obtained from Abcam: Phosphorylated (p)-VAV2 (cat. no.
ab86695), VAV2 (cat. no. ab52640), RAC1 (cat. no. ab155938), NOX4
(cat. no. ab133303), collagen IV (cat. no. ab6586), fibronectin
(cat. no. ab268020), TGF-β1 (cat. no. ab215715), p-SMAD2/3 (cat.
no. ab272332), SMAD2/3 (cat. no. ab217553), CAV-1 (cat. no.
ab32577), p-CAV-1 (cat. no. ab75876), NF-kB p65 (cat. no. ab16502),
goat anti-rabbit IgG H&L (HRP) (cat. no. ab7090) and goat
anti-mouse IgG H&L (HRP) (cat. no. ab205719), goat anti-rabbit
IgG H&L (Alexa Fluor 647) (cat. no. ab150079) and GAPDH (cat.
no. sc-365062) was obtained from Santa Cruz Biotechnology, Inc. The
RAC1 activity assay kit (cat. no. STA-401-1) was obtained from Cell
Biolabs, Inc. Other chemicals and reagents used in the present
study, unless otherwise specified, were obtained from Beyotime
Institute of Biotechnology or Sangon Biotech Co., Ltd.
Cell culture and treatments
The mouse mesangial SV40-Mes13 (Mes13) cells were
purchased from the American Type Culture Collection. Cells were
cultured in Dulbecco's modified Eagle medium (Gibco; Thermo Fisher
Scientific, Inc.) supplemented with 10% (v/v) heat-inactivated
fetal bovine serum (Gibco; Thermo Fisher Scientific, Inc.) at 37˚C
in a humidified atmosphere (95% air, 5% CO2). The
concentration of D-glucose to induce hyperglycemia (at 37˚C for 12
h) was chosen according to previous studies (19,20),
the concentration of PB2 (at 37˚C for 12 h) was chosen according to
another previous study (17), and
the untreated cells were chosen as a control. For evaluating the
involvement of redoxosomes in the effect of PB2, cells were treated
with RAC1 inhibitor (80 µM) or NOX inhibitor (10 µM) under HG
conditions (25 mM) for 12 h at 37˚C, and ROS or
H2O2 generation was assessed by ROS or
H2O2 assay kits. For evaluating the
involvement of TGF-β1/SMAD, cells were treated with TGF-β1/SMAD
inhibitor (LY2109761, 10 µM) under HG conditions (25 mM) for 12 h
at 37˚C. The expression levels of collagen IV and fibronectin were
assessed by western blot analysis.
Cell transfection
The negative control pcDNA3.1-vector was purchased
from Invitrogen; Thermo Fisher Scientific, Inc. and the CAV-1
expression vector pcDNA3.1-CAV-1 was purchased from Shanghai
GenePharma Co., Ltd. Cells were seeded in 96-well plates
(1x104 cells/well) or 6-well plates (1x105
cells/well), and transfected with empty or expression vector (1
µg/ml) using Lipofectamine® 2000 reagent (Invitrogen;
Thermo Fisher Scientific, Inc.) at 37˚C according to the
manufacturer's instructions. After 48 h, western blot analysis was
performed to determine the efficiency of transfection. In addition,
cells underwent subsequent experiments 48 h post-transfection.
Cell proliferation analysis
Cell proliferation was determined using the MTT
assay. Cells (1x104/well) were incubated with 0.5 mg/ml
MTT solution (100 µl) for 3 h at 37˚C. The cultured medium was then
replaced with DMSO solution (100 µl) and the absorbance was
recorded at a wavelength of 490 nm (DTX800; Beckman Coulter,
Inc.).
Oxidative marker analysis
Intracellular ROS generation was assessed using a
ROS assay kit (Beijing Solarbio Science & Technology Co., Ltd.)
with fluorescent probe 2',7'-dichlorofluorescein diacetate
(DCFH-DA). Briefly, cells (1x104/well) with or without
treatment were incubated with DCFH-DA (10 µM) for 15 min at 37˚C.
ROS concentration was analyzed by measuring fluorescence using a
fluorescence microplate reader (SpectraMax M5; Molecular Devices,
LLC) with excitation and emission wavelengths at 488 and 525 nm,
respectively. Intracellular H2O2 levels were
measured using a hydrogen peroxide/peroxidase assay kit (cat. no.
BC3595; Beijing Solarbio Science & Technology Co., Ltd.)
according to the manufacturer's protocol, with excitation at 530 nm
and detection at 590 nm, using a fluorescence microplate reader
(SpectraMax M5; Molecular Devices, LLC).
Rac1 activation assay
RAC1 activity was assessed using RAC1 activation
assay kit (cat. no. STA-401-1; Cell Biolabs, Inc.) according to the
manufacturer's protocol. Briefly, cells after treatment were lysed
by RIPA lysis buffer and PAK PBD agarose beads were used to isolate
and pull-down the active form of RAC1 (RAC1-GTP). Subsequently, the
precipitated RAC1-GTP was detected by western bot analysis using
RAC1 antibody.
Western blot analysis
Total protein was extracted from Mes13 cells by RIPA
lysis buffer (Beyotime Institute of Biotechnology) and quantitively
determined using BCA Protein Assay kit (Beyotime Institute of
Biotechnology). Proteins (50 µg/lane) were loaded and separated by
SDS-PAGE on 10% gels and were then transferred onto polyvinylidene
difluoride membranes. Membranes were blocked with non-fat milk (5%
in 0.1% Tween20-PBS buffer) for 1 h at room temperature and
incubated with primary antibodies (1:1,000) overnight at 4˚C.
Subsequently, membranes were incubated with HRP-conjugated
secondary antibodies (1:500) for 1 h at room temperature. The bands
were detected using Immobilon Western Chemiluminescent HRP
Substrate (Beyotime Institute of Biotechnology) and band
intensities were semi-quantified using ImageJ 1.48 (National
Institutes of Health).
Enzymelinked immunosorbent assay
(ELISA)
The cell supernatant was collected after the
indicated treatment (cells with or without CAV-1 transfection were
treated with PB2 (25 µM) under HG conditions (25 mM) for 12 h) and
processed for ELISA analysis. Culture supernatant (100 µl) was
analyzed using the ELISA kits (IL-1β and TNF-α) (Beijing Solarbio
Science & Technology Co., Ltd.) according to the manufacturers'
protocols. The absorbance was measured at 450 nm using a microplate
reader (Molecular Devices, LLC).
Immunofluorescence analysis
Cells were fixed in 2% paraformaldehyde for 20 min
at room temperature and permeabilized in 0.1% Triton X-100 for 20
min at room temperature, followed by incubation with primary
antibody (NF-κB p65; 1:500) overnight at 4˚C and
fluorescent-conjugated secondary antibody [goat anti-rabbit IgG
H&L (Alexa Fluor 647); 1:250] for 1 h at room temperature.
Fluorescence was observed with a fluorescence microscope (Leica
Microsystems GmbH). DAPI (0.5 µg/ml; 20 min at room temperature)
was used to stain the nuclei.
Statistical analysis
Statistical analysis was performed using SPSS 16.0
software package (SPSS, Inc.). Data are presented as the mean ± SD
of three independent experiments. Statistical comparisons were made
by one-way ANOVA followed by Tukey's post hoc test among multiple
groups. P<0.05 was considered to indicate a statistically
significant difference.
Results
PB2 protects MCs against HG-induced
cell proliferation and oxidative stress
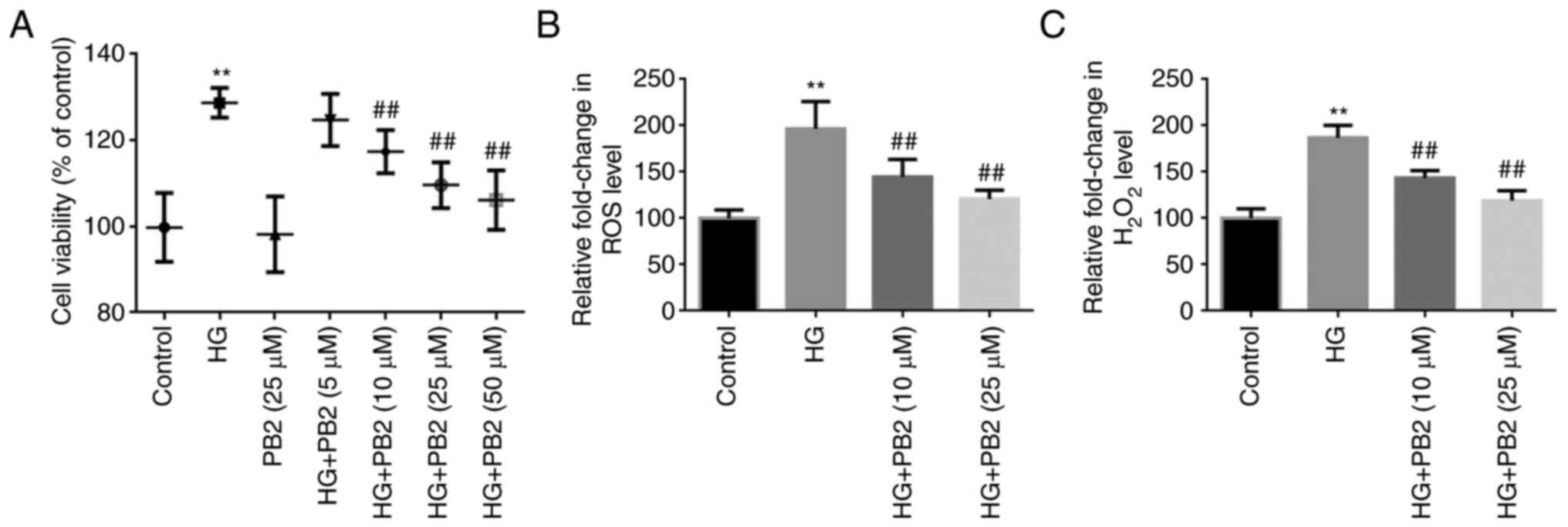
To investigate the effect of PB2 on HG-induced MC
dysfunction, Mes13 cells were treated with PB2 (5, 10, 25 and 50
µM) under HG conditions (25 mM). In the preliminary study,
D-glucose (10, 25 and 30 mM) treatment at 37˚C for 12 h
significantly induced rapid cell proliferation in Mes13 cells, as
determined using MTT assay, whereas there was no significant
difference between 25 and 30 mM concentrations (data not shown).
Therefore, the concentration of 25 mM D-glucose was chosen for
subsequent experiments. In this study, cell proliferation was
determined using the MTT assay, and oxidative stress was assessed
using ROS and H2O2 analyses. As shown in
Fig. 1, HG treatment significantly
increased cell proliferation, and elevated ROS and
H2O2 generation compared with the control
group. However, PB2 treatment (10, 25 and 50 µM) treatment exerted
protective effects on Mes13 cells under HG treatment, as determined
by MTT assay, but there was no significant difference between the
25 and 50 µM concentrations. Therefore, the concentrations of 10
and 25 µM PB2 were chosen for assessing anti-oxidative effect. The
results showed that PB2 treatment (10 and 25) dose-dependently
attenuated elevated ROS and H2O2 generation
induced by HG treatment.
PB2 protects MCs against oxidative
stress by regulating redoxosomes signaling
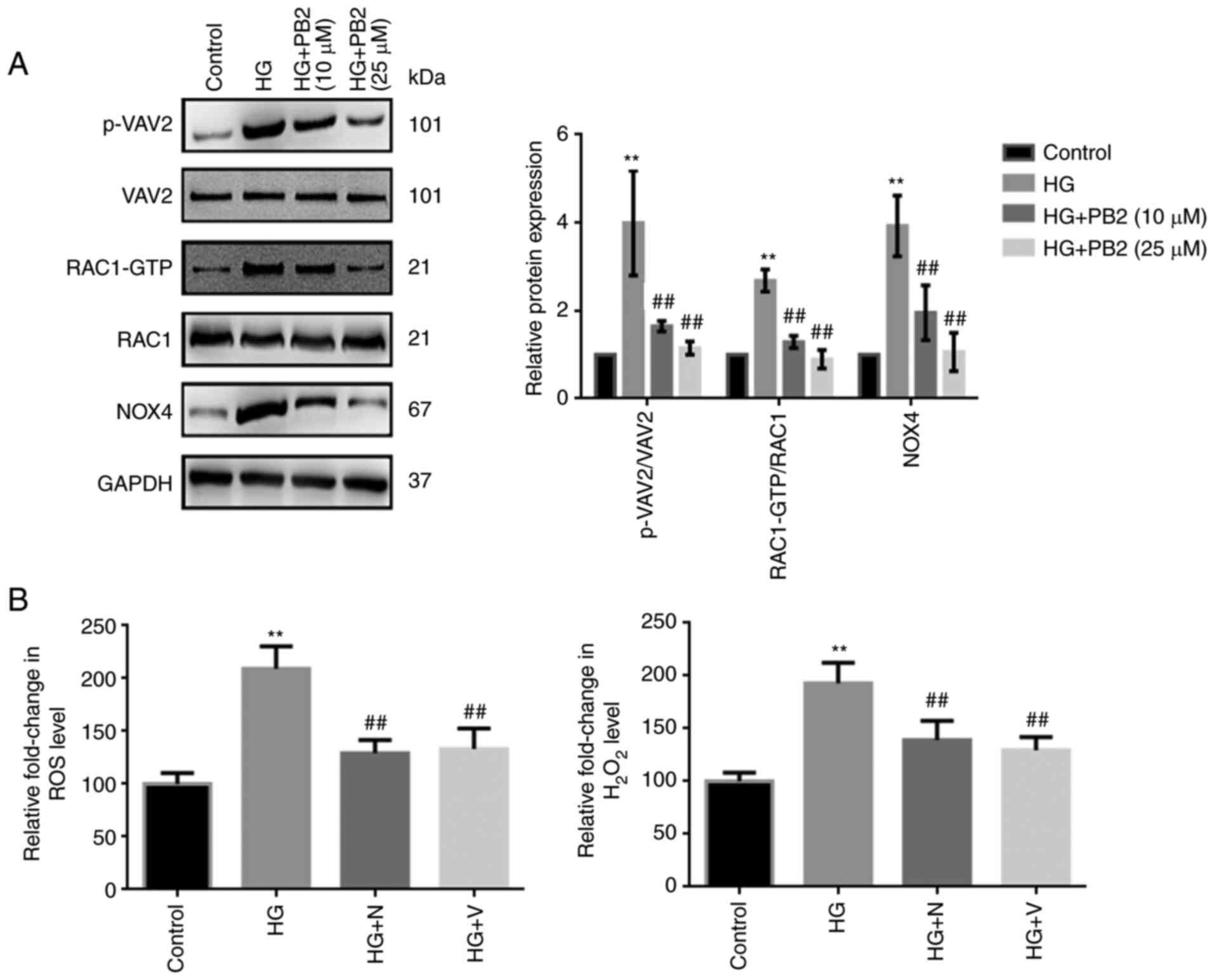
To investigate the molecular mechanism underlying
the cytoprotective effect of PB2 against oxidative stress in MCs,
Mes13 cells were treated with PB2 (10 and 25 µM) under HG
conditions (25 mM) and the expression levels of redoxosome-related
proteins were assessed. As shown in Fig. 2A, HG treatment induced the
upregulated expression of redoxosome-related proteins
[RAC1/VAV2/NOX4] in cells, which may subsequently enhance ROS
levels. However, PB2 treatment significantly attenuated such
changes in Mes13 cells under HG conditions, which was similar to
the effects of RAC1 inhibitor (NSC23766) and NOX inhibitor
(VAS2870) (Fig. 2B). However, the
effects of RAC1 inhibitor (NSC23766) or NOX inhibitor (VAS2870) on
the expression of redoxosome-related proteins were not
assessed.
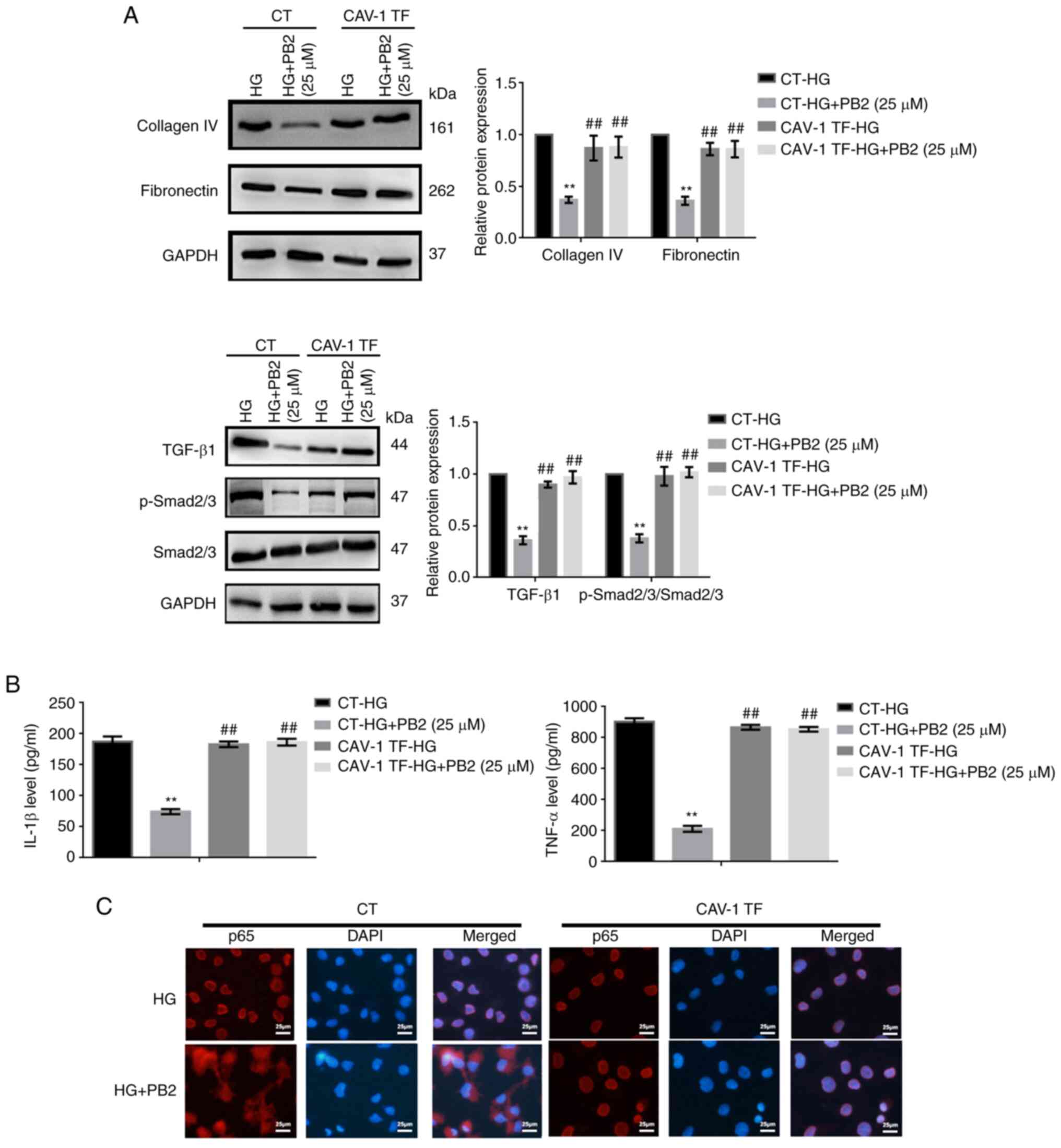
PB2 protects MCs against HG-induced
ECM accumulation by regulating TGF-β1/SMAD signaling
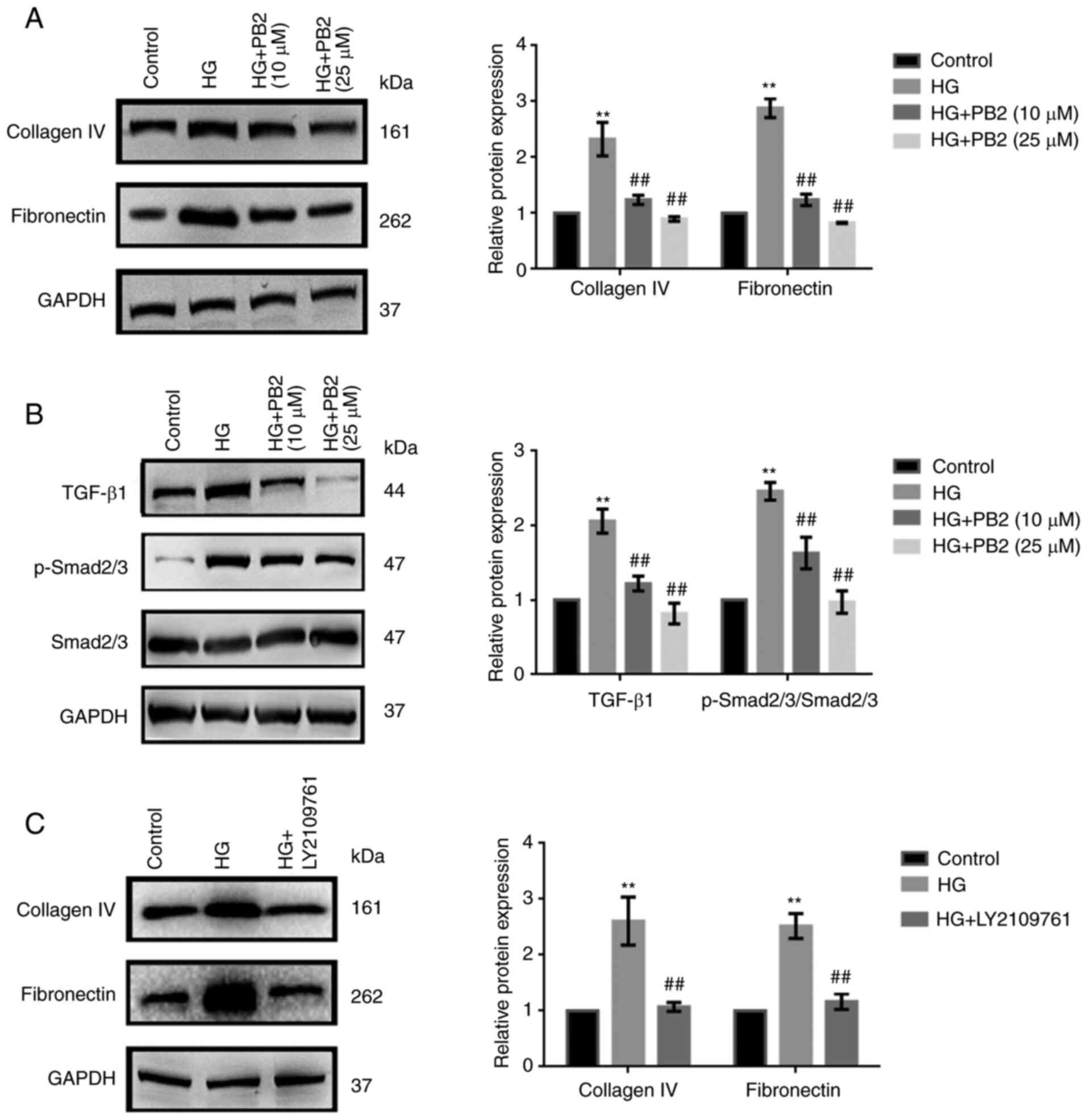
To investigate the effect of PB2 on HG-induced ECM
accumulation in MCs, Mes13 cells were treated with PB2 (10 and 25
µM) under HG conditions (25 mM). ECM accumulation is a
characteristic cellular response to HG (5). The expression of two typical ECM
markers (collagen IV and fibronectin), as well as changes in
TGF-β1/SMAD signaling were assessed. As shown in Fig. 3A and B, HG increased the expression levels of
ECM proteins and activation of TGF-β1/SMAD signaling; however, PB2
treatment markedly attenuated ECM accumulation and inactivated
TGF-β1/SMAD signaling in Mes13 cells under HG conditions. PB2 had
similar effects to the TGF-β1/SMAD signaling inhibitor (LY2109761)
with regard to ECM accumulation (Fig.
3C).
PB2 protects MCs against HG-induced
inflammation by regulating NF-κB signaling
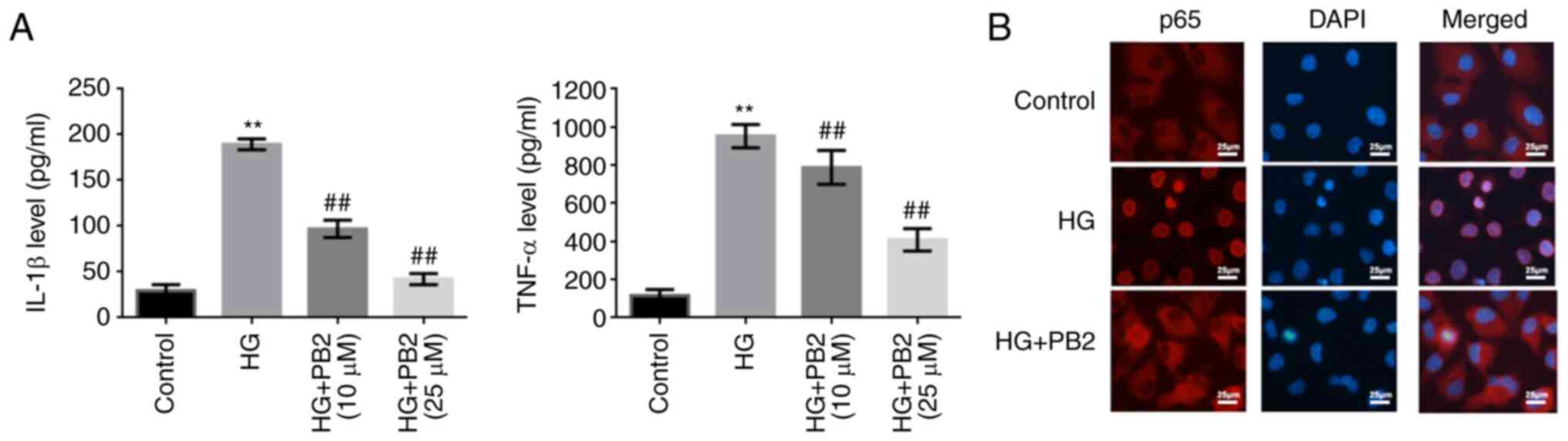
To investigate the effect of PB2 on HG-induced
inflammation in MCs, Mes13 cells were treated with PB2 (10 and 25
µM) under HG conditions (25 mM). The levels of two typical
cytokines (IL-1β and TNF-α) and the activation of NF-κB were
assessed. As shown in Fig. 4, HG
induced increased secretion of TNF-α and IL-1β, and activation of
NF-κB signaling [translocation of p65 (NF-κB) from the cytoplasm to
the nucleus]. However, PB2 treatment markedly reversed these
phenotypes in Mes13 cells under HG conditions.
CAV-1 overexpression reverses the
protective effect of PB2 on HG-induced MC dysfunction
CAV-1 is known to affect cellular redox homeostasis
and inflammation in several cell lines. To investigate the key role
of CAV-1 in the cytoprotective effect of PB2 on MCs, the expression
and activation of CAV-1 were assessed. HG treatment enhanced CAV-1
expression levels and the phosphorylation of CAV-1 (Y14) in Mes13
cells (Fig. 5A). To further assess
the role of CAV-1, Mes13 cells overexpressing CAV-1 were treated
with PB2 (10 and 25 µM) under HG conditions (25 mM). CAV-1
overexpression could reverse the effects of PB2 on cell
proliferation, oxidative stress, ECM accumulation and inflammation
and the related signaling pathways (Figs. 5 and 6). The present data indicated that PB2
protected MCs against hyperglycemia by inactivating redoxosomes,
and TGF-β1/SMAD and NF-κB signaling pathways in a CAV-1 dependent
manner.
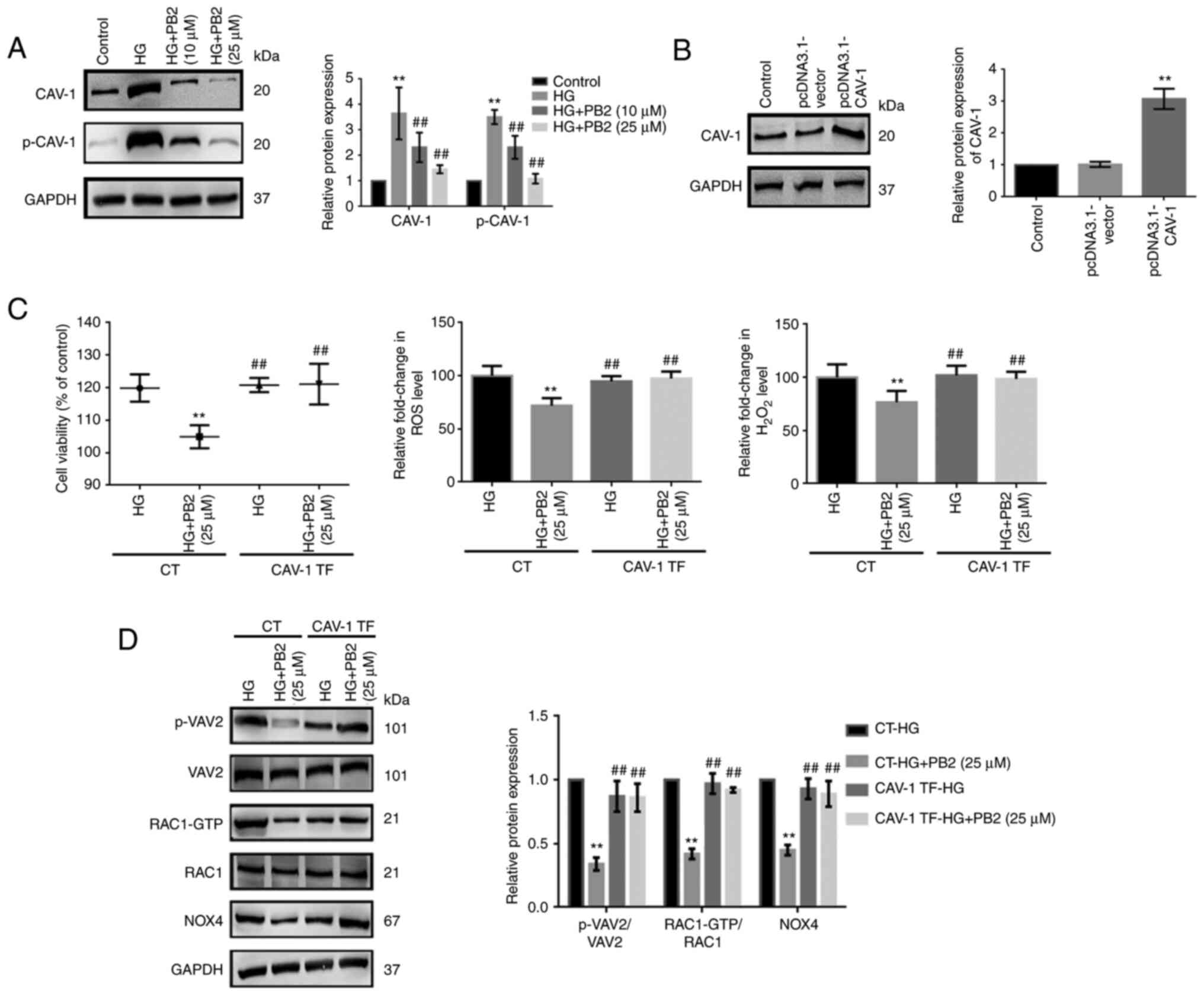 | Figure 5Effect of CAV-1 overexpression on the
cytoprotective effect of PB2 against oxidative stress in Mes13
cells. (A) Cells were treated with PB2 (10 and 25 µM) under HG
conditions (25 mM) for 12 h, and the expression levels of CAV-1 and
p-CAV-1 were assessed by western blot analysis and statistically
analyzed. **P<0.01 vs. control,
##P<0.01 vs. HG. (B) Cells were transfected with
pcDNA3.1-vector (negative control) or pcDNA3.1-CAV-1 for 48 h, and
the expression levels of CAV-1 were assessed by western blot
analysis and statistically analyzed. **P<0.01 vs.
control. (C) Cells with or without pcDNA3.1-CAV-1 transfection were
treated with PB2 (25 µM) under HG conditions (25 mM) for 12 h. Cell
proliferation was assessed by MTT assay. ROS generation was
assessed using a ROS assay kit and H2O2
generation was assessed using a H2O2
detection kit, and the results were statistically analyzed.
**P<0.01 vs. CT-HG; ##P<0.01 vs. CT-HG
+ PB2 (25 µM). (D) Cells transfected with or without pcDNA3.1-CAV-1
were treated with PB2 (25 µM) under HG conditions (25 mM) for 12 h.
The expression levels of redoxosome-related proteins were assessed
by western blot analysis and statistically analyzed.
**P<0.01 vs. CT-HG; ##P<0.01 vs. CT-HG
+ PB2 (25 µM). All data are presented as the mean ± SD of three
independent experiments. PB2, procyanidin B2; HG, high glucose;
CAV-1, caveolin-1; p-, phosphorylated; NOX4, NADPH oxidase 4; CT,
control; TF, CAV-1 transfection. |
Discussion
PB2 has been reported to exert a variety of
pharmacological effects on diabetic complications due to its
antioxidative and anti-inflammatory activities (21,22).
The renal protective effect of PB2 has been suggested in previous
studies on animal models of diabetic nephropathy. Zhang et
al (23) indicated PB2 as a
prospective therapy, which could suppress MFG-E8, along with
ERK1/2, Akt and GSK-3β signaling pathways. Li et al
(24) reported that PB2 inhibited
HG-induced epithelial-mesenchymal transition through the
downregulation of TGF-β/SMAD and mitogen-activated protein
kinase/P38 signaling pathways in renal tubular epithelial cells.
Bao et al (17) revealed
that PB2 markedly ameliorated mitochondrial dysfunction and
inhibited apoptosis in HG-treated rat MCs by activating the
AMPK/SIRT1/PGC-1α axis. To the best of our knowledge, the present
study was the first to reveal that PB2 exerted protective effects
against hyperglycemia-induced MC dysfunction by attenuating cell
proliferation, oxidative stress, ECM accumulation and
inflammation.
Previous studies have revealed that CAV-1, in
response to stimuli, can trigger MCs to secrete cytokines and
produce ECM proteins (25,26). In DKD, CAV-1 has been reported to
facilitate profibrotic signal transduction in response to several
stimuli, including hyperglycemia and angiotensin II (27,28).
The present study demonstrated that hyperglycemia upregulated and
activated CAV-1 (CAV-1 phosphorylation) in MCs, which subsequently
led to marked MC dysfunction. It is important to understand the
role of CAV-1 in MC dysfunction under hyperglycemia. HG levels are
known to stimulate redox signaling in renal MCs, which can lead to
diabetic glomerular changes (29).
Redoxosomes have recently been found to be a fledgling area of
cellular signaling through superoxide-producing endosomes, which
act via specific redox modifications on numerous proteins and enzymes
(30,31). In the present study, using CAV-1
transfection experiments, it was revealed that redoxosomes were the
downstream target of CAV-1, which control ROS generation and NF-κB
activation. However, PB2 treatment significantly suppressed
oxidative stress and inflammation induced by hyperglycemia, which
may be via inhibiting CAV-1-dependent redoxosomes activation. These
results indicated that CAV-1 was involved in redoxosomes signaling
and may be a novel therapeutic target of DKD. The TGF-β1/SMAD
signaling has an important role in renal fibrosis, the activation
of which is crucial to the formation of the ECM (32,33).
The present results indicated that CAV-1 could also activate
TGF-β1/SMAD signaling, which may result in ECM accumulation in MCs
under hyperglycemia. In addition, CAV-1 overexpression
significantly reversed the effects of PB2 on TGF-β1/SMAD signaling
and consequently ECM accumulation in MCs. Thus, CAV-1-dependent
TGF-β1/SMAD signaling may also be a new therapeutic target of DKD
(Fig. 7).
In conclusion, PB2 regulated the proliferation,
oxidative stress, ECM accumulation and inflammation of glomerular
MCs under hyperglycemia potentially by suppressing CAV-1-dependent
signaling pathways. The present study suggested the potential
clinical application of PB2 in treating DKD and indicated that
CAV-1-related signaling pathways may be a potential therapeutic
target of this disease.
Acknowledgements
Not applicable.
Funding
Funding: The present study was supported by the Project of Wuxi
Health Commission (grant nos. Z202009, Z202014 and Z202110), the
Key Project of Medical and Health Science and Technology Plan of
Suzhou New District (grant no. 2019Z005) and the Science and
Technology Development Project of Suzhou (grant no.
SYSD2020087).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
GL, DJ and FZ designed the experiments. JY, KW, XZ
and JQ performed the experiments. JY, KW, XZ and JQ confirm the
authenticity of all the raw data. JY and XZ analyzed the
experimental results. JY and KW wrote the manuscript. All authors
read and approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Alicic RZ, Rooney MT and Tuttle KR:
Diabetic kidney disease: Challenges, progress, and possibilities.
Clin J Am Soc Nephrol. 12:2032–2045. 2017.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Lin YC, Chang YH, Yang SY, Wu KD and Chu
TS: Update of pathophysiology and management of diabetic kidney
disease. J Formos Med Assoc. 117:662–675. 2018.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Anders HJ, Huber TB, Isermann B and
Schiffer M: CKD in diabetes: Diabetic kidney disease versus
nondiabetic kidney disease. Nat Rev Nephrol. 14:361–377.
2018.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Zhao JH: Mesangial cells and renal
fibrosis. Adv Exp Med Biol. 1165:165–194. 2019.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Dong Z, Sun Y, Wei G, Li S and Zhao Z:
Ergosterol ameliorates diabetic nephropathy by attenuating
mesangial cell proliferation and extracellular matrix deposition
via the TGF-β1/Smad2 signaling pathway. Nutrients.
11(483)2019.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Marciano DK: Mesangial cells: The tuft
guys of glomerular development. J Am Soc Nephrol. 30:1551–1553.
2019.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Tung CW, Hsu YC, Shih YH, Chang PJ and Lin
CL: Glomerular mesangial cell and podocyte injuries in diabetic
nephropathy. Nephrology (Carlton). 23 (Suppl 4):S32–S37.
2018.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Parton RG, Tillu VA and Collins BM:
Caveolae. Curr Biol. 28:R402–R405. 2018.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Parton RG, McMahon KA and Wu Y: Caveolae:
Formation, dynamics, and function. Curr Opin Cell Biol. 65:8–16.
2020.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Wang S, Wang N, Zheng Y, Zhang J, Zhang F
and Wang Z: Caveolin-1: An oxidative stress-related target for
cancer prevention. Oxid Med Cell Longev.
2017(7454031)2017.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Peng F, Wu D, Ingram AJ, Zhang B, Gao B
and Krepinsky JC: RhoA activation in mesangial cells by mechanical
strain depends on caveolae and caveolin-1 interaction. J Am Soc
Nephrol. 18:189–198. 2007.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Mehta N, Zhang D, Li R, Wang T, Gava A,
Parthasarathy P, Gao B and Krepinsky JC: Caveolin-1 regulation of
Sp1 controls production of the antifibrotic protein follistatin in
kidney mesangial cells. Cell Commun Signal. 17(37)2019.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Sun LN, Liu XC, Chen XJ, Guan GJ and Liu
G: Curcumin attenuates high glucose-induced podocyte apoptosis by
regulating functional connections between caveolin-1
phosphorylation and ROS. Acta Pharmacol Sin. 37:645–655.
2016.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Rauf A, Imran M, Abu-Izneid T,
Iahtisham-Ul-Haq Patel S, Pan X, Naz S, Sanches Silva A, Saeed F
and Rasul Suleria HA: Proanthocyanidins: A comprehensive review.
Biomed Pharmacother. 116(108999)2019.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Luca SV, Macovei I, Bujor A, Miron A,
Skalicka-Woźniak K, Aprotosoaie AC and Trifan A: Bioactivity of
dietary polyphenols: The role of metabolites. Crit Rev Food Sci
Nutr. 60:626–659. 2020.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Su H, Li Y, Hu D, Xie L, Ke H, Zheng X and
Chen W: Procyanidin B2 ameliorates free fatty acids-induced hepatic
steatosis through regulating TFEB-mediated lysosomal pathway and
redox state. Free Radic Biol Med. 126:269–286. 2018.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Bao L, Cai X, Zhang Z and Li Y: Grape seed
procyanidin B2 ameliorates mitochondrial dysfunction and inhibits
apoptosis via the AMP-activated protein kinase-silent mating type
information regulation 2 homologue 1-PPARγ co-activator-1α axis in
rat mesangial cells under high-dose glucosamine. Br J Nutr.
113:35–44. 2014.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Zhou Y, Li BY, Li XL, Wang YJ, Zhang Z,
Pei F, Wang QZ, Zhang J, Cai YW, Cheng M and Gao HQ: Restoration of
mimecan expression by grape seed procyanidin B2 through regulation
of nuclear factor-kappaB in mice with diabetic nephropathy. Iran J
Kidney Dis. 10:325–331. 2016.PubMed/NCBI
|
|
19
|
Breast Cancer Linkage Consortium. Cancer
risks in BRCA2 mutation carriers. J Natl Cancer Inst. 91:1310–1316.
1999.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Chen B, Li Y, Liu Y and Xu Z: circLRP6
regulates high glucose-induced proliferation, oxidative stress, ECM
accumulation, and inflammation in mesangial cells. J Cell Physiol.
234:21249–21259. 2019.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Fan J, Liu H, Wang J, Zeng J, Tan Y, Wang
Y, Yu X, Li W, Wang P, Yang Z and Dai X: Procyanidin B2 improves
endothelial progenitor cell function and promotes wound healing in
diabetic mice via activating Nrf2. J Cell Mol Med. 25:652–665.
2021.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Yin M, Zhang P, Yu F, Zhang Z, Cai Q, Lu
W, Li B, Qin W, Cheng M, Wang H and Gao H: Grape seed procyanidin
B2 ameliorates hepatic lipid metabolism disorders in db/db mice.
Mol Med Rep. 16:2844–2850. 2017.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Zhang Z, Li BY, Li XL, Cheng M, Yu F, Lu
WD, Cai Q, Wang JF, Zhou RH, Gao HQ and Shen L: Proteomic analysis
of kidney and protective effects of grape seed procyanidin B2 in
db/db mice indicate MFG-E8 as a key molecule in the development of
diabetic nephropathy. Biochim Biophys Acta. 1832:805–816.
2013.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Li D, Zhao T, Meng J, Jing Y, Jia F and He
P: Procyanidin B2 inhibits high glucose-induced
epithelial-mesenchymal transition in HK-2 human renal proximal
tubular epithelial cells. Mol Med Rep. 12:8148–8154.
2015.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Adebiyi A, Soni H, John TA and Yang F:
Lipid rafts are required for signal transduction by angiotensin II
receptor type 1 in neonatal glomerular mesangial cells. Exp Cell
Res. 324:92–104. 2014.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Guan TH, Chen G, Gao B, Janssen MR,
Uttarwar L, Ingram AJ and Krepinsky JC: Caveolin-1 deficiency
protects against mesangial matrix expansion in a mouse model of
type 1 diabetic nephropathy. Diabetologia. 56:2068–2077.
2013.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Moriyama T, Tsuruta Y, Shimizu A, Itabashi
M, Takei T, Horita S, Uchida K and Nitta K: The significance of
caveolae in the glomeruli in glomerular disease. J Clin Pathol.
64:504–509. 2011.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Li CD, Zhao JY, Chen JL, Lu JH, Zhang MB,
Huang Q, Cao YN, Jia GL, Tao YX, Li J and Cao H: Mechanism of the
JAK2/STAT3-CAV-1-NR2B signaling pathway in painful diabetic
neuropathy. Endocrine. 64:55–66. 2019.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Akaishi T, Abe M, Okuda H, Ishizawa K, Abe
T, Ishii T and Ito S: High glucose level and angiotensin II type 1
receptor stimulation synergistically amplify oxidative stress in
renal mesangial cells. Sci Rep. 9(5214)2019.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Spencer NY and Engelhardt JF: The basic
biology of redoxosomes in cytokine-mediated signal transduction and
implications for disease-specific therapies. Biochemistry.
53:1551–1564. 2014.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Oakley FD, Abbott D, Li Q and Engelhardt
JF: Signaling components of redox active endosomes: The
redoxosomes. Antioxid Redox Signal. 11:1313–1333. 2009.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Dong J, Ding L, Wang L, Yang Z, Wang Y,
Zang Y, Cao X and Tang L: Effects of bradykinin on proliferation,
apoptosis, and cycle of glomerular mesangial cells via the
TGF-β1/Smad signaling pathway. Turk J Biol. 45:17–25.
2021.PubMed/NCBI View Article : Google Scholar
|
|
33
|
Ma TT and Meng XM: TGF-β/Smad and renal
fibrosis. Adv Exp Med Biol. 1165:347–364. 2019.PubMed/NCBI View Article : Google Scholar
|