Introduction
Verbascoside (Verb) (1) is naturally occurring in many plants
such as Paulownia tomentosa, Buddleja brasiliensis, Lantana
camara, Olea europea, Striga asiatica, Lippia citriodora,
Lippia javanica. Verb is hydrophilic in nature and has various
pharmacological activities such as antimicrobial, antioxidant,
neuroprotective, anti-inflammatory, and wound healing effects along
with pro-apoptotic, anti-proliferative, and pro-differentiative,
chemopreventive-chemotherapeutic potential effects in cancer
(2-6).
Bcr/Abl protein is encoded by the Bcr/Abl fusion gene, which causes
chronic myeloid leukaemia (CML) due to uncontrolled tyrosine kinase
activity. Hematopoietic cell differentiation and proliferation are
destroyed by Bcr/Abl (7). It was
reported that differentiation of leukemic cells could induced by
Verb towards the macrophage-monocyte lineage (8). Verb has been proposed to be the
investigational drug for the treatment of leukaemia patients
(9,10). Tyrosine kinase inhibitor; Imatinib
mesylate (IM) targets the inhibition of Bcr/Abl and involved in the
stimulation of apoptosis in leukemic cells (11). Unfortunately, relapse and
resistance can be seen in some patients (12). IM resistance is mainly related to
Abl mutations (Val256Gly, p.Thr315Ile, p.Gly250Glu, p.Tyr253His,
p.Phe317Leu and p.Met351Thr) and amplification of the Bcr/Abl
oncogene, but, some patients lacking these mutations could be
unresponsive to IM treatment (13). Bcr/Abl independent resistance
mechanisms are still being investigated (14,15).
To overcome the resistance, 2nd generation Abl tyrosine kinase
inhibitors have been developed. Dasatinib (Das) is 325-fold more
potent against cells expressing wild-type Bcr/Abl compared to IM
(16). Utilization of elevated
concentrations of chemotherapeutic drugs to treat cancers might
possibly cause cytotoxic effects on the healthy cells. So, possible
usage of the least drug concentrations is the main aim in the
treatment of various cancers. Natural compounds with low toxicity
can influence the resistance mechanisms (17). Complete destruction of resistant
leukemic cells may require other therapeutic agents in combination
with Bcr/Abl inhibitors that modify the molecular pathways to
control cell survival. Only a few studies have investigated the
role of Verb in leukaemia. It remains unclear whether Verb
regulates the biological functions of leukaemia cells and overcomes
the resistance. Therapeutic applications targeting the inflammatory
environment might perturb CML cells (18). Tumour necrosis factor (TNFα) is a
cytokine that plays a role in inflammation, stress response, and
apoptosis and produced mainly by activated macrophages (19). In our study Lipopolysaccharide
(LPS) and TNFα was utilized to trigger inflammation in sensitive
(K562) and resistant K562 (R-K562) cells and the effect of Verb on
oxidative stress, apoptosis, and mitogen-activated protein kinase
(MAPK) signalling pathway was investigated. The abnormal activation
of MAPK signalling pathway by Bcr/Abl tyrosine kinase plays an
important role in the progression of leukaemia (20). Extracellular signalling molecules
such as cytokines, cellular stress, and growth factors could
activate the MAPK pathway. MAPK is involved in the regulation of
cell proliferation, differentiation, cell cycle, survival, and
induction of leukemogenesis. p38 MAP kinase, c-Jun N-terminal
kinase (JNK), extracellular signal regulated kinase (ERK) are three
main members of the MAPK pathway (21). As a biological material we used
rapidly proliferating human myelogenous leukaemia cell line K562.
In the present study we aimed to investigate the effects of Verb
alone and its concomitant use with IM, Das, LPS and TNFα on
apoptosis, oxidative stress and MAPK pathway in both IM-sensitive
and IM-resistant K562 CML cells in order to assess possible
interactions between inhibitors of tyrosine kinase and Verb.
Materials and methods
Cell cultures
K562 (CCL-243); human chronic myelogenous leukaemia
cell line were purchased from ATCC (American type culture
collection). Authentication of the cells was performed by the
American Type Culture Collection using short tandem repeat analysis
on the cells. Cells were cultured in Iscove's Modified Dulbecco's
Medium (Wisent Bioproducts) supplemented with 10% foetal bovine
serum (Gibco) and 1% of penicillin-streptomycin antibiotics (Bio
Ind). Cells were maintained under 5% CO2 at 37˚C
conditions. Increasing concentrations of IM were applied to K562
cells stepwise for 12 months between 0.1 µM-10 µM (22). Resistant cells were separated using
a Ficoll-Hypaque gradient, washed with IMDM, and kept in IMDM with
10% FBS and 10 µM IM. Subpopulation cells which were able to grow
in 10 µM IM, selected as resistant and referred as R-K562.
Cell treatment
For the determination of the cytotoxic effect of IM
and Das which are commonly used tyrosine kinase inhibitors,
‘high-dose pulse’ of IM (100 µM) and Das (2,500 nM) were chosen in
our experiments. It was reported that high-dose pulse therapy and
low-dose continuous therapy confer equivalent cytotoxicity to CML
cells. IM and Das concentration used in the high dose pulse therapy
experiments mimics peak serum levels achieved in patients, much
lower concentrations induce cytotoxicity in vitro with
prolonged exposure (23). García
et al determined the cytotoxic activity of essential oils of
Lippia alba (Verbenaceae Family) on chronic myelogenous
leukaemia cells-K562 using concentrations ranging from 45 to 200
µg/ml (24). Essential oils
displayed a cytotoxic effect on tumour cells IC50 was >45 µg/ml
in K562 cells. Cytotoxic activity assay of essential oils of
Lippia citriodora were performed in P815 murine mastocytoma
cells in 68.67, 34.33, 17.16, 8.58, 4.29, 2.14 µg/ml concentrations
(25). In our study cells were
treated with 50 µg/ml Verb and/or 100 µM IM, 2,500 nM Das, 100
ng/ml LPS (26), 10 ng/ml TNFα
(19) for a duration of 48 h at
37˚C.
Cell viability
Cell viability of untreated controls and
drug-treated cells were assessed by the WST-8 Cell Proliferation
Kit (Elabscience). 20x103 cells were seeded into 96-well
plates in 100 ml IMDM medium for each well. At the end of the
assay, absorbance of each sample was measured using a microplate
reader (Biotek).
Abl (Phospho-Tyr412) colorimetric
cell-based analysis
Tyr412 is necessary both for activity and for
regulation of Bcr/Abl, by stabilizing the inactive or the active
conformation of the enzyme in a phosphorylation-dependent manner
(27). The Colorimetric Cell-Based
ELISA Kit (Cat#EKC2643, Boster Biological Technology, USA) allows
for the detection of Abl (Phospho-Tyr412). The effects of Verb
alone and in combination with IM, Das, LPS and TNFa on target
protein expression determined in K562 and R-K562 cells. The Abl
protein is captured by target-specific primary (1˚) antibody while
the HRP-conjugated secondary (2˚) antibody binds to the Fc region
of the 1˚ antibody. Through this binding, the HRP enzyme conjugated
to the 2˚ antibody catalysed a colorimetric reaction upon substrate
addition. The phosphorylated target protein's OD values were
normalized using the non-phosphorylated target protein's OD values
(Anti-ABL2 (Phosphorylated) Antibody)/OD450 (Anti-ABL2 Antibody).
The measured OD450 readings were normalized to the OD595 values
obtained for crystal violet staining (OD450/OD595). Crystal violet
solution was provided as a ready-to-use solution in the kit.
Briefly, 50 µl crystal violet solution was added to each well and
was incubated for 30 min at room temperature on a shaker. Crystal
violet binds to cell nuclei and gives absorbance readings
proportional to cell counts at 595 nm. GAPDH specific monoclonal
antibody was used to serve as an internal positive control in
normalizing the target absorbance values.
Caspase-3 analysis
Caspase-3 enzyme activity was determined using a
colorimetric assay (Elabscience). The cells that had been treated
with IM, Das, LPS and TNFα and/or Verb for 48 h were collected by
centrifugation at 1,000 g for 10 min. Pelleted cells were treated
with 100 µl of cold lysis buffer to obtain the cell lysate. Then,
the cell lysates were incubated on ice for 10 min and were
centrifuged at 14,000 g for 1 min. Following the centrifugation,
supernatants were transferred to new microcentrifuge tubes. For
measuring caspase-3 enzyme activity, 100 µl of the samples were
added to the wells, after the incubation period 100 µl Biotinylated
Detection Ab working solution, 100 µl HRP conjugate working
solution, 90 µl Substrate Reagent and 50 µl Stop Solution applied
to each well respectively. Absorbances of the samples were read
under 450 nm wavelength of light via a microplate reader.
Spheroid formation
It was reported that tumour spheroid formation was
observed only in the resistant K562 cells, independently of the
absence or the presence of IM in the culture medium (28). 1x104 cells were plated
per well in a 6-well plate pre-coated with 1ml of agar-agar (Sigma)
in serum-free IMDM and incubated at 37˚C in 5% CO2 for
10 days. The colonies were photographed every 5 days (Olympus
CKX41).
Cell-based ELISA measuring MAPK
protein expression levels
Total/phosphorylated protein levels of ERK, p38 and
JNK were determined using cell-based ELISA (RayBio® Cell
Based Assay cat. no. CBEL-ERK-SK; RayBiotech Life, Inc.). These
assays can be used for determination of the relative amount of p38
MAPK (Thr180/Tyr182), JNK (Thr183/Tyr185) and ERK1/2
(Thr202/Tyr204) phosphorylation. 50,000 cells were seeded into a 96
well tissue culture plate coated by adding 100 µl poly-L-Lysine.
The cells were fixed after the incubation period. After blocking,
anti-phospho-protein specific antibody or anti-pan-protein specific
antibody was added into the wells and incubated. The wells were
washed, and HRP-conjugated anti-mouse IgG was added. TMB substrate
solution was added to the wells. Colour development was in
proportion to the amount of protein. The intensity of the colour
was measured at 450 nm.
Measurements of total oxidant status
(TOS)
Incubation of K562 and R-K562 cells with the IM,
Das, LPS and TNFα and/or Verb for 48 h, the cell culture media was
discarded. Colorimetric method was applied to the supernatants to
detect the TOS (29). In this
method, the ferrous ion O-dianisidine complex was oxidized to
ferric ion. The glycerol molecules which are abundant in the
reaction medium, enhance the oxidation reaction. A coloured complex
with xylenol orange appeared due to the ferric ion present in the
acidic medium. The colour intensity was assessed by the
spectrophotometry method is associated with the amount of total
oxidant molecules present in the sample. The calibrations were
carried out using hydrogen peroxide, and the obtained results were
explained as mmol H2O2 equiv./l.
Measurements of total antioxidant
status (TAS)
The Fenton reaction produced a hydroxyl radical,
which is the most powerful biological radical. The dianisyl
radical, which has a bright yellowish-brown colour, was also
obtained due to the hydroxyl radical's reaction with the colourless
substrate O-dianisidine (29). Upon the addition of a cell culture
medium sample, the oxidative reactions initiated by the hydroxyl
radicals present in the reaction mix are suppressed by the
antioxidant components of the sample, preventing the colour change
and thereby producing an effective way to measure the TAS level.
The assay results are expressed as mmol Trolox equiv./l. The
protein concentration of the samples obtained for the TAS-TOS
assays were determined by the Bradford method using BSA as a
standard.
Determination of oxidative stress
index (OSI)
The OSI was calculated in accordance with the
following formula:
OSI (arbitrary unit)=TOS (µmol
H2O2 equiv./l)/TAS (mmol Trolox equiv./l)
x10.
Comet assay
DNA damage and any genotoxic effects of IM, Das,
Verb, IM + Verb, Das + Verb were determined using Comet Assay IV
Version 4.3.2 for Basler FireWire in K562 cells. K562 cells were
incubated with the agents for 48 h. Then the cell mediums were
removed and cells were washed in 0.1 M PBS for three times,
separated, and then suspended in 0.1 M PBS (2x103 cells
in 75 µl PBS) with low melting agarose (LMA). The cells of all
groups were divided into three layers. The first layer contained 1%
normal melting agarose. The cells were suspended in a second layer
and 1% LMA was added to the solidified first layer. A third layer
containing 1% LMA was subsequently added. All these stages were
performed in the cold. After the solidification process, all slides
were kept in the lysing solution for 1 h at 4˚C. Next, the slides
were placed in the electrophoresis buffer for 20 min. Then, the
slides were electrophoresed at 25 V (300 mA, approx. 0.74 V/cm) for
30 min at 4˚C. The samples were washed 3 times with the
neutralization buffer for 5 min, and then the slides were immersed
in methanol for 5 min at -20˚C. To prevent exogenous DNA damage,
these processes were carried out in a dark medium. Then, the slides
were placed on a smooth area to dry. The slides were saturated with
ethidium bromide (60 µl) and then monitored the 590 nm emission and
510-560 nm excitation filters using a Nikon fluorescent microscope.
Images of 50 randomly chosen comets on each triplicate slide were
captured per sample using x20 magnification (30).
Statistical analysis
Data are expressed as mean ± standard deviation of
three independent determinations. An error probability with
P<0.05 was selected as significant. All experiments were
performed in triplicate and the mean value was used for analysis. A
one-way analysis of variance (ANOVA) was used to test significant
(P<0.05) differences between the groups (Control, IM, Das, LPS,
TNF and/or Verb) datasets, separately. When significant, the
differences were further tested with a post hoc Tukey-HSD test.
Shapiro-Wilk test was applied, prior to ANOVA routine, to test for
normality and homogeneity of variances, respectively. When those
conditions could not be met, a Kruskal-Wallis analysis and Dunn's
test were used.
Results
Cell viability assessment via WST8
cell proliferation assay
To analyse the effectiveness of Verb on cell
viability we used K562 and R-K562 cells. The cell viability of K562
control cells incubated with no agent was set as 100% and the cell
viability of the other groups was compared to the viability of the
control cells. In order to determine the % cell viability of K562
and R-K562 cells we have treated the cells with different
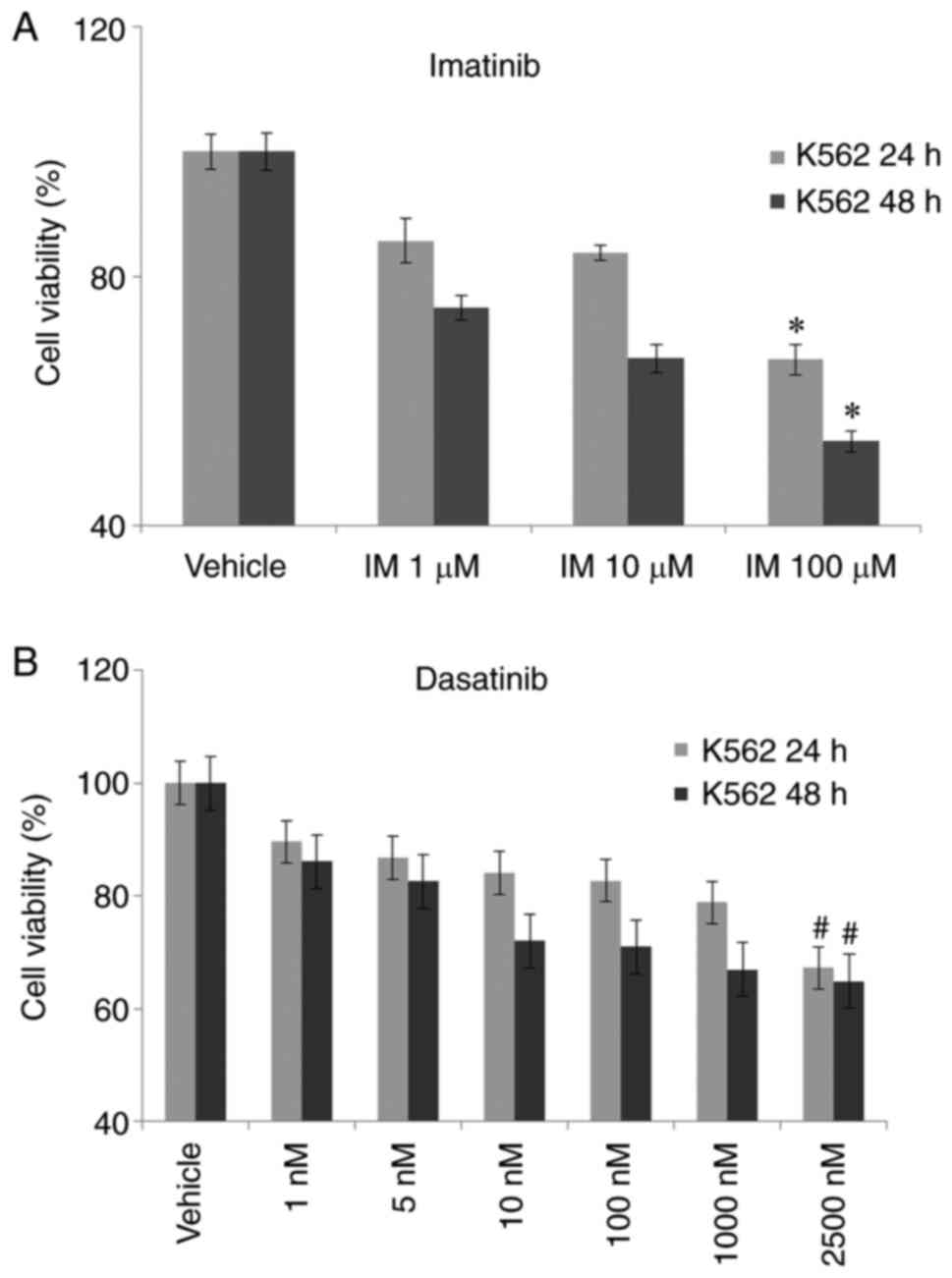
concentrations of IM (1-10-100 µM) for 24 and 48 h. We have
observed a significant change in cell viability in 100 µM IM dosage
after 24 and 48 h of exposure compared to untreated control cells
in K562 cells. IM had a significantly higher action on 48 h
incubation in K562 cells (Fig.
1A). Cell viability was observed as 67.2 and 65% after 2,500 nM
Das, incubation respectively in 24 and 48 h (Fig. 1B). 48 h incubation was selected for
further analysis. And the 48-h treatment was fixed for the
treatment of R-K562 cells. We incubated K562 cells with Verb alone
and in combination with IM, Das, LPS and TNFa for 48 and 72 h. Cell
viability was quantified using the WST8 assay. Relative amount of
cell viability of the Verb and/or IM, Das, LPS and TNFa in K562 and
R-K562 cells were shown in Fig. 2.
Incubation with 100 µM IM, 2,500 nM Das, 100 ng/ml LPS, 10 ng/ml
TNFα and 50 µg/ml Verb led to a significant decrease in cell
viability after 48 and 72 h. Concomitant incubation of Verb with
selected TKI drugs and inflammation inducers, LPS and TNFα, caused
a dramatic diminution in K562 and R-K562 cell viability. Cell
viability was observed as 51, 65, 52, 45% after IM, Das, LPS and
TNFα incubation respectively in 48 h (Fig. 2A). 72 h treatment resulted in 43,
52, 52, 35% cell viability after IM, Das, LPS and TNFa incubation,
respectively. Verb treatment was more effective when combined with
the IM, Das, LPS and TNFa on cell viability than its single use in
48 or 72 h timeline. 48 h treatment of Verb was seen as sufficient
for obtaining the inhibitory effect on cell viability combined with
the selected drugs. Hence, 48 h treatment was applied to the R-K562
cells for further experiments. Treatment with Verb alone or in
combination with IM, Das, LPS and TNFα resulted in reduced cell
viability after 48 h in R-K562 cells (Fig. 2B). Cell viability was observed as
51.5, 42.7, 52.8, 52.2% after IM, Das, LPS and TNFα incubation
respectively in 48 h treatment in R-K562 cells. Cell viability was
observed as 51.5% after IM incubation and 70.23% after Verb
incubation in R-K562 cells (Fig.
2C). Concomitant use of Verb with IM, led to a significant
decrease in cell viability (52.29%) compared to single treatment of
Verb. But the viability of R-K562 cells were not significantly
increased in the IM + Verb treatment compared to single IM
treatment. This observation could result from the increased
proliferation rate of R-K562 cells compared to their non-resistant
counterparts. But Verb shows its anti-leukemic effect by
downregulating of Abl, inhibiting of MAPK/ERK signalling and ROS
mediated DNA damage.
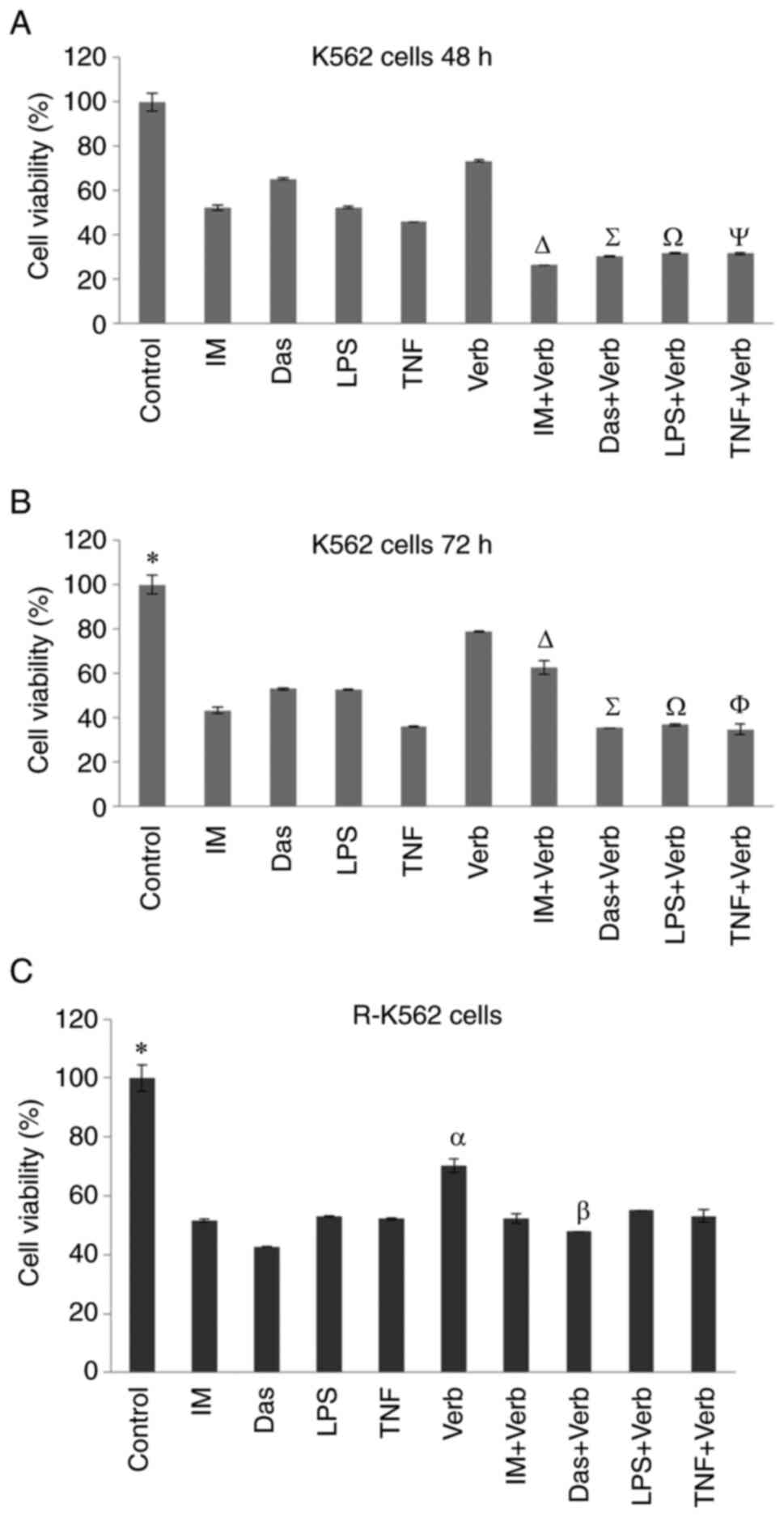 | Figure 2Viability of cells treated with 100
µM IM, 2,500 nM Das, 100 ng/ml LPS, 10 ng/ml TNF and/or 50 µg/ml
Verb K562. Cells were treated for (A) 48 h and (B) 72 h.
*P<0.05 untreated control cells vs. all other groups,
∆P<0.05 IM + Verb vs. IM and Verb,
∑P<0.05 Das + Verb vs. Das and Verb,
ΩP<0.05 LPS + Verb vs. LPS and Verb,
ΨP<0.05 TNF + Verb vs. TNF and Verb,
ΦP<0.05 TNF + Verb vs. Verb. (C) R-K562 cells were
treated for 48 h. *P<0.05 R-562 untreated control
cells vs. all other groups, αP<0.05 Verb vs. IM +
Verb, Das + Verb, LPS + Verb and TNF + Verb, βP<0.05
Das vs. Das + Verb (n=5). IM, imatinib; Das, dasatinib; LPS,
lipopolysaccharide; TNF, tumor necrosis factor α; Verb,
verbascoside. |
Abl colorimetric cell-based assay
Abl (Phospho-Tyr412) levels were determined in K562
and R-K562 cells grown in complete media supplemented with 100 µM
IM, 2,500 nM Das, 100 ng/ml LPS, 10 ng/ml TNFα and/or 50 µg/ml Verb
for 48 h. Bcr/Abl expression levels were found as elevated in
resistant cells according to previous reports (20). We evaluated the inhibition of
tyrosine phosphorylation by IM, Das, LPS, TNFα, Verb and their
combined application in K562 and R-K562 cells. Resistant cell
lines, considering that cells grown in the presence of 10 µM IM,
incubated with higher dosage of IM 100 µM. All the tested TKIs,
inducers of inflammation, Verb showed a significant decrease in the
Tyr-phosphorylation pattern after 48 h treatment in K562 and R-K562
cells (Fig. 3). This set of
experiments indicates that in our experimental system resistance
correlates with an increase in Bcr/Abl expression level. Verb
potentiates the inhibitory effect of TNF on Abl expression in K562
cells. Verb with its low toxicity to cell had revealed a
significant lowering effect on Abl expression in K562 and R-K562
cells. Consequently, we confirmed that the Verb's antileukemic
activity was related to the expression of Bcr/Abl. Verb
significantly reduced Abl levels in R-K562 cells and potentiated
the Abl expression-lowering effect of IM. However, Verb failed to
demonstrate this success with Das.
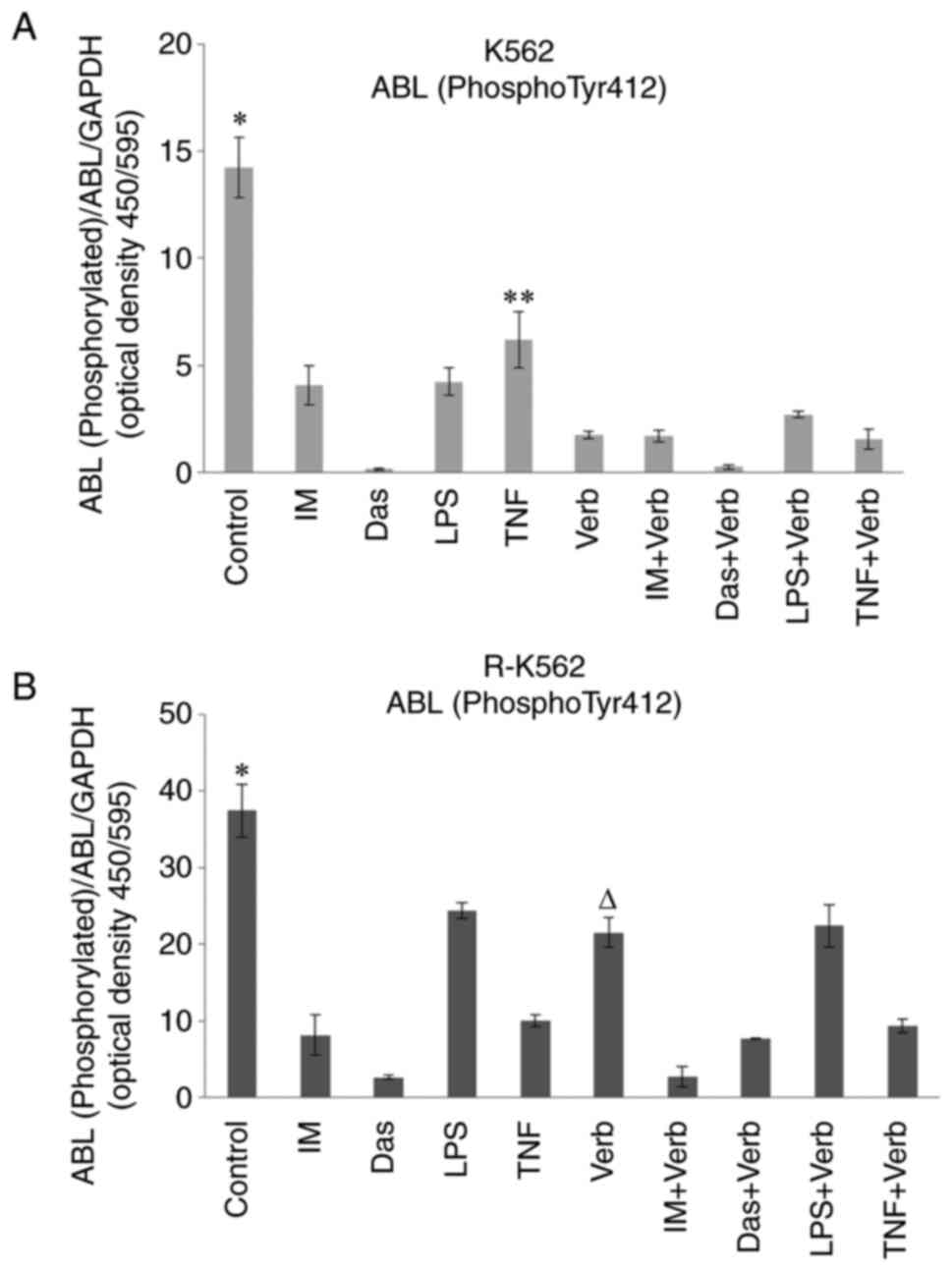 | Figure 3Bcr/Abl (Phospho-Tyr412) levels in
(A) K562 and (B) R-K562 cells incubated with 100 µM IM, 2,500 nM
Das, 100 ng/ml LPS, 10 ng/ml TNF and/or 50 µg/ml Verb for 48 h.
*P<0.05 untreated control cells vs. cells treated
with IM, Das, LPS, TNF and/or Verb, **P<0.05 TNF vs.
TNF + Verb; ∆P0.05 Verb vs. IM + Verb, Das + Verb and
TNF + Verb (n=5). IM, imatinib; Das, dasatinib; LPS,
lipopolysaccharide; TNF, tumor necrosis factor α; Verb,
verbascoside. |
Caspase-3 analysis
Caspase-3 levels were evaluated in K562 and R-K562
cells grown in complete media supplemented with 100 µM IM, 2,500 nM
Das, 100 ng/ml LPS, 10 ng/ml TNF and/or 50 µg/ml Verb for 48 h
(Fig. 4). Caspase-3 levels were
elevated by the Das, TNF, IM + Verb, Das + Verb, TNF + Verb
incubation in both cells compared to untreated control cells.
Treatment with Verb alone led to a significant increase in
caspase-3 levels in K562 cells compared to untreated control cells.
The concomitant incubation of Verb and IM, Das, LPS significantly
elevated the caspase-3 levels compared to their single use in K562
cells. Verb stimulated the caspase-3 activity in K562 cells, which
is the key factor for the initiation of the apoptotic pathway. The
data obtained from our results show Verb significantly potentiated
the effects of IM, Das and LPS on caspase-3 activity in K562 cells.
This effect was insignificantly observed in R-K562 cells.
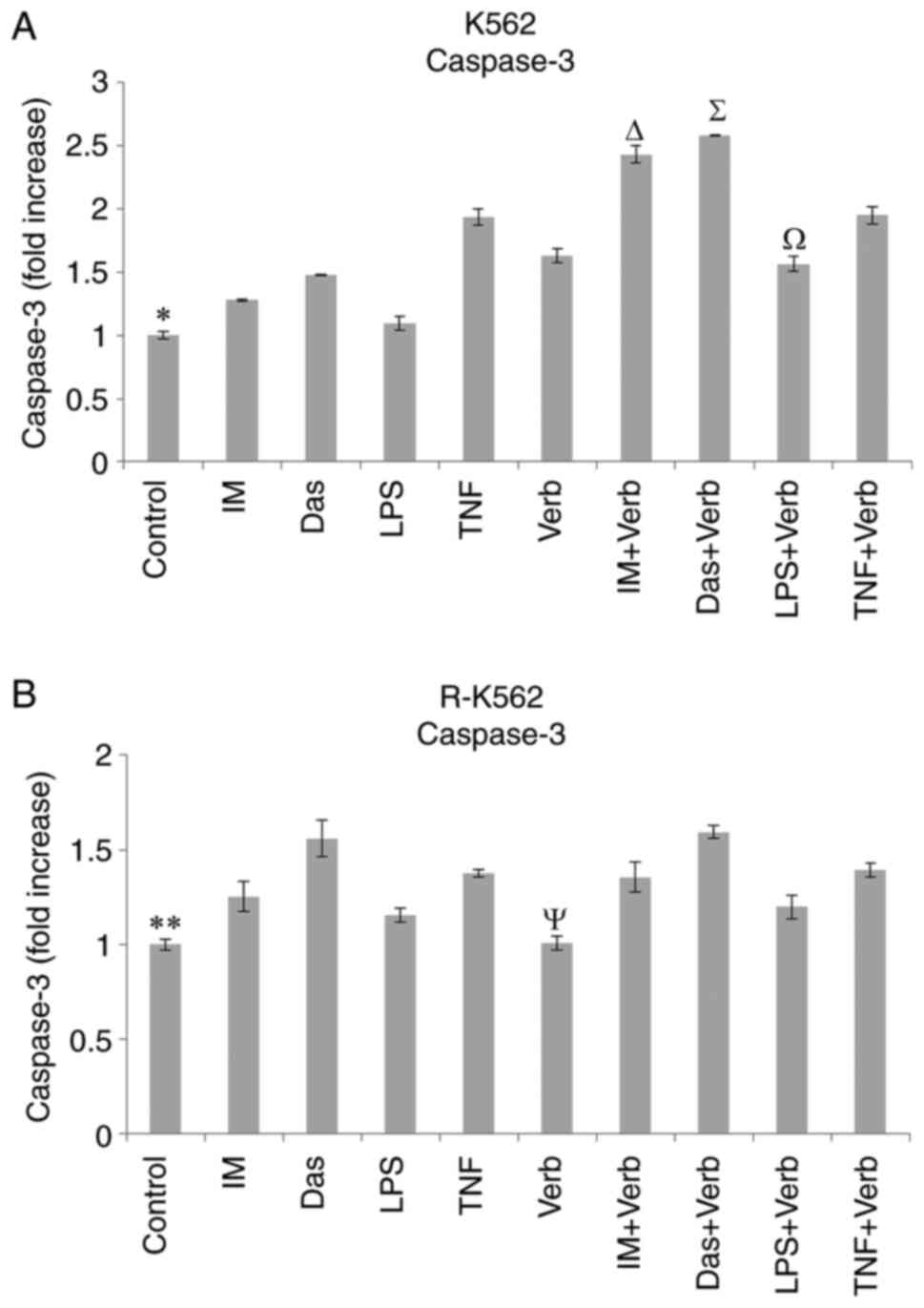 | Figure 4Caspase-3 levels in (A) K562 and (B)
R-K562 cells incubated with 100 µM IM, 2,500 nM Das, 100 ng/ml LPS,
10 ng/ml TNF and/or 50 µg/ml Verb for 48 h. *P<0.05
untreated K562 control cells vs. cells treated with Das, TNF, Verb,
IM + Verb, Das + Verb, LPS + Verb and TNF + Verb,
∆P<0.05 IM + Verb vs. IM, ∑P<0.05 Das +
Verb vs. Das, ΩP<0.05 LPS + Verb vs. LPS,
**P<0.05 untreated R-K562 control cells vs. cells
treated with IM, Das, TNF, IM + Verb, Das + Verb and TNF + Verb,
ΨP<0.05 Verb vs. IM + Verb, Das + Verb and TNF + Verb
(n=5). IM, imatinib; Das, dasatinib; LPS, lipopolysaccharide; TNF,
tumor necrosis factor α; Verb, verbascoside. |
Total/phosphorylated ERK, JNK, p38
protein expression
Cell-based sandwich ELISA was used to determine
total and phosphorylated p38, JNK, ERK protein expression levels in
R-K562 cells. Bcr/Abl is activated through the activation of the
ERK/MAPK pathway (31). There is
no study in the literature investigating ERK, p38MAPK, JNK pathway
regulated by Verb in resistant CML cells. Regarding MAPK signalling
pathway family members, Verb and its combinational therapy with IM,
Das, LPS and TNF cause a significant downregulation in ERK1/2
regard to the untreated R-K562 control cells but the Verb and/or
TKIs caused an upregulation in the phospho-JNK and phospho-p38
levels in 48 h treatment. Verb increased ERK/MAPK signalling
(Fig. 5). There was no significant
difference in the total protein levels of ERK1/2 and p-38 following
the incubations, compared to the untreated control. Total protein
level of JNK increased by the incubations of Verb and/or IM, Das,
LPS, TNF. Verb reinforced the TKIs, LPS and TNF's activity in
upregulating p-JNK and p-p38. Concomitant incubation of Verb with
IM, Das, LPS and TNF decreased ER1/2 levels.
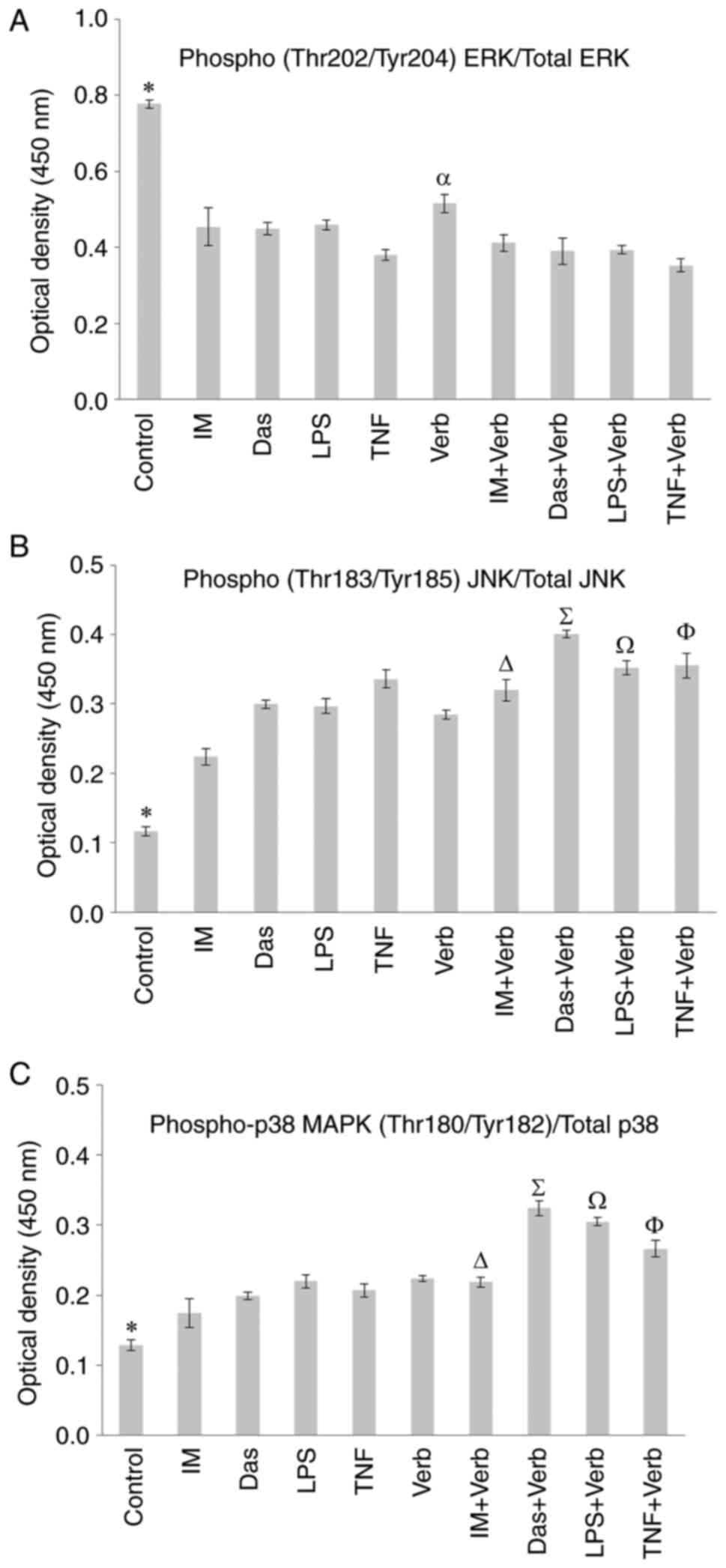 | Figure 5Phosphorylated/total protein levels
of (A) ERK1/2, (B) JNK and (C) p38. R-K562 cells were incubated
with 100 µM IM, 2,500 nM Das, 100 ng/ml LPS, 10 ng/ml TNF and/or 50
µg/ml Verb for 48 h. *P<0.05 untreated control cells
vs. all other groups, αP<0.05 Verb vs. IM + Verb, Das
+ Verb, LPS + Verb and TNF + Verb, ∆P<0.05 IM + Verb
vs. IM, ∑P<0.05 Das + Verb vs. Das and Verb,
ΩP<0.05 LPS + Verb vs. LPS and Verb,
ΦP<0.05 TNF + Verb vs. TNF and Verb (n=5). IM,
imatinib; Das, dasatinib; LPS, lipopolysaccharide; TNF, tumor
necrosis factor a; Verb, verbascoside; p38, p38 MAP kinase; JNK,
c-Jun N-terminal kinase; ERK, extracellular signal regulated
kinase. |
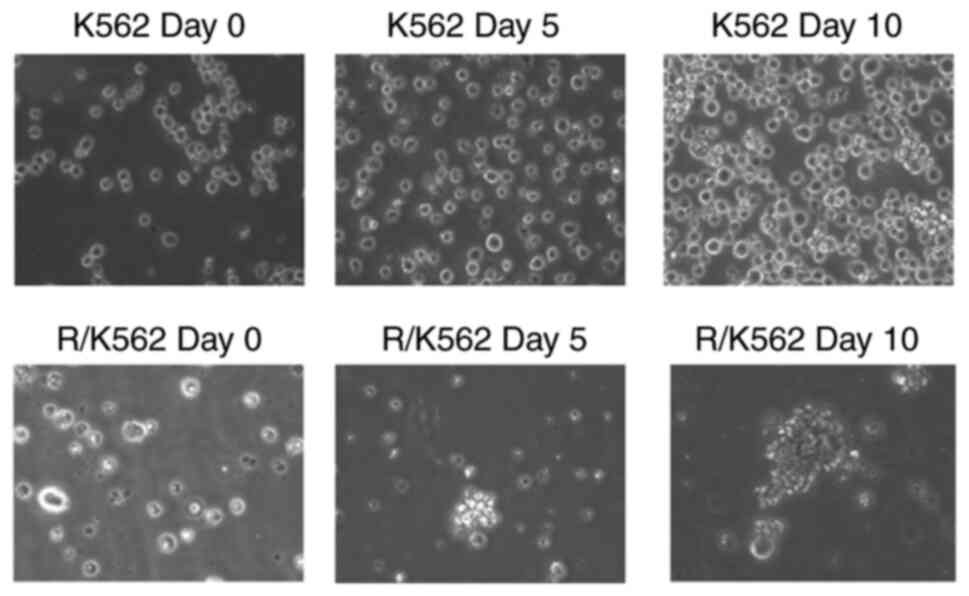
Spheroid formation assay
The in vitro spheroid formation assay is a
common assay used to measure the self-renewal and multipotent
nature of the cancer cell subpopulations within cancer cell line.
In our study we incubated K562 and R-K562 cells in soft agar
culture medium for 10 days. Multicellular tumour spheroid formation
was observed on 5 and 10th days. K562 cells do not form tumour
spheroids whereas R-K562 cells show spheroid formation. We have
observed the spheroid formation in R-K562 cells in time dependent
manner (Fig. 6). Spheroid
formation assay confirms the resistant characteristics of the
R-K562 cells.
TAS, TOS and OSI
The effects of Verb and/or IM, Das, LPS, TNF for 48
h on oxidative stress were determined in K562 and R-K562 cells by
using TAS-TOS assay. TAS levels significantly decreased in K562
cells treated with IM, Das, LPS and TNF compared to untreated K562
control cells. Verb and its combinational treatment with IM, Das,
LPS and TNF also led to a significant decrease in TAS levels in
K562 cells. The antioxidant capacity of R-K562 cells was
significantly reduced by IM, Das, LPS and TNF. Verb decreased the
TAS levels when used together with the Das, LPS and TNF. The TOS
levels significantly increased in K562 and R-K562 cells treated
with IM, Das, LPS, TNF, Das + Verb, LPS + Verb and TNF + Verb
groups. Verb potentiated the TOS inducing activity of Das, LPS and
TNF in K562 cells. OSI levels were elevated by all the tested
substances (Table I).
 | Table IOxidative stress parameters of K562
and R-K562 cells. |
Table I
Oxidative stress parameters of K562
and R-K562 cells.
| | TAS, mmol Trolox
equiv./l | TOS, µmol
H2O2 equiv./l | OSI, arbitrary
unit |
|---|
| Experimental
groups | K562 | R-K562 | K562 | R-K562 | K562 | R-K562 |
|---|
| Control | 0.73±0.03 | 1.34±0.02 | 0.25±0.06 | 5.69±1.10 | 0.03 | 0.43 |
| IM |
0.54±0.08a |
1.08±0.12a |
5.70±2.13a |
18.91±1.42a | 1.06 | 1.75 |
| Das |
0.31±0.17a |
0.59±0.09a |
16.03±2.47a |
28.19±2.53a | 5.23 | 4.81 |
| LPS |
0.21±0.05a |
0.70±0.06a |
15.42±2.15a |
33.87±5.81a | 7.43 | 4.81 |
| TNF |
0.28±0.11a |
0.64±0.20a |
23.82±4.37a |
30.77±1.11a | 8.49 | 4.80 |
| Verb |
0.30±0.01a | 1.17±0.09 |
17.00±2.27a | 6.49±2.01 | 5.66 | 0.56 |
| IM + Verb |
0.20±0.00a,b | 1.10±0.03 | 10.99±5.59 |
19.98±8.93a,b | 5.50 | 1.82 |
| Das + Verb |
0.14±0.05a |
0.73±0.05a,b |
37.25±2.05a,c |
31.98±7.02a,b | 7.26 | 4.37 |
| LPS + Verb |
0.14±0.03a |
0.88±0.15a,b |
26.36±5.11a,d |
32.6±2.93a,b | 18.42 | 3.72 |
| TNF + Verb |
0.19±0.10a |
0.69±0.12a,b |
37.93±9.44a-e |
27.56±7.75a,b | 19.76 | 4.02 |
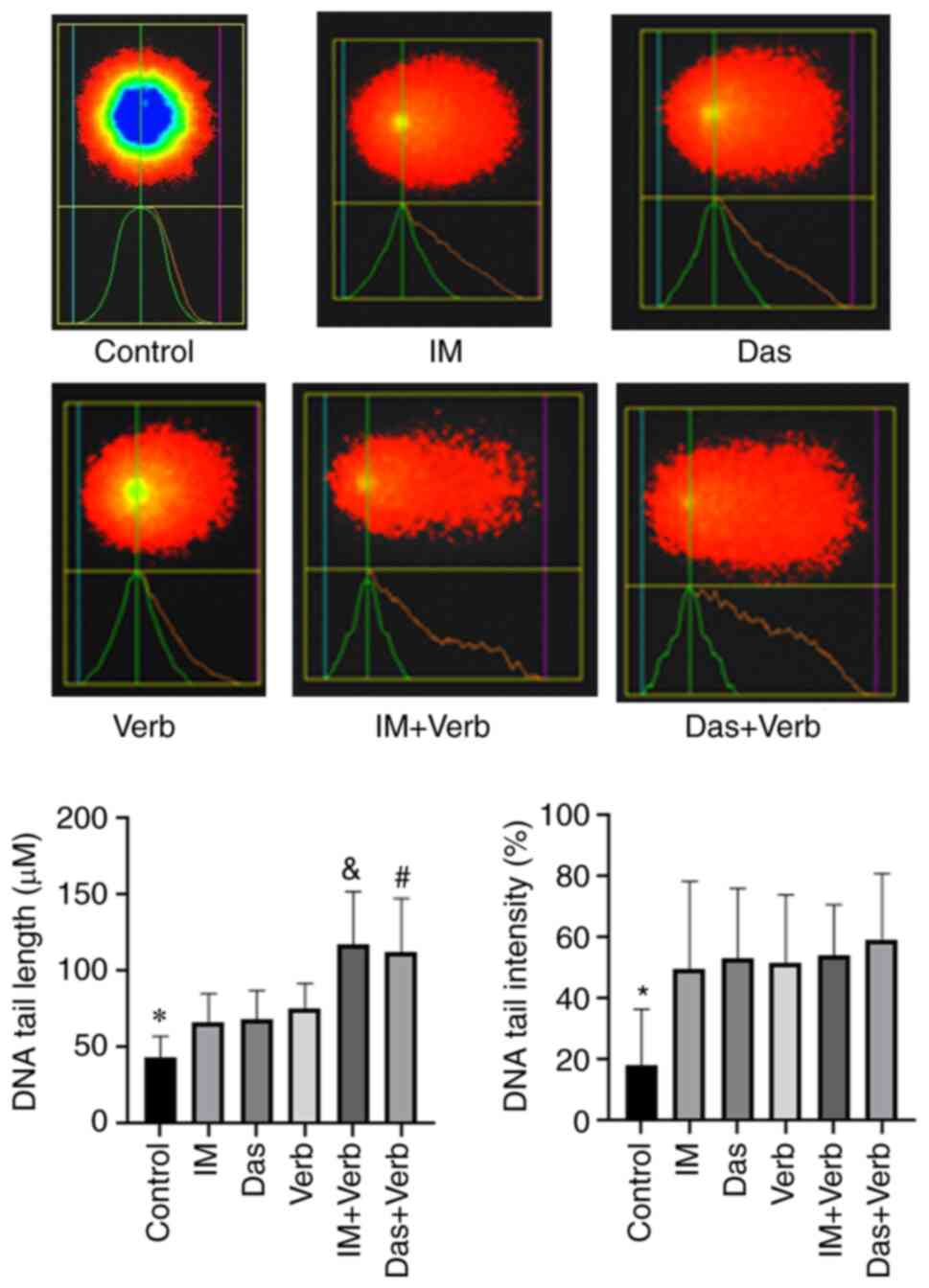
Comet assay
A Comet assay was measured DNA damage in K562 cells
treated with IM, Das, Verb, IM + Verb, Das + Verb for 48 h.
Findings were evaluated in accordance with DNA tail length and DNA
tail intensity. In K562 cells treated with IM, Das, IM + Verb, Das
+ Verb tail length, tail intensity of DNA were found to be elevated
with respect to the control cells. Verb enhanced the DNA tail
length when combined with IM or Das compared to IM or Das alone
groups (Fig. 7).
Discussion
Bcr/Abl tyrosine kinase inhibitors have been
demonstrated to have significant therapeutic effects in leukaemia.
But CML is a life-threatening disease for patients with blast
phases, mainly resulting from TKI resistance (32). Resistance and toxicity of tyrosine
kinase inhibitors have been frequently reported. Resistant
CML cells could have improved DNA repair, impaired apoptosis,
modification in membrane transport systems, proteins and enzymes
(33). Thus, a TKI in combination
with additional drugs, potentiating the efficacy of Bcr/Abl
inhibitors for the treatment of leukaemia seems to be an
encouraging strategy to tackle the challenges related to CML
treatment. There is a demand for continued research into
alternative pharmacological approaches with not only more effective
but also with less toxic effects. Because of the anticancer effect
of natural extracts, they have drawn attention and been
investigated in many studies. Verb is hydrophilic in nature and
possesses pharmacologically beneficial activities for human health,
including anti-oxidant, anti-inflammatory and anti-neoplastic
properties (1). It has been
reported that Verb prevented the proliferation of HL-60 leukaemia
cell line in a time and concentration dependent manner.
Additionally, cell cycle progression is blocked by Verb at the G1
phase (9). A new delivery system
is being investigated by the researchers for boosting Verb's
chemotherapeutic effect against drug-resistant leukaemia cells.
Verb containing nanoproduct showed visible tumour suppressor
effects by elevating caspase 3 expression (34). The accumulated evidence shows that
Verb could have potential antitumour activities in leukaemia, but
its activity against sensitive and resistant K562 cells in the
presence of TKIs is yet to be investigated. K562 cells have been
largely used as a model system for investigating drug development
technologies against CML. Different compounds are under
investigation to show cell viability changes in K562 cells to
induce apoptosis. We explored the cytotoxic activity of Verb and/or
TKIs, LPS and TNF against IM sensitive and resistant leukaemia cell
lines and found that Verb possesses a remarkable antiproliferative
effect on the both K562 and R-K562 cells additively with the TKIs;
IM and Das or inducers of inflammation and apoptosis LPS and TNF
(Fig. 2).
Cell viability was observed as 51.5% after IM
incubation and 70.23% after Verb incubation in R-K562 cells
(Fig. 2C). Concomitant use of Verb
with IM, led to a significant decrease in cell viability (52.29%)
compared to single treatment of Verb. But the viability of R-K562
cells was not significantly increased in the IM + Verb treatment
compared to single the IM treatment. This observation could result
from the increased proliferation rate of R-K562 cells compared to
their non-resistant counterparts. The expression of Ki67 is
strongly associated with tumour cell proliferation and growth and
is widely used in the routine pathological investigation as a
proliferation marker. Limitations of this study are that the lack
of cell viability testing on the range of Verb concentrations on
K562 and R-K562 cells and the viability of K562 cells with that of
R-K562 cells in response to different concentrations of IM was not
compared. We could not evaluate the Ki67 expression levels in our
study groups and the establishment of R-K562 cells was confirmed
only by Spheroid formation assay and Bcr/Abl expression levels.
Another limitation of our study was Verb failed to reduce Abl
levels when combined with the Das in R-K562 cells compared to Das
alone treatment and did not potentiate the Abl expression-lowering
effect of Das.
Bcr/Abl contributes to the development of CML by
enhancing the proliferation potential of hematopoietic progenitors
and preventing progenitor cells from apoptosis. To get more
insights into Verb's molecular mechanism, we assessed its effect on
Abl1 activity, which is amplified in R-K562 cells. As anticipated,
Verb inhibited Abl1 phosphorylation at 50 µg/ml concentration. Our
results demonstrate that Bcr/Abl protein levels are downregulated
by Verb (Fig. 3), indicating that
Verb's anti-leukemic activity is related to Bcr/Abl expression. In
our study Verb treatment significantly inhibited Abl expressions,
showing that Abl inhibition by Verb contributed to the growth
inhibitions in both K562 and K-562 cells. Although Abl is
considered the trigger of leukemogenesis, targeting Abl kinase
activity alone may be insufficient because of Abl independent
downstream pathways. TKIs, LPS, TNF and/or Verb showed a
significant decrease in the Tyr-412 phosphorylation of Abl in K562
and R-K562 cells. Verb insignificantly potentiates the inhibitory
effect of TKIs, LPS and TNF on Abl expression in R-K562 cells. Src
family kinases (SFK) lead to the activation of Bcr/Abl in the
progression of CML. SFK inhibitor Das, may suppress Bcr/Abl
phosphorylation and decrease the tyrosine kinase activity of
Bcr/Abl. Accordingly, Das treatment significantly alleviated the
Abl phosphorylation in K562 and R-K562 cells compared to untreated
control cells. SFKs are also involved in IM resistance. To overcome
resistance problems in CML, combinations of drugs with IM provided
an emerging therapeutic concept. Verb induced programmed cell death
in CML cells more efficiently than IM, Das, LPS or TNF compared to
their single use at 48 h of treatment. It has been reported that
Das induces the apoptosis of leukemic cells (17,35).
De Martino et al revealed that citrals containing Verb were
also apoptotic against leukaemia cells by activation of
caspase-3(36). Our data showed
that the combination of Verb with IM or Das induces apoptosis
additively in both cells. Verb combined TKIs significantly
stimulated caspase-3 expression in K562 and R-K562 cells (Fig. 4), which might offer a new
chemotherapeutic approach in leukaemia treatment. As a driving
force for leukaemia, Bcr/Abl is able to trigger the MAPK pathway.
Tyr177 phosphorylation results in the activation of the MAPK
pathway to improve lymphoid transformation (37). One of the aims of our study was to
determine whether Verb exerted its anti-leukemic effects by
inhibiting the MAPK signalling in R-K562 cells. p38 and JNK result
in growth arrest and cell death whereas the activation of ERK
results in cell division and differentiation. In this study, we
found that treatment with Verb and its TKI combination did not
influence on the total protein expression of p38, ERK1/2 and JNK.
Our data reveals that phosphorylated ERK1/2 is suppressed by Verb
and/or IM, Das, LPS, TNF (Fig. 5).
Verb's inhibitory effects on the phospho-ERK levels may be
regulated by the suppression of Bcr/Abl upstream protein tyrosine
kinases. According to the results of our study Verb's anti-leukemic
activity is linked to the inhibition of MAPK/ERK signalling and
decreased Bcr/Abl expression. Verb is found to inhibit the ERK1/2
activation, suggesting that Verb may be an effective MAPK
signalling pathway inhibitor. ERK1/2 phosphorylation plays
important role in Bcr/Abl controlled resistance response and
apoptosis (20). It has been shown
that Erk1/2 phosphorylation is interrupted in R-K562 cells that are
cultured continuously in 10 µM IM (38). The phosphorylated p-38 and JNK were
significantly elevated by Verb and/or IM, Das, LPS, TNF suggesting
that inhibition of Src and Abl could be involved in MAPK signalling
in R-K562 cells which is affected by Verb. Dumka et al
reported that Das related p38 activation plays an important role in
the antileukemic responses, while inhibition or knockdown of p38
reverses Das-related apoptosis, cell cycle arrest, and
anti-proliferative effects (39).
Decreased levels of phosphorylation may result from the protein
tyrosine kinase inhibition or protein tyrosine phosphatase
activation. Bcr/Abl had been noted to be an important subject of
MAPK mediated resistance to IM (40). Phospho-ERK (key molecule of
oncogenic MAPK signalling) levels were significantly diminished by
Verb treatment, explaining the inhibitory effects of Verb on
MAPK/ERK signalling. It has been shown that the MAPK signalling
cascade is a possible mechanism underlying the effect of Verb in
inflammation-activated monocytes (41). Verb treatment restores the kinase
activity which is fundamental for the resumption of cell function
in response to inflammatory damage. Authors reported that Verb
attenuates LPS-induced proinflammatory mediator production in
activated U937 cells, preventing cell damage. We have determined
the MAPK signalling family members in leukaemia cells. We have
found that phosphorylation of JNK and p38 was elevated by the Verb
and/or IM, Das, LPS, TNF. Verb acts as an activator of JNK and p38
which result in cell damage mediated by MAPK family members. We
have investigated the colony forming capacity of K562 and R-K562
cells (Fig. 6). In vitro
spheroid formation capacity evaluate tumour-initiating potential.
RK562 cells generate compact spheroids, whereas K562 cells are not
capable of this type of spheroid formation, according to a recent
study by Baykal-Kose and Yalcin (38). Puissant et al report that
IM-resistant and highly adherent K562 cells can produce spheroid,
whereas IM-sensitive counterparts cannot (42). Decreased ROS levels elevate tumour
cell survival, while high levels of ROS can overcome tumour growth
by triggering inhibitors of cell cycle (43). In our study Verb elevated ROS
levels additively with TKIs in both sensitive and resistant cells
by increasing the oxidant capacity and decreasing the antioxidant
capacity. It is also shown by the other researchers reporting that
the Verb has the potential to elevate ROS levels inside the tumour
cells (44). Taken together, these
data suggest that TKIs and Verb, exert cytotoxic effects on K562
and induce apoptosis by triggering ROS. Inflammation was triggered
by LPS in K562 and R-K562 cells and the role of Bcr/Abl, caspase-3
and ROS axis was investigated in our study. The results are
compatible with previous studies obtained by Wang et al,
whose data confirmed that LPS treatment increased ROS production in
K562 cells (45). Verb increased
the TOS levels of K562 cells compared to LPS's single treatment.
TAS levels diminished by the LPS and/or Verb in both cells.
Speranza et al reported that Verb has anti-inflammatory
activity on LPS treated THP-1, human myelomonocytic leukaemia
cells, the anti-inflammatory effects of Verb are related to
inhibition of intracellular O2.- production
and the suppression of anti-oxidant enzymes at the
post-translational level (46).
ROS regulates TNF induced signalling pathways. In our findings it
can be observed that TNF significantly increased TOS levels alone
or in combination with Verb. González-Flores et al reported
that treatment of K562 cells with TNFα stimulated intracellular ROS
levels followed by raised caspase-3(19). Similarly caspase-3 levels were
elevated by TNF and TNF + Verb in both cells in our study. TNFα
triggered intracellular ROS generation and led to caspase
activation and apoptosis in K562 and R-K562 cells.
The production of ‘comet tails’ during agarose gel
electrophoresis can be used to identify DNA damage. Tails with a
greater DNA content indicate a more severe DNA damage profile. We
have evaluated DNA damage induced by IM, Das and/or Verb in K562
cells. The comet experiment revealed that Verb was potentiated the
effect of IM and Das on DNA damage by increasing the length and
intensity of DNA tails (Fig. 7).
IM and Das, which are commonly used in the treatment of
haematological malignancies, and their Verb combination led to
increased ROS levels shown by the elevated TOS levels (Table I). Besides the damaging effects on
DNA IM, Das and their Verb combination treatment also resulted in
apoptosis by activating the caspase-3. Our results indicate that
the K562 cells are vulnerable to the genotoxic stress of IM, Das
and/or Verb, when compared with K562 control cells. The increased
DNA damage with the tested compounds can probably be explained by
the fact that the mechanism of DNA damage is oxidative.
Mutations in the Abl kinase domain occur by
conformational changes that reduce the affinity of the ATP-binding
pocket of the TKI, thus leading to drug resistance. The resistance
of IM occurs due to point mutations. T212R mutation in Abl kinase
domain is determined in the SH2-SH3 domain that generates
conformation changes to prevent the binding of TKIs. Abl kinase
gatekeeper T315I mutant has proven challenging to inhibit with ATP
mimetics. Therefore, severe mutations have urged the second-line
therapy (Das, nilotinib, bosutinib) against these particular
Bcr-Abl mutants present in the patients (47). Tyr177 and Tyr412 are the two
constitutive active fusions of Bcr-Abl. Tyr177 serves as a binding
site for the Grb2 adaptor and Tyr412 is required for the activity
and the control of c-Abl, which stabilizes the enzyme's inactive or
active conformation in a phosphorylation-dependent manner (48). The catalytic domain includes a
substrate-binding site. Inhibitors targeting this site would be
less affected by mutations. IM works by binding to the ATP cleft of
the inactive form of Bcr-Abl and preventing the conformational
change required for kinase activation. The literature reported that
the amide substituent on the phenyl ring of IM and Das provided the
molecule with inhibitory activity against Abl tyrosine kinase. The
core phenyl or heterocyclic rings of these drugs occupy the adenine
pocket of Abl (49). Verb has many
reactive sites with high selectivity for combinatorial chemistry.
Verb is a phenylethanoid consisting of a cinnamic acid and
hydroxyphenylethyl moieties attached to a β-glucopyranose through a
glycosidic bond. In a recent study the strong inhibition potency of
Verb had been shown against Carbonic Anhydrase Enzyme. Docking
results confirmed the strong interactions between Verb and the
active site of the enzyme (50). Similar binding properties
could be found between Abl and the Verb, although further research
is needed. Although the biochemical roles and stereochemistry of
the Bcr-Abl enzyme active sites have been well identified, and
numerous natural and synthetic compounds have been tested for
inhibitory action, there is no one-size-fits-all structural type
that is the most effective.
Verb is suggested to be possible for the future use
of chemotherapy and co-therapy in the clinic (51). We revealed that Verb could repress
cell growth and induce apoptosis in K562 and R-K562 cells. The
mechanisms involve the inhibition of the Abl oncoprotein and
regulation of its downstream p38-MAPK/JNK pathway. Verb effectively
suppressed the crosstalk between MAPK and Bcr/Abl signalling,
thereby increasing the sensitivity of CML cells towards TKIs
without suppressing the inflammation. Thus, Verb may be used
additively with Das or IM in the treatment of CML. More research is
needed to elucidate the precise mechanisms by that these molecules
exert their effects.
Acknowledgements
Not applicable.
Funding
Funding: This study was supported by Pamukkale University (grant
nos. 2020BSP009, 2020HZDP005, 2020HZDP007 and 2021HZDP008).
Availability of data and materials
The datasets used and/or analysed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
ACD designed and conducted all the experimental
procedures of the study. GAC interpreted the results. EKT conducted
the measurement of TAS/TOS. FA performed the comet assay. ACD and
EKT confirm the authenticity of all the raw data. All authors read
and approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Alipieva K, Korkina L, Orhan IE and
Georgiev MI: Verbascoside-a review of its occurrence,
(bio)synthesis and pharmacological significance. Biotechnol Adv.
32:1065–1076. 2014.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Song HS and Sim SS: Acteoside inhibits
alpha-MSH-induced melanin production in B16 melanoma cells by
inactivation of adenyl cyclase. J Pharm Pharmacol. 61:1347–1351.
2009.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Akdemir Z, Kahraman C, Tatlı II, Küpeli
Akkol E, Süntar I and Keles H: Bioassay-guided isolation of
anti-inflammatory, antinociceptive and wound healer glycosides from
the flowers of Verbascum mucronatum Lam. J Ethnopharmacol.
136:436–443. 2011.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Georgiev M, Pastore S, Lulli D, Alipieva
K, Kostyuk V, Potapovich A, Panetta M and Korkina L: Verbascum
xanthophoeniceum-derived phenylethanoid glycosides are potent
inhibitors of inflammatory chemokines in dormant and
interferon-gamma-stimulated human keratinocytes. J Ethnopharmacol.
144:754–760. 2012.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Wartenberg M, Budde P, De Mareés M,
Grünheck F, Tsang SY, Huang Y, Chen ZY, Hescheler J and Sauer H:
Inhibition of tumor-induced angiogenesis and
matrix-metalloproteinase expression in confrontation cultures of
embryoid bodies and tumor spheroids by plant ingredients used in
traditional chinese medicine. Lab Invest. 83:87–98. 2003.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Attia YM, El-Kersh DM, Wagdy HA and
Elmazar MM: Verbascoside: Identification, quantification, and
potential sensitization of colorectal cancer cells to 5-FU by
targeting PI3K/AKT pathway. Sci Rep. 8(16939)2018.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Qin YZ and Huang XJ: Molecular detection
of BCR-ABL in chronic myeloid leukemia. Methods Mol Biol.
1465:1–15. 2016.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Zhang F, Jia Z, Deng Z, Wei Y, Zheng R and
Yu L: In vitro modulation of telomerase activity, telomere length
and cell cycle in MKN45 cells by verbascoside. Planta Med.
68:115–118. 2002.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Lee KW, Kim HJ, Lee YS, Park HJ, Choi JW,
Ha J and Lee KT: Acteoside inhibits human promyelocytic HL-60
leukemia cell proliferation via inducing cell cycle arrest at G0/G1
phase and differentiation into monocyte. Carcinogenesis.
28:1928–1936. 2007.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Bardellini E, Amadori F, Schumacher RF,
D'Ippolito C, Porta F and Majorana A: Efficacy of a solution
composed by verbascoside, polyvinylpyrrolidone (PVP) and sodium
hyaluronate in the treatment of chemotherapy-induced oral mucositis
in children with acute lymphoblastic leukemia. J Pediatr Hematol
Oncol. 38:559–562. 2016.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Yilmaz M and Jabbour E: Tyrosine kinase
inhibitors early in the disease course: Lessons from chronic
myelogenous leukemia. Semin Oncol. 42:876–886. 2015.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Assouline S and Lipton JH: Monitoring
response and resistance to treatment in chronic myeloid leukemia.
Curr Oncol. 18:e71–e83. 2011.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Jabbour EJ, Cortes JE and Kantarjian HM:
Resistance to tyrosine kinase inhibition therapy for chronic
myelogenous leukemia: A clinical perspective and emerging treatment
options. Clin Lymphoma Myeloma Leuk. 13:515–529. 2013.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Bixby D and Talpaz M: Mechanisms of
resistance to tyrosine kinase inhibitors in chronic myeloid
leukemia and recent therapeutic strategies to overcome resistance.
Hematology Am Soc Hematol Educ Program. 461–476. 2009.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Mitchell R, Hopcroft LEM, Baquero P, Allan
EK, Hewit K, James D, Hamilton G, Mukhopadhyay A, O'Prey J, Hair A,
et al: Targeting BCR-ABL-independent TKI resistance in chronic
myeloid leukemia by mTOR and autophagy inhibition. J Natl Cancer
Inst. 110:467–478. 2018.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Olivieri A and Manzione L: Dasatinib: A
new step in molecular target therapy. Ann Oncol. 18 (Suppl
6):vi42–vi46. 2007.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Damiano S, Montagnaro S, Puzio MV,
Severino L, Pagnini U, Barbarino M, Cesari D, Giordano A, Florio S
and Ciarcia R: Effects of antioxidants on apoptosis induced by
dasatinib and nilotinib in K562 cells. J Cell Biochem.
119:4845–4854. 2018.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Welner RS, Amabile G, Bararia D, Czibere
A, Yang H, Zhang H, Pontes LL, Ye M, Levantini E, Di Ruscio A, et
al: Treatment of chronic myelogenous leukemia by blocking cytokine
alterations found in normal stem and progenitor cells. Cancer Cell.
27:671–681. 2015.PubMed/NCBI View Article : Google Scholar
|
|
19
|
González-Flores D, Rodriguez AB and
Pariente JA: TNFα-induced apoptosis in human myeloid cell lines
HL-60 and K562 is dependent of intracellular ROS generation. Mol
Cell Biochem. 390:281–287. 2014.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Yu C, Krystal G, Varticovksi L, McKinstry
R, Rahmani M, Dent P and Grant S: Pharmacologic mitogen-activated
protein/extracellular signal-regulated kinase
kinase/mitogen-activated protein kinase inhibitors interact
synergistically with STI571 to induce apoptosis in
Bcr/Abl-expressing human leukemia cells. Cancer Res. 62:188–199.
2002.PubMed/NCBI
|
|
21
|
Wu GS: Role of mitogen-activated protein
kinase phosphatases (MKPs) in cancer. Cancer Metastasis Rev.
26:579–585. 2007.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Deininger MW and Druker BJ: Specific
targeted therapy of chronic myelogenous leukemia with imatinib.
Pharmacol Rev. 55:401–423. 2003.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Shah NP, Kasap C, Weier C, Balbas M,
Nicoll JM, Bleickardt E, Nicaise C and Sawyers CL: Transient potent
BCR-ABL inhibition is sufficient to commit chronic myeloid leukemia
cells irreversibly to apoptosis. Cancer Cell. 14:485–493.
2008.PubMed/NCBI View Article : Google Scholar
|
|
24
|
García LT, Leal AF, Morenoa ÉM, Stashenko
EE and Arteaga J: Differential anti-proliferative effect on K562
leukemia cells of Lippia alba (Verbenaceae) essential oils
produced under diverse growing, collection and extraction
conditions. sInd Crops Prod. 96:140–148. 2017.
|
|
25
|
Oukerrou MA, Tilaoui M, Mouse HA, Bouchmaa
N and Zyad A: Differential cytotoxic activity of essential oil of
Lippia citriodora from different regions in morocco. Chem
Biodivers. 14:2017.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Yang X, Feng W, Wang R, Yang F, Wang L,
Chen S, Ru Y, Cheng T and Zheng G: Repolarizing heterogeneous
leukemia-associated macrophages with more M1 characteristics
eliminates their pro-leukemic effects. Oncoimmunology.
7(e1412910)2017.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Dorey K, Engen JR, Kretzschmar J, Wilm M,
Neubauer G, Schindler T and Superti-Furga G: Phosphorylation and
structure-based functional studies reveal a positive and a negative
role for the activation loop of the c-Abl tyrosine kinase.
Oncogene. 20:8075–8084. 2001.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Baykal-Köse S, Acikgoz E, Yavuz AS, Gönül
Geyik Ö, Ateş H, Sezerman OU, Özsan GH and Yüce Z: Adaptive
phenotypic modulations lead to therapy resistance in chronic
myeloid leukemia cells. PLoS One. 15(e0229104)2020.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Erel O: A novel automated method to
measure total antioxidant response against potent free radical
reactions. Clin Biochem. 37:112–119. 2004.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Olive PL and Banáth JP: The comet assay: A
method to measure DNA damage in individual cells. Nat Protoc.
1:23–29. 2006.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Modi H, Li L, Chu S, Rossi J, Yee JK and
Bhatia R: Inhibition of Grb2 expression demonstrates an important
role in BCR-ABL-mediated MAPK activation and transformation of
primary human hematopoietic cells. Leukemia. 25:305–312.
2011.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Flis S and Chojnacki T: Chronic
myelogenous leukemia, a still unsolved problem: Pitfalls and new
therapeutic possibilities. Drug Des Devel Ther. 13:825–843.
2019.PubMed/NCBI View Article : Google Scholar
|
|
33
|
Alfarouk KO, Stock CM, Taylor S, Walsh M,
Muddathir AK, Verduzco D, Bashir AH, Mohammed OY, Elhassan GO,
Harguindey S, et al: Resistance to cancer chemotherapy: Failure in
drug response from ADME to P-gp. Cancer Cell Int.
15(71)2015.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Ma Z, Zhao X, Jiang C, Yu J, Wu J and Zeng
X: Gold nanoshells with verbascoside induce the apoptosis of
drug-resistant leukemia cells through caspases pathway and inhibit
tumor growth. J Nanosci Nanotechnol. 16:7118–7124. 2016.
|
|
35
|
Dalgıç CT, Kaymaz BT, Özkan MC, Dalmızrak
A, Şahin F and Saydam G: Investigating the role of JAK/STAT pathway
on dasatinib-induced apoptosis for CML cell model K562. Clin
Lymphoma Myeloma Leuk. 15 (Suppl):S161–S166. 2015.PubMed/NCBI View Article : Google Scholar
|
|
36
|
De Martino L, D'Arena G, Minervini MM,
Deaglio S, Fusco BM, Cascavilla N and De*Feo V: Verbena officinalis
essential oil and its component citral as apoptotic-inducing agent
in chronic lymphocytic leukemia. Int J Immunopathol Pharmacol.
22:1097–1104. 2009.PubMed/NCBI View Article : Google Scholar
|
|
37
|
Ma L, Xu Z, Wang J, Zhu Z, Lin G, Jiang L,
Lu X and Zou C: Matrine inhibits BCR/ABL mediated ERK/MAPK pathway
in human leukemia cells. Oncotarget. 8:108880–108889.
2017.PubMed/NCBI View Article : Google Scholar
|
|
38
|
Baykal-Kose S and Yalcin P: Altered
apoptotic protein expressions characterize the survival of
Bcr-Abl-independent drug-resistant chronic myeloid leukemia cell
line. 1 J Basic Clin Health Sci. 1:1–5. 2021.
|
|
39
|
Dumka D, Puri P, Carayol N, Lumby C,
Balachandran H, Schuster K, Verma AK, Terada LS, Platanias LC and
Parmar S: Activation of the p38 Map kinase pathway is essential for
the antileukemic effects of dasatinib. Leuk Lymphoma. 50:2017–2029.
2009.PubMed/NCBI View Article : Google Scholar
|
|
40
|
Aceves-Luquero CI, Agarwal A,
Callejas-Valera JL, Arias-González L, Esparís-Ogando A, del Peso
Ovalle L, Bellón-Echeverria I, de la Cruz-Morcillo MA, Galán Moya
EM, Moreno Gimeno I, et al: ERK2, but not ERK1, mediates acquired
and ‘de novo’ resistance to imatinib mesylate: implication for CML
therapy. PLoS One. 4(e6124)2009.PubMed/NCBI View Article : Google Scholar
|
|
41
|
Pesce M, Franceschelli S, Ferrone A, De
Lutiis MA, Patruno A, Grilli A, Felaco M and Speranza L:
Verbascoside down-regulates some pro-inflammatory signal
transduction pathways by increasing the activity of tyrosine
phosphatase SHP-1 in the U937 cell line. J Cell Mol Med.
19:1548–1556. 2015.PubMed/NCBI View Article : Google Scholar
|
|
42
|
Puissant A, Dufies M, Fenouille N, Ben
Sahra I, Jacquel A, Robert G, Cluzeau T, Deckert M, Tichet M, Chéli
Y, et al: Imatinib triggers mesenchymal-like conversion of CML
cells associated with increased aggressiveness. J Mol Cell Biol.
4:207–220. 2012.PubMed/NCBI View Article : Google Scholar
|
|
43
|
Cort A, Ozben T, Saso L, De Luca C and
Korkina L: Redox control of multidrug resistance and its possible
modulation by antioxidants. Oxid Med Cell Longev.
2016(4251912)2016.PubMed/NCBI View Article : Google Scholar
|
|
44
|
Vasincu A, Neophytou CM, Luca SV,
Skalicka-Woźniak K, Miron A and Constantinou AI:
6-O-(3',4'-di-O-trans-cinnamoyl)-α-l-rhamnopyranosylcatalpol and
verbascoside: Cytotoxicity, cell cycle kinetics, apoptosis, and ROS
production evaluation in tumor cells. J Biochem Mol Toxicol.
34(e22443)2020.PubMed/NCBI View Article : Google Scholar
|
|
45
|
Wang L, Wang M, Dou H, Lin W and Zou L:
Sirtuin 1 inhibits lipopolysaccharide-induced inflammation in
chronic myelogenous leukemia k562 cells through interacting with
the Toll-like receptor 4-nuclear factor kappa B-reactive oxygen
species signaling axis. Cancer Cell Int. 20(73)2020.PubMed/NCBI View Article : Google Scholar
|
|
46
|
Speranza L, Franceschelli S, Pesce M,
Reale M, Menghini L, Vinciguerra I, De Lutiis MA, Felaco M and
Grilli A: Antiinflammatory effects in THP-1 cells treated with
verbascoside. Phytother Res. 24:1398–1404. 2010.PubMed/NCBI View Article : Google Scholar
|
|
47
|
Huang YH, Henriques ST, Wang CK,
Thorstholm L, Daly NL, Kaas Q and Craik DJ: Design of
substrate-based BCR-ABL kinase inhibitors using the cyclotide
scaffold. Sci Rep. 5(12974)2015.PubMed/NCBI View Article : Google Scholar
|
|
48
|
Trela E, Glowacki S and Błasiak J: Therapy
of chronic myeloid leukemia: Twilight of the imatinib era? ISRN
Oncol. 2014(596483)2014.PubMed/NCBI View Article : Google Scholar
|
|
49
|
Rossari F, Minutolo F and Orciuolo E:
Past, present, and future of Bcr-Abl inhibitors: From chemical
development to clinical efficacy. J Hematol Oncol.
11(84)2018.PubMed/NCBI View Article : Google Scholar
|
|
50
|
Aggul AG, Taslimi P, Kuzu M, Uzun N,
Bilginer S and Gulcin I: Oleuropein and verbascoside-their
inhibition effects on carbonic anhydrase and molecular docking
studies. J Oleo Sci. 70:1275–1283. 2021.PubMed/NCBI View Article : Google Scholar
|
|
51
|
Cheimonidi C, Samara P, Polychronopoulos
P, Tsakiri EN, Nikou T, Myrianthopoulos V, Sakellaropoulos T,
Zoumpourlis V, Mikros E, Papassideri I, et al: Selective
cytotoxicity of the herbal substance acteoside against tumor cells
and its mechanistic insights. Redox Biol. 16:169–178.
2018.PubMed/NCBI View Article : Google Scholar
|