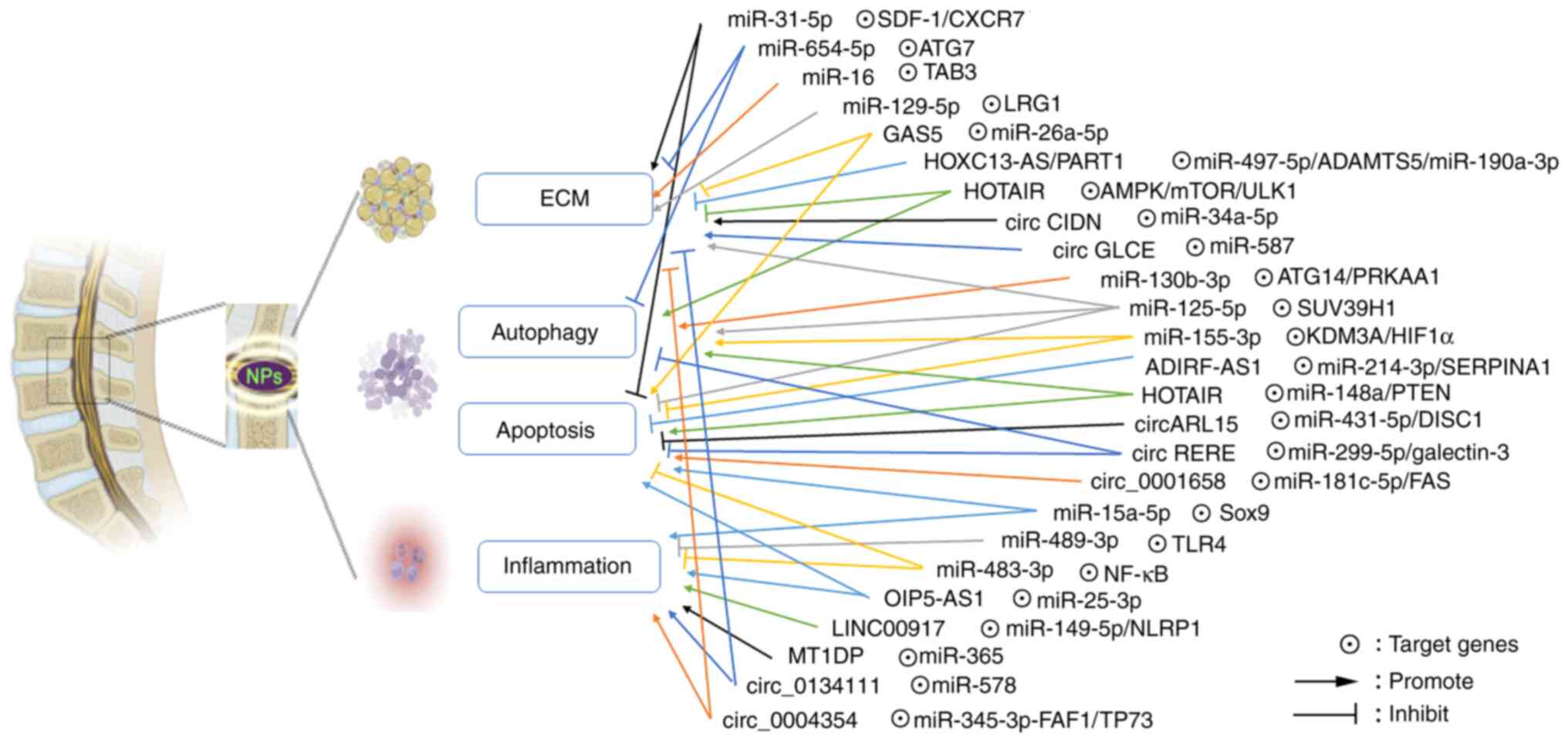
Intervertebral disc degeneration (IDD) is one of the
primary causes of lower back pain, which poses serious social and
economic burdens (1). The global
prevalence of IDD is >60%, which leads to high costs for society
(2). The pathogenesis of IDD is
complex and involves genetic, aging and environmental (3) and inflammatory factors (4). The gross anatomy of the ID primarily
comprises interdependent tissue compartments: The nucleus pulposus
(NP) is surrounded by a fibrocartilaginous annulus fibrosus
(5). The NP serves an essential
role in maintaining homeostasis of the ID and the key cellular
change during the development of IDD concerns centrally located NP
cells that undergo phenotypical transition, compromising the
structural integrity of discs (5).
Although numerous researchers have performed experimental studies
on the potential mechanism of IDD in recent years (6,7), the
underlying precise pathological mechanisms of IDD remain unclear.
Therefore, a further investigation of the molecular regulatory
mechanisms underlying the homeostasis of the ID may contribute to
identifying novel potential therapeutic targets for IDD.
A series of genetic and biological regulators are
associated with IDD pathogenesis and non-coding RNAs (ncRNAs) are
the critical factors (8,9). ncRNAs, primarily consisting of
microRNAs (miRNAs or miRs), long ncRNAs (lncRNAs) and circular RNAs
(circRNAs), are critical regulators in various cellular processes,
such as cell proliferation, autophagy and apoptosis. miRNAs, a
class of ncRNAs ~22 nucleotides in length, can recognize the
3'-untranslated regions of target mRNAs via complementary base
pairing and suppress gene expression at the post-transcriptional
level (10). lncRNA is a type of
ncRNA with a length >200 nucleotides that binds to proteins or
mRNAs using its nucleotide sequence or folded secondary structure
and regulates gene expression through multiple mechanisms at the
transcriptional and post-transcriptional levels (11). circRNA, primarily formed through
reverse splicing, is a type of endogenous covalently closed RNA
molecule that serves as a miRNA sponge to regulate the
miRNA-associated cell processes (12). It has been previously documented
that the level of aberrant ncRNAs is involved in various aspects of
IDD, including NP cell proliferation (13), autophagy (14), apoptosis (15), extracellular matrix (ECM)
regeneration (16) and
inflammatory response (17).
Therefore, dysregulation of ncRNAs in NP cells may be a crucial
pathological mechanism underlying the initiation and development of
IDD. Lan et al (18)
reported 76 pairs of differentially expressed circRNAs in IDD using
microarray datasets. Liu et al (19) documented that 636 circRNAs are
differentially expressed in IDD compared with normal controls.
lncRNA human leukocyte antigen complex group 18 (HCG18) is
upregulated in patients with IDD and luciferase reporter assays
have indicated that HCG18 may act as an endogenous sponge to
decrease expression of miR-146a-5p in NP cells, thus upregulating
the target gene of miR-146a-5p, decreasing the proliferation of NP
cells and finally resulting in cell apoptosis (20). Zhu et al (21) reported that circVMA21, serving as a
sponge for miR-200c, downregulates expression of X-linked inhibitor
of apoptosis (XIAP). Thus, decreased expression of XIAP in
inflammatory cytokine-treated NP cells is directly associated with
excessive cell apoptosis. Furthermore, intradiscal injection of
circVMA21 may alleviate IDD in a rat model (21). Accumulating evidence has shown the
key regulatory role of ncRNAs in IDD (22,23).
In addition, it has been reported that ncRNAs, targeted to IDs
using viruses and other vectors, can reverse the pathological
process of IDD and rescue ID function at the genetic level
(22). These findings indicate the
therapeutic potential of ncRNAs in IDD.
The present review aimed to summarize the latest
studies on ncRNAs involved in IDD pathology and clarify the key
roles of ncRNAs in the IDD process though regulating ECM
degradation, autophagy, apoptosis and inflammation, which could
provide a novel biotherapeutic strategy for clinical treatment of
IDD (Fig. 1).
Autophagy, a highly conserved biological phenomenon,
is the primary intracellular degradation process by which
cytoplasmic organelles, macromolecules and proteins are transported
and degraded in lysosomes, so it plays a critical role in
maintaining cellular homeostasis of NP in response to stressful
stimulation (52). Dysregulation
of autophagy may lead to human diseases such as obesity,
atherosclerosis, metabolic disorder and osteoporosis (53). The latest research indicates that
autophagy may serve a role in the pathophysiology of IDD (54). Emerging evidence has shown that
ncRNAs are essential for cellular functions of the NP, including
autophagy (54).
Normal apoptosis, which is autonomous programmed
cell death regulated by genes, can preserve the stability of the
intracellular environment (55).
The apoptosis of NP cells is a key pathological feature of IDD and
high rates of apoptosis are associated with degenerative processes
involving IDs, which reduce the number of cells in NP tissue,
damage ID structure and function and induce a disturbance of tissue
homeostasis (55). Degenerated IDs
exhibit higher rates of apoptosis, but the underlying mechanisms
remain unclear (55). Importantly,
evidence indicates that ncRNAs are involved in abnormal apoptosis
of NP cells during IDD (56). It
is reported that numerous ncRNAs can simultaneously affect the
process of apoptosis and autophagy during development of IDD
(57,58). Several studies have indicated that
autophagy alleviates IDD to protect NP cells from apoptosis and the
imbalance between autophagy and apoptosis is involved in IDD
progress (6,59) (Table
II).
miR-130b is involved in various pathophysiological
processes, such as regulation of cell proliferation and metastasis
in cancer as an oncogene (60,61),
affecting macrophage polarization (62) and modulation of
epithelial-mesenchymal crosstalk in pulmonary fibrosis (63). Recently, Wu et al (57) discovered that miR-130b-3p is
dysregulated in degenerative NP obtained from humans and rats and
that miR-130b-3p significantly influences IDD progression both
in vivo and in vitro. In addition, they found that
inhibiting miR-130b-3p protects NP cells from oxidative
stress-stimulated dysfunction and miR-130b-3p inhibition promotes
autophagy of NP cells. Mechanistically, they illustrated that
miR-130b-3p regulated autophagy-related 14 (ATG14) and protein
kinase AMP-activated catalytic subunit α 1 (PRKAA1) directly and
inhibited expression of ATG14 or PRKAA1 while the application of
the autophagy inhibitor (3-Methyladenine)suppressed autophagic flux
and attenuated the protective effects of miR-130b-3p inhibition in
tertbutyl hydro-peroxide (TBHP)-stimulated NP cells. Notably, IDD
was improved in a rat model by the injection of AAV-miR-130b-3p,
which inhibits miR-130b-3p (57).
These findings showed that by focusing on ATG14 and PRKAA1,
miR-130b-3p inhibition may increase autophagic flow and decrease
IDD, which suggested that miR-130b-3p may serve as a useful
treatment approach for IDD.
Exosomes are protein and nucleic acid-containing
membrane vesicles with a diameter of 40-100 nm that are released
from cells via exocytosis following fusion with the plasma membrane
(64). A recent report indicated
that exosomes serve a key role in NP apoptosis and autophagy
(65). Recently, Chen and Jiang
(66) found that cartilage
endplate stem cell-derived exosomes induce NP cell autophagy but
suppress apoptosis and ECM degradation. Furthermore, they
discovered that expression of miR-125-5p is significantly
upregulated in exosomes and miR-125-5p regulates suppressor of
variegation 3-9 homolog 1 (SUV39H1) to improve autophagy and
inhibit apoptosis and as ECM degradation in NP cells. They found
that miR-125-5p derived from cartilage endplate stem cell-derived
exosomes improves microtubule associated protein 1 light chain 3
(LC3)B expression in NP tissue of IDD rats, which suggested that
miR-125-5p may serve as a therapeutic target for IDD. In addition,
miR-125-5p could influence the inflammatory response and be
involved in the apoptosis of intestinal epithelial cells via the
Janus kinase 1/STAT3 and NF-κB pathways in ulcerative colitis
(67). miR-125a-5p was also
reported to downregulate TNF receptor superfamily member 1B gene
(TNFRSF1B) expression and improve osteoclast differentiation, which
indicated that miR-125a-5p is involved in osteoclast
differentiation (68).
Studies have shown that miR-155-3p, derived from
miR-155, is associated with IDD. Wang et al (69) documented that Fas-induced apoptosis
is promoted when miR-155 is suppressed and inhibited when miR-155
is upregulated in human NP cells. Zhang et al (70) reported that upregulation of miR-155
decreases IDD, while its suppression worsens IDD. Recently, Zhou
et al (71) revealed that
miR-155-3p is downregulated in NP tissues and cells of IDD. In
addition, upregulation of the expression of miR-155-3p or
suppression of lysine demethylase 3A (KDM3A) contribute to
autophagy and inhibit apoptosis of NP cells. Mechanistically,
miR-155-3p suppresses expression of KDM3A and Hypoxia-inducible
factor 1-α in NP cells from IDD samples, which may be a potential
approach for the treatment of IDD. According to a recent study,
miR-328-5p is abnormally expressed in numerous types of cancer
(colorectal cancer and lung cancer) and is associated with
apoptosis of cancer cells (72).
Yan et al (73) found that
the expression of miR-328-5p is upregulated significantly, while
the expression of its target gene WW domain containing E3 ubiquitin
protein ligase 2 (WWP2) is downregulated in IDD tissues compared
with normal tissues. miR-328-5p upregulation is positively, whereas
WWP2 downregulation is inversely, linked with the degeneration
grade of IDD. In addition, they identified that miR-328-5p
suppresses proliferation and causes apoptosis of NP cells via
regulation of the expression of Bcl-2, Bax and caspase-3.
Mechanistically, miR-328-5p may affect NP cell apoptosis by
targeting WWP2 and consequently be involved in the development of
IDD, which may provide a novel potential target for IDD
therapy.
lncRNAs are involved in the processes of apoptosis
and autophagy of NP cells in IDD (74). Growth arrest-specific transcript 5
(GAS5), which has been documented to serve as a tumor suppressor in
various types of cancer, is related to the process of
osteoarthritis as a pathogenic lncRNA (75). A previous study revealed the
ability of GAS5 to promote apoptosis of NP cells via targeting
miR-26-5p and suppression of GAS5 expression is a potential
therapeutic target for the treatment of IDD (39). In addition, GAS5 has also been
shown to target other miRNAs, such as miR-18a-5p and miR-17-5p
(39,76). Recently, Yu et al (58) found that lncRNA GAS5 is upregulated
in the NP tissue obtained from patients with IDD and it could
target miR-17-3p and affect Ang-2 expression. Furthermore, they
reported that downregulation of GAS5 suppresses levels of activated
caspase-3, activated caspase-7, activated caspase-9, MMP13 and
ADAMTS4, whereas GAS5 upregulates collagen II expression.
Importantly, they discovered that GAS5 suppression inhibits
apoptosis and ECM degradation in NP cells while promoting
proliferation via upregulating miR-17-3p. In addition, GAS5
inhibition or miR-17-3p upregulation relieves the degree of IDD in
mouse models, which demonstrates that inhibition of GAS5 suppresses
NP cell apoptosis and improves ECM remodeling, ultimately
alleviating IDD via miR-17-3p-dependent inhibition of Ang-2. These
results indicated that GAS5 could serve as a novel therapeutic
target for IDD treatment.
lncRNA adipogenesis regulatory factor-antisense RNA
1 (ADIRF-AS1), located at 10q23.2, is broadly expressed in numerous
types of cell, such as endothelial and epithelial cells.
Previously, it has been reported that expression of ADIRF-AS1 is
downregulated in more than 100 cancer cell lines (77). Xu et al (78) reported that ADIRF-AS1 is
upregulated in osteosarcoma (OS) and the overall survival of
patients with OS and high ADIRF-AS1 expression levels is shorter
than that of those with low ADIRF-AS1 expression levels.
Furthermore, ADIRF-AS1 suppression inhibits migration and
invasiveness of OS cells and promotes apoptosis. Mechanistically,
ADIRF-AS1 could target miR-761 and upregulate insulin receptor
substrate 1. Recently, Zhong et al (79) showed that ADIRF-AS1 expression is
inhibited in high-grade degenerated NP tissues and is positively
correlated with expression of serpin family A member 1 (SERPINA1).
In addition, ADIRF-AS1 upregulation decreases degenerative changes
in NP cells. miR-214-3p directly binds to SERPINA1 and ADIRF-AS1
and negatively regulates ADIRF-AS1 expression. Furthermore,
upregulation of ADIRF-AS1 relieves IDD via targeting miR-214-3p to
promote the expression of SERPINA1, which suggested that ADIRF-AS1
may be used as a potential target for IDD treatment.
HOTAIR has been reported to be involved in the
development of numerous types of cancer, such as breast and lung
cancer as well as hepatocellular carcinoma (80-82).
Wang et al (83) reported
that HOTAIR is involved in autophagy. Recently, HOTAIR was
documented to participate in progression of IDD by regulating NP
cell apoptosis, senescence and autophagy (44). Zhan et al (44) found that expression of HOTAIR is
positively correlated with IDD grade and its upregulation improves
autophagy. Further in vivo experiments demonstrated that
downregulation of HOTAIR ameliorates IDD in rat model of IDD, which
indicated that suppression of HOTAIR expression may serve as a
treatment for IDD. Zhang et al (84) reported that HOTAIR is significantly
upregulated in degenerative NP cells and its downregulation
suppresses degenerative NP cell apoptosis and autophagy. In
addition, they showed that HOTAIR increases expression of PTEN via
sponging miR-148a. In vivo assay confirmed that HOTAIR
suppression inhibits autophagy and apoptosis in ID tissues, which
ameliorates pathological injury in the IDD model.
Studies have reported that circRNAs, acting as
competing endogenous RNAs, are associated with pathogenesis of IDD
by affecting NP cell apoptosis and autophagy. Recently, Wang et
al (85) analyzed the dataset
GSE67567 from the Gene Expression Omnibus database and found that
the expression of circARL15 was significantly downregulated in IDD
tissues. They also found that circARL15 expression was negatively
associated with miR-431-5p and positively correlated with DISC1
scaffold protein (DISC1). In addition, they showed that circARL15
suppresses NP cell apoptosis but contributes to NP cell
proliferation via regulating the miR-431-5p/DISC1 signaling axis,
which suggested that circARL15 may serve as a biomarker and provide
a promising therapeutic target for patients with IDD.
Aggravation of inflammatory cytokine levels
contributes to IDD. Various studies have demonstrated upregulation
of the proinflammatory cytokines, such as Tumor necrosis factor
(TNF)-α, IL-6, IL-1β and IL-1α, in IDD (91,92).
These proinflammatory cytokines stimulate ECM degradation and
reformation of the phenotype of NP cells, thus causing degeneration
of the ID (93). Studies (Table III) have indicated that ncRNAs
are associated with production of inflammatory cytokines in NP
tissue.
Previously, OIP5 antisense RNA 1 (OIP5-AS1) was
reported to induce apoptosis of oxidized low-density lipoprotein
(ox-LDL)-stimulated vascular endothelial cells by regulating
glycogen synthase kinase 3 β (GSK-3β) via recruiting EZH2(97), while inhibition of OIP5-AS1
improves cell viability and suppresses apoptosis in
ox-LDL-stimulated human endothelial cells (98). Moreover, it was demonstrated that
lncRNA OIP5-AS1 serves an important role in suppressing osteoblast
differentiation of valve interstitial cells via the
miR-137/twist-related protein 11 signaling pathway (99). Further study indicated that lncRNA
OIP5-AS1 is associated with occurrence and development of
osteoarthritis and downregulation of lncRNA OIP5-AS1 worsens
osteoarthritis by regulating the miR-29b-3p/progranulin axis
(100). Recently, Che et
al (97) found that OIP5-AS1
is upregulated in IDD tissues and silencing of OIP5-AS1 promotes
cell proliferation but inhibits apoptosis and ECM degradation. In
addition, OIP5-AS1 suppression inhibits the inflammatory response
in LPS-induced NP cells. Mechanistically, OIP5-AS1 regulates
proliferation, apoptosis, inflammation and ECM degradation via
targeting miR-25-3p, which suggested that the OIP5-AS1/miR-25-3p
axis may be a potential target for IDD treatment. Previously, it
was reported that 135 lncRNAs are increased and 170 lncRNAs are
decreased in IDD samples, among which LINC00917 is the most
upregulated lncRNA and is hypothesized to regulate IDD progression
(101). Recently, Li et al
(102) found that LINC00917 is
significantly increased in TBHP-stimulated NP cells and inhibition
of LINC00917 improves proliferation and suppresses the inflammatory
response and pyroptosis of NP cells. Furthermore, they revealed
that LINC00917 silencing restores NP cellular function and inhibits
IDD progression by regulating the miR-149-5p/NLR family pyrin
domain containing 1 signaling pathway. Liao et al (103) reported that metallothionein 1D
pseudogene (MT1DP) and miR-365 are significantly upregulated in
human IDD NP tissue and NP cells, while the expression of nuclear
factor erythroid 2-related factor 2 (NRF-2) is significantly
downregulated. They found that upregulation of MT1DP and miR-365
and restriction of NRF-2 inhibit NP cell viability and cause
apoptosis and inflammation. Furthermore, MT1DP and miR-365 induce
inflammation in NP cells by damaging the mitochondrial membrane and
mitochondrial function.
circRNAs have been suggested as treatment targets
for several types of disease, such as cancer and metabolic and
cardiovascular diseases (104).
Liu and Zhang (105) reported
that expression of circ_0134111 is increased in IL-1β-activated
chondrocytes and knockdown of circ_0134111 reverses IL-1β-induced
cell decreases by suppressing apoptosis. In addition, they reported
that circ_0134111 causes osteoarthritis progression via targeting
miR-224-5p and regulating C-C motif chemokine ligand 1. Recently,
Yan et al (106) found
that circ_0134111 expression is significantly upregulated in IDD
tissue and the elevation of circ_0134111 is greater in severe IDD
cases. In addition, they discovered that IL-1β and TNF-α
significantly increase circ_0134111 expression in NP cells.
Furthermore, circ_0134111 overexpression increases proliferation,
inflammatory cytokine production and ECM degradation in NP cells.
circ_0134111 promotes progression of IDD by improving NP cell
inflammation and ECM degradation partly by regulating miR-578,
which suggested that circ_0134111 may serve as a potential target
for IDD treatment. Li et al (107) found that circ_0004354 competes
with circ_0040039 to stimulate the progression of IDD by regulating
miR-345-3p-FAF1/tumour protein P73 pathway-mediated apoptosis,
inflammatory response and ECM degradation of NP cells. These
results provide insight into the circ_0004354-mediated
post-transcriptional regulation of IDD, thus contributing to the
development of a promising treatment target for IDD in future.
Over the past few years, emerging studies have
indicated that a series of ncRNAs are involved in the progression
of IDD, particularly NP cell phenotypical regulation, which
provides insight into the pathogenesis of IDD (8,9).
Certain dysregulated ncRNAs may be useful diagnostic biomarkers and
therapeutic targets in the future with continued research (108). The present review discussed
progression of IDD, the role of miRNAs and lncRNAs and control of
downstream target genes to regulate ECM degradation, inflammation,
autophagy and apoptosis of NP cells. The present section discusses
the key regulatory role of ncRNAs in the intervention and treatment
of IDD from a macro perspective, as well as limitations of research
in clinical practice and potential future research directions.
ncRNAs have potential for investigation as
medications or biomarkers for the treatment of IDD according to the
findings of prior studies (108,109). However, to the best of our
knowledge, no clinical trials have been performed on ncRNAs in IDD
and all discussed studies have only used cell/animal experiments.
Nonetheless, the theoretical approach needs to translate into a
real course of action. Based on animal models, it is possible to
slow down or stop the progression of IDD by boosting, lowering or
eliminating the expression of one or more particular miRNAs,
lncRNAs and circRNAs in vivo (21,44).
To date, most studies have primarily focused on disease samples,
cells and small animals, which are dissimilar from the biomechanics
of humans (106,107). Therefore, there is an urgent need
for an improved animal model of the human spine. Additionally, the
regulatory mechanism of humans is complex. Thus, delivering
therapeutic ncRNAs to nearby degenerating IDs via a carrier appears
to be a workable treatment plan for IDD in the future (108).
Numerous obstacles still need to be overcome, such
as the fact that an ncRNA binding to its target gene is not
entirely complementary (44,84),
indicating that one ncRNA may control several targets or that one
target can be controlled by multiple ncRNAs. The first challenge to
be addressed in clinical application of ncRNAs will be improvement
of the specificity of ncRNAs toward the targets. Therefore, highly
selective ncRNAs that serve a role in numerous IDD-associated
processes may provide opportunities to prevent IDD. Thus, ncRNAs,
such as miRNAs, lncRNAs and circRNAs, as targets for the therapy of
IDD may become a unique research area with the development of
nanoparticle technology which could delivery ncRNAs and a thorough
understanding of the etiology of IDD. Several delivery methods have
been created to minimize off-target effects, particularly
concerning nanoparticles, which stand out due to their stability,
small size, biocompatibility and self-assembly properties (110,111). Thus, the potential of
nanoparticles as efficient ncRNA delivery systems is appealing and
merits further research.
Not applicable.
Funding: The present study was supported by the Key R&D
Project of Shandong Province (grant no. 2018GSF119011), the Natural
Science Foundation of Shandong Province (grant no. ZR2021MH254) and
the Project of Jinan Science and Technology Planning (grant no.
201821091).
Not applicable.
CG and YC wrote the manuscript. YW and YH edited the
manuscript. All authors have read and approved the final
manuscript. Data authentication is not applicable.
Not applicable.
Not applicable.
The authors declare that they have no competing
interests.
|
1
|
Maher C, Underwood M and Buchbinder R:
Non-specific low back pain. Lancet. 389:736–747. 2017.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Gianola S, Castellini G, Andreano A,
Corbetta D, Frigerio P, Pecoraro V, Redaelli V, Tettamanti A,
Turolla A, Moja L and Valsecchi MG: Effectiveness of treatments for
acute and sub-acute mechanical non-specific low back pain: Protocol
for a systematic review and network meta-analysis. Syst Rev.
8(196)2019.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Tessier S and Risbud MV: Understanding
embryonic development for cell-based therapies of intervertebral
disc degeneration: Toward an effort to treat disc degeneration
subphenotypes. Dev Dyn. 250:302–317. 2021.PubMed/NCBI View
Article : Google Scholar
|
|
4
|
Lyu FJ, Cui H, Pan H, Mc Cheung K, Cao X,
Iatridis JC and Zheng Z: Painful intervertebral disc degeneration
and inflammation: From laboratory evidence to clinical
interventions. Bone Res. 9(7)2021.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Ji ML, Jiang H, Zhang XJ, Shi PL, Li C, Wu
H, Wu XT, Wang YT, Wang C and Lu J: Preclinical development of a
microRNA-based therapy for intervertebral disc degeneration. Nat
Commun. 9(5051)2018.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Xu G, Liu C, Jiang J, Liang T, Yu C, Qin
Z, Zhang Z, Lu Z and Zhan X: A novel mechanism of intervertebral
disc degeneration: Imbalance between autophagy and apoptosis.
Epigenomics. 12:1095–1108. 2020.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Dong W, Liu J, Lv Y, Wang F, Liu T, Sun S,
Liao B, Shu Z and Qian J: miR-640 aggravates intervertebral disc
degeneration via NF-κB and WNT signalling pathway. Cell Prolif.
52(e12664)2019.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Memczak S, Jens M, Elefsinioti A, Torti F,
Krueger J, Rybak A, Maier L, Mackowiak SD, Gregersen LH, Munschauer
M, et al: Circular RNAs are a large class of animal RNAs with
regulatory potency. Nature. 495:333–338. 2013.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Mitra A, Pfeifer K and Park KS: Circular
RNAs and competing endogenous RNA (ceRNA) networks. Transl Cancer
Res. 7 (Suppl 5):S624–S628. 2018.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Kabekkodu SP, Shukla V, Varghese VK,
D'Souza J, Chakrabarty S and Satyamoorthy K: Clustered miRNAs and
their role in biological functions and diseases. Biol Rev Camb
Philos Soc. 93:1955–1986. 2018.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Bridges MC, Daulagala AC and Kourtidis A:
LNCcation: LncRNA localization and function. J Cell Biol.
220(e202009045)2021.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Zheng S, Zhang X, Odame E, Xu X, Chen Y,
Ye J, Zhou H, Dai D, Kyei B, Zhan S, et al: CircRNA-Protein
interactions in muscle development and diseases. Int J Mol Sci.
22(3262)2021.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Liu H, Huang X, Liu X, Xiao S, Zhang Y,
Xiang T, Shen X, Wang G and Sheng B: miR-21 promotes human nucleus
pulposus cell proliferation through PTEN/AKT signaling. Int J Mol
Sci. 15:4007–4018. 2014.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Xie L, Huang W, Fang Z, Ding F, Zou F, Ma
X, Tao J, Guo J, Xia X, Wang H, et al: CircERCC2 ameliorated
intervertebral disc degeneration by regulating mitophagy and
apoptosis through miR-182-5p/SIRT1 axis. Cell Death Dis.
10(751)2019.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Wang T, Li P, Ma X, Tian P, Han C, Zang J,
Kong J and Yan H: MicroRNA-494 inhibition protects nucleus pulposus
cells from TNF-α-induced apoptosis by targeting JunD. Biochimie.
115:1–7. 2015.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Jing W and Jiang W: MicroRNA-93 regulates
collagen loss by targeting MMP3 in human nucleus pulposus cells.
Cell Prolif. 48:284–292. 2015.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Gu SX, Li X, Hamilton JL, Chee A, Kc R,
Chen D, An HS, Kim JS, Oh CD, Ma YZ, et al: MicroRNA-146a reduces
IL-1 dependent inflammatory responses in the intervertebral disc.
Gene. 555:80–87. 2015.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Lan PH, Liu ZH, Pei YJ, Wu ZG, Yu Y, Yang
YF, Liu X, Che L, Ma CJ, Xie YK, et al: Landscape of RNAs in human
lumbar disc degeneration. Oncotarget. 7:63166–63176.
2016.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Liu X, Che L, Xie YK, Hu QJ, Ma CJ, Pei
YJ, Wu ZG, Liu ZH, Fan LY and Wang HQ: Noncoding RNAs in human
intervertebral disc degeneration: An integrated microarray study.
Genom Data. 5:80–81. 2015.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Xi Y, Jiang T, Wang W, Yu J, Wang Y, Wu X
and He Y: Long non-coding HCG18 promotes intervertebral disc
degeneration by sponging miR-146a-5p and regulating TRAF6
expression. Sci Rep. 7(13234)2017.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Zhu J, Zhang X, Gao W, Hu H, Wang X and
Hao D: lncRNA/circRNA-miRNA-mRNA ceRNA network in lumbar
intervertebral disc degeneration. Mol Med Rep. 20:3160–3174.
2019.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Fontana G, See E and Pandit A: Current
trends in biologics delivery to restore intervertebral disc
anabolism. Adv Drug Deliv Rev. 84:146–158. 2015.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Xu D, Ma X, Sun C, Han J, Zhou C, Wong SH,
Chan MTV and Wu WKK: Circular RNAs in intervertebral disc
degeneration: An updated review. Front Mol Biosci.
8(781424)2022.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Cazzanelli P and Wuertz-Kozak K: MicroRNAs
in intervertebral disc degeneration, apoptosis, inflammation, and
mechanobiology. Int J Mol Sci. 21(3601)2020.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Dowdell J, Erwin M, Choma T, Vaccaro A,
Iatridis J and Cho SK: Intervertebral disk degeneration and repair.
Neurosurgery. 80 (3S):S46–S54. 2017.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Li G, Ma L, He S, Luo R, Wang B, Zhang W,
Song Y, Liao Z, Ke W, Xiang Q, et al: WTAP-mediated m6A
modification of lncRNA NORAD promotes intervertebral disc
degeneration. Nat Commun. 13(1469)2022.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Zhou Y, Deng M, Su J, Zhang W, Liu D and
Wang Z: The role of miR-31-5p in the development of intervertebral
disc degeneration and its therapeutic potential. Front Cell Dev
Biol. 9(633974)2021.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Xie L, Chen Z, Liu M, Huang W, Zou F, Ma
X, Tao J, Guo J, Xia X, Lyu F, et al: MSC-Derived exosomes protect
vertebral endplate chondrocytes against apoptosis and calcification
via the miR-31-5p/ATF6 Axis. Mol Ther Nucleic Acids. 22:601–614.
2020.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Mi B, Li Q, Li T, Liu G and Sai J: High
miR-31-5p expression promotes colon adenocarcinoma progression by
targeting TNS1. Aging (Albany NY). 12:7480–7490. 2020.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Li P, Cai JX, Han F, Wang J, Zhou JJ, Shen
KW and Wang LH: Expression and significance of miR-654-5p and
miR-376b-3p in patients with colon cancer. World J Gastrointest
Oncol. 12:492–502. 2020.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Lu M, Wang C, Chen W, Mao C and Wang J:
miR-654-5p Targets GRAP to promote proliferation, metastasis, and
chemoresistance of oral squamous cell carcinoma through Ras/MAPK
signaling. DNA Cell Biol. 37:381–388. 2018.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Wang S, Guo Y, Zhang X and Wang C:
miR-654-5p inhibits autophagy by targeting ATG7 via mTOR signaling
in intervertebral disc degeneration. Mol Med Rep.
23(444)2021.PubMed/NCBI View Article : Google Scholar
|
|
33
|
Du K, He X and Deng J: MicroRNA-16
inhibits the lipopolysaccharide-induced inflammatory response in
nucleus pulposus cells of the intervertebral disc by targeting
TAB3. Arch Med Sci. 17:500–513. 2018.PubMed/NCBI View Article : Google Scholar
|
|
34
|
He J, Xue R, Li S, Lv J, Zhang Y, Fan L,
Teng Y and Wei H: Identification of the potential molecular targets
for human intervertebral disc degeneration based on bioinformatic
methods. Int J Mol Med. 36:1593–1600. 2015.PubMed/NCBI View Article : Google Scholar
|
|
35
|
Li N, Gao Q, Zhou W, Lv X, Yang X and Liu
X: MicroRNA-129-5p affects immune privilege and apoptosis of
nucleus pulposus cells via regulating FADD in intervertebral disc
degeneration. Cell Cycle. 19:933–948. 2020.PubMed/NCBI View Article : Google Scholar
|
|
36
|
Yang W and Sun P: Downregulation of
microRNA-129-5p increases the risk of intervertebral disc
degeneration by promoting the apoptosis of nucleus pulposus cells
via targeting BMP2. J Cell Biochem. 120:19684–19690.
2019.PubMed/NCBI View Article : Google Scholar
|
|
37
|
Zhou M, He SJ, Liu W, Yang MJ, Hou ZY,
Meng Q and Qian ZL: EZH2 upregulates the expression of MAPK1 to
promote intervertebral disc degeneration via suppression of
miR-129-5p. J Gene Med. 24(e3395)2022.PubMed/NCBI View Article : Google Scholar
|
|
38
|
Cui S and Zhang L: microRNA-129-5p
shuttled by mesenchymal stem cell-derived extracellular vesicles
alleviates intervertebral disc degeneration via blockade of
LRG1-mediated p38 MAPK activation. J Tissue Eng.
12(20417314211021679)2021.PubMed/NCBI View Article : Google Scholar
|
|
39
|
Tan L, Xie Y, Yuan Y and Hu K: LncRNA GAS5
as miR-26a-5p sponge regulates the PTEN/PI3K/Akt axis and affects
extracellular matrix synthesis in degenerative nucleus pulposus
cells in vitro. Front Neurol. 12(653341)2021.PubMed/NCBI View Article : Google Scholar
|
|
40
|
Jing W and Liu W: HOXC13-AS induced
extracellular matrix loss via targeting miR-497-5p/ADAMTS5 in
intervertebral disc. Front Mol Biosci. 8(643997)2021.PubMed/NCBI View Article : Google Scholar
|
|
41
|
Bao X, Ren T, Huang Y, Sun K, Wang S, Liu
K, Zheng B and Guo W: Knockdown of long non-coding RNA HOTAIR
increases miR-454-3p by targeting Stat3 and Atg12 to inhibit
chondrosarcoma growth. Cell Death Dis. 8(e2605)2017.PubMed/NCBI View Article : Google Scholar
|
|
42
|
Li E, Zhao Z, Ma B and Zhang J: Long
noncoding RNA HOTAIR promotes the proliferation and metastasis of
osteosarcoma cells through the AKT/mTOR signaling pathway. Exp Ther
Med. 14:5321–5328. 2017.PubMed/NCBI View Article : Google Scholar
|
|
43
|
Mercer TR, Dinger ME and Mattick JS: Long
non-coding RNAs: Insights into functions. Nat Rev Genet.
10:155–159. 2009.PubMed/NCBI View Article : Google Scholar
|
|
44
|
Zhan S, Wang K, Xiang Q, Song Y, Li S,
Liang H, Luo R, Wang B, Liao Z, Zhang Y and Yang C: lncRNA HOTAIR
upregulates autophagy to promote apoptosis and senescence of
nucleus pulposus cells. J Cell Physiol. 235:2195–2208.
2020.PubMed/NCBI View Article : Google Scholar
|
|
45
|
Zhan S, Wang K, Song Y, Li S, Yin H, Luo
R, Liao Z, Wu X, Zhang Y and Yang C: Long non-coding RNA HOTAIR
modulates intervertebral disc degenerative changes via
Wnt/β-catenin pathway. Arthritis Res Ther. 21(201)2019.PubMed/NCBI View Article : Google Scholar
|
|
46
|
Zhu D, Yu Y, Wang W, Wu K, Liu D, Yang Y,
Zhang C, Qi Y and Zhao S: Long noncoding RNA PART1 promotes
progression of non-small cell lung cancer cells via JAK-STAT
signaling pathway. Cancer Med. 8:6064–6081. 2019.PubMed/NCBI View Article : Google Scholar
|
|
47
|
Zhou T, Wu L, Ma N, Tang F, Zong Z and
Chen S: LncRNA PART1 regulates colorectal cancer via targeting
miR-150-5p/miR-520h/CTNNB1 and activating Wnt/β-catenin pathway.
Int J Biochem Cell Biol. 118(105637)2020.PubMed/NCBI View Article : Google Scholar
|
|
48
|
Zhao B, Lu M, Wang D, Li H and He X:
Genome-Wide identification of long noncoding RNAs in human
intervertebral disc degeneration by RNA sequencing. Biomed Res Int.
2016(3684875)2016.PubMed/NCBI View Article : Google Scholar
|
|
49
|
Zhang Z, Huo Y, Zhou Z, Zhang P and Hu J:
Role of lncRNA PART1 in intervertebral disc degeneration and
associated underlying mechanism. Exp Ther Med.
21(131)2021.PubMed/NCBI View Article : Google Scholar
|
|
50
|
Xiang Q, Kang L, Wang J, Liao Z, Song Y,
Zhao K, Wang K, Yang C and Zhang Y: CircRNA-CIDN mitigated
compression loading-induced damage in human nucleus pulposus cells
via miR-34a-5p/SIRT1 axis. EBioMedicine. 53(102679)2020.PubMed/NCBI View Article : Google Scholar
|
|
51
|
Chen Z, Zhang W, Deng M, Li Y and Zhou Y:
CircGLCE alleviates intervertebral disc degeneration by regulating
apoptosis and matrix degradation through the targeting of
miR-587/STAP1. Aging (Albany NY). 12:21971–21991. 2020.PubMed/NCBI View Article : Google Scholar
|
|
52
|
Mizushima N and Komatsu M: Autophagy:
Renovation of cells and tissues. Cell. 147:728–741. 2011.PubMed/NCBI View Article : Google Scholar
|
|
53
|
Kim KH and Lee MS: Autophagy-a key player
in cellular and body metabolism. Nat Rev Endocrinol. 10:322–337.
2014.PubMed/NCBI View Article : Google Scholar
|
|
54
|
Kritschil R, Scott M, Sowa G and Vo N:
Role of autophagy in intervertebral disc degeneration. J Cell
Physiol. 237:1266–1284. 2022.PubMed/NCBI View Article : Google Scholar
|
|
55
|
Wang F, Cai F, Shi R, Wang XH and Wu XT:
Aging and age related stresses: A senescence mechanism of
intervertebral disc degeneration. Osteoarthritis Cartilage.
24:398–408. 2016.PubMed/NCBI View Article : Google Scholar
|
|
56
|
Li H, Tian L, Li J, Li Y, Du L, Huo Z and
Xu B: The roles of circRNAs in intervertebral disc degeneration:
Inflammation, extracellular matrix metabolism, and apoptosis. Anal
Cell Pathol (Amst). 2022(9550499)2022.PubMed/NCBI View Article : Google Scholar
|
|
57
|
Wu T, Jia X, Zhu Z, Guo K, Wang Q, Gao Z,
Li X, Huang Y and Wu D: Inhibition of miR-130b-3p restores
autophagy and attenuates intervertebral disc degeneration through
mediating ATG14 and PRKAA1. Apoptosis. 27:409–425. 2022.PubMed/NCBI View Article : Google Scholar
|
|
58
|
Yu X, Liu Q, Wang Y, Bao Y, Jiang Y, Li M,
Li Z, Wang B, Yu L, Wang S, et al: Depleted Long Noncoding RNA GAS5
relieves intervertebral disc degeneration via microRNA-17-3p/Ang-2.
Oxid Med Cell Longev. 2022(1792412)2022.PubMed/NCBI View Article : Google Scholar
|
|
59
|
Wen F, Yu J, He CJ, Zhang ZW and Yang AF:
β-ecdysterone protects against apoptosis by promoting autophagy in
nucleus pulposus cells and ameliorates disc degeneration. Mol Med
Rep. 19:2440–2448. 2019.PubMed/NCBI View Article : Google Scholar
|
|
60
|
Yu X, Wang ZL, Han CL, Wang MW, Jin Y, Jin
XB and Xia QH: LncRNA CASC15 functions as an oncogene by sponging
miR-130b-3p in bladder cancer. Eur Rev Med Pharmacol Sci.
24(7203)2020.PubMed/NCBI View Article : Google Scholar
|
|
61
|
Liao Y, Wang C, Yang Z, Liu W, Yuan Y, Li
K, Zhang Y, Wang Y, Shi Y, Qiu Y, et al: Dysregulated
Sp1/miR-130b-3p/HOXA5 axis contributes to tumor angiogenesis and
progression of hepatocellular carcinoma. Theranostics.
10:5209–5224. 2020.PubMed/NCBI View Article : Google Scholar
|
|
62
|
Guo Q, Zhu X, Wei R, Zhao L, Zhang Z, Yin
X, Zhang Y, Chu C, Wang B and Li X: miR-130b-3p regulates M1
macrophage polarization via targeting IRF1. J Cell Physiol.
236:2008–2022. 2021.PubMed/NCBI View Article : Google Scholar
|
|
63
|
Li S, Geng J, Xu X, Huang X, Leng D, Jiang
D, Liang J, Wang C, Jiang D and Dai H: miR-130b-3p modulates
epithelial-mesenchymal crosstalk in lung fibrosis by targeting
IGF-1. PLoS One. 11(e0150418)2016.PubMed/NCBI View Article : Google Scholar
|
|
64
|
Kalluri R and LeBleu VS: The biology,
function, and biomedical applications of exosomes. Science.
367(eaau6977)2020.PubMed/NCBI View Article : Google Scholar
|
|
65
|
Luo L, Jian X, Sun H, Qin J, Wang Y, Zhang
J, Shen Z, Yang D, Li C, Zhao P, et al: Cartilage endplate stem
cells inhibit intervertebral disc degeneration by releasing
exosomes to nucleus pulposus cells to activate Akt/autophagy. Stem
Cells. 39:467–481. 2021.PubMed/NCBI View Article : Google Scholar
|
|
66
|
Chen D and Jiang X: Exosomes-derived
miR-125-5p from cartilage endplate stem cells regulates autophagy
and ECM metabolism in nucleus pulposus by targeting SUV38H1. Exp
Cell Res. 414(113066)2022.PubMed/NCBI View Article : Google Scholar
|
|
67
|
Yao D, Zhou Z, Wang P, Zheng L, Huang Y,
Duan Y, Liu B and Li Y: MiR-125-5p/IL-6R axis regulates macrophage
inflammatory response and intestinal epithelial cell apoptosis in
ulcerative colitis through JAK1/STAT3 and NF-κB pathway. Cell
Cycle. 20:2547–2564. 2021.PubMed/NCBI View Article : Google Scholar
|
|
68
|
Sun L, Lian JX and Meng S: MiR-125a-5p
promotes osteoclastogenesis by targeting TNFRSF1B. Cell Mol Biol
Lett. 24(23)2019.PubMed/NCBI View Article : Google Scholar
|
|
69
|
Wang HQ, Yu XD, Liu ZH, Cheng X, Samartzis
D, Jia LT, Wu SX, Huang J, Chen J and Luo ZJ: Deregulated miR-155
promotes Fas-mediated apoptosis in human intervertebral disc
degeneration by targeting FADD and caspase-3. J Pathol.
225:232–242. 2011.PubMed/NCBI View Article : Google Scholar
|
|
70
|
Zhang WL, Chen YF, Meng HZ, Du JJ, Luan
GN, Wang HQ, Yang MW and Luo ZJ: Role of miR-155 in the regulation
of MMP-16 expression in intervertebral disc degeneration. J Orthop
Res. 35:1323–1334. 2017.PubMed/NCBI View Article : Google Scholar
|
|
71
|
Zhou X, Li J, Teng J, Liu Y, Zhang D, Liu
L and Zhang W: microRNA-155-3p attenuates intervertebral disc
degeneration via inhibition of KDM3A and HIF1α. Inflamm Res.
70:297–308. 2021.PubMed/NCBI View Article : Google Scholar
|
|
72
|
Liu Z, Xu L, Zhang K, Guo B, Cui Z and Gao
N: LINC00210 plays oncogenic roles in non-small cell lung cancer by
sponging microRNA-328-5p. Exp Ther Med. 19:3325–3331.
2020.PubMed/NCBI View Article : Google Scholar
|
|
73
|
Yan J, Wu LG, Zhang M, Fang T, Pan W, Zhao
JL and Zhou Q: miR-328-5p induces human intervertebral disc
degeneration by targeting WWP2. Oxid Med Cell Longev.
2022(3511967)2022.PubMed/NCBI View Article : Google Scholar
|
|
74
|
Li Z, Li X, Chen C, Li S, Shen J, Tse G,
Chan MTV and Wu WKK: Long non-coding RNAs in nucleus pulposus cell
function and intervertebral disc degeneration. Cell Prolif.
51(e12483)2018.PubMed/NCBI View Article : Google Scholar
|
|
75
|
Mayama T, Marr AK and Kino T: Differential
expression of glucocorticoid receptor noncoding RNA Repressor Gas5
in autoimmune and inflammatory diseases. Horm Metab Res.
48:550–557. 2016.PubMed/NCBI View Article : Google Scholar
|
|
76
|
Kolenda T, Guglas K, Kopczyńska M,
Sobocińska J, Teresiak A, Bliźniak R and Lamperska K: Good or not
good: Role of miR-18a in cancer biology. Rep Pract Oncol Radiother.
25:808–819. 2020.PubMed/NCBI View Article : Google Scholar
|
|
77
|
Klijn C, Durinck S, Stawiski EW, Haverty
PM, Jiang Z, Liu H, Degenhardt J, Mayba O, Gnad F, Liu J, et al: A
comprehensive transcriptional portrait of human cancer cell lines.
Nat Biotechnol. 33:306–312. 2015.PubMed/NCBI View Article : Google Scholar
|
|
78
|
Xu L, Tan Y, Xu F and Zhang Y: Long
noncoding RNA ADIRF antisense RNA 1 upregulates insulin receptor
substrate 1 to decrease the aggressiveness of osteosarcoma by
sponging microRNA-761. Bioengineered. 13:2028–2043. 2022.PubMed/NCBI View Article : Google Scholar
|
|
79
|
Zhong H, Zhou Z, Guo L, Liu FS, Wang X, Li
J, Lv GH and Zou MX: SERPINA1 is a hub gene associated with
intervertebral disc degeneration grade and affects the nucleus
pulposus cell phenotype through the ADIRF-AS1/miR-214-3p axis.
Transl Res. 245:99–116. 2022.PubMed/NCBI View Article : Google Scholar
|
|
80
|
Zhao W, Geng D, Li S, Chen Z and Sun M:
LncRNA HOTAIR influences cell growth, migration, invasion, and
apoptosis via the miR-20a-5p/HMGA2 axis in breast cancer. Cancer
Med. 7:842–855. 2018.PubMed/NCBI View Article : Google Scholar
|
|
81
|
Loewen G, Jayawickramarajah J, Zhuo Y and
Shan B: Functions of lncRNA HOTAIR in lung cancer. J Hematol Oncol.
7(90)2014.PubMed/NCBI View Article : Google Scholar
|
|
82
|
Yang L, Peng X, Li Y, Zhang X, Ma Y, Wu C,
Fan Q, Wei S, Li H and Liu J: Long non-coding RNA HOTAIR promotes
exosome secretion by regulating RAB35 and SNAP23 in hepatocellular
carcinoma. Mol Cancer. 18(78)2019.PubMed/NCBI View Article : Google Scholar
|
|
83
|
Wang X, Liu W, Wang P and Li S: RNA
interference of long noncoding RNA HOTAIR suppresses autophagy and
promotes apoptosis and sensitivity to cisplatin in oral squamous
cell carcinoma. J Oral Pathol Med. 47:930–937. 2018.PubMed/NCBI View Article : Google Scholar
|
|
84
|
Zhang S, Song S, Cui W, Liu X and Sun Z:
Mechanism of long Noncoding RNA HOTAIR in nucleus pulposus cell
autophagy and apoptosis in intervertebral disc degeneration. Evid
Based Complement Alternat Med. 2022(8504601)2022.PubMed/NCBI View Article : Google Scholar
|
|
85
|
Wang H, Zhu Y, Cao L, Guo Z, Sun K, Qiu W
and Fan H: circARL15 plays a critical role in intervertebral disc
degeneration by modulating miR-431-5p/DISC1. Front Genet.
12(669598)2021.PubMed/NCBI View Article : Google Scholar
|
|
86
|
Wang R, Zhou X, Luo G, Zhang J, Yang M and
Song C: CircRNA RERE Promotes the oxidative stress-induced
apoptosis and autophagy of nucleus pulposus cells through the
miR-299-5p/Galectin-3 Axis. J Healthc Eng.
2021(2771712)2021.PubMed/NCBI View Article : Google Scholar
|
|
87
|
Liu Y, Yang Y, Lin Y, Wei B, Hu X, Xu L,
Zhang W and Lu J: N6 -methyladenosine-modified circRNA
RERE modulates osteoarthritis by regulating β-catenin
ubiquitination and degradation. Cell Prolif: Jun 22, 2022 (Epub
ahead of print).
|
|
88
|
Wang L, Wang P, Su X and Zhao B:
Circ_0001658 promotes the proliferation and metastasis of
osteosarcoma cells via regulating miR-382-5p/YB-1 axis. Cell
Biochem Funct. 38:77–86. 2020.PubMed/NCBI View Article : Google Scholar
|
|
89
|
Meng GD and Xu BS: Circular RNA
hsa_circ_0001658 inhibits intervertebral disc degeneration
development by regulating hsa-miR-181c-5p/FAS. Comput Math Methods
Med. 2021(7853335)2021.PubMed/NCBI View Article : Google Scholar
|
|
90
|
Duan X, Yu X and Li Z: Circular RNA
hsa_circ_0001658 regulates apoptosis and autophagy in gastric
cancer through microRNA-182/Ras-related protein Rab-10 signaling
axis. Bioengineered. 13:2387–2397. 2022.PubMed/NCBI View Article : Google Scholar
|
|
91
|
Cosamalón-Gan I, Cosamalón-Gan T,
Mattos-Piaggio G, Villar-Suárez V, García-Cosamalón J and
Vega-Álvarez JA: Inflammation in the intervertebral disc
herniation. Neurocirugia (Astur: Engl Ed). 32:21–35.
2021.PubMed/NCBI View Article : Google Scholar
|
|
92
|
Le Maitre CL, Hoyland JA and Freemont AJ:
Catabolic cytokine expression in degenerate and herniated human
intervertebral discs: IL-1beta and TNFalpha expression profile.
Arthritis Res Ther. 9(R77)2007.PubMed/NCBI View
Article : Google Scholar
|
|
93
|
Risbud MV and Shapiro IM: Role of
cytokines in intervertebral disc degeneration: Pain and disc
content. Nat Rev Rheumatol. 10:44–56. 2014.PubMed/NCBI View Article : Google Scholar
|
|
94
|
Zhang S, Song S, Zhuang Y, Hu J, Cui W,
Wang X, Zhao Z, Liu X and Sun Z: Role of microRNA-15a-5p/Sox9/NF-κB
axis in inflammatory factors and apoptosis of murine nucleus
pulposus cells in intervertebral disc degeneration. Life Sci.
277(119408)2021.PubMed/NCBI View Article : Google Scholar
|
|
95
|
Dong L and Dong B: miR-489-3p
overexpression inhibits lipopolysaccharide-induced nucleus pulposus
cell apoptosis, inflammation and extracellular matrix degradation
via targeting Toll-like receptor 4. Exp Ther Med.
22(1323)2021.PubMed/NCBI View Article : Google Scholar
|
|
96
|
Ji Z, Guo R, Ma Z and Li H: Arctigenin
inhibits apoptosis, extracellular matrix degradation, and
inflammation in human nucleus pulposus cells by up-regulating
miR-483-3p. J Clin Lab Anal. 36(e24508)2022.PubMed/NCBI View Article : Google Scholar
|
|
97
|
Che Z, Xueqin J and Zhang Z: LncRNA
OIP5-AS1 accelerates intervertebral disc degeneration by targeting
miR-25-3p. Bioengineered. 12:11201–11212. 2021.PubMed/NCBI View Article : Google Scholar
|
|
98
|
Zhang C, Yang H, Li Y, Huo P and Ma P:
LNCRNA OIP5-AS1 regulates oxidative low-density
lipoprotein-mediated endothelial cell injury via miR-320a/LOX1
axis. Mol Cell Biochem. 467:15–25. 2020.PubMed/NCBI View Article : Google Scholar
|
|
99
|
Zheng D, Wang B, Zhu X, Hu J, Sun J, Xuan
J and Ge Z: LncRNA OIP5-AS1 inhibits osteoblast differentiation of
valve interstitial cells via miR-137/TWIST11 axis. Biochem Biophys
Res Commun. 511:826–832. 2019.PubMed/NCBI View Article : Google Scholar
|
|
100
|
Zhi L, Zhao J, Zhao H, Qing Z, Liu H and
Ma J: Downregulation of LncRNA OIP5-AS1 Induced by IL-1β aggravates
osteoarthritis via regulating miR-29b-3p/PGRN. Cartilage. 13
(2_suppl):1345S–1355S. 2021.PubMed/NCBI View Article : Google Scholar
|
|
101
|
Chen Y, Ni H, Zhao Y, Chen K, Li M, Li C,
Zhu X and Fu Q: Potential role of lncRNAs in contributing to
pathogenesis of intervertebral disc degeneration based on
microarray data. Med Sci Monit. 21:3449–3458. 2015.PubMed/NCBI View Article : Google Scholar
|
|
102
|
Li T, Peng Y, Chen Y, Huang X, Li X, Zhang
Z and Du J: Long intergenic non-coding RNA-00917 regulates the
proliferation, inflammation, and pyroptosis of nucleus pulposus
cells via targeting miR-149-5p/NOD-like receptor protein 1 axis.
Bioengineered. 13:6036–6047. 2022.PubMed/NCBI View Article : Google Scholar
|
|
103
|
Liao ZW, Fan ZW, Huang Y, Liang CY, Liu C,
Huang S and Chen CW: Long non-coding RNA MT1DP interacts with
miR-365 and induces apoptosis of nucleus pulposus cells by
repressing NRF-2-induced anti-oxidation in lumbar disc herniation.
Ann Transl Med. 9(151)2021.PubMed/NCBI View Article : Google Scholar
|
|
104
|
Huang JG, Tang X, Wang JJ, Liu J, Chen P
and Sun Y: A circular RNA, circUSP36, accelerates endothelial cell
dysfunction in atherosclerosis by adsorbing miR-637 to enhance WNT4
expression. Bioengineered. 12:6759–6770. 2021.PubMed/NCBI View Article : Google Scholar
|
|
105
|
Liu Y and Zhang Y: Hsa_circ_0134111
promotes osteoarthritis progression by regulating miR-224-5p/CCL1
interaction. Aging (Albany NY). 13:20383–20394. 2021.PubMed/NCBI View Article : Google Scholar
|
|
106
|
Yan P, Sun C, Luan L, Han J, Qu Y, Zhou C
and Xu D: Hsa_circ_0134111 promotes intervertebral disc
degeneration via sponging miR-578. Cell Death Discov.
8(55)2022.PubMed/NCBI View Article : Google Scholar
|
|
107
|
Li Y, Wu X, Li J, Du L, Wang X, Cao J, Li
H, Huo Z, Li G, Pan D, et al: Circ_0004354 might compete with
circ_0040039 to induce NPCs death and inflammatory response by
targeting miR-345-3p-FAF1/TP73 axis in intervertebral disc
degeneration. Oxid Med Cell Longev. 2022(2776440)2022.PubMed/NCBI View Article : Google Scholar
|
|
108
|
Xin J, Wang Y, Zheng Z, Wang S, Na S and
Zhang S: Treatment of intervertebral disc degeneration. Orthop
Surg. 14:1271–1280. 2022.PubMed/NCBI View Article : Google Scholar
|
|
109
|
Guo HY, Guo MK, Wan ZY, Song F and Wang
HQ: Emerging evidence on noncoding-RNA regulatory machinery in
intervertebral disc degeneration: A narrative review. Arthritis Res
Ther. 22(270)2020.PubMed/NCBI View Article : Google Scholar
|
|
110
|
Fekrazad R, Naghdi N, Nokhbatolfoghahaei H
and Bagheri H: The combination of laser therapy and metal
nanoparticles in cancer treatment originated from epithelial
tissues: A literature review. J Lasers Med Sci. 7:62–75.
2016.PubMed/NCBI View Article : Google Scholar
|
|
111
|
Zhao K, Li D, Shi C, Ma X, Rong G, Kang H,
Wang X and Sun B: Biodegradable polymeric nanoparticles as the
delivery carrier for drug. Curr Drug Deliv. 13:494–499.
2016.PubMed/NCBI View Article : Google Scholar
|