Introduction
Breast cancer (BC) is the most common cancer in
women globally (1). However, the
early stages of BC are considered curable (2); therefore, early screening and
diagnostic marker detection are critical for effective BC
treatment. As BC progresses, the growth of cancer cells accelerates
and causes distant migration, which greatly shortens the survival
expectancy of BC patients and increases the difficulty of
establishing effective treatment strategies (3). Given the current limited treatment
options, new drugs are urgently required to treat BC. The primary
aim of this study was to explore the therapeutic effect and
internal mechanism of scoparone (SCO; 6,7-dimethoxycoumarin) in
BC.
Traditional Chinese medicine (TCM) has the
remarkable ability to reduce the side effects of chemotherapy
alongside being used to prevent and treat cancer (4). The anticancer effects of Chinese
herbal medicines have also been verified against BC. For example,
plantamajoside can inhibit BC cell growth and pulmonary metastasis
by decreasing the activity of matrix metalloproteinase-9 and
-2(5). Liew et al (6) determined that treatment with Chinese
herbal medicine during chemotherapy reduced fatigue, nausea, and
anorexia in BC patients; therefore, TCM can improve quality of life
in cancer patients. SCO is the primary component of the Chinese
herb Artemisia capillaris; this component specifically has
antioxidant, lipid-lowering, and anti-inflammatory effects
(7,8). Recent studies have demonstrated that
scopolamine (SCO) can play a significant role in cancer treatment.
For example, SCO shows significant anti-tumor effects against
prostate cancer cells by inhibiting the activity of signal
transducer and activator of transcription 3(9). SCO also reduces migration and induces
apoptosis of laryngeal cancer cells in a dose-dependent manner
(10). However, no systematic
study has been conducted on the potential anti-tumor effect of SCO
in BC.
Long noncoding RNAs (lncRNAs) are important players
in the progression of BC (11),
and their dysregulation may alter intercellular signal transduction
and affect the growth of BC cells (12). In addition, lncRNAs can act as
competing endogenous RNA (ceRNAs) that regulate the transcriptional
activity and translation level of downstream mRNA by adsorbing
microRNAs (miRNAs/miRs). For example, LINC00673 acts as a
ceRNA that adsorbs miR-515-5p to regulate microtubule
affinity-regulating kinase 4, thereby affecting the transduction of
Hippo signaling and the growth of BC cells (13). Small nucleolar RNA host gene 12
(SNHG12) promotes cell proliferation, migration, and
invasion, and inhibits BC cell apoptosis through the
miR-15a-5p/spalt-like transcription factor 4 axis (14). In addition, miRNAs regulate the
transcriptional activity and translation level of downstream mRNA
by binding to the 3'-untranslated region (UTR) of mRNA. For
example, the expression of miR-140-3p is reduced in BC, which
promotes the expression of tripartite motif containing 28 and
accelerates the viability, migration, and invasion of BC cells
(15). Additionally, tumor
necrosis factor (TNF) receptor-associated factor 2 (TRAF2)
is upregulated in BC; nonetheless, miR-502-5p can bind to the
3'-UTR of TRAF2, thereby inhibiting TRAF2 and
reducing the progression of BC (16). TRAF2 also promotes the
proliferation and metastasis of BC cells through the nuclear factor
κB (NF-κB) pathway by directly interacting with various TNF
receptors (17,18).
Overall, this study aims to investigate the effect
and mechanism of SCO on BC cells and provides a theoretical basis
for its potential clinical application in BC treatment.
Materials and methods
Cell culture and treatment
All cell lines used in this study were obtained from
the Procell (Wuhan, China). These cell lines included the normal
human mammary epithelial cell line MCF-10A (culture conditions:
Dulbecco's modified eagle medium [DMEM]/nutrient mixture F-12
containing 5% horse serum) and the human BC cell lines MCF-7
(culture conditions: minimum essential medium containing 10% fetal
bovine serum [FBS]), MDA-MB-231 (culture conditions: DMEM
containing 5% FBS), ZR-75-1 (culture conditions: Roswell Park
Memorial Institute (RPMI)-1640 medium containing 10% FBS), HCC1937
(culture conditions: RPMI-1640 containing 10% FBS), and HEK293
(culture conditions: DMEM containing 10% FBS). All cells were
seeded in 6-well plates at a density of 2x105; gradient
concentrations of SCO (0, 100, 200, 500, and 1,000 µM) were
purchased from Sigma-Aldrich (St. Louis, MO, USA) and used to treat
cells for 0, 24, 48, or 72 h.
Cell transfection
Initially, ov-SNHG12 (50 nM; pcDNA3.1),
miR-140-3p mimic/inhibitor (100 nM), and three
TRAF2-targeting small interfering RNAs (si-TRAF2) (50
nM) were transfected into MCF-7 and MDA-MB-231 cells (density of
3x105) using Lipofectamine 3000 (Invitrogen; Thermo
Fisher Scientific, Inc., Massachusetts, USA), according to the
manufacturer's instructions. ov-SNHG12, miR-140-3p
mimic/inhibitor, si-TRAF2, and their negative controls (NCs)
were purchased from Sangon Biotech (Shanghai, China); the
corresponding sequences are listed in Table SI.
Cell viability assay
MCF-10A and BC cells were seeded in 96-well plates
at a density of 3x103 cells/well and cultured for 12 h.
Optical density values were measured at 450 nm for 60 min at 37˚C;
samples were taken at 24 h intervals across 72 h and measurements
were conducted using a Cell Counting Kit-8 (CCK8; Beijing Solarbio
Science & Technology Co., Ltd., Beijing, China) and an enzyme
labeling instrument (Thermo Fisher Scientific).
Nucleocytoplasmic separation and
reverse transcription-quantitative PCR (RT-qPCR) assay
The Cytoplasmic and Nuclear RNA Purification Kit
(Norgen Biotek, Ontario, Canada) was used to isolate RNA from the
cells according to the manufacturer's instructions; total RNA was
extracted from BC cells using TRIzol (Invitrogen). After
centrifugation at 12,000 x g (4˚C, 10 min) in a high-speed
refrigerated centrifuge (JIDI-17RS; Guangzhou JiDi Instrument Co.,
Ltd., Guangzhou, China), the precipitate was resuspended in diethyl
pyrocarbonate water (Invitrogen); aliquots of these precipitates
were then reverse-transcribed to complementary DNA using a
PrimeScript RT kit (Takara Bio, Kyoto, Japan). The SYBR Premix Ex
Taq II kit (Invitrogen) was used to perform RT-qPCR analysis using
an ABI 7500 Real-Time PCR system (Applied Biosystems, California,
USA). PCR amplification was performed under the following
conditions: 95˚C for an initial 30 sec, followed by 40 cycles at
95˚ for 5 sec and 60˚C for 30 sec. The sequences of the primer
pairs used for amplification are listed in Table SII. SNHG12, miR-140-3p, and
TRAF2 RNA levels were normalized to those of the
housekeeping genes β-actin or U6 and were determined using
the 2-ΔΔCq method (19).
RNA pull-down assay
Biotin-labeled SNHG12 and miR-140-3p were
transfected into BC cells. After 24 h of transfection, cells were
lysed with RIPA lysis buffer and incubated with Dynabeads M-280
Streptavidin (Invitrogen) for 15 min, followed by RT-qPCR analysis.
Biotinylated RNA was obtained from Sangon Biotech.
Flow cytometry (FCM) analysis of
apoptosis
Apoptosis was detected using the Annexin
V-Fluorescein Isothiocyanate (FITC) Apoptosis Detection Kit (BD
Biosciences, New Jersey, USA) according to the manufacturer's
instructions. After incubation with annexin V-FITC (5 µl) and
propidium iodide (PI; 10 µl) in the dark (15 min, 25˚C), cell
apoptosis was assessed by FCM (FACSCanto II; BD FACSChorus
software, version: 1.0; BD Biosciences).
Transwell migration and invasion
assays
For the Transwell migration assay, BC cells were
transferred to the upper chamber of a Transwell plate (Corning, New
York, USA) containing 100 µl of culture medium without FBS. The
lower chamber contained 600 µl of culture medium containing FBS.
After 48 h of incubation, BC cells were immobilized with 4%
paraformaldehyde for 15 min, washed with PBS, and stained with 0.1%
crystal violet for 10 min. The migrated cells were counted in five
randomly selected visual fields. For the Transwell invasion assay,
the experimental procedure was similar to that of the Transwell
migration assay, except for the replacement of the Transwell
migration plate with the Transwell invasion plate (Corning).
Western blot analysis
A Beyotime Biotechnology kit (Shanghai, China) was
used to detect the protein concentration in each sample. We mixed
40 µg protein with SDS buffer (5X), boiled the sample for 10 min,
and added these samples to 10% SDS-PAGE gels for electrophoresis.
Proteins were then transferred from the SDS-PAGE gel to a PVDF
membrane (Millipore, Billerica, Massachusetts, USA). The membrane
was blocked with 5% skim milk powder in Tris-buffered saline with
Tween 20 (0.1%, v/v; Solarbio) for 1 h at 25˚C. The membrane was
then incubated overnight at 4˚C, according to the optimum
conditions of the primary antibodies. After washing thrice, the
membrane was incubated with the appropriate horseradish peroxidase
secondary antibody for 1 h. Finally, enhanced chemiluminescence
(Millipore) was used to detect the blots. Antibodies against TRAF2
(1:1,000, ab126758), NF-κB (1:100, ab16502), and p-NF-κB (1:1,000,
ab76302) were obtained from Abcam (Cambridge, UK).
Dual luciferase assays
The SNHG12 or TRAF2 3'-UTR sequences
were inserted into the psi-CHECK2 construct and co-transfected into
HEK293 cells with miR-140-3p mimic/inhibitor using Lipofectamine
3000 at 37˚C for 4 h. After 48 h of culture, the Dual-Luciferase
Reporter assay system (Promega Corp., Wisconsin, USA) was used to
measure luciferase activity at 490 nm. The ratio of firefly to
Renilla luciferase activity was used to normalize the
firefly luciferase values.
Statistical analysis
Three separate experiments were conducted for each
measurement. The corresponding results are expressed as mean ±
standard deviation. Statistical analysis was performed using an
unpaired Student's t-test or one-way analysis of variance followed
by Tukey's post hoc test using SPSS (Chicago, USA). Statistical
significance was set at P<0.05.
Results
SCO inhibits BC cell growth and SNHG12
expression
Overall, SCO exhibited time- and dose-dependent
inhibitory effects on the viability of the four BC cell types to
varying degrees (Fig. 1A-D).
However, when the SCO dose reached 1000 µM, the viability of
MCF-10A cells significantly decreased (Fig. 1E). Therefore, we used a relatively
safe dose of 500 µM in subsequent experiments to explore the
molecular mechanism by which SCO inhibits BC cell viability.
Compared with the normal mammary epithelial cell line MCF-10A,
lncRNA SNHG12 expression in BC cells was upregulated to
varying degrees (Fig. 1F). Among
these cell lines, the highest expression of SNHG12 was
observed in MCF-7 and MDA-MB-231 cells. Therefore, these two cell
lines were used in subsequent experiments to study cellular
function. In addition, the expression of SNHG12 gradually
decreased with increasing SCO concentrations (Fig. 1G and H). These results suggested that the
inhibition of BC cell viability by SCO may be involved in
corresponding inhibition of SNHG12 expression.
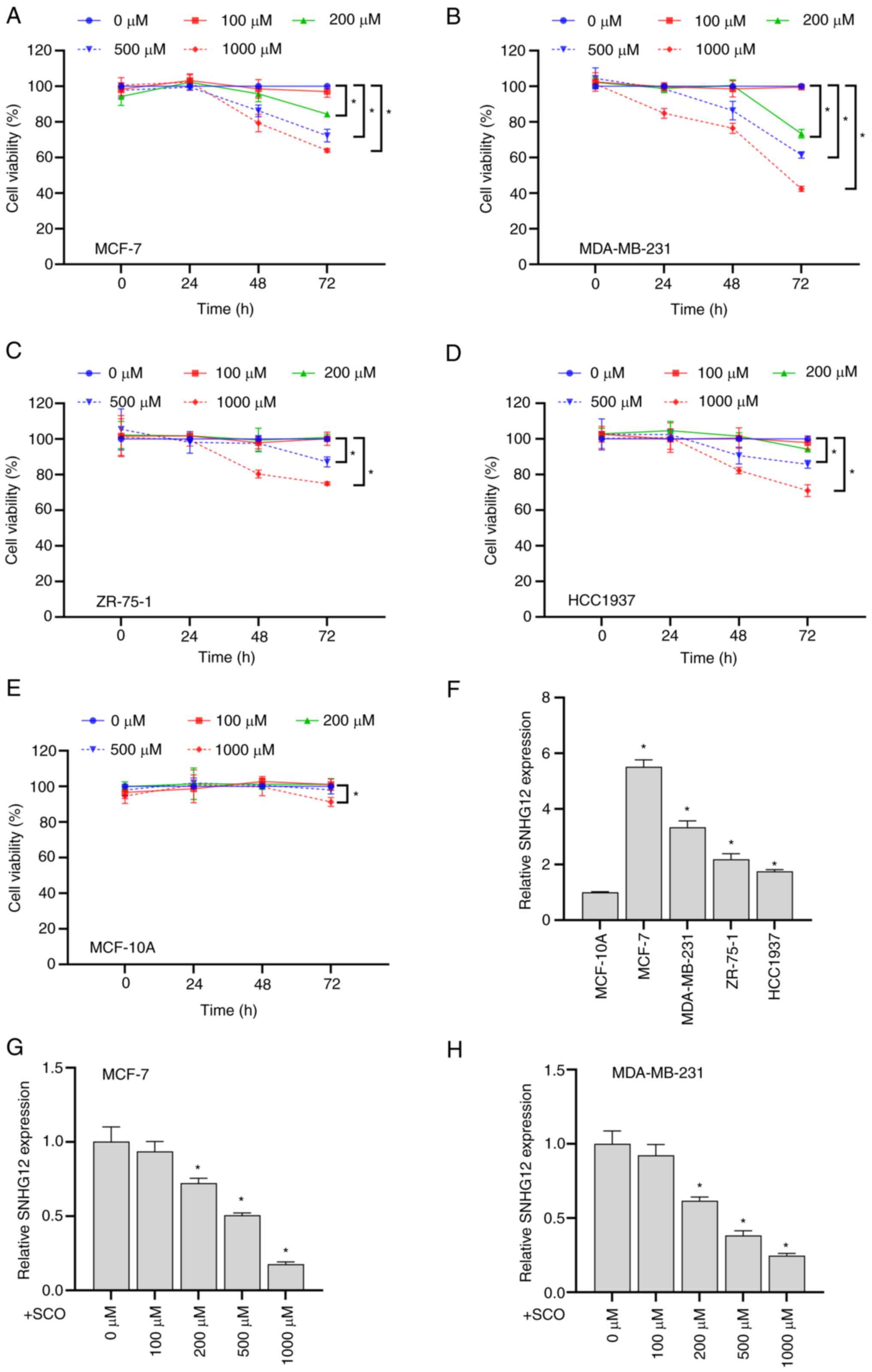 | Figure 1SCO inhibits BC cell growth and
SNHG12 expression. CCK8 assay to analyze the effects of
different doses of SCO on the cell viability of (A) MCF-7, (B)
MDA-MB-231, (C) ZR-75-1, (D) HCC1937 and (E) MCF-10A cells.
*P<0.05. (F) RT-qPCR analysis of the differential
expression of lncRNA SNHG12 in MCF-7, MDA-MB-231, ZR-75-1
and HCC1937 cells compared to MCF-10A. *P<0.05 vs.
MCF-10A. RT-qPCR analysis of the differing expression levels of
lncRNA SNHG12 in (G) MCF-7 and (H) MDA-MB-231 cells under
different SCO doses. *P<0.05 vs. 0 µM. CCK8, Cell
Counting Kit-8; SCO, scoparone; BC, breast cancer; lncRNA, long
non-coding RNA; SNHG12, small nucleolar RNA host gene 12;
RT-qPCR, reverse transcription-quantitative PCR. |
Overexpression of SNHG12 alleviated
the SCO-mediated growth inhibition of BC cells
We constructed a SNHG12 overexpression
plasmid and transfected it into BC cells. The corresponding results
demonstrated that the expression of SNHG12 was significantly
upregulated in ov-SNHG12 compared to that in ov-NC (Fig. 2A). Additionally, SCO inhibited
SNHG12 expression, cell viability, and migration/invasion
ability, and increased apoptosis, all of which were reversed by
overexpression of SNHG12 (Fig.
2B-E). To further understand the functional principle of
SNHG12, we conducted a nuclear plasma separation experiment.
Similar to the positive control GAPDH, SNHG12 was highly
expressed in the cytoplasm, whereas the positive control U6 was
highly expressed in the nucleus (Fig.
2F). This finding suggests that SNHG12 exerts its
downstream effects through post-transcriptional epigenetic
regulation.
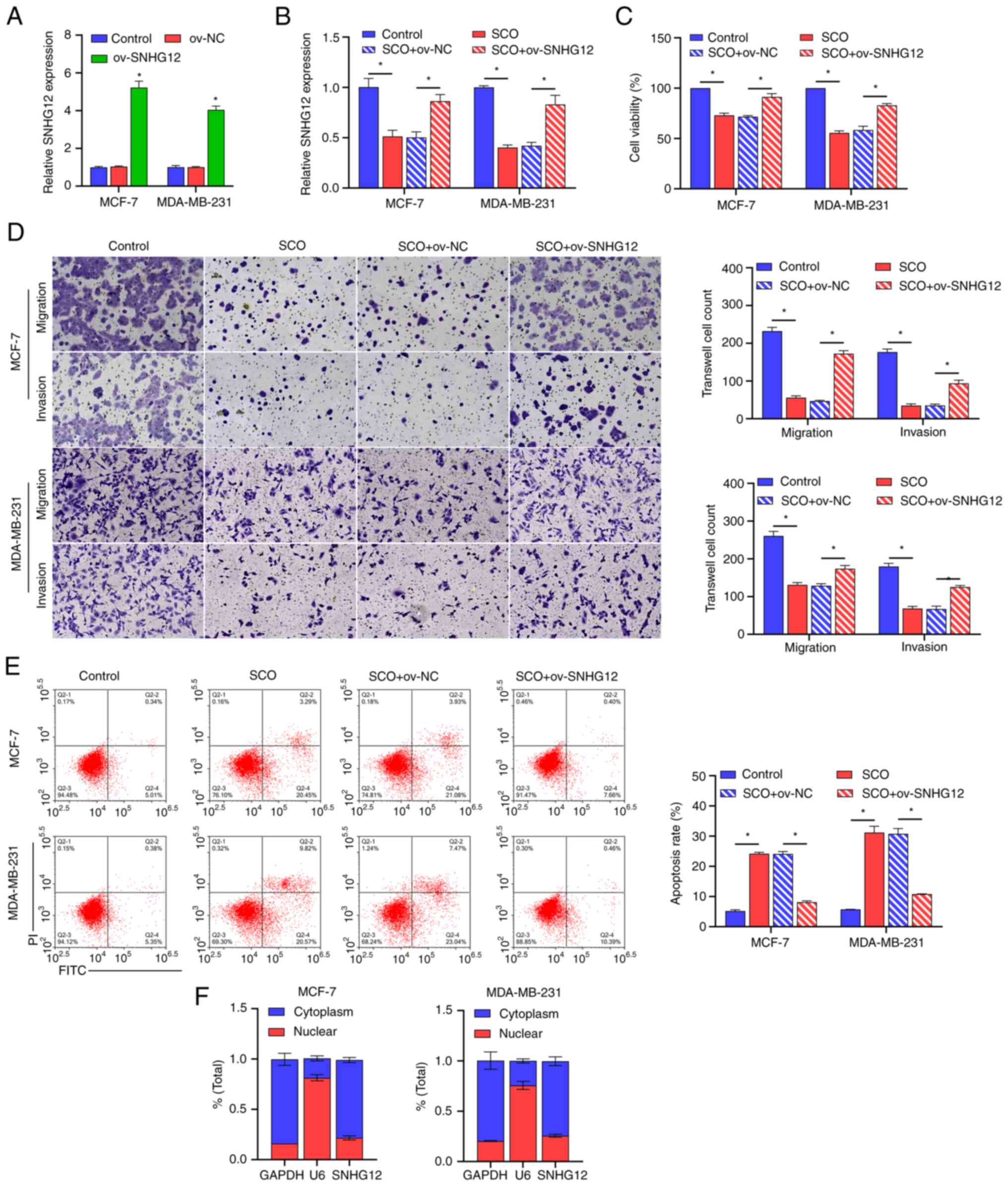 | Figure 2Overexpression of SNHG12
alleviates the SCO-mediated growth inhibition of BC cells. (A)
RT-qPCR analysis verifying the effectiveness of SNHG12
overexpression plasmids. *P<0.05 vs. ov-NC. (B)
RT-qPCR analysis of the effect of SNHG12 overexpression on
SNHG12 expression in SCO-treated cells. (C) CCK8 analysis of
the effect of SNHG12 overexpression on BC cell viability in
SCO-treated cells. (D) Transwell analysis of the effect of
SNHG12 overexpression on BC cell migration/invasion in
SCO-treated cells. Magnification, x200. (E) FCM analysis of the
effect of SNHG12 overexpression on apoptosis in SCO-treated
cells. (F) Nucleocytoplasmic separation analysis of the
localization of SNHG12 in MCF-7 and MDA-MB-231 cells.
*P<0.05. CCK8, Cell Counting Kit-8; SCO, scoparone;
BC, breast cancer; lncRNA, long non-coding RNA; SNHG12,
small nucleolar RNA host gene 12; ov, overexpression; RT-qPCR,
reverse transcription-quantitative PCR; NC, negative control; FCM,
flow cytometry; FITC, fluorescein isothiocyanate; PI, propidium
iodide. |
SNHG12 acts as a ceRNA to adsorb
miR-140-3p
Through joint analysis of the LncBase and Starbase
databases, we determined that miR-361-3p and miR-140-3p may be
potential targets of SNHG12 (Fig. 3A). Studies have shown that
miR-140-3p acts as a tumor suppressor gene in BC and when this gene
is suppressed, BC progression is promoted (20,21);
however, it has been established that miR-361-3p may be unfavorable
for the treatment and prognosis of BC (22,23).
Further, miR-140-3p expression was observed to be downregulated in
both MCF-7 and MDA-MB-231 cells compared with that in the normal
mammary epithelial cell line MCF-10A (Fig. 3B). However, with an increasing
concentration of SCO, the miR-140-3p expression gradually increased
(Fig. 3C and D). In this study, the miR-140-3p
mimic/inhibitor was synthesized and transfected into MCF-7 and
MDA-MB-231 cells. The corresponding expression of miR-140-3p was
up-regulated in the mimic-transfected group and downregulated in
the inhibitor-transfected group, suggesting that the synthesis of
the miR-140-3p mimic/inhibitor was effective (Fig. 3E). Bioinformatic analysis
demonstrated that there were potential binding sites between
SNHG12 and miR-140-3p (Fig.
3F). Subsequent dual luciferase results showed that the
fluorescence activity of the WT SNHG12 + miR-140-3p mimic
co-transfected group was significantly decreased compared with that
of the WT SNHG12 + miR-140-3p mimic NC co-transfected group.
Nonetheless, there was no significant difference observed between
the fluorescence activity of the mutant SNHG12 + miR-140-3p
inhibitor NC co-transfection group and the mutant SNHG12 +
miR-140-3p inhibitor co-transfection group (Fig. 3G). Subsequent RNA pull-down assays
showed that miR-140-3p and SNHG12 were enriched in the miR-140-3p
and SNHG12 groups, further establishing the binding efficiency
between SNHG12 and miR-140-3p (Fig.
3H and I). In addition,
ov-SNHG12 inhibited the expression of miR-140-3p compared
with that of the o-NC group (Fig.
3J). However, changes in miR-140-3p expression had no
significant effect on the expression of SNHG12 (Fig. 3K).
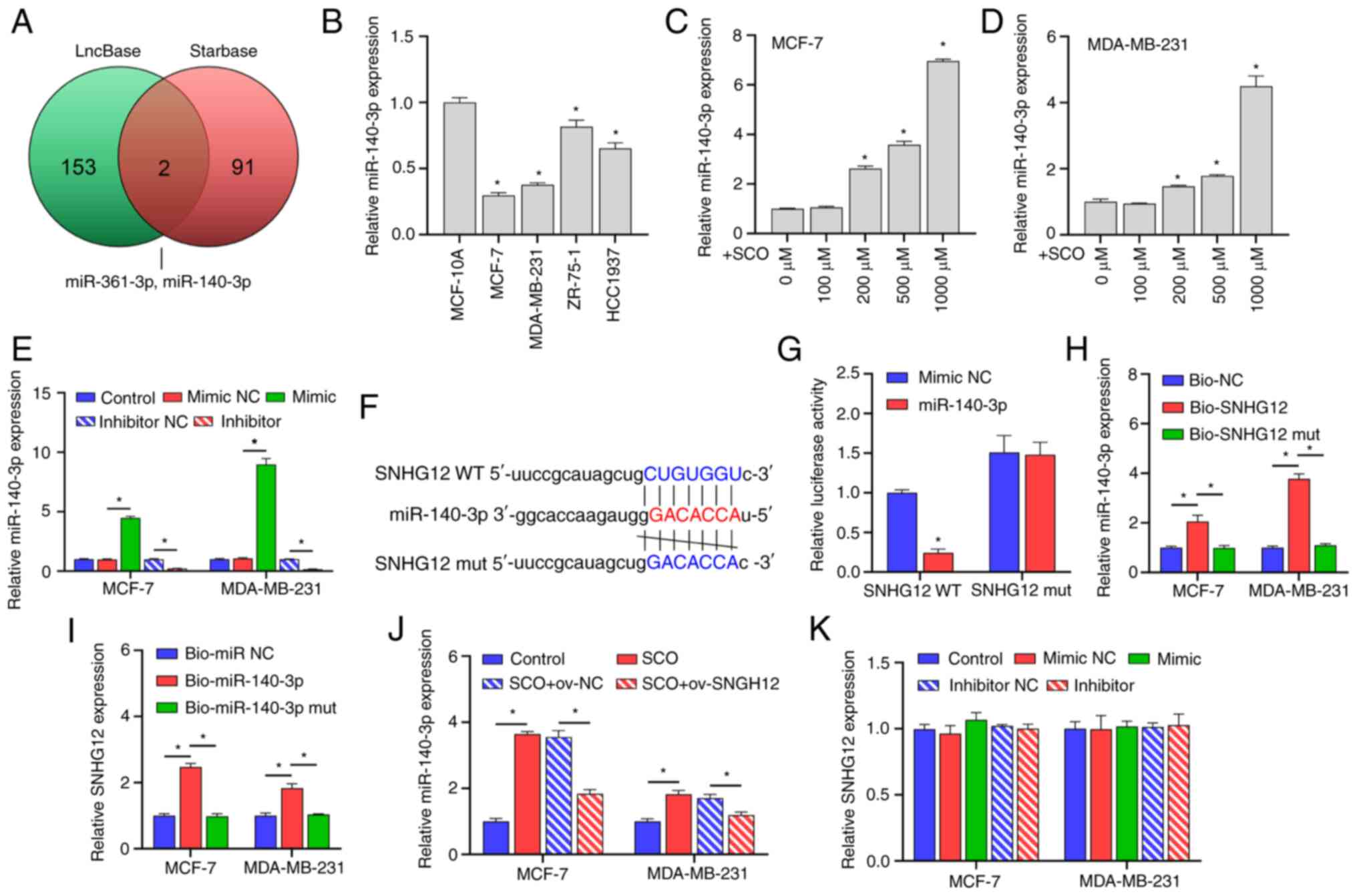 | Figure 3SNHG12 acts as a ceRNA to
adsorb miR-140-3p. (A) LncBase and Starbase joint analysis of
potential targets for SNHG12. (B) RT-qPCR analysis of the
differential expression of miR-140-3p in MCF-7, MDA-MB-231, ZR-75-1
and HCC1937 cells compared to MCF-10A. *P<0.05 vs.
MCF-10A. RT-qPCR analysis of the difference in expression of
miR-140-3p in (C) MCF-7 and (D) MDA-MB-231 cells under different
SCO doses. *P<0.05 vs. 0 µM. (E) RT-qPCR analysis
verifying the effectiveness of the miR-140-3p mimic/inhibitor.
*P<0.05. (F) Bioinformatic analysis of the potential
binding sites between SNHG12 and miR-140-3p. (G) Dual
luciferase analysis of the binding between SNHG12 and
miR-140-3p. *P<0.05 vs. mimic NC. RNA pull-down
analysis was used to examine the binding between (H) bio-SNHG12 and
miR-140-3p, as well as (I) bio-miR-140-3p and SNHG12,
respectively, in MCF-7 and MDA-MB-231 cells. (J) RT-qPCR analysis
of miR-140-3p expression under the combined action of SCO and
ov-SNHG12. (K) RT-qPCR analysis of the effect of miR-140-3p
mimic/inhibitor on SNHG12 expression. *P<0.05.
SCO, scoparone; lncRNA, long non-coding RNA; SNHG12, small
nucleolar RNA host gene 12; RT-qPCR, reverse
transcription-quantitative PCR; NC, negative control; miR,
microRNA; Bio, biotinylated; ceRNA, competing endogenous RNA. |
miR-140-3p directly targets TRAF2 to
regulate the NF-κB signaling pathway
miRNAs are typically used to bind to the 3'-UTR of
the downstream target to inhibit transcription and translation,
thereby affecting signal transduction between BC cells (24,25).
Using the miRDB and Starbase databases, we found that many putative
target genes of miR-140-3p, such as TRAF2, are upstream
regulators of NF-κB signaling (Fig.
4A). Prior studies have shown that TRAF2 is involved in
the activation of NF-κB (26) and
induces the growth and distal migration of breast cancer cells
(27). In the current study,
TRAF2 expression was upregulated in both MCF-7 and
MDA-MB-231 cells compared to that in the normal mammary epithelial
cell line MCF-10A (Fig. 4B).
However, as the concentration of SCO increased, TRAF2
expression gradually decreased (Fig.
4C and D). Bioinformatic
analysis indicated that there were potential binding sites between
miR-140-3p and TRAF2 (Fig.
4E). The dual luciferase assay results showed that the
fluorescence activity of the WT TRAF2 + miR-140-3p mimic
co-transfected group was significantly decreased compared with that
of the WT TRAF2 + miR-140-3p mimic NC co-transfected group.
Nonetheless, there was no significant difference observed between
the fluorescence activity of the mutant TRAF2 + miR-140-3p
inhibitor NC co-transfection and mutant TRAF2 + miR-140-3p
inhibitor co-transfection groups (Fig.
4F). In addition, SCO inhibited TRAF2 protein levels, and NF-κB
activation was partially reversed by the miR-140-3p inhibitor.
Overall, miR-140-3p negatively regulated TRAF2 expression (Fig. 4G).
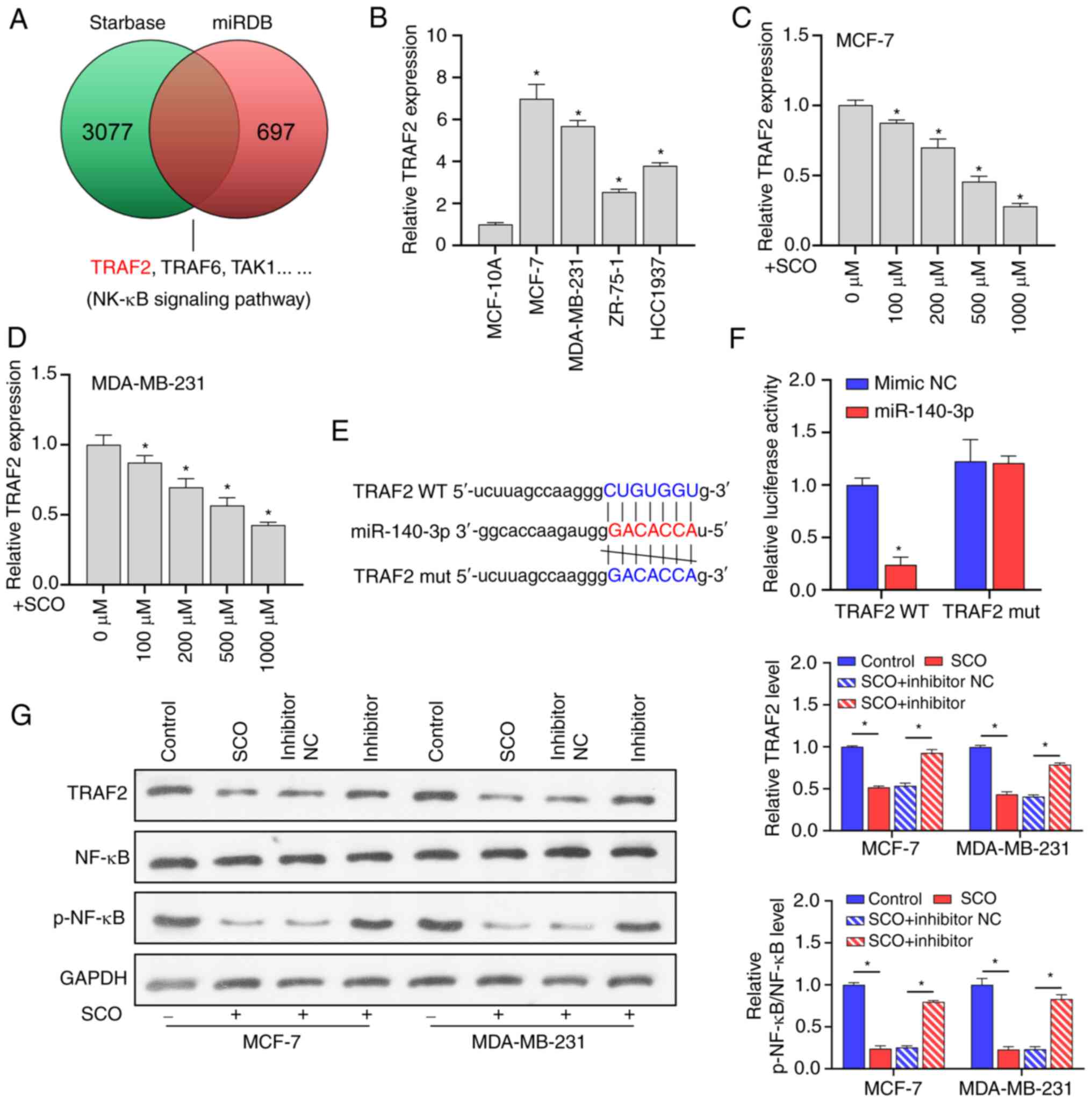 | Figure 4miR-140-3p directly targets TRAF2 to
regulate the NF-κB signaling pathway. (A) miRDB and Starbase
database joint analysis of potential targets for miR-140-3p. (B)
RT-qPCR analysis of the differential expression of TRAF2 in
MCF-7, MDA-MB-231, ZR-75-1 and HCC1937 cells compared to MCF-10A.
*P<0.05 vs. MCF-10A. RT-qPCR analysis of TRAF2
expression in (C) MCF-7 and (D) MDA-MB-231 cells under different
SCO doses. *P<0.05 vs. 0 µM. (E) Bioinformatic
analysis of the potential binding sites between TRAF2 and
miR-140-3p. (F) Dual luciferase analysis of the binding between
TRAF2 and miR-140-3p. *P<0.05 vs. mimic NC.
(G) Western blot analysis of the effect of SCO combined with
miR-140-3p inhibitor on protein levels of TRAF2, NF-κB, and
p-NF-κB. *P<0.05. SCO, scoparone; TRAF2,
receptor-associated factor 2; RT-qPCR, reverse
transcription-quantitative PCR; NC, negative control; miR,
microRNA; Bio, biotinylated; ceRNA, competing endogenous RNA;
NF-κB, nuclear factor κB. |
miR-140-3p is negatively correlated
with TRAF2 expression
To confirm the negative correlation between
miR-140-3p and TRAF2, we constructed three siRNAs that
targeted TRAF2 and transfected MCF-7 and MDA-MB-231 cells
with these siRNAs. Corresponding TRAF2 gene expression and protein
levels were downregulated in BC cells after siRNA transfection
(Fig. 5A and B). Among these siRNAs, si-TRAF2-3
possessed the best inhibition efficiency; therefore,
si-TRAF2-3 was used in subsequent experiments. Subsequent
RT-qPCR results demonstrated that, compared with the control group,
the miR-140-3p inhibitor upregulated the expression of TRAF2
(Fig. 5C). In the presence of SCO,
inhibition of TRAF2 expression had no significant effect on
the expression of miR-140-3p (Fig.
5D). Nonetheless, in the presence of SCO and miR-140-3p
inhibitor, the inhibition of TRAF2 gene expression reduced
corresponding TRAF2 protein levels (Fig. 5E). These results indicated that
miR-140-3p is negatively correlated with the expression of the
downstream target gene TRAF2.
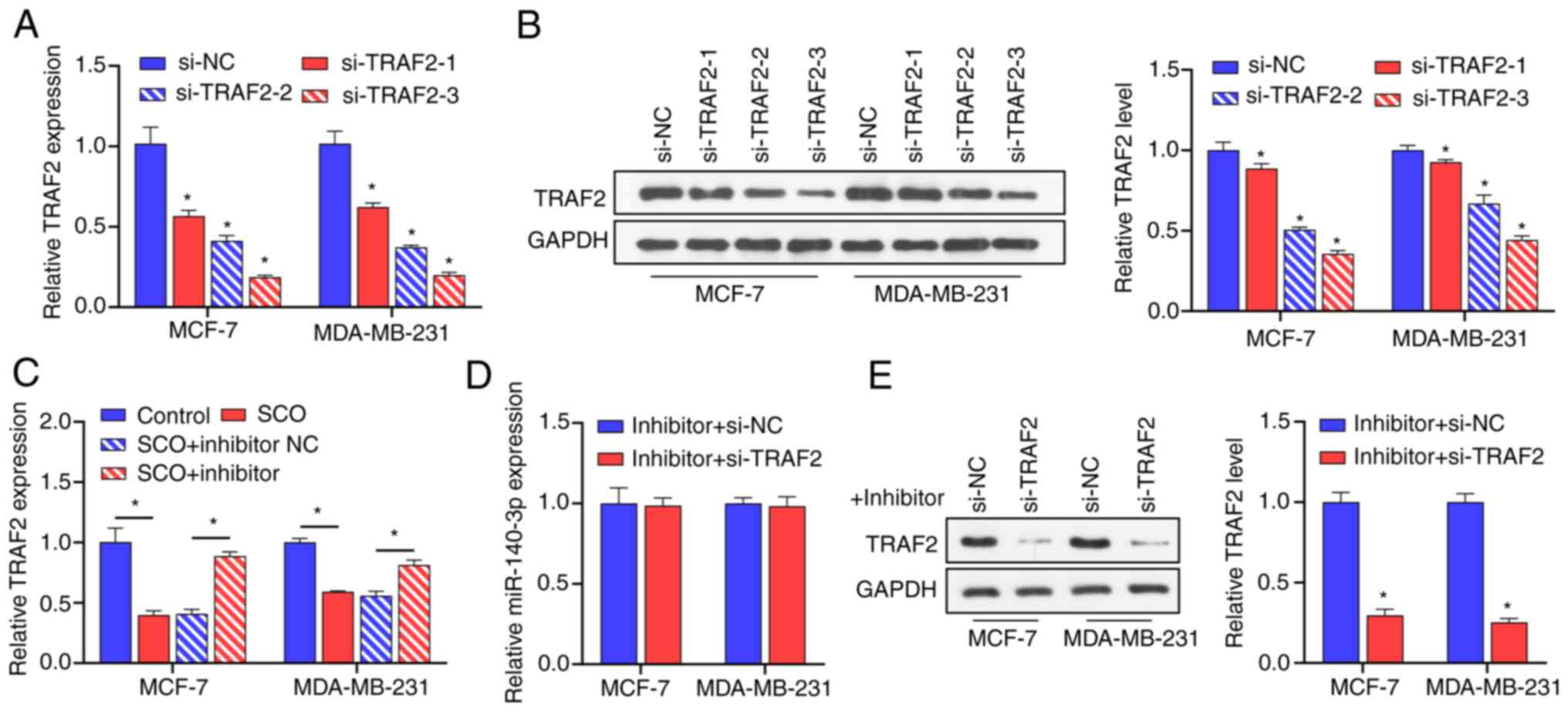 | Figure 5miR-140-3p is negatively correlated
with the expression of TRAF2. (A) RT-qPCR analysis verifying the
effectiveness of 3 siRNAs against TRAF2 in MCF-7 and
MDA-MB-231 cells. *P<0.05 vs. si-NC. (B) Western blot
analysis verifying the effectiveness of 3 siRNAs against
TRAF2 in MCF-7 and MDA-MB-231 cells. *,
si-TRAF2-1, si-TRAF2-2,si-TRAF2-3 groups vs.
si-NC group. (C) RT-qPCR analysis of the effect of SCO combined
with miR-140-3p inhibitor on the expression of TRAF2.
*P<0.05. (D) RT-qPCR analysis of the effect of
si-TRAF2 on miR-140-3p expression. (E) Western blot analysis of the
effect of si-TRAF2 on protein levels of TRAF2 in the presence of
miR-140-3p inhibitor. *P<0.05 vs. inhibitor + si-NC.
SCO, scoparone; TRAF2, receptor-associated factor 2; RT-qPCR,
reverse transcription-quantitative PCR; NC, negative control; miR,
microRNA; Bio, biotinylated; ceRNA, competing endogenous RNA; si,
small interfering. |
SCO regulates the NF-κB signaling
pathway through the SNHG12/miR-140-3p/TRAF2 axis to inhibit BC cell
growth
To understand the mechanism of the
SNHG12/miR-140-3p/TRAF2/NF-κB axis in SCO treatment of BC,
ov-SNHG12 was co-transfected with an miR-140-3p inhibitor
and si-TRAF2 in BC cells in the presence of SCO. Overall, we
found that the promotion of TRAF2 protein levels and NF-κB
activation by the miR-140-3p inhibitor was enhanced by
ov-SNHG12. The combined effect of ov-SNHG12 and the
miR-140-3p inhibitor was partially mitigated in the presence of
si-TRAF2 (Fig. 6A). In
addition, the miR-140-3p inhibitor-mediated promotion of activity,
migration/invasion ability, and apoptosis inhibition in BC cells
was enhanced by overexpression of SNHG12. Further,
inhibition of TRAF2 expression partially prevented the
combined effect of the ov-SNHG12/miR-140-3p inhibitor
(Fig. 6B-D).
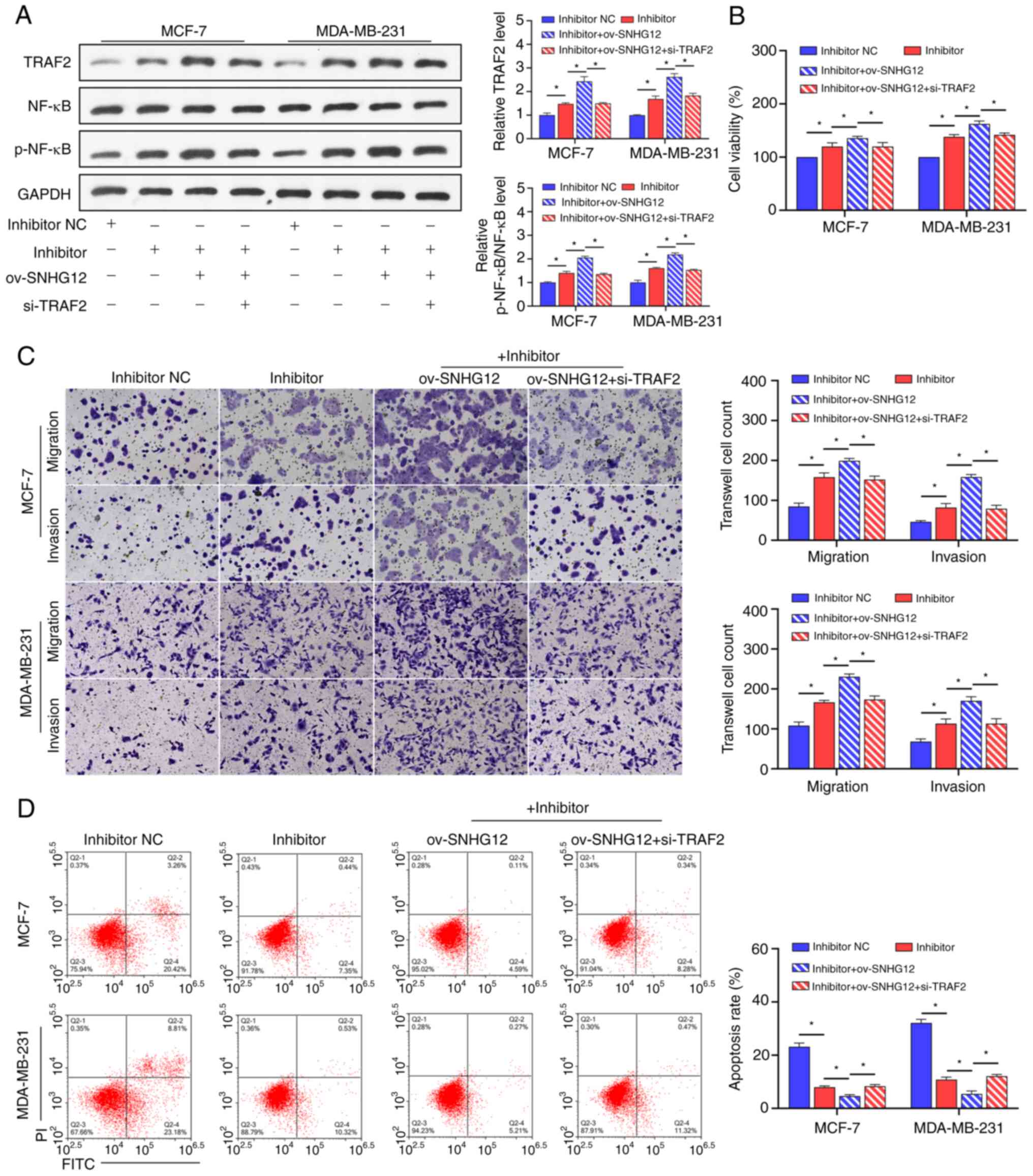 | Figure 6SCO regulates the NF-κB signaling
pathway through the SNHG12/miR-140-3p/TRAF2 axis to
inhibit BC cell growth. (A) Western blot analysis of the effect of
the interaction between ov-SNHG12, miR-140-3p inhibitor, and
si-TRAF2 on protein levels of TRAF2, NF-κB, and p-NF-κB in the
presence of SCO. (B) CCK8 analysis of the effect of the interaction
between ov-SNHG12, miR-140-3p inhibitor, and si-TRAF2 on BC
cell viability in the presence of SCO. (C) Transwell analysis of
the effect of the interaction between ov-SNHG12, miR-140-3p
inhibitor, and si-TRAF2 on BC cell migration/invasion in the
presence of SCO. Magnification, x200. (D) FCM analysis of the
effect of the interaction between ov-SNHG12, miR-140-3p
inhibitor, and si-TRAF2 on BC cell apoptosis in the presence of
SCO. *P<0.05. SCO, scoparone; BC, breast cancer;
lncRNA, long non-coding RNA; SNHG12, small nucleolar RNA
host gene 12; ov, overexpression; TRAF2, receptor-associated factor
2; RT-qPCR, reverse transcription-quantitative PCR; NC, negative
control; miR, microRNA; Bio, biotinylated; ceRNA, competing
endogenous RNA; si, small interfering; NF-κB, nuclear factor κB;
FCM, flow cytometry; FITC, fluorescein isothiocyanate; PI,
propidium iodide. |
Discussion
Prior studies have shown that TCM can effectively
increase treatment efficiency, improve survival rates, and reduce
the side effects of chemotherapy in patients with BC (28,29).
SCO is a major component of the Chinese herbal medicine A.
capillaris (30) and plays an
important role in cancer therapy (7,31).
In the current study, our results confirmed that SCO inhibited BC
cell viability and promoted apoptosis, aligning with those of a
previous study (9). This indicated
that SCO had a significant anti-BC effect. Notably, our data also
indicated that SCO is less toxic to normal mammary epithelial cells
and is suitable for the treatment of BC.
Our finding that lncRNA SNHG12 is upregulated
in BC cells and plays an oncogenic role in BC is consistent with a
previous study (32). Furthermore,
SCO inhibited the expression of SNHG12 in a dose-dependent manner,
whereas the overexpression of SNHG12 reversed the effect of SCO on
BC cells. Therefore, we determined that SCO inhibits the growth of
BC cells through SNHG12. Cytoplasmic lncRNA acts as a ceRNA;
therefore, we first conducted a nucleocytoplasmic separation
experiment and confirmed that SNHG12 is predominantly located in
the cytoplasm of BC cells, which is consistent with a previous
study (33). We then demonstrated
for the first time that SNHG12 adsorbs miR-140-3p, a tumor
suppressor gene in BC (20),
thereby downregulating the expression of miR-140-3p. Therefore,
SCO-mediated inhibition of SNHG12 expression promotes the
expression of miR-140-3p, leading to a reduction in BC cell
growth.
It is well understood that TRAF2 acts as an
oncogene in various cancers, such as gastric cancer (34) and prostate cancer (35), and plays a key role in the
promotion of cell viability. Additionally, high TRAF2
expression promotes the growth and invasion of BC cells (27,36).
The current study confirmed, for the first time, the direct binding
relationship between miR-140-3p and TRAF2. TRAF2 is highly
expressed in BC, and the growth and migration of BC cells promoted
by the miR-140-3p inhibitor were reversed by si-TRAF2. Prior
studies have demonstrated that abnormal NF-κB regulation
contributes to autoimmune diseases, chronic inflammation, and many
cancers (37,38). In BC progression, NF-κB activation
is an important contributor to tumor development (39). As an upstream effector of NF-κB,
the main pathway through which TRAF2 mediates BC progression
is via NF-κB activation (40). In
the present study, we demonstrated that SCO suppresses NF-κB
signaling, but this signaling was partially restored by the
miR-140-3p inhibitor. The promoting effect of the miR-140-3p
inhibitor on NF-κB signaling was enhanced by overexpression of
SNHG12 and decreased by treatment with si-TRAF2. Thus far,
we have confirmed that SCO inhibits the NF-κB signaling pathway
through the SNHG12/miR-140-3p/TRAF2 axis, inhibits BC cell
viability, and promotes apoptosis.
Nonetheless, this study had some limitations. First,
we did not analyze the therapeutic effect of SCO in BC patients or
the correlation of clinical data. Second, the development of BC
involves many RNA and protein interactions and signaling pathway
alterations, which have not been assessed here. Third, SCO directly
or indirectly affects the stability or processing of SNHG12,
potentially through post-transcriptional modification or through
interactions with RNA-binding proteins or other regulatory factors;
however, these mechanisms were not explored within this study.
Finally, it is unknown whether SCO can increase the susceptibility
of BC cells to drug resistance. Overall, these research areas
remain unclear; therefore, we aim to focus on these factors in
future research of SCO.
In conclusion, our study demonstrated that SCO
inhibits BC cell survival by inhibiting intercellular NF-κB
signaling, which is related to the regulation of the
SNHG12/miR-140-3p/TRAF2 axis. These results indicated
that SCO may be a promising anti-BC therapeutic drug, which
provides a strong theoretical basis for the treatment of BC.
Supplementary Material
Sequence of miR-140-3p
mimic/inhibitor, si-TRAF2 and their NCs.
Sequences of miR-140-3p, TRAF2,
β-actin and U6 primers for RT-qPCR.
Acknowledgements
Not applicable.
Funding
Funding: No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
CE designed experiments. XW and XL performed
experiments and drafted the manuscript. JL, XZ and YC collected and
analyzed the data. CE and JL confirm the authenticity of all the
raw data. All authors read and approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Azamjah N, Soltan-Zadeh Y and Zayeri F:
Global trend of breast cancer mortality rate: A 25-year study.
Asian Pac J Cancer Prev. 20:2015–2020. 2019.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Harbeck N and Gnant M: Breast cancer.
Lancet. 389:1134–1150. 2017.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Libson S and Lippman M: A review of
clinical aspects of breast cancer. Int Rev Psychiatry. 26:4–15.
2014.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Tang JL, Liu BY and Ma KW: Traditional
Chinese medicine. Lancet. 372:1938–1940. 2008.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Pei S, Yang X, Wang H, Zhang H, Zhou B,
Zhang D and Lin D: Plantamajoside, a potential anti-tumor herbal
medicine inhibits breast cancer growth and pulmonary metastasis by
decreasing the activity of matrix metalloproteinase-9 and -2. BMC
Cancer. 15(965)2015.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Liew AC, Peh KK, Tan BS, Zhao W and
Tangiisuran B: Evaluation of chemotherapy-induced toxicity and
health-related quality of life amongst early-stage breast cancer
patients receiving Chinese herbal medicine in Malaysia. Support
Care Cancer. 27:4515–4524. 2019.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Jung SH, Lee GB, Ryu Y, Cui L, Lee HM, Kim
J, Kim B and Won KJ: Inhibitory effects of scoparone from chestnut
inner shell on platelet-derived growth factor-BB-induced vascular
smooth muscle cell migration and vascular neointima hyperplasia. J
Sci Food Agric. 99:4397–4406. 2019.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Wang Y, Wang M, Chen B and Shi J:
Scoparone attenuates high glucose-induced extracellular matrix
accumulation in rat mesangial cells. Eur J Pharmacol. 815:376–380.
2017.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Kim JK, Kim JY, Kim HJ, Park KG, Harris
RA, Cho WJ, Lee JT and Lee IK: Scoparone exerts anti-tumor activity
against DU145 prostate cancer cells via inhibition of STAT3
activity. PLoS One. 8(e80391)2013.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Kielbus M, Skalicka-Wozniak K, Grabarska
A, Jeleniewicz W, Dmoszynska-Graniczka M, Marston A, Polberg K,
Gawda P, Klatka J and Stepulak A: 7-substituted coumarins inhibit
proliferation and migration of laryngeal cancer cells in vitro.
Anticancer Res. 33:4347–4356. 2013.PubMed/NCBI
|
|
11
|
Zhao Z, Guo Y, Liu Y, Sun L, Chen B, Wang
C, Chen T, Wang Y, Li Y, Dong Q, et al: Individualized lncRNA
differential expression profile reveals heterogeneity of breast
cancer. Oncogene. 40:4604–4614. 2021.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Venkatesh J, Wasson MD, Brown JM, Fernando
W and Marcato P: LncRNA-miRNA axes in breast cancer: Novel points
of interaction for strategic attack. Cancer Lett. 509:81–88.
2021.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Qiao K, Ning S, Wan L, Wu H, Wang Q, Zhang
X, Xu S and Pang D: LINC00673 is activated by YY1 and promotes the
proliferation of breast cancer cells via the miR-515-5p/MARK4/Hippo
signaling pathway. J Exp Clin Cancer Res. 38(418)2019.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Yuan JH, Li WX, Hu C and Zhang B:
Upregulation of SNHG12 accelerates cell proliferation, migration,
invasion and restrain cell apoptosis in breast cancer by enhancing
regulating SALL4 expression via sponging miR-15a-5p. Neoplasma.
67:861–870. 2020.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Zhou Y, Wang B, Wang Y, Chen G, Lian Q and
Wang H: miR-140-3p inhibits breast cancer proliferation and
migration by directly regulating the expression of tripartite motif
28. Oncol Lett. 17:3835–3841. 2019.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Sun LL, Wang J, Zhao ZJ, Liu N, Wang AL,
Ren HY, Yang F, Diao KX, Fu WN, Wan EH and Mi XY: Suppressive role
of miR-502-5p in breast cancer via downregulation of TRAF2. Oncol
Rep. 31:2085–2092. 2014.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Taminiau A, Draime A, Tys J, Lambert B,
Vandeputte J, Nguyen N, Renard P, Geerts D and Rezsohazy R: HOXA1
binds RBCK1/HOIL-1 and TRAF2 and modulates the TNF/NF-κB pathway in
a transcription-independent manner. Nucleic Acids Res.
44:7331–7349. 2016.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Jang KW, Lee KH, Kim SH, Jin T, Choi EY,
Jeon HJ, Kim E, Han YS and Chung JH: Ubiquitin ligase CHIP induces
TRAF2 proteasomal degradation and NF-κB inactivation to regulate
breast cancer cell invasion. J Cell Biochem. 112:3612–3620.
2011.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408.
2001.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Onyeisi JOS, Greve B, Espinoza-Sanchez NA,
Kiesel L, Lopes CC and Gotte M: microRNA-140-3p modulates
invasiveness, motility, and extracellular matrix adhesion of breast
cancer cells by targeting syndecan-4. J Cell Biochem.
122:1491–1505. 2021.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Dou D, Ren X, Han M, Xu X, Ge X, Gu Y,
Wang X and Zhao S: Circ_0008039 supports breast cancer cell
proliferation, migration, invasion, and glycolysis by regulating
the miR-140-3p/SKA2 axis. Mol Oncol. 15:697–709. 2021.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Zamarbide Losada JN, Sulpice E, Combe S,
Almeida GS, Leach DA, Choo J, Protopapa L, Hamilton MP, McGuire S,
Gidrol X, et al: Apoptosis-modulatory miR-361-3p as a novel
treatment target in endocrine-responsive and endocrine-resistant
breast cancer. J Endocrinol. 256(e220229)2023.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Hua B, Li Y, Yang X, Niu X, Zhao Y and Zhu
X: MicroRNA-361-3p promotes human breast cancer cell viability by
inhibiting the E2F1/P73 signalling pathway. Biomed Pharmacother.
125(109994)2020.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Jin T, Zhang Y and Zhang T: MiR-524-5p
Suppresses migration, invasion, and EMT progression in breast
cancer cells through targeting FSTL1. Cancer Biother Radiopharm.
35:789–801. 2020.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Cheng S, Huang Y, Lou C, He Y, Zhang Y and
Zhang Q: FSTL1 enhances chemoresistance and maintains stemness in
breast cancer cells via integrin β3/Wnt signaling under miR-137
regulation. Cancer Biol Ther. 20:328–337. 2019.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Gu M, Zhou W, Chen J, Zhao Y, Xie C and
Zhou Z: TRAF2 gene silencing induces proliferation and represses
apoptosis of nucleus pulposus cells in rats with intervertebral
disc degeneration. Life Sci. 279(119670)2021.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Yao Y, Zhao K, Yu Z, Ren H, Zhao L, Li Z,
Guo Q and Lu N: Wogonoside inhibits invasion and migration through
suppressing TRAF2/4 expression in breast cancer. J Exp Clin Cancer
Res. 36(103)2017.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Li QW, Zhang GL, Hao CX, Ma YF, Sun X,
Zhang Y, Cao KX, Li BX, Yang GW and Wang XM: SANT, a novel Chinese
herbal monomer combination, decreasing tumor growth and
angiogenesis via modulating autophagy in heparanase overexpressed
triple-negative breast cancer. J Ethnopharmacol.
266(113430)2021.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Qi F, Zhao L, Zhou A, Zhang B, Li A, Wang
Z and Han J: The advantages of using traditional Chinese medicine
as an adjunctive therapy in the whole course of cancer treatment
instead of only terminal stage of cancer. Biosci Trends. 9:16–34.
2015.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Liu B, Deng X, Jiang Q, Li G, Zhang J,
Zhang N, Xin S and Xu K: Scoparone improves hepatic inflammation
and autophagy in mice with nonalcoholic steatohepatitis by
regulating the ROS/P38/Nrf2 axis and PI3K/AKT/mTOR pathway in
macrophages. Biomed Pharmacother. 125(109895)2020.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Li N, Yang F, Liu DY, Guo JT, Ge N and Sun
SY: Scoparone inhibits pancreatic cancer through PI3K/Akt signaling
pathway. World J Gastrointest Oncol. 13:1164–1183. 2021.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Liu Y, Cheng G, Huang Z, Bao L, Liu J,
Wang C, Xiong Z, Zhou L, Xu T, Liu D, et al: Long noncoding RNA
SNHG12 promotes tumour progression and sunitinib resistance by
upregulating CDCA3 in renal cell carcinoma. Cell Death Dis.
11(515)2020.PubMed/NCBI View Article : Google Scholar
|
|
33
|
Zhou S, Yu L, Xiong M and Dai G: LncRNA
SNHG12 promotes tumorigenesis and metastasis in osteosarcoma by
upregulating Notch2 by sponging miR-195-5p. Biochem Biophys Res
Commun. 495:1822–1832. 2018.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Ye Y, Ye F, Li X, Yang Q, Zhou J, Xu W,
Aschner M, Lu R and Miao S: 3,3'-diindolylmethane exerts
antiproliferation and apoptosis induction by TRAF2-p38 axis in
gastric cancer. Anticancer Drugs. 32:189–202. 2021.PubMed/NCBI View Article : Google Scholar
|
|
35
|
Wei B, Liang J, Hu J, Mi Y, Ruan J, Zhang
J, Wang Z, Hu Q, Jiang H and Ding Q: TRAF2 is a valuable prognostic
biomarker in patients with prostate cancer. Med Sci Monit.
23:4192–4204. 2017.PubMed/NCBI View Article : Google Scholar
|
|
36
|
Jiao C, Chen W, Tan X, Liang H, Li J, Yun
H, He C, Chen J, Ma X, Xie Y and Yang BB: Ganoderma lucidum spore
oil induces apoptosis of breast cancer cells in vitro and in vivo
by activating caspase-3 and caspase-9. J Ethnopharmacol.
247(112256)2020.PubMed/NCBI View Article : Google Scholar
|
|
37
|
Poma P: NF-κB and disease. Int J Mol Sci.
21(9181)2020.PubMed/NCBI View Article : Google Scholar
|
|
38
|
Yu H, Lin L, Zhang Z, Zhang H and Hu H:
Targeting NF-kappaB pathway for the therapy of diseases: Mechanism
and clinical study. Signal Transduct Target Ther.
5(209)2020.PubMed/NCBI View Article : Google Scholar
|
|
39
|
Tan Y, Sun R, Liu L, Yang D, Xiang Q, Li
L, Tang J, Qiu Z, Peng W, Wang Y, et al: Tumor suppressor DRD2
facilitates M1 macrophages and restricts NF-κB signaling to trigger
pyroptosis in breast cancer. Theranostics. 11:5214–5231.
2021.PubMed/NCBI View Article : Google Scholar
|
|
40
|
Jiang L, Yu L, Zhang X, Lei F, Wang L, Liu
X, Wu S, Zhu J, Wu G, Cao L, et al: miR-892b silencing activates
NF-κB and promotes aggressiveness in breast cancer. Cancer Res.
76:1101–1111. 2016.PubMed/NCBI View Article : Google Scholar
|














