Obesity is a condition in which adipose tissues
accumulates excessively in the body to an extent that it exerts
detrimental effects on health (16). It is characterized by weight gain,
which is caused by excessive fat accumulation due to excessive
daily food intake and insufficient calorific expenditure (17). Obesity increases the risk of
coronary artery disease (18),
hypertension (19), type 2
diabetes (20), asthma (21), cancer (17), venous thromboembolism (22), periodontal disease (23) and Coronavirus disease 2019
(Covid-19) (24,25).
For the treatment of obesity, reducing daily intake
whilst increasing daily calorific expenditure and increasing the
metabolic rate in the body can confer a significant effects. In a
previous study on the effects of AO on obesity, hepatic lipid
metabolism and insulin sensitivity using high-fat diet-induced
obese mice, AO was found to significantly reduce body weight and
inhibit lipid accumulation in the liver and the white adipose
tissue (WAT) (26). The mechanism
of action was found to be mainly due to AO increasing peroxisome
proliferator-activated receptor (PPAR)-α mRNA expression and
decreasing PPAR-γ mRNA expression in the liver. PPAR-α expression
can inhibit triglyceride synthesis and promote fatty acid oxidation
(27) in another study, while
decreasing PPAR-γ expression can reduce the differentiation of
preadipocytes into adipocytes to decrease fatty acid storage
(28). This suggests to a certain
extent the inhibitory effects AO can exert against obesity.
Diabetes is a condition in which the combination of
genetic and environmental factors contributes to either absolute or
relative insulin deficiency and reduced insulin sensitivity in
target tissue cells (29). This
results in metabolic disorders in the body, which are characterized
by hyperglycemia (29). Diabetic
complications caused by the prolonged exposure to hyperglycemic
conditions are the main causes of organ dysfunction and even
mortality in patients with diabetes (30). Diabetic complications can affect
almost all organs of the body, including the nervous system, heart,
kidney, eyes and blood vessels (29), which can be classified as
macroangiopathy and microangiopathy. Macroangiopathy includes
cardiovascular and heart disease, whereas microangiopathy includes
diabetic nephropathy, cataract and retinopathy (31,32).
Diabetes has also been reported to predispose patients to the more
severe forms of Covid-19, which increases the risk of poorer
prognosis (33). Advanced
glycation end products (AGEs) and aldose reductase (AR) are two
important contributing components to the complications of diabetes
(34). Therefore, AR inhibitors
have been proposed to be a viable option for the treatment of
diabetes mellitus. In the AGE formation and rat lens aldose
reductase (RLAR) inhibition assay (35), AO showed no inhibitory activity on
AGE formation, but showed significant inhibitory activity against
RLAR with an IC50 value of 13.6 µM, suggesting that AO
can exert inhibitory effects against AR. In addition, it was found
in another study that AO can activate the insulin signaling pathway
to increase sensitivity to insulin whilst also improving obesity
(26). Therefore, AO is a
potential candidate for the treatment of diabetic complications and
associated diseases.
NAFLD is defined as a disease caused by excessive
hepatic adipose accumulation associated with insulin resistance
(IR) (36). It is a general term
used for a range of diseases with histological hepatic alterations,
including simple hepatic steatosis, non-alcoholic hepatitis
characterized by hepatocellular damage with inflammation and
varying degrees of fibrosis, cirrhosis and hepatocellular carcinoma
(37,38). IR is therefore a key factor in the
pathogenesis of NAFLD. It can, on the one hand, lead to lipolysis
in adipose tissue, thus providing free fatty acids to the liver,
and on the other hand, it can promote de novo synthesis,
leading to further accumulation of fatty acids in the liver
(39). However, the specific
mechanism driving the pathogenesis of NAFLD remains unclear, where
the main strategy of treatment is to target the IR and intrahepatic
lipid accumulation (40). Under
conditions of high ester and high glucose conditions, AO has been
found to improve IR by downregulating the mRNA expression of genes
associated with lipid metabolism such as PPAR-γ and FAS, whilst
suppressing the mRNA expression of inflammatory cytokines such as
IL-6, IL-1β, MCP-1 and TNF-α in WAT (26). In addition, in a mouse model of
NAFLD induced by high-sugar and high-fat conditions and in oleic
and palmitic acid-treated mouse primary hepatocytes, AO was found
to significantly promote autophagic flow and activate the
transcription factor EB (41).
This inhibited ab initio lipid synthesis and suppressed
lipid accumulation to improve hepatic steatosis (41). Altogether, this provides a
pharmacological treatment avenue for NAFLD and related
complications.
Allergy is an acquisitive hypersensitivity by the
immune system to harmless environment substances (42). This spectrum includes allergic
rhinitis, allergic asthma, food allergies and atopic dermatitis
(also known as eczema) (43,44).
Immunoglobulin E (IgE) is one of the key drivers of allergic
responses (45). Although it is
the least abundant antibody in the human serum, it can induce an
effective inflammatory immune response in various tissues and
organs, whilst also serving as a Th2 biomarker involved
in the regulation of Th2 inflammatory responses
(46,47). Previous studies on the effects of
AO on IgE-mediated allergic responses and lipopolysaccharide
(LPS)-induced RAW264.7 cells have found that AO can inhibit the
expression of TNF-α and IL-4 mRNA whilst also suppressing the
expression of prostaglandin E2 and cyclooxygenase-2 (48,49).
Aryl hydrocarbon receptors (AhRs) is a
ligand-activated transcription factor and is present in important
signaling pathways in the mammalian immune system (50). In addition, they can regulate the
differentiation of monocytes into dendritic cells and that of T
cells into regulatory T cells and Th17 cells (51). AhRs also serves an important role
in anticancer effects, energy metabolism, immunity and drug
metabolism (50,52,53),
such that it has been shown that activation of AhRs decreases the
immune response; AO exhibits significant AhRs activity and it may
be a significant natural AhR agonist (54). It has also been found that the
natural plant extract mixture AF-343, obtained from Cassia tora
L., Ulmus pumila L. and Taraxacum officinale, is
potentially a natural candidate for the prevention and treatment of
mast cell-induced allergic diseases, such as allergic inflammation
(55,56). Since the natural active compounds
of AF-343 also include AO, this suggests the possible benefits of
using AO for the treatment of allergy-related disorders.
In 2019, chronic obstructive pulmonary disease
(COPD) and asthma were respiratory diseases with high morbidity and
mortality rates in China, the United States and other regions
(57,58). Airway smooth muscle contraction is
one of the causes of both of the aforementioned diseases, rendering
bronchodilators to be an effective drug for their treatment
(59). In a previous study on the
effects of Cassia seeds on airway smooth muscle contraction
(60), the ethanolic extract of
Cassia seeds have been found to inhibit the contraction of
airway smooth muscle by inhibiting voltage-dependent
L-type-mediated Ca2+ influx. Further studies have
demonstrated that the main component of the ethanol extract of
Cassia seeds that can induce the relaxation of airway smooth
muscle is AO. Therefore, AO may serve as a viable therapeutic agent
for the treatment of asthma and COPD.
Mosquitoes are vectors of a number of diseases, such
as malaria, dengue fever, dengue shock syndrome and yellow fever
(61,62). Therefore, controlling their
population can control these aforementioned infectious diseases
(63). Generally, control is done
at their larval stages because they are more accessible compared
with adults, where they are more concentrated and less likely to
change their habitat (64). AO has
been shown to be effective for controlling the larvae of
Anopheles gambiae, with a median lethal dose of 1 mg/ml
(65,66). In addition, besides killing this
species of mosquitoes, AO can also protect against cowpea weevil
beetle infestation, which can be applied as a protective agent for
stored cowpea seeds and other crops (67). Furthermore, a previous review of
plant-based insecticides from 2000 to 2018 showed that AO can be
used as a potential larvicide (68).
In addition, AO showed >15% inhibition on the
senescence-associated secretory phenotype (SASP) (69). SASP is a bioactive secretion
produced during senescence of cells which can mediate
non-cell-autonomous effects of senescence (70,71).
Therefore, inhibition of SASP formation may have a role in slowing
down cellular senescence. It has also been shown that AO can
significantly reduce total serum cholesterol levels, triglyceride
and low-density lipoprotein levels in hyperlipidemic rats (12). In Xuezhiling tablets, a Chinese
patented medicine used for the treatment of hyperlipidemia, tests
have identified AO to be one of the active ingredients, which is
associated with the anti-hyperlipidemic effect of Xuezhiling
(13). This suggests AO to be a
potential drug for the treatment of hyperlipidemia. Bate-site
amyloid precursor protein cleaving enzyme-1 (BACE1) inhibitors can
significantly reduce the concentration of cerebrospinal fluid
amyloid plaques of β-protein and are promising drugs for the
treatment of Alzheimer's disease (72). It has been found that AO has a
strong inhibitory effect on BACE1 with IC50 values of
50.9-190 µg/ml (73), which
suggests that AO has some application in the development of drugs
for the prevention and treatment of Alzheimer's disease.
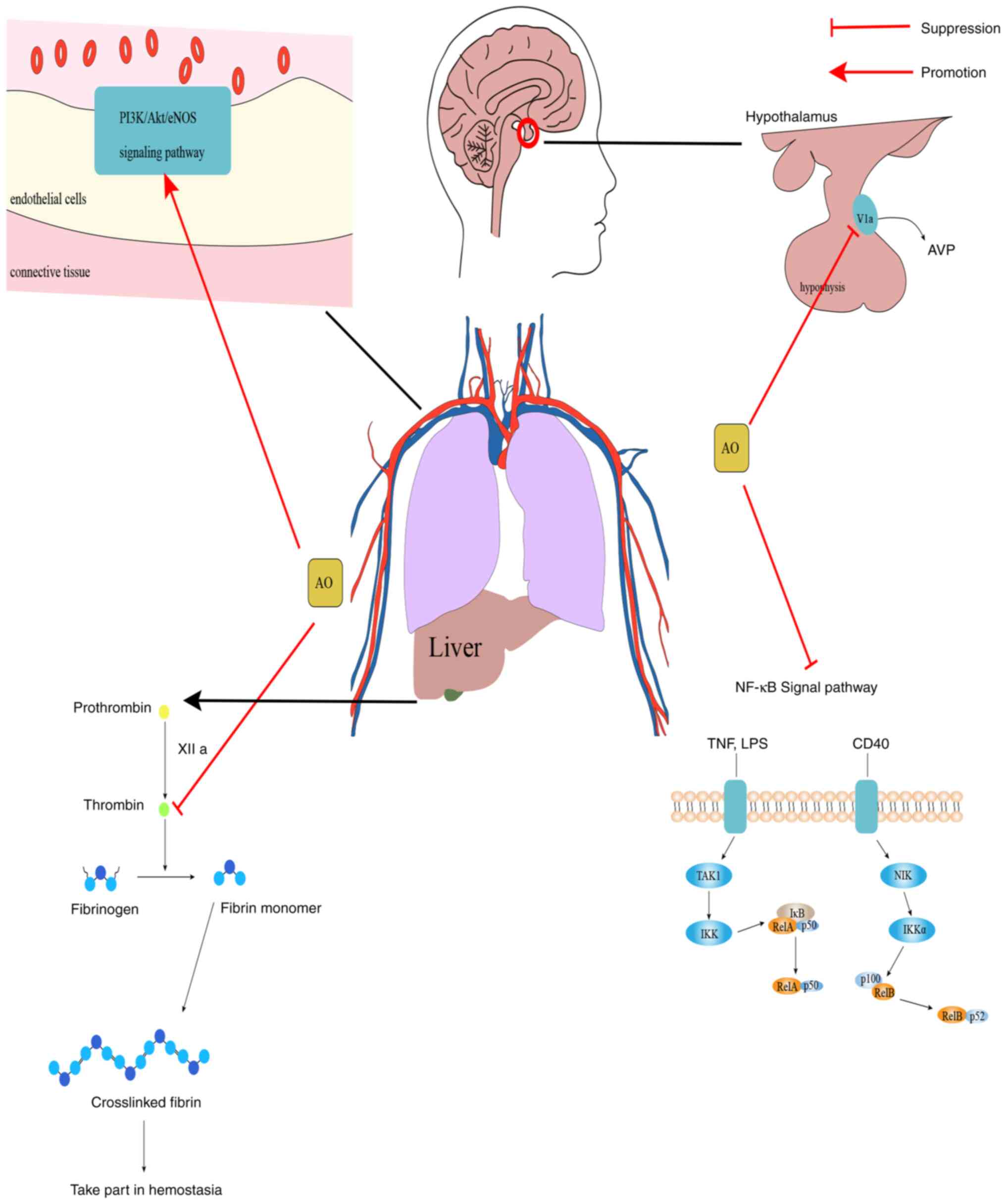
Vasopressin (AVP), a nonapeptide, is mainly
synthesized in the hypothalamic supraoptic nucleus, paraventricular
nucleus and the supraoptic nucleus (74). It can also be produced in other
areas of the brain and organs, such as the medial amygdala, the
nucleus of the terminal bed and adrenal chromophores (74,75).
AVP can only be found in mammals and is involved in the regulation
of blood pressure, water and salt balance, social behavior (such as
learning and cognition) and regulation of emotion (such as anxiety,
fear and depression) (76,77). AVP acts through three different
vasopressin receptors (78):
V1a, V1b and V2 receptors, all of
which belong to different isoforms of G protein-coupled receptors
(79). A previous study has
demonstrated that V1a is the most abundant and widely
distributed vasopressin receptor (80). V1a-knockout mice show a
significant reduction in anxious behavior but also severely
impaired social cognition performance (81,82).
V1a is mainly distributed in the brain and is involved
in regulation of emotional and adaptive behaviors, pain, circadian
rhythm cortisol synthesis and secretion (82).
PI3K/Akt/eNOS is an important regulatory signaling
pathway in endothelial cells, and previous studies have also found
that activation of the PI3K/Akt/eNOS signaling pathway can not only
promote angiogenesis (87,88), improve renal microcirculation
(89) and protect endothelial
cells from injury (90), but can
also suppress diabetes-induced atrial remodeling and atrial
fibrillation (91), and can
improve cardiac function in rats with myocardial infarction
(92). Thus, these data suggest
that the PI3K/Akt/eNOS signaling pathway plays an important role in
regulating vascular activity. In a study investigating the effects
of AO on isolated mesenteric arteries and its mechanism of action,
AO was previously found to have an important role in activating the
PI3K/Akt/eNOS signaling pathway by phosphorylating
Ser473 to activate Akt. This enhanced eNOS activation by
phosphorylating Ser1177 and Thr495 to
stimulate nitric oxide (NO) production in endothelial cells
(93). Therefore, these
observations suggest that AO can serve as a potential
vasodilator.
In summary, AO can exert varying degrees of
inhibitory effects on vasopressin and thrombin signaling, whilst
also conferring agonistic effects on the PI3K/Akt/eNOS and NF-κB
signaling pathways (Fig. 3). These
can potentially be exploited for the development of therapeutic
agents for the corresponding diseases.
Anthraquinones generally have some hepatic and renal
toxicity, where AO is of no exception (103,104). Whilst examining the effects of
oral administration of different doses of AO on hepatotoxicity in
rats, it was previously found that medium (40 mg/kg) and high doses
(200 mg/kg) of AO can cause liver damage (5). Furthermore, in a study in which
Cassia seed aqueous extract was administered orally to rats
at doses of 4.37, 15.75 and 47.30 g/kg for 28 days,
histopathological changes in the livers of male rats (47.30 g/kg
group) and female rats (15.75 and 47.30 g/kg groups) were found; it
was demonstrated that the aqueous extract can induce hepatotoxicity
in rats, where AO was one of the components that caused the
hepatotoxicity (105). AO induces
hepatotoxicity by activating the nucleotide-binding oligomerization
domain, leucine rich repeat and pyrin domain-containing 3
inflammatory vesicle signaling pathway (106), which also causes nephrotoxicity
and colorectal melanosis (107).
In addition to this, AO may increase the toxicity of certain drugs.
AO has been observed to significantly increase the toxicity of
irinotecan compared with glucoaurantio-obtusin (108). Therefore, the biosafety of AO
should be considered when developing it for the treatment of
various diseases in the body.
Not applicable.
Funding: The present study was supported by the Project of
Science and Technology Department of Sichuan Province (grant no.
2021YJ0445), Foundation of Affiliated Hospital of Southwest Medical
University (grant no. 21072) and Innovation and Entrepreneurship
Training Program for Students of Southwest Medical University
(grant no. 202210632176).
Not applicable.
ZW and YL conceived the study. YL and XS designed
the study, drafted, reviewed and edited the manuscript, and
produced all the figures. XH, YX and TL wrote the manuscript. YL,
XS and TL analyzed the relevant literature. All authors read and
approved the final manuscript. Data sharing is not applicable.
Not applicable.
Not applicable.
The authors declare that they have no competing
interests.
|
1
|
Lee GY, Cho BO, Shin JY, Jang SI, Cho IS,
Kim HY, Park JS, Cho CW, Kang JS, Kim JH and Kim YH: Tyrosinase
inhibitory components from the seeds of Cassia tora. Arch Pharm
Res. 41:490–496. 2018.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Luo H, Wu H, Wang L, Xiao S, Lu Y, Liu C,
Yu X, Zhang X, Wang Z and Tang L: Hepatoprotective effects of
Cassiae Semen on mice with non-alcoholic fatty liver disease based
on gut microbiota. Commun Biol. 4(1357)2021.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Kim YJ, Lee S, Jin J, Woo H, Choi YK and
Park KG: Cassiaside C Inhibits M1 polarization of macrophages by
downregulating glycolysis. Int J Mol Sci. 23(1696)2022.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Yang B, Xie L, Peng S, Sun K, Jin J, Zhen
Y, Qin K and Cai B: Nine components pharmacokinetic study of rat
plasma after oral administration raw and prepared Semen Cassiae in
normal and acute liver injury rats. J Sep Sci. 42:2341–2350.
2019.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Xu L, Li J, Tang X, Wang Y, Ma Z and Gao
Y: Metabolomics of aurantio-obtusin-induced hepatotoxicity in rats
for discovery of potential biomarkers. Molecules.
24(3452)2019.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Mugas ML, Calvo G, Marioni J, Céspedes M,
Martinez F, Vanzulli S, Sáenz D, Di Venosa G, Nuñez Montoya S and
Casas A: Photosensitization of a subcutaneous tumour by the natural
anthraquinone parietin and blue light. Sci Rep.
11(23820)2021.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Zou Y, Cao Z, Wang J, Chen X, Chen YQ, Li
Y, Liu J, Zhao Y, Wang A and He B: A Series of Novel HDAC
inhibitors with anthraquinone as a cap group. Chem Pharm Bull
(Tokyo). 68:613–617. 2020.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Watroly MN, Sekar M, Fuloria S, Gan SH,
Jeyabalan S, Wu YS, Subramaniyan V, Sathasivam KV, Ravi S, Mat Rani
NNI, et al: Chemistry, biosynthesis, physicochemical and biological
properties of rubiadin: A promising natural anthraquinone for new
drug discovery and development. Drug Des Devel Ther. 15:4527–4549.
2021.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Li Y, Guo F, Guan Y, Chen T, Ma K, Zhang
L, Wang Z, Su Q, Feng L, Liu Y and Zhou Y: Novel anthraquinone
compounds inhibit colon cancer cell proliferation via the reactive
oxygen Species/JNK Pathway. Molecules. 25(1672)2020.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Szymańska M and Majerz I: Effect of
substitution of hydrogen atoms in the molecules of anthrone and
anthraquinone. Molecules. 26(502)2021.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Panigrahi GK, Verma N, Singh N, Asthana S,
Gupta SK, Tripathi A and Das M: Interaction of anthraquinones of
Cassia occidentalis seeds with DNA and Glutathione. Toxicol Rep.
5:164–172. 2018.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Zhang N, Dong N, Pang L, Xu H and Ji H:
Quantitative determination and pharmacokinetic study of
aurantio-obtusin in rat plasma by liquid chromatography-mass
spectrometry. J Chromatogr Sci. 52:1059–1064. 2014.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Nie C, Zhang F, Ma X, Guo R, Zhou S, Zhao
L, Xu H, Xiao X and Wang Z: Determination of quality markers of
Xuezhiling tablet for hyperlipidemia treatment. Phytomedicine.
44:231–238. 2018.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Kwon KS, Lee JH, So KS, Park BK, Lim H,
Choi JS and Kim HP: Aurantio-obtusin, an anthraquinone from cassiae
semen, ameliorates lung inflammatory responses. Phytother Res.
32:1537–1545. 2018.PubMed/NCBI View
Article : Google Scholar
|
|
15
|
He YQ, Zhou CC, Yu LY, Wang L, Deng JL,
Tao YL, Zhang F and Chen WS: Natural product derived phytochemicals
in managing acute lung injury by multiple mechanisms. Pharmacol
Res. 163(105224)2021.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Ahmed B, Sultana R and Greene MW: Adipose
tissue and insulin resistance in obese. Biomed Pharmacother.
137(111315)2021.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Munafò A, Frara S, Perico N, Di Mauro R,
Cortinovis M, Burgaletto C, Cantarella G, Remuzzi G, Giustina A and
Bernardini R: In search of an ideal drug for safer treatment of
obesity: The false promise of pseudoephedrine. Rev Endocr Metab
Disord. 22:1013–1025. 2021.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Stadler JT and Marsche G: Obesity-Related
changes in high-density lipoprotein metabolism and function. Int J
Mol Sci. 21(8985)2020.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Mongioì LM, La Vignera S, Cannarella R,
Cimino L, Compagnone M, Condorelli RA and Calogero AE: The role of
resveratrol administration in human obesity. Int J Mol Sci.
22(4362)2021.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Mayoral LP, Andrade GM, Mayoral EP, Huerta
TH, Canseco SP, Rodal Canales FJ, Cabrera-Fuentes HA, Cruz MM,
Pérez Santiago AD, Alpuche JJ, et al: Obesity subtypes, related
biomarkers & heterogeneity. Indian J Med Res. 151:11–21.
2020.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Kuder MM and Nyenhuis SM: Optimizing
lifestyle interventions in adult patients with comorbid asthma and
obesity. Ther Adv Respir Dis. 14(1753466620906323)2020.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Purdy JC and Shatzel JJ: The hematologic
consequences of obesity. Eur J Haematol. 106:306–319.
2021.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Ganesan SM, Vazana S and Stuhr S:
Waistline to the gumline: Relationship between obesity and
periodontal disease-biological and management considerations.
Periodontol 2000. 87:299–314. 2021.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Landecho MF, Marin-Oto M, Recalde-Zamacona
B, Bilbao I and Frühbeck G: Obesity as an adipose tissue
dysfunction disease and a risk factor for infections-Covid-19 as a
case study. Eur J Intern Med. 91:3–9. 2021.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Gammone MA and D'Orazio N: COVID-19 and
Obesity: Overlapping of two pandemics. Obes Facts. 14:579–585.
2021.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Guo CY, Liao WT, Qiu RJ, Zhou DS, Ni WJ,
Yu CP and Zeng Y: Aurantio-obtusin improves obesity and insulin
resistance induced by high-fat diet in obese mice. Phytother Res.
35:346–360. 2021.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Yamashita S, Masuda D and Matsuzawa Y:
Pemafibrate, a new selective PPARα modulator: Drug concept and its
clinical applications for dyslipidemia and metabolic diseases. Curr
Atheroscler Rep. 22(5)2020.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Ciavarella C, Motta I, Valente S and
Pasquinelli G: Pharmacological (or Synthetic) and nutritional
agonists of PPAR-γ as candidates for cytokine storm modulation in
COVID-19 disease. Molecules. 25(2076)2020.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Prabhakar PK: Pathophysiology of diabetic
secondary complication and their management. Curr Diabetes Rev.
17:395–396. 2021.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Wang Z, Shen S, Cui Z, Nie H, Han D and
Yan H: Screening and isolating major aldose reductase inhibitors
from the seeds of evening primrose (Oenothera biennis). Molecules.
24(2709)2019.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Julius A, Renuka RR, Hopper W, Babu Raghu
P, Rajendran S, Srinivasan S, Dharmalingam K, Alanazi AM,
Arokiyaraj S and Prasath S: Inhibition of aldose reductase by novel
phytocompounds: A heuristic approach to treating diabetic
retinopathy. Evid Based Complement Alternat Med.
2022(9624118)2022.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Thakur S, Gupta SK, Ali V, Singh P and
Verma M: Aldose Reductase: A cause and a potential target for the
treatment of diabetic complications. Arch Pharm Res. 44:655–667.
2021.PubMed/NCBI View Article : Google Scholar
|
|
33
|
Zhou Y, Chi J, Lv W and Wang Y: Obesity
and diabetes as high-risk factors for severe coronavirus disease
2019 (Covid-19). Diabetes Metab Res Rev. 37(e3377)2021.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Dodda D, Rama Rao A and Veeresham C: In
vitro and in vivo evaluation of pterostilbene for the management of
diabetic complications. J Ayurveda Integr Med. 11:369–375.
2020.PubMed/NCBI View Article : Google Scholar
|
|
35
|
Jang DS, Lee GY, Kim YS, Lee YM, Kim CS,
Yoo JL and Kim JS: Anthraquinones from the seeds of Cassia tora
with inhibitory activity on protein glycation and aldose reductase.
Biol Pharm Bull. 30:2207–2210. 2007.PubMed/NCBI View Article : Google Scholar
|
|
36
|
Younossi ZM, Marchesini G, Pinto-Cortez H
and Petta S: Epidemiology of nonalcoholic fatty liver disease and
nonalcoholic steatohepatitis: Implications for liver
transplantation. Transplantation. 103:22–27. 2019.PubMed/NCBI View Article : Google Scholar
|
|
37
|
Arrese M, Arab JP, Barrera F, Kaufmann B,
Valenti L and Feldstein AE: Insights into nonalcoholic fatty-liver
disease heterogeneity. Semin Liver Dis. 41:421–434. 2021.PubMed/NCBI View Article : Google Scholar
|
|
38
|
American Family Physician: Nonalcoholic
Fatty Liver Disease. https://www.aafp.org/pubs/afp/issues/2020/1115/p603-s1.html.
Accessed April 2, 2023.
|
|
39
|
Makri E, Goulas A and Polyzos SA:
Epidemiology, pathogenesis, diagnosis and emerging treatment of
nonalcoholic fatty liver disease. Arch Med Res. 52:25–37.
2021.PubMed/NCBI View Article : Google Scholar
|
|
40
|
Friedman SL, Neuschwander-Tetri BA,
Rinella M and Sanyal AJ: Mechanisms of NAFLD development and
therapeutic strategies. Nat Med. 24:908–922. 2018.PubMed/NCBI View Article : Google Scholar
|
|
41
|
Zhou F, Ding M, Gu Y, Fan G, Liu C, Li Y,
Sun R, Wu J, Li J, Xue X, et al: Aurantio-Obtusin attenuates
non-alcoholic fatty liver disease through AMPK-Mediated autophagy
and fatty acid oxidation pathways. Front Pharmacol.
12(826628)2021.PubMed/NCBI View Article : Google Scholar
|
|
42
|
Galli SJ, Tsai M and Piliponsky AM: The
development of allergic inflammation. Nature. 454:445–454.
2008.PubMed/NCBI View Article : Google Scholar
|
|
43
|
Hu T, Dong Y, Yang C, Zhao M and He Q:
Pathogenesis of children's allergic diseases: Refocusing the role
of the gut microbiota. Front Physiol. 12(749544)2021.PubMed/NCBI View Article : Google Scholar
|
|
44
|
Yao Y, Chen CL, Yu D and Liu Z: Roles of
follicular helper and regulatory T cells in allergic diseases and
allergen immunotherapy. Allergy. 76:456–470. 2021.PubMed/NCBI View Article : Google Scholar
|
|
45
|
Nguyen SMT, Rupprecht CP, Haque A,
Pattanaik D, Yusin J and Krishnaswamy G: Mechanisms governing
anaphylaxis: Inflammatory cells, mediators, endothelial gap
junctions and beyond. Int J Mol Sci. 22(7785)2021.PubMed/NCBI View Article : Google Scholar
|
|
46
|
Xu-De Z, Bei-Bei G, Xi-Juan W, Hai-Bo L,
Li-Li Z and Feng-Xia L: Serum IgE Predicts difference of population
and allergens in allergic diseases: Data from Weifang City, China.
Mediators Inflamm. 2021(6627087)2021.PubMed/NCBI View Article : Google Scholar
|
|
47
|
Zellweger F and Eggel A: IgE-associated
allergic disorders: Recent advances in etiology, diagnosis, and
treatment. Allergy. 71:1652–1661. 2016.PubMed/NCBI View Article : Google Scholar
|
|
48
|
Kim M, Lim SJ, Lee HJ and Nho CW: Cassia
tora seed extract and its active compound aurantio-obtusin inhibit
allergic responses in IgE-Mediated mast cells and anaphylactic
models. J Agric Food Chem. 63:9037–9046. 2015.PubMed/NCBI View Article : Google Scholar
|
|
49
|
Hou J, Gu Y, Zhao S, Huo M, Wang S, Zhang
Y, Qiao Y and Li X: Anti-Inflammatory effects of aurantio-obtusin
from seed of cassia obtusifolia L. through Modulation of the NF-κB
pathway. Molecules. 23(3093)2018.PubMed/NCBI View Article : Google Scholar
|
|
50
|
Song JY, Casanova-Nakayama A, Möller AM,
Kitamura SI, Nakayama K and Segner H: Aryl hydrocarbon receptor
signaling is functional in immune cells of rainbow trout
(Oncorhynchus mykiss). Int J Mol Sci. 21(6323)2020.PubMed/NCBI View Article : Google Scholar
|
|
51
|
Disner GR, Lopes-Ferreira M and Lima C:
Where the Aryl hydrocarbon receptor meets the microRNAs: Literature
review of the last 10 years. Front Mol Biosci.
8(725044)2021.PubMed/NCBI View Article : Google Scholar
|
|
52
|
Yamashita N, Kanno Y, Yoshikawa M, Ozawa
M, Sanada N, Nemoto K and Kizu R: Polycyclic aromatic hydrocarbons
induce CYP3A5 gene expression via aryl hydrocarbon receptor in
HepG2 cells. J Toxicol Sci. 46:25–29. 2021.PubMed/NCBI View Article : Google Scholar
|
|
53
|
Vogel CFA, Van Winkle LS, Esser C and
Haarmann-Stemmann T: The aryl hydrocarbon receptor as a target of
environmental stressors-Implications for pollution mediated stress
and inflammatory responses. Redox Biol. 34(101530)2020.PubMed/NCBI View Article : Google Scholar
|
|
54
|
Amakura Y, Yoshimura M, Takaoka M, Toda H,
Tsutsumi T, Matsuda R, Teshima R, Nakamura M, Handa H and Yoshida
T: Characterization of natural aryl hydrocarbon receptor agonists
from cassia seed and rosemary. Molecules. 19:4956–4966.
2014.PubMed/NCBI View Article : Google Scholar
|
|
55
|
Xu H, Bin NR and Sugita S: Diverse
exocytic pathways for mast cell mediators. Biochem Soc Trans.
46:235–247. 2018.PubMed/NCBI View Article : Google Scholar
|
|
56
|
Lee EK, Song J, Seo Y, Koh EM, Kim SH and
Jung KJ: Inhibitory Effects of AF-343, a Mixture of Cassia tora L.,
Ulmus pumila L., and Taraxacum officinale, on Compound
48/80-Mediated Allergic Responses in RBL-2H3 Cells. Molecules.
25(2434)2020.PubMed/NCBI View Article : Google Scholar
|
|
57
|
WHO: Global health estimates:leading
causes of death. Cause specific mortality 2000-2019. WHO, Genova,
Switzerland, 2019.
|
|
58
|
Dumitrache MD, Jieanu AS, Scheau C,
Badarau IA, Popescu GDA, Caruntu A, Costache DO, Costache RS,
Constantin C, Neagu M and Caruntu C: Comparative effects of
capsaicin in chronic obstructive pulmonary disease and asthma
(Review). Exp Ther Med. 22(917)2021.PubMed/NCBI View Article : Google Scholar
|
|
59
|
Chen YY, Yu MF, Zhao XX, Shen J, Peng YB,
Zhao P, Xue L, Chen W, Ma LQ, Qin G, et al: Paracetamol inhibits
Ca2+ permeant ion channels and Ca(2+) sensitization
resulting in relaxation of precontracted airway smooth muscle. J
Pharmacol Sci. 142:60–68. 2020.PubMed/NCBI View Article : Google Scholar
|
|
60
|
She YS, Ma LQ, Liu BB, Zhang WJ, Qiu JY,
Chen YY, Li MY, Xue L, Luo X, Wang Q, et al: Semen cassiae extract
inhibits contraction of airway smooth muscle. Front Pharmacol.
9(1389)2018.PubMed/NCBI View Article : Google Scholar
|
|
61
|
Duong V, Lambrechts L, Paul RE, Ly S, Lay
RS, Long KC, Huy R, Tarantola A, Scott TW, Sakuntabhai A and Buchy
P: Asymptomatic humans transmit dengue virus to mosquitoes. Proc
Natl Acad Sci USA. 112:14688–14693. 2015.PubMed/NCBI View Article : Google Scholar
|
|
62
|
Lee H, Halverson S and Ezinwa N:
Mosquito-Borne Diseases. Prim Care. 45:393–407. 2018.PubMed/NCBI View Article : Google Scholar
|
|
63
|
Gan SJ, Leong YQ, Bin Barhanuddin MFH,
Wong ST, Wong SF, Mak JW and Ahmad RB: Dengue fever and insecticide
resistance in Aedes mosquitoes in Southeast Asia: A review. Parasit
Vectors. 14(315)2021.PubMed/NCBI View Article : Google Scholar
|
|
64
|
Fillinger U and Lindsay SW: Larval source
management for malaria control in Africa: Myths and reality. Malar
J. 10(353)2011.PubMed/NCBI View Article : Google Scholar
|
|
65
|
Raman Ibrahim NBB, Puchooa D,
Govinden-Soulange J and Facknath S: Cassia species: a potential
source of biopesticides. Journal of Plant Diseases and Protection.
128:339–351. 2021.
|
|
66
|
Mbatchou VC, Tchouassi DP, Dickson RA,
Annan K, Mensah AY, Amponsah IK, Jacob JW, Cheseto X, Habtemariam S
and Torto B: Mosquito larvicidal activity of Cassia tora seed
extract and its key anthraquinones aurantio-obtusin and obtusin.
Parasit Vectors. 10(562)2017.PubMed/NCBI View Article : Google Scholar
|
|
67
|
Mbatchou VC, Dickson RA, Amponsah IK,
Mensah AY and Habtemariam S: Protection effect of the
anthraquinones, cassiatorin and aurantio-obtusin from seeds of
Senna tora against cowpea weevil attack. Asian Pac J Trop Biomed.
8:98–105. 2018.
|
|
68
|
Piplani M, Bhagwat DP, Singhvi G,
Sankaranarayanan M, Balana-Fouce R, Vats T and Chander S:
Plant-based larvicidal agents: An overview from 2000 to 2018. Exp
Parasitol. 199:92–103. 2019.PubMed/NCBI View Article : Google Scholar
|
|
69
|
Lim H, Park BK, Shin SY, Kwon YS and Kim
HP: Methyl caffeate and some plant constituents inhibit age-related
inflammation: Effects on senescence-associated secretory phenotype
(SASP) formation. Arch Pharm Res. 40:524–535. 2017.PubMed/NCBI View Article : Google Scholar
|
|
70
|
Birch J and Gil J: Senescence and the
SASP: Many therapeutic avenues. Genes Dev. 34:1565–1576.
2020.PubMed/NCBI View Article : Google Scholar
|
|
71
|
Ohtani N: The roles and mechanisms of
senescence-associated secretory phenotype (SASP): Can it be
controlled by senolysis? Inflamm Regen. 42(11)2022.PubMed/NCBI View Article : Google Scholar
|
|
72
|
Scheltens P, De trooper B, Kivipelto M,
Holstege H, Chételat G, Teunissen CE, Cummings J and van der Flier
WM: Alzheimer's disease. Lancet. 397:1577–1590. 2021.PubMed/NCBI View Article : Google Scholar
|
|
73
|
Jung HA, Ali MY, Jung HJ, Jeong HO, Chung
HY and Choi JS: Inhibitory activities of major anthraquinones and
other constituents from Cassia obtusifolia against β-secretase and
cholinesterases. J Ethnopharmacol. 191:152–160. 2016.PubMed/NCBI View Article : Google Scholar
|
|
74
|
Török B, Fazekas CL, Szabó A and Zelena D:
Epigenetic modulation of vasopressin expression in health and
disease. Int J Mol Sci. 22(9415)2021.PubMed/NCBI View Article : Google Scholar
|
|
75
|
Watanabe J, Takayanagi Y, Yoshida M,
Hattori T, Saito M, Kohno K, Kobayashi E and Onaka T: Conditional
ablation of vasopressin-synthesizing neurons in transgenic rats. J
Neuroendocrinol. 33(e13057)2021.PubMed/NCBI View Article : Google Scholar
|
|
76
|
Glavaš M, Gitlin-Domagalska A, Dębowski D,
Ptaszyńska N, Łęgowska A and Rolka K: Vasopressin and its
analogues: From natural hormones to multitasking peptides. Int J
Mol Sci. 23(3068)2022.PubMed/NCBI View Article : Google Scholar
|
|
77
|
Al-Kuraishy HM, Al-Gareeb AI, Qusti S,
Alshammari EM, Atanu FO and Batiha GE: Arginine vasopressin and
pathophysiology of COVID-19: An innovative perspective. Biomed
Pharmacother. 143(112193)2021.PubMed/NCBI View Article : Google Scholar
|
|
78
|
Mondritzki T, Mai TA, Vogel J, Pook E,
Wasnaire P, Schmeck C, Hüser J, Dinh W, Truebel H and Kolkhof P:
Cardiac output improvement by pecavaptan: A novel dual-acting
vasopressin V1a/V2 receptor antagonist in experimental heart
failure. Eur J Heart Fail. 23:743–750. 2021.PubMed/NCBI View Article : Google Scholar
|
|
79
|
Szczepanska-Sadowska E, Wsol A,
Cudnoch-Jedrzejewska A and Żera T: Complementary role of oxytocin
and vasopressin in cardiovascular regulation. Int J Mol Sci.
22(11465)2021.PubMed/NCBI View Article : Google Scholar
|
|
80
|
Che K, Muttenthaler M and Kurzbach D:
Conformational selection of vasopressin upon V1a
receptor binding. Comput Struct Biotechnol J. 19:5826–5833.
2021.PubMed/NCBI View Article : Google Scholar
|
|
81
|
Sparapani S, Millet-Boureima C, Oliver J,
Mu K, Hadavi P, Kalostian T, Ali N, Avelar CM, Bardies M, Barrow B,
et al: The biology of vasopressin. Biomedicines.
9(89)2021.PubMed/NCBI View Article : Google Scholar
|
|
82
|
Lago TR, Brownstein MJ, Page E, Beydler E,
Manbeck A, Beale A, Roberts C, Balderston N, Damiano E, Pineles SL,
et al: The novel vasopressin receptor (V1aR) antagonist SRX246
reduces anxiety in an experimental model in humans: A randomized
proof-of-concept study. Psychopharmacology (Berl). 238:2393–2403.
2021.PubMed/NCBI View Article : Google Scholar
|
|
83
|
Paudel P, Kim DH, Jeon J, Park SE, Seong
SH, Jung HA and Choi JS: Neuroprotective effect of
aurantio-obtusin, a putative vasopressin V(1A) receptor antagonist,
on transient forebrain ischemia mice model. Int J Mol Sci.
22(3335)2021.PubMed/NCBI View Article : Google Scholar
|
|
84
|
Lemmens-Gruber R and Kamyar M: Vasopressin
antagonists. Cell Mol Life Sci. 63:1766–1779. 2006.PubMed/NCBI View Article : Google Scholar
|
|
85
|
Lemmens-Gruber R and Kamyar M:
Pharmacology and clinical relevance of vasopressin antagonists.
Internist (Berl). 49:628. 629–630, 632-4. 2008.PubMed/NCBI View Article : Google Scholar
|
|
86
|
Ripoll GV, Pifano M, Garona J and Alonso
DF: Commentary: Arginine vasopressin receptor 1a is a therapeutic
target for castration-resistant prostate cancer. Front Oncol.
9(1490)2020.PubMed/NCBI View Article : Google Scholar
|
|
87
|
Hu Y, Tao R, Chen L, Xiong Y, Xue H, Hu L,
Yan C, Xie X, Lin Z, Panayi AC, et al: Exosomes derived from
pioglitazone-pretreated MSCs accelerate diabetic wound healing
through enhancing angiogenesis. J Nanobiotechnology.
19(150)2021.PubMed/NCBI View Article : Google Scholar
|
|
88
|
Li S, Wang Y, Wang Z, Chen L, Zuo B, Liu C
and Sun D: Enhanced renoprotective effect of GDNF-modified
adipose-derived mesenchymal stem cells on renal interstitial
fibrosis. Stem Cell Res Ther. 12(27)2021.PubMed/NCBI View Article : Google Scholar
|
|
89
|
Chen C, Sun L, Zhang W, Tang Y, Li X, Jing
R and Liu T: Limb ischemic preconditioning ameliorates renal
microcirculation through activation of PI3K/Akt/eNOS signaling
pathway after acute kidney injury. Eur J Med Res.
25(10)2020.PubMed/NCBI View Article : Google Scholar
|
|
90
|
Chen J, Huang Y, Hu X, Bian X and Nian S:
Gastrodin prevents homocysteine-induced human umbilical vein
endothelial cells injury via PI3K/Akt/eNOS and Nrf2/ARE pathway. J
Cell Mol Med. 25:345–357. 2021.PubMed/NCBI View Article : Google Scholar
|
|
91
|
Xue X, Ling X, Xi W, Wang P, Sun J, Yang Q
and Xiao J: Exogenous hydrogen sulfide reduces atrial remodeling
and atrial fibrillation induced by diabetes mellitus via activation
of the PI3K/Akt/eNOS pathway. Mol Med Rep. 22:1759–1766.
2020.PubMed/NCBI View Article : Google Scholar
|
|
92
|
Song W, Liang Q, Cai M and Tian Z:
HIF-1α-induced up-regulation of microRNA-126 contributes to the
effectiveness of exercise training on myocardial angiogenesis in
myocardial infarction rats. J Cell Mol Med. 24:12970–12979.
2020.PubMed/NCBI View Article : Google Scholar
|
|
93
|
Li S, Li Q, Lv X, Liao L, Yang W, Li S, Lu
P and Zhu D: Aurantio-obtusin relaxes systemic arteries through
endothelial PI3K/AKT/eNOS-dependent signaling pathway in rats. J
Pharmacol Sci. 128:108–115. 2015.PubMed/NCBI View Article : Google Scholar
|
|
94
|
Yu X, Wei LH, Zhang JK, Chen TR, Jin Q,
Wang YN, Zhang SJ, Dou TY, Cao YF, Guo WZ, et al: Anthraquinones
from Cassiae semen as thrombin inhibitors: In vitro and in silico
studies. Phytochemistry. 165(112025)2019.PubMed/NCBI View Article : Google Scholar
|
|
95
|
Tarandovskiy ID, Artemenko EO, Panteleev
MA, Sinauridze EI and Ataullakhanov FI: Antiplatelet agents can
promote two-peaked thrombin generation in platelet rich plasma:
Mechanism and possible applications. PLoS One.
8(e55688)2013.PubMed/NCBI View Article : Google Scholar
|
|
96
|
Varghese R, George Priya Doss C, Kumar RS,
Almansour AI, Arumugam N, Efferth T and Ramamoorthy S:
Cardioprotective effects of phytopigments via multiple signaling
pathways. Phytomedicine. 95(153859)2022.PubMed/NCBI View Article : Google Scholar
|
|
97
|
Alkarithi G, Duval C, Shi Y, Macrae FL and
Ariëns RAS: Thrombus structural composition in cardiovascular
disease. Arterioscler Thromb Vasc Biol. 41:2370–2383.
2021.PubMed/NCBI View Article : Google Scholar
|
|
98
|
Wu H, Wang Y, Zhang Y, Xu F, Chen J, Duan
L, Zhang T, Wang J and Zhang F: Breaking the vicious loop between
inflammation, oxidative stress and coagulation, a novel
anti-thrombus insight of nattokinase by inhibiting LPS-induced
inflammation and oxidative stress. Redox Biol.
32(101500)2020.PubMed/NCBI View Article : Google Scholar
|
|
99
|
Zhao Y, Xie R, Yodsanit N, Ye M, Wang Y,
Wang B, Guo LW, Kent KC and Gong S: Hydrogen peroxide-responsive
platelet membrane-coated nanoparticles for thrombus therapy.
Biomater Sci. 9:2696–2708. 2021.PubMed/NCBI View Article : Google Scholar
|
|
100
|
Williams LM and Gilmore TD: Looking Down
on NF-κB. Mol Cell Biol. 40:e00104–20. 2020.PubMed/NCBI View Article : Google Scholar
|
|
101
|
Harrold AP, Cleary MM, Bharathy N, Lathara
M, Berlow NE, Foreman NK, Donson AM, Amani V, Zuercher WJ and
Keller C: In vitro benchmarking of NF-κB inhibitors. Eur J
Pharmacol. 873(172981)2020.PubMed/NCBI View Article : Google Scholar
|
|
102
|
Son M, Wang AG, Tu HL, Metzig MO, Patel P,
Husain K, Lin J, Murugan A, Hoffmann A and Tay S: NF-κB responds to
absolute differences in cytokine concentrations. Sci Signal.
14(eaaz4382)2021.PubMed/NCBI View Article : Google Scholar
|
|
103
|
Yang B, Xie Y, Guo M, Rosner MH, Yang H
and Ronco C: Nephrotoxicity and Chinese Herbal Medicine. Clin J Am
Soc Nephrol. 13:1605–1611. 2018.PubMed/NCBI View Article : Google Scholar
|
|
104
|
Liu Y, Mapa MST and Sprando RL: Liver
toxicity of anthraquinones: A combined in vitro cytotoxicity and in
silico reverse dosimetry evaluation. Food Chem Toxicol.
140(111313)2020.PubMed/NCBI View Article : Google Scholar
|
|
105
|
Yang J, Zhu A, Xiao S, Zhang T, Wang L,
Wang Q and Han L: Anthraquinones in the aqueous extract of Cassiae
semen cause liver injury in rats through lipid metabolism disorder.
Phytomedicine. 64(153059)2019.PubMed/NCBI View Article : Google Scholar
|
|
106
|
Hu M, Lin L, Liu J, Zhong Y, Liang B,
Huang Y, Li Z, Lin X, Wang B, Zhang B, et al: Aurantio-obtusin
induces hepatotoxicity through activation of NLRP3 inflammasome
signaling. Toxicol Lett. 354:1–13. 2022.PubMed/NCBI View Article : Google Scholar
|
|
107
|
Wang J, Zhao Y, Xiao X, Li H, Zhao H,
Zhang P and Jin C: Assessment of the renal protection and
hepatotoxicity of rhubarb extract in rats. J Ethnopharmacol.
124:18–25. 2009.PubMed/NCBI View Article : Google Scholar
|
|
108
|
Yu J, Han JC and Gao YJ: Biotransformation
of glucoaurantio-obtusin towards aurantio-obtusin increases the
toxicity of irinotecan through increased inhibition towards SN-38
glucuronidation. Phytother Res. 28:1577–1580. 2014.PubMed/NCBI View Article : Google Scholar
|