Introduction
Hypoxia is a common pathological process involving
abnormal changes in the morphological structure, function and
metabolism of tissue due to the lack of oxygen or clinically, to
the barrier of oxygen use (1–3).
Hypoxia of major organs, such as the brain and heart (4–7),
often results in the death of the organism. The combination of the
extent, speed, and duration of hypoxia and the organism's metabolic
state determine the influence of hypoxia on the organism. Acute
severe hypoxia often causes organ compensatory insufficiency,
metabolic disorders, irreversible damage and even death (8–10),
and chronic mild hypoxia often causes slight compensatory reactions
(11–13).
Brain hypoxia often causes severe changes to the
internal and external environments of cells, often leading to
necrosis or apoptosis and also often activating self-repair
mechanisms in response to hypoxic injury (14,15). Recently, although neuronal
apoptosis has been well documented at the molecular level, the
mechanism remains unknown and the chaos of apoptosis may be closely
associated with the genesis and development of multiple nervous
system diseases, such as stroke (16,17), cerebral injury (18,19), neurologic tumors and
neurodegenerative recession diseases (20–23).
Neuro-oncological ventral antigen (Nova) is one of
the first validated mammalian neuron-specific splicing factors, and
has been identified as an antigen of paraneoplastic opsoclonus
myoclonus ataxia (POMA) (24,25). Members of the Nova superfamily,
including Nova1 and Nova2, are a type of RNA binding protein that
is expressed only in the central nervous system and that can bind
the YCAY motif of pre-mRNA of target genes both in vivo and
in vitro via its KH domain, thereby regulating alternative
splicing (26,27). Based on the differences in the
target binding positions, Nova has a dual function as a splicing
regulating factor, both promoting and inhibiting splicing. Nova can
recruit other splicing factors when bound to the YCAY motif of
pre-mRNA to promote the assembly of the spliceosome, or it can
indirectly enhance the assembly of the spliceosome at weak splicing
sites (28). In addition, due to
steric hindrance, Nova can also inhibit the assembly of the
spliceosome (25). Within the
Nova superfamilly, Nova1 was initially speculated to regulate
alternative splicing due to its KH domain, similar to hnRNP K and
MER1. Nova1 is highly evolutionarily conserved and its amino acid
homology among human, rat and mouse is 99%, indicating its
significant function in organismal biology (29). As demonstrated by several studies,
Nova1 can regulate the alternative splicing of neuronal
transcripts, a process that is closely associated with all types of
brain physiology and biochemistry, as well as complex nerve systems
and elaborate regulatory processes (30). We also showed that Nova1 may
mediate neuronal responsiveness after ischemia-reperfusion insults
in the rat brain (29).
Therefore, to ascertain whether Nova1 is involved in hypoxia injury
and has a protective effect on neural cells after hypoxia injury,
the eukaryotic expression vector pCMV-Myc-Nova1 was
constructed. The results showed that Nova1 may protect against
hypoxia-induced apoptosis in PC12 cells via Bax/Bcl-2/caspase-3
pathway. As expected, after overexpression of Nova1 protein in PC12
cells subjected to 48 h hypoxia, the rate of apoptosis was
significantly decreased, the mRNA and protein expression levels of
Bax and caspase-3 were significantly decreased, and the expression
of Bcl-2 was significantly increased, indicating that Nova1
protects against the hypoxia-induced apoptosis of PC12 cells via
the Bax/Bcl-2/caspase-3 pathway.
Materials and methods
Construction of eukaryotic expression
vector pCMV-Myc-Nova1
Primers based on the full-length cDNA sequence of
Nova1 (NM_002515.2) were designed according to GeneBank data
(http://www.ncbi.nlm.nih.gov/genbank)
and chemically synthesized by Invitrogen (Carlsbad, CA, USA) as
follows: pU, 5′-TACGTCGACTATGATGGCGGCAGCTCCC-3′
(the SalI restriction site is underlined), and pD,
5′-TCCCTCGAGTCAACCCACTTTCTGAGG-3′
(the XhoI restriction site is underlined). Subsequently, a
Nova1 cDNA fragment (1,503 bp) with SalI and
XhoI restriction endonuclease sites was amplified from
pCR4-TOPO-Nova1 by polymerase chain reaction (PCR) using the
above-designed primers and then subcloned into the eukaryotic
expression vector pCMV-Myc followed by direct sequencing.
Culturing of PC12 cells and
transfection
PC12 cells purchased from the American Type Culture
Collection (ATCC; Manassas, VA, USA) (CRL-1721™) were thawed from
storage in liquid nitrogen using Dulbecco's modified Eagle's medium
(DMEM) with 100 ml/l horse serum and 50 ml/l fetal bovine serum.
The cells were then cultured to the logarithmic growth phase in a
CO2-incubator with 5% CO2 at 37°C. After
digestion with 0.25% Trypsin, the cell solution was diluted and
incubated in a 6-well plate (3×105 cells/well) followed
by culturing in a CO2-incubator with 5% CO2
at 37°C. When the cells reached ~80–90% confluence, a total of 2
µg of pCMV-Myc and pCMV-Myc-Nova1 plasmids was
transfected into PC12 cells using Lipofectamine 2000 (Invitrogen)
according to the manufacturer's instructions. First, 2 µg of
plasmids, including pCMV-Myc and pCMV-Myc-Nova1, was mixed
with 50 µl of Opti-MEM serum-free medium (solution A; Gibco,
Grand Island, NY, USA). Then, 5 µl of Lipofectamine 2000 was
mixed with 50 µl of Opti-MEM serum-free medium (solution B)
and incubated at room temperature (RT) for 5 min. Finally, solution
A and solution B were mixed and incubated at RT for 20 min, after
which they were added to 6-well plates for transfection. At 24, 48,
72 and 96 h after transfection, the cells were collected for
extraction of total RNA and total proteins as described below.
Extraction of total RNA
PC12 cells were washed twice (3 min each) with
phosphate-buffered saline (PBS) to extract the total RNA according
to the manufacturer's instructions. RNAiso Plus (9108; Takara,
Otsu, Japan) was added to a 6-well plate (500 µl/well) and
mixed, followed by being transferred to a new Eppendorf (EP) tube
and incubated at 4°C for 5 min and then centrifuged at 12,000 rpm
for 10 min. The supernatant was transferred to a new EP tube; 100
µl of chloroform was added followed by vortexing for 15 sec,
incubation at RT for 3 min and centrifugation at 12,000 rpm for 15
min. The upper water phase was transferred to a new EP tube, and
250 µl of isopropanol was added and mixed. Then, the tube
was placed in a −20°C refrigerator for 1–2 h and centrifuged at
12,000 rpm for 20 min at 4°C. The supernatant was discarded, and 1
ml of 75% ethanol was added, followed by centrifugation at 12,000
rpm for 5 min at 4°C, and this step was repeated once. The EP tube
was incubated at RT for 5–10 min until the residual liquid had
completely volatilized, and the pellet was dissolved with 20
µl DEPC ddH2O. The purity and the concentration
of RNA were measured using ultraviolet spectroscopy.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR) assay
The above-extracted total RNA was used as a template
in a reverse transcription reaction using a ReverTra Ace qPCR RT
kit (FSQ-101; Toyobo, Osaka, Japan), according to the
manufacturer's instructions. The reaction mixture, including 10
µl of 2X loading buffer, 1.2 µl of oligo(dT), 2
µl of RNA, 0.2 µl of MMLV, and 6.6 µl of DEPC
ddH2O, was prepared and reacted at 65°C for 30 min, then
at 42°C for 30 min, and then at 85°C for 10 min. Subsequently, a
total of 100 ng of cDNA was used as the template in a RT-qPCR
reaction using the SYBR® Premix Ex Taq™ kit (DRR420A;
Takara), according to the manufacturer's instructions. The reaction
mixture, including 10 µl of 2X master mix, 0.1 µl of
forward primer, 0.1 µl of reverse primer, 2 µl of
cDNA, 0.4 µl of Taq DNA polymerase, and 7.4 µl
of ddH2O, was prepared and used to performed RT-qPCR
according to the following program: one cycle of 95°C for 3 min; 40
cycles of 95°C for 12 sec, 62°C for 30 sec, and 72°C for 30 sec,
using primers as follows: Nova1, pU, 5′-GGTCTCAGCCAAGCAGCAGCAA-3′
and pD, 5′-TTGCAGCAGTAGCAGCAGCCAG-3′; glyceraldehyde 3-phosphate
dehydrogenase (GAPDH), pU, 5′-TGGATCTGACATGCCGCCTGGA-3′ and pD,
5′-AGGTCCACCACCCTGTTGCTGT-3′. The results were analyzed using SDS
1.4 software (Applied Biosystems, Foster City, CA, USA) based on
the 2−ΔΔCt method, and histogram analysis was performed
using Origin 9.5 software (http://www.originlab.com/).
Extraction of total protein
PC12 cells were washed with PBS twice (3 min each)
to extract the total proteins according to the manufacturer's
instructions [RIPA lysis buffer (P0013C); Beyotime, China]. Then,
50 µl cell lysis buffer [50 mmol/l Tris-Cl (pH 6.8), 2% SDS,
10% glycerol, 10 mM PMSF] was added, followed by incubation for 30
min on ice. Cells were collected using a cell scraper, and then
transferred to an EP tube and centrifuged at 12,000 rpm for 15 min
at 4°C. The supernatant was collected, and the protein
concentration was measured using BCA protein assay reagent (Thermo
Fisher Scientific Inc., Waltham, MA, USA).
Western blot assay
Samples of equal mass (35 µg) were
fractionated by electrophoresis through 12% polyacrylamide gels and
transferred to polyvinylidene difluoride (PVDF) membranes (GE
Healthcare, Piscataway, NJ, USA) according to the manufacturer's
instructions. The membranes were probed with rabbit-derived
anti-Nova1 polyclonal antibody (HPA004155; 1:500 in TBST; Sigma,
St. Louis, MO, USA) and mouse-derived anti-Bax (BA0315)/Bcl-2
(BA0412)/caspase-3 (BM3957)/GAPDH (BM1985) monoclonal antibody
(1:500 in TBST) for 1.5 h at room temperature, followed by an
HRP-conjugated goat anti-rabbit (BA1054) or anti-mouse (BA1050)
secondary antibody (1:5,000 in TBST) (both from Wuhan Boster
Biological Engineering Co., Ltd., Wuhan, China). Incubation was
then carried out at RT for 1 h. After that, the chemiluminescent
substrate luminal reagent (GE) and exposure to X-ray film were used
to examine the immunolabeled bands. The optical densities of the
bands were scanned and quantified using ImageJ software version
1.46 (http://rsb.info.nih.gov/ij/).
Cell immunocytochemistry assay
PC12 cells were plated on polylysine-coated
converslips in 6-well plates at an inoculation density of
2×105/ml and were cultured in a CO2-incubator
with 5% CO2 at 37°C for 15–30 min, and then 1 ml of DMEM
was added and cells were cultured overnight. The following day, the
plates were placed on ice, and the medium was discarded. The cells
were washed 3 times (5 min each) with PBS. Cells were fixed with 4%
paraformaldehyde for 30 min and then washed once with
ddH2O (5 min) and once with PBS (5 min). Next, 3%
H2O2 was added, incubation at RT for 30 min,
and the cells were washed 3 times with PBS. Then, 0.3% Triton X-100
was applied, followed by incubation at RT for 30 min. Three washes
(5 min each) with PBS and incubation with 5% BSA for 20 min at RT
were carried out. The slides were then placed in a cassette,
incubated overnight with rabbit-derived anti-Nova1 polyclonal
antibody at 4°C, and washed 3 times (5 min each) with PBS.
Subsequently, the slides were treated with biotinylated goat
anti-rabbit IgG, and incubated at 37°C for 20 min followed by 3
washes with PBS. SABC reagent was applied for 20 min of incubation
at 37°C, followed by 3 washes (5 min each) with PBS and development
of DAB. The slides were dehydrated, cleared and mounted, and images
were captured by microscopy. The average optical density was
calculated using ImageJ software version 1.46 (http://rsb.info.nih.gov/ij/), and histogram analysis
was performed using Origin 9.5 software (http://www.originlab.com/).
Establishment of a hypoxia model
PC12 cells were diluted, inoculated into 96-well
plates at 5×104/ml, and cultured in a
CO2-incubator with 5% CO2 at 37°C for 24 h.
Then, the plates were placed in a hypoxic chamber, and a gas
mixture of 95% N2 and 5% CO2 was pumped in
for 1–2 min. The air outlet was closed, and then a gas mixture of
95% N2 and 5% CO2 was pumped in for 4 min.
Then, the air inlet was closed, and cells were incubated in a
CO2 incubator with 5% CO2 at 37°C for
different durations of hypoxia, 0, 2, 4, 6, 8, 12, 24, 48 and 72 h.
Cells were collected for cell proliferation assays using methyl
thiazolyl tetrazolium (MTT) and apoptosis assays using flow
cytometry as follows. In addition, images of cells at 0 and 48 h of
hypoxia were captured using an inverted microscope.
To examine the effect hypoxia on the expression of
Nova1, PC12 cells transfected with pCMV-Myc or
pCMV-Myc-Nova1 were subjected to 48 h of hypoxia. Then the
mRNA expression of Nova1 was examined by RT-qPCR, and the protein
expression of Nova1 was examined by western blot analysis, and the
distribution of Nova1 in PC12 cells was examined using
immunocytochemistry. In addition, the apoptosis rate was examined
by flow cytometry (Ex=488 nm, Em=530 nm). The mRNA expression
levels of Bax, Bcl-2 and caspase-3 were examined by RT-qPCR and WB
using the following primers: Bax, pU, 5′-CCAAGAAGCTGAGCGAGTGTC-3′
and pD, 5′-TGAGGACTCCAGCCACAAAGA-3′; Bcl-2, pU,
5′-TGAACCGGCATCTGCACAC-3′ and pD, 5′-CGTCTTCAGAGACAGCCAGGAG-3′;
caspase-3, pU, 5′-GTGGAACTGACGATGATATGGC-3′ and pD,
5′-CGCAAAGTGACTGGATGAACC-3′; GAPDH, pU,
5′-TGGATCTGACATGCCGCCTGGA-3′ and pD,
5′-AGGTCCACCACCCTGTTGCTGT-3′.
Methyl thiazolyl tetrazolium (MTT)
assay
After hypoxia as described above, 20 µl of
MTT (final concentration, 5 mg/ml) was added to the cells, followed
by incubation at 37°C for 4 h. The supernatant was then discarded;
cells were treated with 150 µl of dimethyl sulfoxide (DMSO)
to shock them for 5 min, and the optical density (OD) at 490 nm was
recorded using a microplate reader, with cell-free conditions as a
control. The cellular survival rate was calculated based on the
following formula: Cellular survival rate = (OD490 of
hypoxia group/OD490 of control group) × 100%.
Flow cytometric assay of apoptosis
Apoptosis was examined using the Annexin v-fitc
apoptosis kit (BU-AP101; Biouniquer, San Diego, CA, USA) according
to the manufacturer's instructions. After digestion with 0.25%
trypsin, PC12 cells were washed twice with PBS and then centrifuged
at 2,000 rpm for 5 min. Next, the cells were resuspended in 500
µl Annexin V binding buffer, and 5 µl of Annexin
V-FITC was added to each sample, followed by the addition of 5
µl PI and reaction at RT for 10 min in the dark.
Subsequently, the apoptosis rate was examined using flow cytometry
(Ex=488 nm, Em=530 nm) and analyzed using Origin 9.5 software
(http://www.originlab.com/).
Statistical analysis
All data are expressed as the mean ± SD. Statistical
analysis was performed using SPSS software (version 21.0) (SPSS,
Inc., Chicago, IL, USA) (http://spss.en.softonic.com/). In the Student's
t-test, p<0.05 and p<0.01 indicate significant and highly
significant differences, respectively.
Results
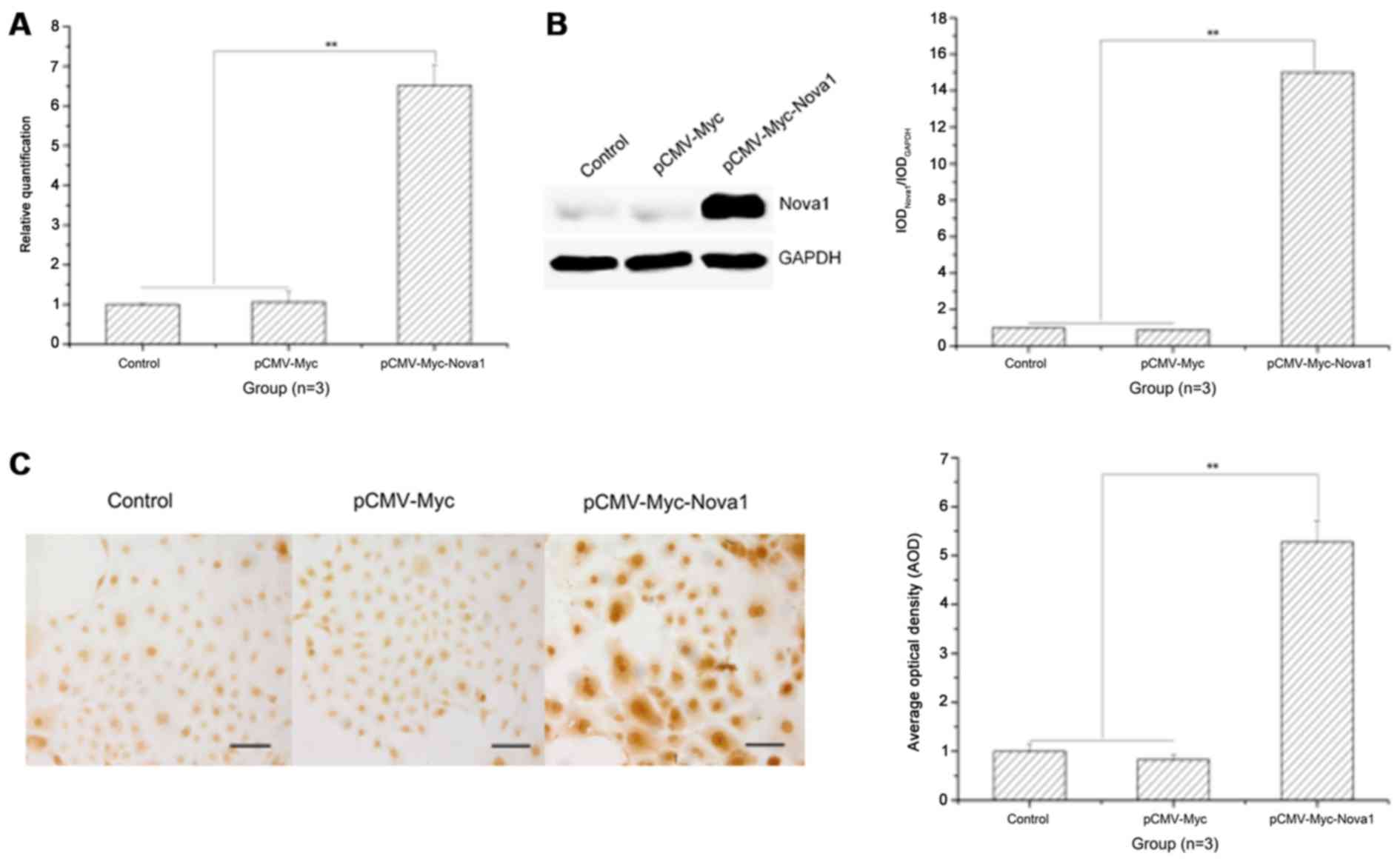
mRNA and protein expression levels of
Nova1 are significantly increased in PC12 cells after transfection
with pCMV-Myc-Nova1, and Nova1 is widely distributed in the
cytoplasm and nucleus
To identify the time of optimal expression of Nova1
in PC12 cells, Nova1 mRNA and protein expression was examined by
RT-qPCR, western blot analysis and cell immunocytochemistry at 24,
48, 72 and 96 h after transfection, and the optimal expression time
was 48 h after transfection (data not shown). At 48 h after
transfection with pCMV-Myc-Nova1, the mRNA and protein
expression levels of Nova1 were significantly increased compared to
those in the control and pCMV-Myc groups (p<0.01) (Fig. 1A and B), and Nova1 protein was
also increased in the cytoplasm and nucleus (Fig. 1C).
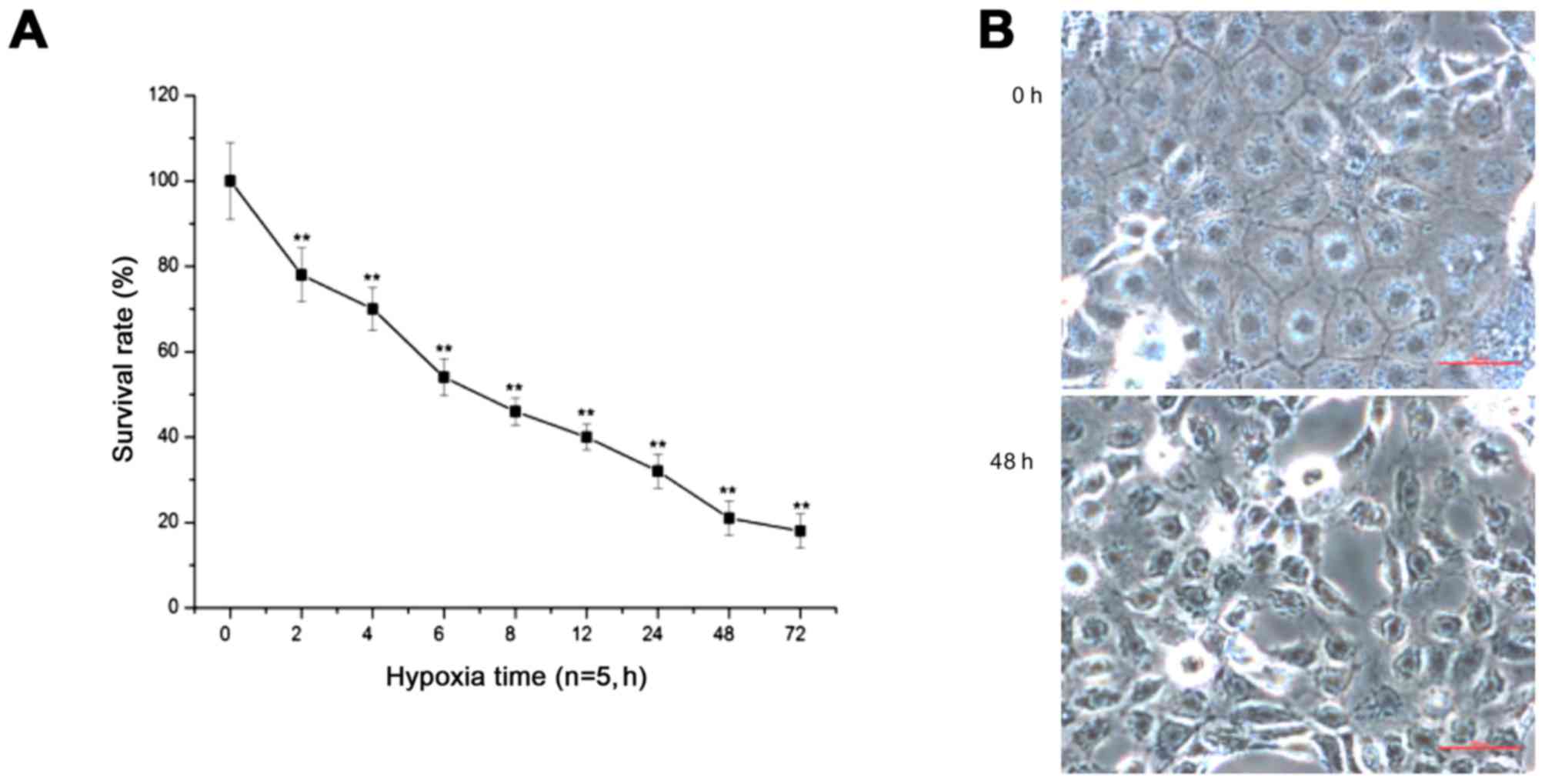
Prolonged hypoxia significantly decreases
the cell survival rate and increases the apoptosis rate
With prolonged duration of hypoxia, the survival
rate of PC12 cells was significantly decreased (p<0.01)
(Fig. 2A). After 48 h of hypoxia,
the cells showed shrinkage, the intercellular space was wider, and
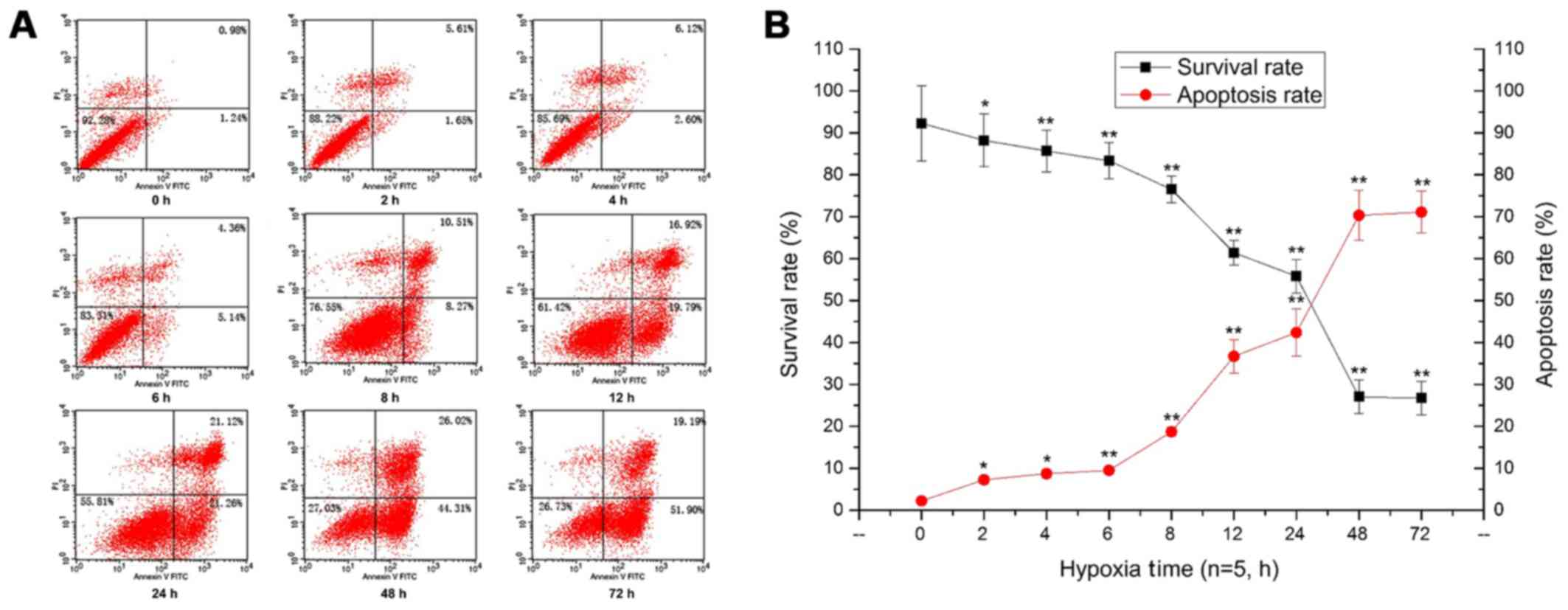
some cells had detached, floating in the medium (Fig. 2B). Additionally, the flow
cytometry results showed that the survival rate of PC12 cells was
significantly decreased, and the apoptosis rate of PC12 cells was
constantly increased with prolonged hypoxia, peaking at 48 h of
hypoxia (p<0.05, p<0.01) (Fig.
3).
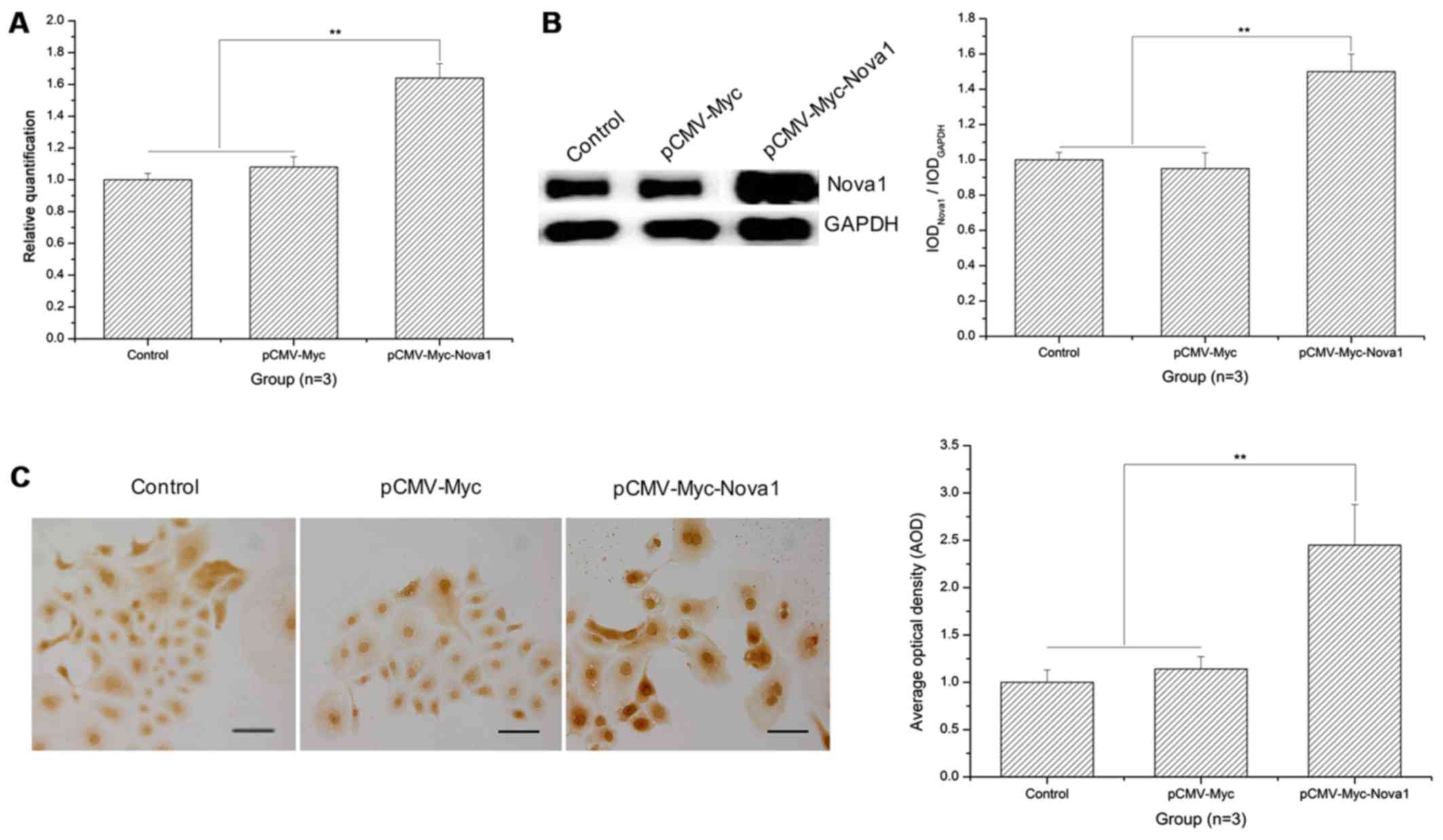
mRNA and protein expression levels of
Nova1 are significantly increased in PC12 cells following
transfection with pCMV-Myc-Nova1 after 48 h of hypoxia and the
level of Nova1 is higher than the control groups
As shown in the RT-qPCR results, after 48 h of
hypoxia, the mRNA expression of Nova1 was significantly increased
in the PC12 cells following transfection with pCMV-Myc-Nova1
in contrast to the control and pCMV-Myc groups (p<0.01)
(Fig. 4A). Similarly, as shown by
the western blot analysis and cell immunocytochemistry results, the
protein expression of Nova1 was also significantly increased in the
PC12 cells transfected with pCMV-Myc-Nova1 in contrast to
the control and pCMV-Myc groups, and the level of Nova1 was much
higher than that in the control and pCMV-Myc groups (p<0.01)
(Fig. 4B and C).
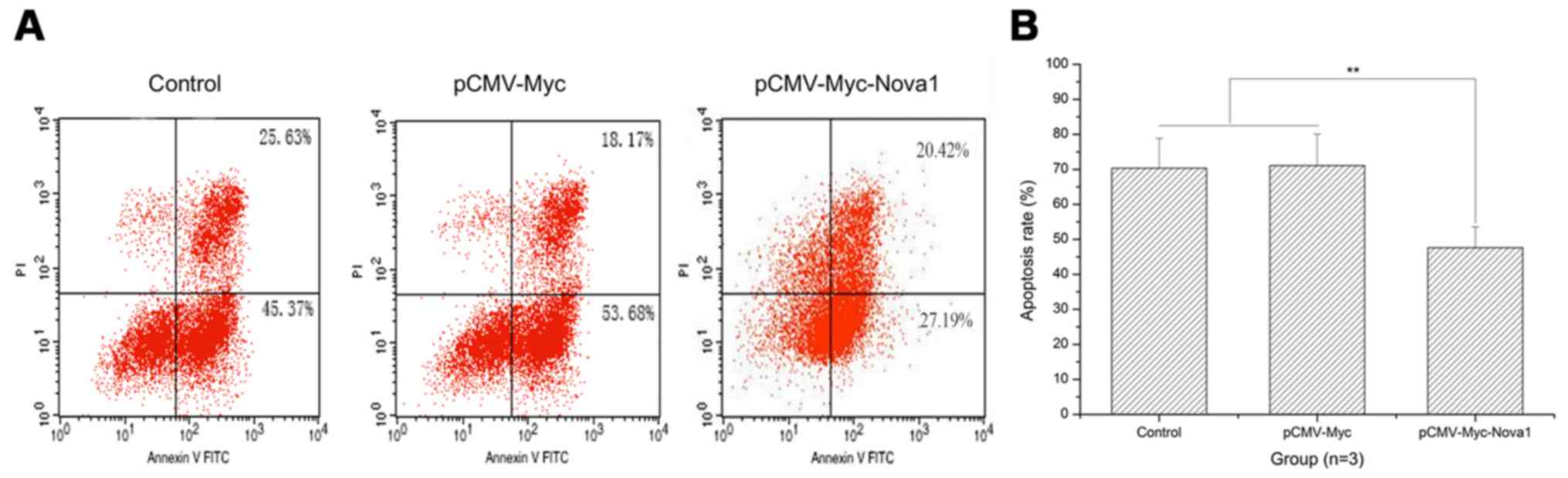
The apoptosis rate after 48 h of hypoxia
is significantly decreased in PC12 cells following transfection
with pCMV-Myc-Nova1
As shown by the flow cytometry results, after 48 h
of hypoxia, the apoptosis rate of PC12 cells was significantly
decreased by transfection with pCMV-Myc-Nova1 compared with
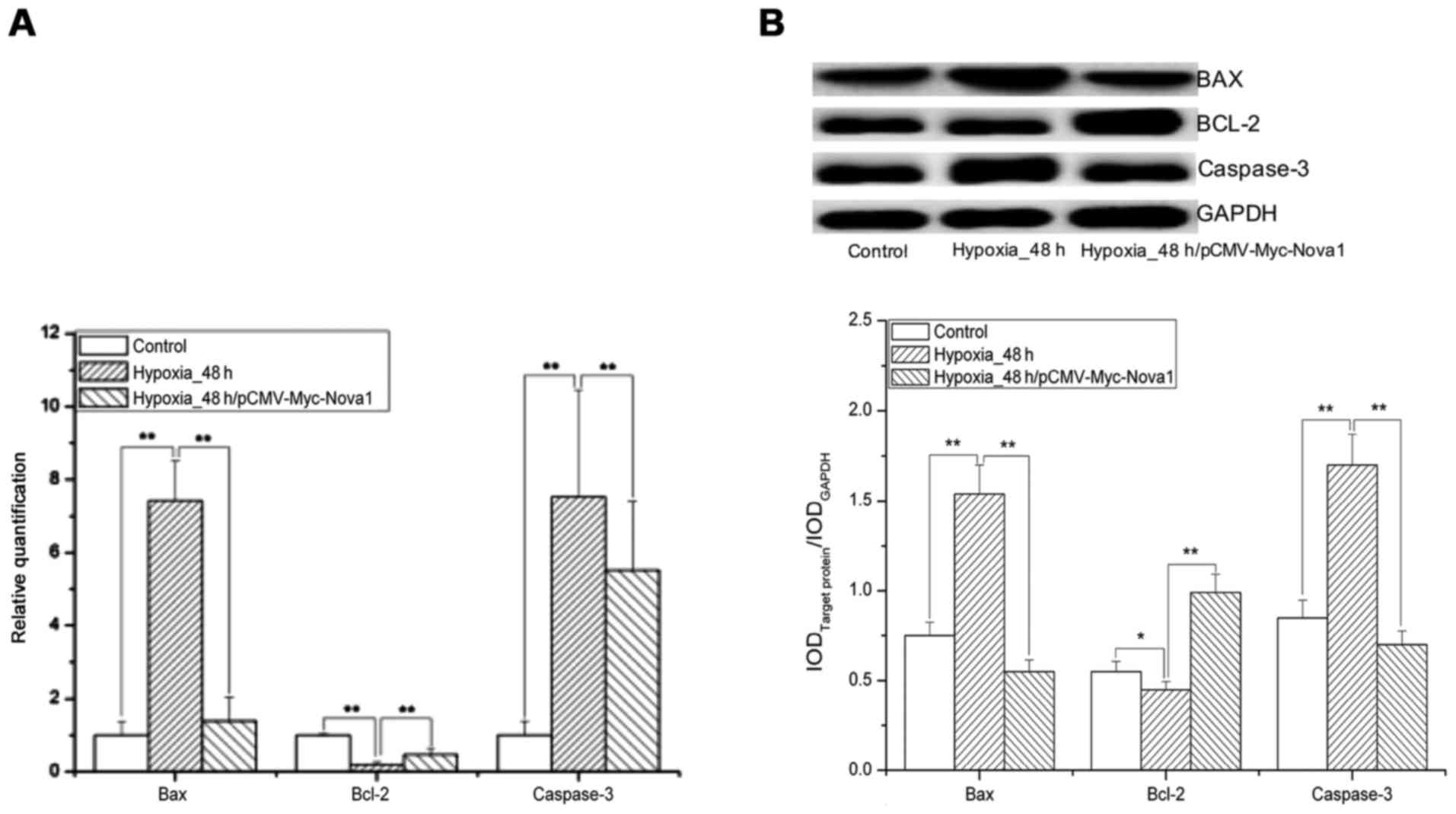
the control and pCMV-Myc groups (p<0.01) (Fig. 5). Significantly, after 48 h of
hypoxia, the mRNA and protein expression levels of Bax and
caspase-3 were significantly increased in the pCMV-Myc group in
comparison to the control and significantly decreased in the
pCMV-Myc-Nova1 group in comparison to the pCMV-Myc group
(p<0.01) (Fig. 6).
Furthermore, the mRNA and protein expression levels of Bcl-2 were
significantly decreased in the pCMV-Myc group in comparison to the
control, and significantly increased in the pCMV-Myc-Nova1
group in comparison to the pCMV-Myc group (p<0.01) (Fig. 6). These results indicate that
Nova1 induces resistance to hypoxia-induced apoptosis in PC12 cells
via the Bax/Bcl-2/caspase-3 pathway.
Discussion
The present study demonstrated that Nova1 was highly
expressed in PC12 cells at 48 h after transfection and was widely
distributed in the cytoplasm and nucleus. After continuous hypoxia
of untransfected PC12 cells, the cell survival rate was gradually
decreased, and the apoptosis rate was gradually increased compared
to the control. Upon 48 h of hypoxia in PC12 cells overexpressing
Nova1, the cell apoptosis rate was significantly decreased and the
survival rate was significantly increased. The mRNA and protein
expression levels of Bax and caspase-3 were also significantly
decreased, and the mRNA and protein expression levels of Bcl-2 were
significantly increased, indicating that Nova1 could induce
resistance to hypoxia-induced apoptosis in PC12 cells via the
Bax/Bcl-2/caspase-3 pathway.
Recently, alternative splicing has been regarded as
a major mechanism for increasing proteomic complexity and protein
function diversity, and alternative splicing has been shown to play
a significant role in the regulation of gene expression and protein
function diversity (31–33). Alternative splicing is much more
common in the brain than in other organs (34,35), and it is also often involved in
ischemia-reperfusion injury to the brain as described in our
previous study (29). However,
the mechanism by which Nova1 participates in ischemia-reperfusion
injury of the brain remains unknown. To address this question, the
eukaryotic expression vector pCMV-Myc was chosen for the expression
of Nova1 cDNA. pCMV-Myc contains a strong promoter from
cytomegalovirus (CMV) that can transcribe exogenous genes early and
shows strong expression efficiency. Regarding the regulation of the
CMV promoter, at 48 h after transfection with
pCMV-Myc-Nova1, the mRNA and protein expression levels of
Nova1 were significantly increased, and Nova1 was also increased in
the cytoplasm and nucleus.
The brain accounts for only 2% of the body weight in
humans but uses ~15% of the cardiac output and ~20% of oxygen
consumption of the whole body. In addition, hypoxia produces even
greater damage in the brain than in other tissues, and it severely
affects the normal physiological functions of various organs
(36,37). In the study of hypoxic injury of
the brain and its associated diseases, stabilized neuronal hypoxia
models have often been established in vitro, including the
PC12 hypoxia model using a hypoxic chamber. PC12 cells are derived
from rat pheochromocytoma cells, separated from a rat
pheochromocytoma by Greene and Tischler in 1976. They show many
characteristics of nerve cells (38). PC12 cells are therefore often used
as an ideal in vitro cell model for nervous system diseases
(39–41). Here, PC12 cells were subjected to
oxygen deprivation using a hypoxic chamber to mimic hypoxia, and
the cell survival and apoptosis rates were examined by MTT assay
and flow cytometry, respectively. After continuous hypoxia, the
cell survival rate was gradually declined, and the apoptosis rate
was steadily increased by hypoxia in a time-dependent manner. These
results indicated that the PC12 cell hypoxia model was correctly
established. After hypoxia, Nova1 protein expression was
significantly increased, indicating that hypoxia could induce the
strong splicing activity of Nova1 and that cells may activate a
compensatory reaction. In addition, Nova1 protein was widely
expressed in the cytoplasm and nuclei, especially in the nuclei
indicating that Nova1 may be involved in the processing of RNAs in
neurons.
Several studies have documented that apoptosis is
induced both in the delayed neuronal cell death after transient
brain hypoxia and by acute focal brain ischemia (42–46). Normally, the anti-apoptosis and
pro-apoptosis proteins are metastable in organisms, and it requires
several pro-apoptosis elements to induce apoptosis. Bcl-2 is an
anti-apoptosis protein and can regulate apoptosis by binding
pro-apoptosis proteins, such as Bax, Bad or Bcl-xl; therefore the
ratio of Bcl-2 to Bax has been regarded as a critical factor in
cell survival or death (47,48). In this study, after 48 h of
hypoxia, the mRNA expression level of Bcl-2 was significantly
decreased, that of Bax was obviously increased, and the ratio of
Bcl-2 to Bax was decreased. This indicated that the pro-apoptotic
effect of Bax was greater than the anti-apoptotic effect of Bcl-2,
finally resulting in apoptosis. After transfection of
pCMV-Myc-Nova1, the mRNA and protein expression levels of
Bcl-2 were increased, levels of Bax were significantly decreased,
and the ratio of Bcl-2 to Bax was increased, thus indicating that
Nova1 could protect against hypoxia-induced neuronal injury by
regulating the ratio of Bcl-2 to Bax. Additionally, in comparison
to the control, the mRNA and protein expression levels of caspase-3
were significantly increased after 48 h of hypoxia and
significantly decreased upon transfection of pCMV-Myc-Nova1,
indicating that Nova1 may be involved in regulating hypoxia-induced
apoptosis via the caspase-3 pathway.
In conclusion, the eukaryotic expression vector
pCMV-Myc-Nova1 was constructed and transfected into the PC12
cells during hypoxia. Our study demonstrated that Nova1 is
associated with resistance to the hypoxia-induced apoptosis of PC12
cells via the Bax/Bcl-2/caspase-3 pathway. This discovery provides
evidence of Nova1 to explore the novel mechanism during
hypoxia.
Abbreviations:
|
Nova1
|
neuro-oncological ventral antigen
1
|
|
POMA
|
paraneoplastic opsoclonus myoclonus
ataxia
|
|
Bax
|
Bcl-2 associated gene X
|
|
Bcl-2
|
B cell lymphoma gene-2
|
|
caspase-3
|
cysteinyl aspartic acid specific
protease-3
|
|
MTT
|
methyl thiazolyl tetrazolium
|
Acknowledgments
We would like to thank Gorospe Myraim of the
National Institute on Aging (NIA) (National Institutes of Health,
Bethesda, MD, USA) for the antibody and plasmid pCMV-myc. This
study was supported by the National Science Foundation of China
(nos. 81671056, 81372237 and 81100862) and the Yangzhou University
and CSC (no. 2015CXJ070).
References
|
1
|
Belo AI, van Vliet SJ, Maus A, Laan LC,
Nauta TD, Koolwijk P, Tefsen B and van Die I: Hypoxia inducible
factor 1α down regulates cell surface expression of
α1,2-fucosylated glycans in human pancreatic adenocarcinoma cells.
FEBS Lett. 589:2359–2366. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Chen HC, Lee JT, Shih CP, Chao TT, Sytwu
HK, Li SL, Fang MC, Chen HK, Lin YC, Kuo CY, et al: Hypoxia induces
a metabolic shift and enhances the stemness and expansion of
cochlear spiral ganglion stem/progenitor cells. Biomed Res Int.
2015:3595372015.PubMed/NCBI
|
|
3
|
de Oliveira JT, Ribeiro C, Barros R, Gomes
C, de Matos AJ, Reis CA, Rutteman GR and Gärtner F: Hypoxia
up-regulates galectin-3 in mammary tumor progression and
metastasis. PLoS One. 10:e01344582015. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Bonestroo HJ, Nijboer CH, van Velthoven
CT, van Bel F and Heijnen CJ: The neonatal brain is not protected
by osteopontin peptide treatment after hypoxia-ischemia. Dev
Neurosci. 37:142–152. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Cui C, Zhou T, Li J, Wang H, Li X, Xiong
J, Xu P and Xue M: Proteomic analysis of the mouse brain after
repetitive exposure to hypoxia. Chem Biol Interact. 236:57–66.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Kimura W, Xiao F, Canseco DC, Muralidhar
S, Thet S, Zhang HM, Abderrahman Y, Chen R, Garcia JA, Shelton JM,
et al: Hypoxia fate mapping identifies cycling cardiomyocytes in
the adult heart. Nature. 523:226–230. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Siebenmann C, Rasmussen P, Sørensen H,
Bonne TC, Zaar M, Aachmann-Andersen NJ, Nordsborg NB, Secher NH and
Lundby C: Hypoxia increases exercise heart rate despite combined
inhibition of β-adrenergic and muscarinic receptors. Am J Physiol
Heart Circ Physiol. 308:H1540–H1546. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Calbet JA, Boushel R, Rådegran G,
Søndergaard H, Wagner PD and Saltin B: Determinants of maximal
oxygen uptake in severe acute hypoxia. Am J Physiol Regul Integr
Comp Physiol. 284:R291–R303. 2003. View Article : Google Scholar
|
|
9
|
Gonchar O and Mankovskaya I: Effect of
moderate hypoxia/reoxygenation on mitochondrial adaptation to acute
severe hypoxia. Acta Biol Hung. 60:185–194. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Sanotskaya NV, Matsievskii DD and Lebedeva
MA: Effect of picrotoxin on organism's resistance to acute severe
hypoxia. Bull Exp Biol Med. 145:177–180. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Beltran-Parrazal L, Acuna D, Ngan AM, Kim
E, Ngan A, Kawakami K, Edmond J and Lopez IA: Neuroglobin,
cytoglobin, and transcriptional profiling of hypoxia-related genes
in the rat cerebellum after prenatal chronic very mild carbon
monoxide exposure (25 ppm). Brain Res. 1330:61–71. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Dore-Duffy P, Wencel M, Katyshev V and
Cleary K: Chronic mild hypoxia ameliorates chronic inflammatory
activity in myelin oligodendrocyte glycoprotein (MOG) peptide
induced experimental autoimmune encephalomyelitis (EAE). Adv Exp
Med Biol. 701:165–173. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Lima-Ojeda JM, Vogt MA, Richter SH,
Dormann C, Schneider M, Gass P and Inta D: Lack of protracted
behavioral abnormalities following intermittent or continuous
chronic mild hypoxia in perinatal C57BL/6 mice. Neurosci Lett.
577:77–82. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Hu Q, Wu C, Chen JY, Yan F, Li JR and Chen
G: The relationship between hypoxia-inducible factor-1α expression
and apoptosis in early brain injury after subarachnoid hemorrhage.
Zhejiang Da Xue Xue Bao Yi Xue Ban. 43:58–65. 2014.In Chinese.
PubMed/NCBI
|
|
15
|
Xuan M, Okazaki M, Iwata N, Asano S,
Kamiuchi S, Matsuzaki H, Sakamoto T, Miyano Y, Iizuka H and Hibino
Y: Chronic treatment with a water-soluble extract from the culture
medium of Ganoderma lucidum mycelia prevents apoptosis and
necroptosis in hypoxia/ischemia-induced injury of type 2 diabetic
mouse brain. Evid Based Complement Alternat Med. 2015:8659862015.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Liu B, Zhang YH, Jiang Y, Li LL, Chen Q,
He GQ, Tan XD and Li CQ: Gadd45b is a novel mediator of neuronal
apoptosis in ischemic stroke. Int J Biol Sci. 11:353–360. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Lv H, Wang L, Shen J, Hao S, Ming A, Wang
X, Su F and Zhang Z: Salvianolic acid B attenuates apoptosis and
inflammation via SIRT1 activation in experimental stroke rats.
Brain Res Bull. 115:30–36. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Fang L, Gao H, Zhang W, Zhang W and Wang
Y: Resveratrol alleviates nerve injury after cerebral ischemia and
reperfusion in mice by inhibiting inflammation and apoptosis. Int J
Clin Exp Med. 8:3219–3226. 2015.PubMed/NCBI
|
|
19
|
Zhao P, Zhou R, Zhu XY, Hao YJ, Li N, Wang
J, Niu Y, Sun T, Li YX and Yu JQ: Matrine attenuates focal cerebral
ischemic injury by improving antioxidant activity and inhibiting
apoptosis in mice. Int J Mol Med. 36:633–644. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Daulatzai MA: Death by a thousand cuts in
Alzheimer's disease: Hypoxia - the prodrome. Neurotox Res.
24:216–243. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Liu H, Qiu H, Yang J, Ni J and Le W:
Chronic hypoxia facilitates Alzheimer's disease through
demethylation of γ-secretase by downregulating DNA
methyltransferase 3b. Alzheimers Dement. 12:130–143. 2016.
View Article : Google Scholar
|
|
22
|
Schmid D, Fay F, Small DM, Jaworski J,
Riley JS, Tegazzini D, Fenning C, Jones DS, Johnston PG, Longley
DB, et al: Efficient drug delivery and induction of apoptosis in
colorectal tumors using a death receptor 5-targeted nanomedicine.
Mol Ther. 22:2083–2092. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Walter RF, Werner R, Ting S, Vollbrecht C,
Theegarten D, Christoph DC, Schmid KW, Wohlschlaeger J and
Mairinger FD: Identification of deregulation of apoptosis and cell
cycle in neuroendocrine tumors of the lung via NanoString nCounter
expression analysis. Oncotarget. 6:24690–24698. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Darnell RB and Posner JB: Paraneoplastic
syndromes involving the nervous system. N Engl J Med.
349:1543–1554. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Ratti A, Fallini C, Colombrita C, Pascale
A, Laforenza U, Quattrone A and Silani V: Post-transcriptional
regulation of neuro-oncological ventral antigen 1 by the neuronal
RNA-binding proteins ELAV. J Biol Chem. 283:7531–7541. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Buckanovich RJ and Darnell RB: The
neuronal RNA binding protein Nova-1 recognizes specific RNA targets
in vitro and in vivo. Mol Cell Biol. 17:3194–3201. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Zhang YA, Zhu JM, Yin J, Tang WQ, Guo YM,
Shen XZ and Liu TT: High expression of neuro-oncological ventral
antigen 1 correlates with poor prognosis in hepatocellular
carcinoma. PLoS One. 9:e909552014. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Dredge BK and Darnell RB: Nova regulates
GABA(A) receptor gamma2 alternative splicing via a distal
downstream UCAU-rich intronic splicing enhancer. Mol Cell Biol.
23:4687–4700. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Li H, Sun C, Wang Y, Gao Y, Liu Y, Gao Y,
Li X and Zhang C: Dynamic expression pattern of neuro-oncological
ventral antigen 1 (Nova1) in the rat brain after focal cerebral
ischemia/reperfusion insults. J Histochem Cytochem. 61:45–54. 2013.
View Article : Google Scholar :
|
|
30
|
Dredge BK, Stefani G, Engelhard CC and
Darnell RB: Nova autoregulation reveals dual functions in neuronal
splicing. EMBO J. 24:1608–1620. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Nishida A, Minegishi M, Takeuchi A, Awano
H, Niba ET and Matsuo M: Neuronal SH-SY5Y cells use the
C-dystrophin promoter coupled with exon 78 skipping and display
multiple patterns of alternative splicing including two intronic
insertion events. Hum Genet. 134:993–1001. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Stepankiw N, Raghavan M, Fogarty EA,
Grimson A and Pleiss JA: Widespread alternative and aberrant
splicing revealed by lariat sequencing. Nucleic Acids Res.
43:8488–8501. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Stephan-Otto Attolini C, Peña V and
Rossell D: Designing alternative splicing RNA-seq studies. Beyond
generic guidelines. Bioinformatics. 31:3631–3637. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Schreiner D, Nguyen TM, Russo G, Heber S,
Patrignani A, Ahrné E and Scheiffele P: Targeted combinatorial
alternative splicing generates brain region-specific repertoires of
neurexins. Neuron. 84:386–398. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Trifonov S, Yamashita Y, Kase M, Maruyama
M and Sugimoto T: Glutamic acid decarboxylase 1 alternative
splicing isoforms: Characterization, expression and quantification
in the mouse brain. BMC Neurosci. 15:1142014. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Cantagrel S, Krier C, Ducrocq S, Bodard S,
Payen V, Laugier J, Guilloteau D and Chalon S: Hypoxic
preconditioning reduces apoptosis in a rat model of immature brain
hypoxia-ischaemia. Neurosci Lett. 347:106–110. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Williamson DJ, Ejaz S, Sitnikov S, Fryer
TD, Sawiak SJ, Burke P, Baron JC and Aigbirhio FI: A comparison of
four PET tracers for brain hypoxia mapping in a rodent model of
stroke. Nucl Med Biol. 40:338–344. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Greene LA and Tischler AS: Establishment
of a noradrenergic clonal line of rat adrenal pheochromocytoma
cells which respond to nerve growth factor. Proc Natl Acad Sci USA.
73:2424–2428. 1976. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Dichter MA, Tischler AS and Greene LA:
Nerve growth factor-induced increase in electrical excitability and
acetylcholine sensitivity of a rat pheochromocytoma cell line.
Nature. 268:501–504. 1977. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
McGuire JC, Greene LA and Furano AV: NGF
stimulates incorporation of fucose or glucosamine into an external
glycoprotein in cultured rat PC12 pheochromocytoma cells. Cell.
15:357–365. 1978. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Rudy B, Kirschenbaum B and Greene LA:
Nerve growth factor-induced increase in saxitoxin binding to rat
PC12 pheochromocytoma cells. J Neurosci. 2:1405–1411.
1982.PubMed/NCBI
|
|
42
|
Gutsaeva DR, Carraway MS, Suliman HB,
Demchenko IT, Shitara H, Yonekawa H and Piantadosi CA: Transient
hypoxia stimulates mitochondrial biogenesis in brain subcortex by a
neuronal nitric oxide synthase-dependent mechanism. J Neurosci.
28:2015–2024. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Huang NC, Yongbi MN and Helpern JA: The
influence of preischemic hyperglycemia on acute changes in brain
water ADCw following focal ischemia in rats. Brain Res.
788:137–143. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Liu KF, Li F, Tatlisumak T, Garcia JH,
Sotak CH, Fisher M and Fenstermacher JD: Regional variations in the
apparent diffusion coefficient and the intracellular distribution
of water in rat brain during acute focal ischemia. Stroke.
32:1897–1905. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Roehl AB, Zoremba N, Kipp M, Schiefer J,
Goetzenich A, Bleilevens C, Kuehn-Velten N, Tolba R, Rossaint R and
Hein M: The effects of levosimendan on brain metabolism during
initial recovery from global transient ischaemia/hypoxia. BMC
Neurol. 12:812012. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Shi Q, Zhang P, Zhang J, Chen X, Lu H,
Tian Y, Parker TL and Liu Y: Adenovirus-mediated brain-derived
neurotrophic factor expression regulated by hypoxia response
element protects brain from injury of transient middle cerebral
artery occlusion in mice. Neurosci Lett. 465:220–225. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Carpio MA, Michaud M, Zhou W, Fisher JK,
Walensky LD and Katz SG: BCL-2 family member BOK promotes apoptosis
in response to endoplasmic reticulum stress. Proc Natl Acad Sci
USA. 112:7201–7206. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Lee HJ, Lee EK, Seo YE, Shin YH, Kim HS,
Chun YH, Yoon JS, Kim HH, Han MY, Kim CK, et al: Roles of Bcl-2 and
caspase-9 and -3 in CD30-induced human eosinophil apoptosis. J
Microbiol Immunol Infect. 50:145–152. 2017. View Article : Google Scholar
|