1. Polyamine metabolism
Cellular polyamines
Polyamines (PAs) are ubiquitous, short, positively
charged aliphatic amines essential for normal cell growth,
proliferation and differentiation. Although PAs occur in
prokaryotic and eukaryotic cells, from plants and animals (1,2),
the typical PAs synthesized by mammals are spermidine (Spd), and
spermine (Spm), from the diamine putrescine (Put), (3). Some studies have demonstrated an
alternative pathway of PA synthesis, where agmatine (Agm) can
slightly contribute as a potential source of Put (4-6).
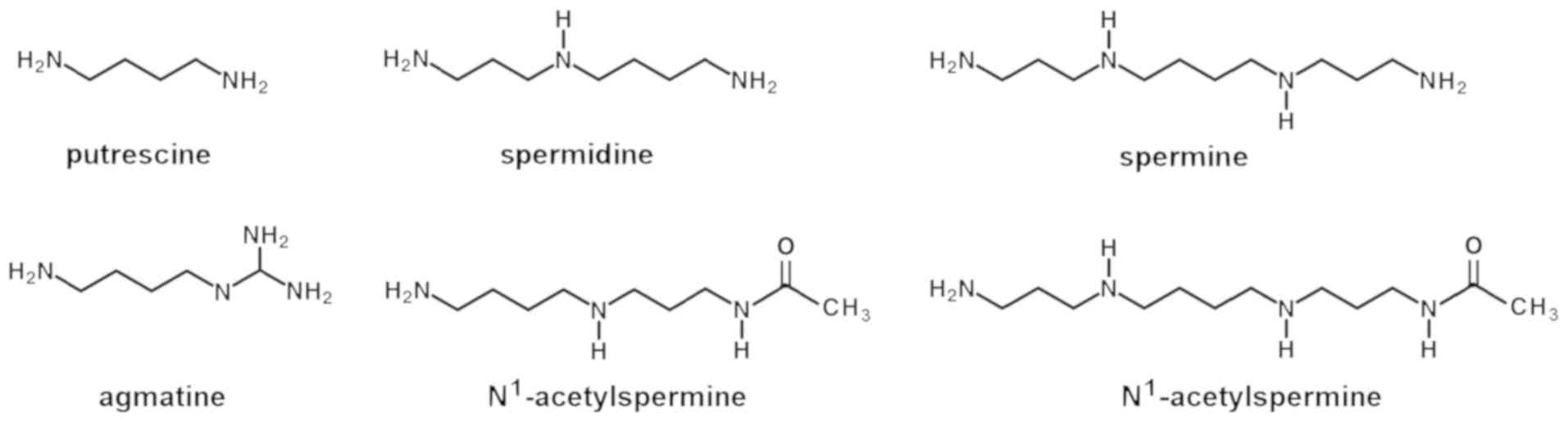
Agmatine, Put, Spd, Spm and the acetylated derivatives
N1-acetylspermidine (AcSpd) and N1-acetylspermine (AcSpm; Fig. 1) have fundamental roles in
cellular homeostasis and their optimal cellular content is
maintained via complex regulatory mechanisms (7). Among the various functional roles of
the natural PAs, the regulation of gene transcription and
post-transcriptional modifications, as well as the modulation of
synaptic activity are noticeable. For this reason, these molecules
are continually under active investigation in wide-ranging research
areas, from neuroscience, cancer, cell differentiation and
development (8-10).
Polyamine interconvertion pathway
Two step-limiting enzymes tightly regulate PA
biosynthesis: ornithine decarboxylase (ODC), which produces Put by
decarboxylation of ornithine and S-adenosylmethionine
decarboxylase, which synthesizes S-adenosylmethioninamine from
S-adenosyl-L-methionine. Two specific aminopropyl transferases, Spd
synthase and Spm synthase are also involved in the biosynthesis of
PAs, adding the aminopropyl group to Put and Spd, respectively
(11-13). Conversely, PA catabolism is finely
regulated by the three enzymes: Spd and Spm N1-acetyltransferase
(SSAT), N1-acetylpolyamine oxidase (PAOX) and spermine oxidase
(SMOX) (Fig. 2). The enzyme SSAT
transfers acetyl groups from acetyl-coenzyme A to the N1 position
of both Spd and Spm to produce AcSpd and AcSpm, respectively. These
substrates are oxidized by the flavoprotein PAOX to produce Spd and
Put, respectively, and 3-aceto-aminopropanal (3-AP) and hydrogen
peroxide (H2O2) (14,15). Another enzyme involved in the PA
catabolism is SMOX that specifically oxidases Spm producing Spd,
3-aminopropanal and H2O2 (15,16). Polyamines can be biosynthesized
and imported into cells by ubiquitous PA transporters (PTSs). As a
result of the multiple enzymes involved in the PA metabolism and
taking also into account the cellular uptake and export of PAs, the
individual content of each PA is constantly well buffered within
the mammalian cells. The typical PTS is saturable, strongly energy
dependent and has a high affinity for its substrates (generally
ranging from 10−7 to 10−5 M, depending on the
PA species and cell type) (17).
Polyamines content can be regulated by internalization through
plasma membrane PA permeases (17,18). Notwithstanding the strong cellular
PA homeostasis, this metabolism can be altered by physiological
stimuli, as well as in many pathological conditions like cancer,
inflammation, neurodegeneration and others (9). Mammalian cell PA metabolism is
summarized in Fig. 2.
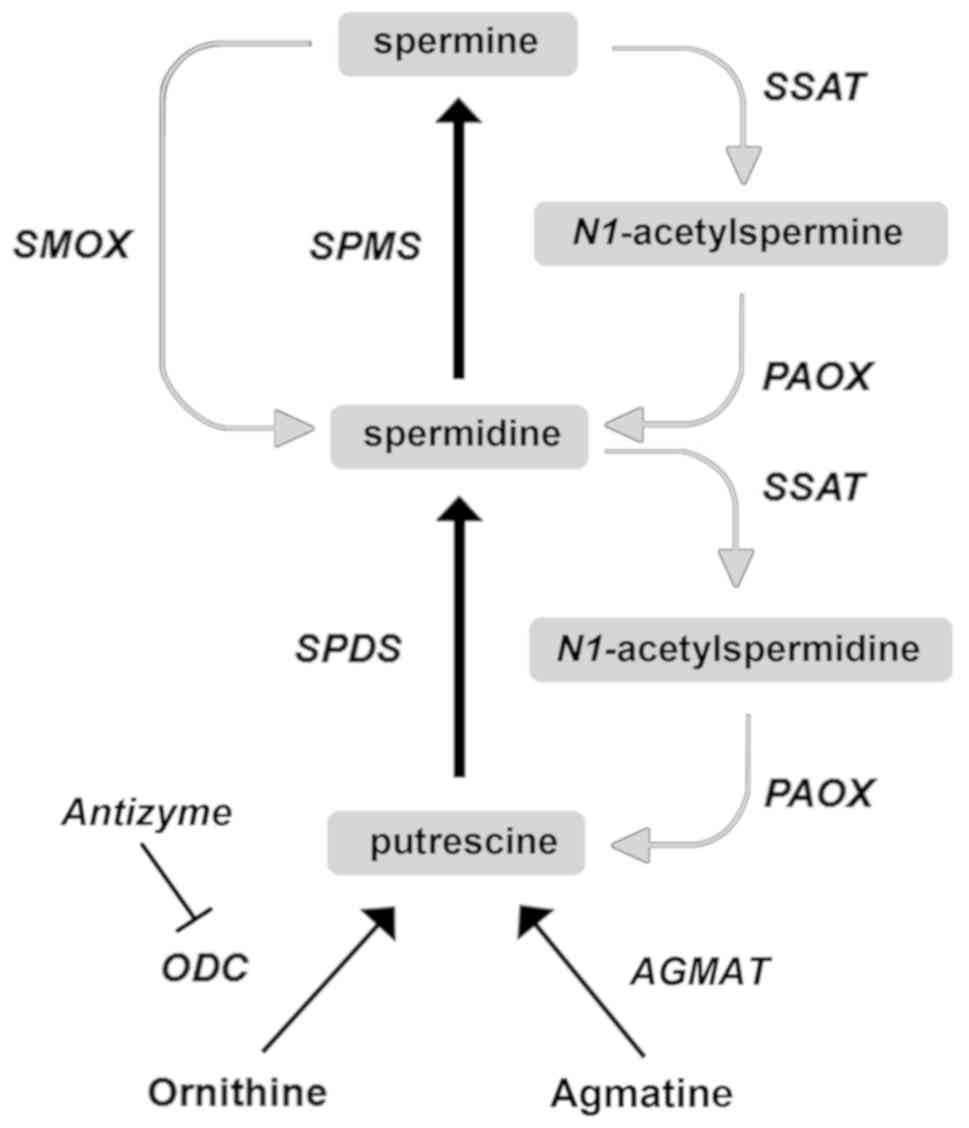 | Figure 2Mammalian PA metabolism. Schematic
representation of PA metabolism showing enzyme network and
substrate interconversion pathways. Anabolic and catabolic pathways
are indicated by black and grey arrows, respectively. AGMT,
agmatinase; ODC, ornithine decarboxylase; PAOX,
N1-acetylpolyamine oxidase; SAMDC,
S-adenosyl-methionine decarboxylase; SMOX, spermine oxidase; SPMS,
spermine synthase; SPDS, spermidine synthase; SSAT, Spd and Spm
N1-acetyltransferase; PA, polyamine. |
2. Polyamines and modulation of ion
channels
Polyamines and the brain
The concentration of PAs varies greatly in different
tissues. In vertebrates, the brain behaves as an autonomous and
closed system with regard to the PA metabolism. There is a high
variation in PA levels between the different brain regions and in
general, the levels of Spd and Spm are much higher than those of
Put (19). Different studies have
shown a neuroprotective role of PAs, nevertheless in pathological
conditions they can cause neurotoxicity due to their oxidation and
conversion into aldehydes and reactive oxygen species (20-23). In recent years, the key role of
PAs in different brain syndromes and diseases has been increasingly
recognized. This role seems to be mainly due to their ability to
modulate and regulate different ion channels (24).
Ion channel modulation by polyamines
Endogenous PAs, in particular Spm, are able to
interact and modulate different ion channels and receptors involved
in the maintenance of homeostasis of calcium, sodium and potassium.
Intracellular Spm, at µM concentrations, acts as an
important blocker of inwardly rectifying potassium channels, by
specifically plugging the ion channel pore. In fact, an increase of
its content is responsible for an increase of the channel gating
and rectification, which leads to a rise in cellular excitability
(25). Polyamines can also
modulate the activities of ion channels responsible for the flux of
cations through the cellular membrane. Additionally, PAs, by
different intra/extra cellular interaction sites, can modulate
voltage activated calcium channels. A specific interaction exists
between PAs and the a1 subunit of L-type calcium channel. Only Put
induces an increase in the flux of current via the protein kinase C
pathway, while Spd and Spd are unable to produce this effect
(26). These channels are
involved in synaptic transmission and plasticity and respond to
ligands such as glutamate. Ionotropic glutamate receptors are
divided into three groups: N-methyl-D-aspartate receptor (NMDAR);
α-amino-3- hydroxy-5-methyl-4-isoxazolepropionic acid receptor
(AMPAR) and kainate receptor (KAR). Intracellular PAs can block the
pore of the AMPAR and KAR, altering sodium and calcium flux,
causing channels rectification, in particular affecting AMPAR
lacking a GluR2 subunit and KAR lacking a GluR6 subunit (27). The AMPAR rectification by Spm is
dependent on the voltage and can regulate the calcium flux and the
excitability threshold of synapses (28). Polyamines can regulate another
glutamate receptor, the NMDAR, a channel gate calcium and sodium
containing two copies of NR1 and NR2 subunits. Polyamines, Spm in
particular, are able either to stimulate or inhibit the NMDAR,
according to the concentrations of glutamate, glycine and
magnesium. Spermine can increase the frequency of the channel
opening causing a strengthening of the NMDAR current in presence of
saturating concentration of glycine. Only in the absence of
magnesium extracellular Spm can block NMDAR in a voltage dependent
manner. The inhibitory effect may take place because the
extracellular Spm interacts with the negatively charged residues of
the NMDAR, creating a steric obstruction for the passage of ions
leading to a reduction of current through the channel. At
physiological magnesium concentrations, only Spm-driven NMDAR
stimulation can occur (29).
3. Polyamines and mental disorders
Polyamines and neurological
disorders
According to the World Health Organization one in
four people will be affected by mental or neurological disorders at
some point in their lives; in the USA nearly 50% of the adult
population have experienced depression or anxiety disorder, which
are the most common and debilitating forms of mental illnesses
associated with a substantially decreased of quality life (30). Currently ~450 million people
suffer from such conditions, for this reason mental illnesses are a
type of worldwide disabilities (31). In fact, this type of disability is
widely recognized as a major illness responsible for indirect costs
because of its high economic impact on society (32). According to the Bureau of Economic
Analysis's Health Care Satellite Account, in 2013 in the USA ~$89
billion has been spent on the treatment of mental illnesses
(33). In the past years, many
studies have focused on understanding the mechanisms underlying
mental illness (34-39); much of the literature has analyzed
the role of the monoaminergic system, in particular, the serotonin
and catecholamine involvement in the etiology of these pathologies
(40-42). The impairment of the monoaminergic
system alone cannot explain all the aspects related to these
diseases, since over the years it has become increasingly clear the
contribution of other players such as PAs (43). One of the early hints of the
neurobiological role of PAs was the serendipitous discovery that
antimalarial drugs, with psychosis side effects, contained a Spd
moiety in their structure (44).
As described above, PAs can affect neuronal excitability since they
interact with different transmembrane channels (27), in the light of this important role
in central nervous system (CNS), over the last three decades
extensive research has pointed out their implications in different
psychiatric conditions. In fact, an alteration of the PA content
and their metabolic enzymes have been found in different mental
illness, such as schizophrenia, mood and anxiety disorders
(45).
Polyamines and schizophrenia
Schizophrenia animal models and human patients have
shown dysregulation of the PA metabolism (46). In patients, the blood's PA content
alteration seems to be due to the pharmacological treatment
response to neuroleptics drugs since these changes have not been
observed in untreated patients and in neuroleptic-resistant
schizophrenia patients (45).
Early studies have pointed out the involvement of PAOX in
schizophrenia, where an increased level of this enzyme has been
found in schizophrenic patient serums (47-49). The contribution of ODC is still
poorly understood, since no differences regarding its levels were
found in patients while an increase of ODC activity was observed in
cortical neurons from a rat model of schizophrenia (50). Nonetheless, several works confirm
the importance of ornithine metabolism in the genesis of
schizophrenic disease because ornithine aminotransferase, antizyme
inhibitor and ornithine cyclodeaminase were found to be decreased
in the prefrontal cortex of patients (51-53). One of the hypothesises proposed to
validate the role of brain PAs in the aetiology of schizophrenia
was that they can modulate dopamine pathway; the latter shares with
the higher PA Spd and Spm, the S-adenosylmethioninamine common
precursor in the biosynthetic pathway. Furthermore, NMDAR
modulation by Spd and Spm, as described above, could explain the
hypofunctional NMDAR signalling in schizophrenia.
Polyamines and depression
Alteration in PA systems have also been found in
animal models of depression. In rats affected by depression a
hippocampal decrease of Put, Spd and Spm has been observed, while
only a decrease of Put levels was observed in the nucleus accumbens
(54). Putrescine was shown to
possess antidepressant properties, since its administration by
injection can reduce immobility time in forced swimming and tail
suspension tests. Analysis of plasma from humans suffering from
depression showed a high level of Agm that is restored to normal
levels after antidepressant treatments, highlighting the critical
role of this molecule in depression (45). Previously, the role of
neurotransmitter has been proposed for Agm in the CNS. This was
confirmed by its accumulation in synaptic vesicles and by the
ability of Agm to be secreted following depolarization (55). Moreover, it has been proved that
Agm is a selective antagonist of the NMDA polyamine-binding site
(56). All these data confirm the
involvement of the PA system in depression. In a similar way to
what was observed in schizophrenic patients, high levels of PAs
were also found in the plasma of patients suffering from depression
(49). Evidence showed that the
transcript and protein levels of different elements of the PA
system are altered in several brain regions of suicide completers;
in particular post-mortem studies have highlighted changes in the
SSAT enzyme which shows a lower level of expression compared with
healthy people (43,57). It has been proposed that in the
brains of depressed people, the lowering of the expression of SSAT
could be a compensatory mechanism to cope with the excessive
presence of PA (57).
Polyamines and stress response
Over the past decade, great attention has been
focused on the role of PAs in the context of stress response and
particularly on its causal relationship with the morbidity of
anxiety and psychiatric disorders. This pathological condition is
called PAs stress response (PSR), it can be triggered by different
types of stressors and this response can be modulated in accordance
with the stressor intensity (58). In the CNS acute stressors do not
increase the concentrations of all PAs but rather they lead to an
accumulation of Put and Agm as well as an increase of ODC activity.
Contrary to the events that have been observed in the CNS, in which
the PSR can be activated independently, in the peripheral nervous
system PSR trigger occurs only after the activation of the
hypothalamic-pituitary axis. Following acute stress, changes are
only appreciated in Put and ODC, while Spd and Spm remain unchanged
resulting an apparently incomplete PSR. When stressors become
chronic, they lead to a complete PSR and changes are also observed
in Spd and Spm levels. However, persistent chronic stressors can
activate a maladaptive PSR since they cause PA accumulation.
Repeated stress events predispose to an increased risk of
developing mental pathologies including depression, anxiety and in
suicide, which often has comorbidity with mood and personality
disorders (57).
4. Polyamines and epilepsy
Link between epilepsy and polyamines
Epilepsy is a neurological disorder characterized by
the manifestation of spontaneous and repeated seizures, which are
caused by disproportionate and simultaneous electrical activity of
neuronal networks. This disorder is one of the most common
worldwide neurological diseases; its incidence in the population is
estimated to be 1-2% (59). In
most cases, the disease is idiopathic, since the factors triggering
epilepsy are unknown; however, some cases can be related to
hereditary factors or to a brain injury. Epilepsy is classified
into different categories: childhood absence epilepsy, benign focal
epilepsy, juvenile myoclonic epilepsy and temporal lobe epilepsy
(TLE). The latter is the most common epilepsy occurring in adults
(60-62). Over the years, different research
groups have focused their studies on the role that PAs could have
in the molecular mechanisms underlying epilepsy. Since the
knowledge, derived from clinical human studies, was not sufficient
to improve understanding of the epileptic pathways, it was
necessary to use the correct genetic models. In fact, extensive
studies have been carried out using animal models in order to
characterize the different form of epilepsy and better define the
physiological importance of PAs in order to develop treatment
therapies to be applied in epilepsy (22,63-71).
Mouse models of epilepsy
The epilepsy animal models used provide seizure
induction by chemoconvulsants, traumatic brain injury and
electrical or sound stimuli. Among the most used chemoconvulsants
are included Pentylenetetrazole (PTZ) and kainic acid (KA).
Epileptic seizures induced by PTZ and KA have also been used in
transgenic mouse models characterized by a deregulation of the PA
system caused by the overexpression of PA metabolic enzymes. The
transgenic mouse line K2 (63)
overexpressing ODC in the brain and consequently having a high Put
cellular content, was shown to be neuroprotected from physically
(electroshock) and chemically (PTZ) induced seizure activity
(64), but displayed impaired
spatial learning, vision, swimming ability and lack of motivation
(64). The deficit in spatial
learning was demonstrated to be associated with a constitutively
high Put level as a result of its antagonistic effect on the NMDA
receptor. Additionally, in transgenic rats overexpressing ODC
(65) subjected to transient
focal cerebral ischaemia, significantly smaller stroke lesions were
observed in comparison with control rats, confirming that induction
of ODC and the subsequent accumulation of Put are neuroprotective
responses in the transient cerebral ischaemia (72). The transgenic mouse line
ubiquitously overexpressing SSAT and generated by Pietilä et
al (66), likewise the ODC
over-expressing line mentioned above, displayed a high level of Put
in the brain as a result of SSAT overexpression. These transgenic
mice showed neuroprotection from KA-induced neuronal toxicity
(67) and an elevated threshold
to PTZ-induced convulsions in comparison with wild-type animals
(68). Neurobehavioral profiling
of SSAT overexpressing mice showed impaired spatial learning and
the mice were revealed to be hypomotoric and less aggressive than
wild-type animals (69). Although
Put is a weak antagonist of the NMDA receptor (29), its elevated content in the brain
could cause a partial blockade of this receptor, therefore giving
protection to the transgenic animals from seizure activity,
ischaemia reperfusion damage while producing impaired spatial
learning (68,69). A mouse Cre/loxP-based genetic
model overexpressing SMOX specifically in neocortex neurons
(Dach-SMOX, former JoSMOrec) has been engineered by Cervelli et
al (70) to investigate the
role of this enzyme and its substrate Spm, which is the strongest
PA modulator of glutamatergic receptors, and certain types of
K+ channels and Na+ channels (27,73,74). Interestingly, Dach-SMOX mice
showed a phenotype with significant astroglial and microglial
activation in the neocortex of old animals, showing more pronounced
brain damage during ageing. Furthermore, in excitotoxic conditions
induced by KA injection, Dach-SMOX mice were more sensitive than
control animals (22,71). Compared with transgenic ODC and
SSAT overexpressing mice, which displayed a neuroprotective
response to different insults, Dach-SMOX animals showed the
opposite phenotype, since increased neurodegeneration was observed
during ageing and following KA injection (70,72). The production of
H2O2 and 3-AP, derived from Spm oxidation,
together with direct effects of Spm on AMPA and KA receptors, are
synergistically involved in ROS increase and in the end, to
neuronal degeneration and death. In conclusion, all these
engineered transgenic rodent lines may represent useful in
vivo genetic models for studying PA metabolism dysregulation in
brain pathological conditions due to various physically and
chemically induced excitotoxic insults that induce epilepsy
(70,75).
5. Mood disorders and epilepsy
The coexistence of two or more pathologies in the
same person is defined as comorbidity. The link between these
altered conditions can be associated with genetic and environmental
factors as well as with common pathogenic mechanisms. The
comorbidity between epilepsy and psychiatric diseases has not been
considered for a long time. Over the last few years, several
epidemiological studies have shown a strong correlation between
epilepsy and different mental illnesses. Several studies
demonstrated that among the many mental illnesses, those prevalent
in epileptic patients are mood and anxiety disorders (76-82). In particular, anxiety appears to
have a greater incidence (83).
Of the people suffering from epilepsy, ~6% of them present
psychiatric comorbidity; this percentage increases in patients with
TLE and/or refractory epilepsy reaching peaks of 60-80% (82). It is very interesting that the
amygdala, a region of the brain often involved in some types of
epilepsy, is also implicated in the neurobiology of anxiety. The
amygdala plays a key role in the manifestation of emotional and
behavioural responses and by different hypothalamic pathways can
activate neuroendocrine and autonomic responses (81). These responses take place during
stressful circumstances. One of the most reliable hypotheses is
that activation of these neuronal circuits together with an
excessive excitability of the related neurons may be responsible of
the manifestation of anxiety (79,84). Notably, the amygdala plays a key
role also in the most common form of epilepsy, TLE, which is also
the most resistant to pharmacological treatments (85). Moreover, a proof of the
relationship between anxiety and epilepsy is that the
pharmacological treatment of anxiety is often carried out using
specific anti-epileptic drugs such as benzodiazepines (86). It has been demonstrated that
psychiatric factors could be predictive of response to treatments
of seizure disorders. It has been demonstrated that epileptic
patients with psychiatric comorbidity show a reduced response to
pharmacological treatments. In fact, a study conducted by Hitiris
et al (87) on 780
patients with new-onset epilepsy showed that if they had a previous
diagnosis of psychiatric illness they were twice as likely to
develop drug resistance to anti-epileptic treatments. Furthermore,
the study by Kanner (88) proved
that mental diseases could cause worse responses to surgical
procedures for epilepsy.
6. Conclusion
For a long time, the link between epilepsy and
mental disorders has been attributed exclusively to the
serotonergic system. Several studies, in fact, have shown how an
epilepsy animal model displayed alterations in serotonergic system
both in terms of release of serotonin and of deficit in pre/post
synaptic transmission of serotonergic neurons (89-91). The connection between
noradrenaline and serotonin deficiencies and epilepsy have been
found also in humans. The noradrenaline/serotonin theory proposed
by Jobe and Browning (92)
pointed out that mood disorder and epilepsy share a common
background. In particular, the two pathologies display different
neuronal circuits, defined intrinsic fabricators that start and
maintain dysfunctional events. Nevertheless, they have the same
exterior defensive shields in which noradrenaline and serotonin
play a crucial role in the protection of the system against a
malfunction of the intrinsic fabricators. Depending on which
neuronal circuits are unbalanced, the development of epileptic or
mental illnesses can be generated. As described previously, the
monoaminergic system fails to explain all the events or all the
pathways that lead to the development of mental illnesses, just as
it is not possible to confirm that epilepsy is caused only by
changes in monoamines. In this review, the authors wanted to focus
on the role that PAs play in the CNS, proposing an evolution of the
Jobe and Browning's theory in which, in addition to the
noradreanalin/serotonin system, PAs could also be considered
exterior defensive shields important to protect the brain from the
development of epilepsy and mental illnesses (Fig. 3). In this context, the modulation
of PA metabolism may be a new important target for the development
of anti-epileptic, anxiolytic and anti-depressive drugs for the
treatment of these diseases that affect health and quality of life
and are important driver of health care costs.
Funding
No funding was received.
Availability of data and materials
Not applicable.
Authors' contributions
GB, JRS, EA, PM and MC conceived the present review,
performed the literature search and wrote the manuscript. All
authors have read and approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Acknowledgments
The authors would like to thank the ‘International
Polyamine Foundation-ONLUS' for the availability to look up the
poly-amines documentation. The Grant to Department of Science, Roma
Tre University (MIUR-Italy Dipartimenti di Eccellenza, ARTICOLO 1,
COMMI 314 - 337 LEGGE 232/2016) is gratefully acknowledged.
References
|
1
|
Thomas T and Thomas TJ: Polyamines in cell
growth and cell death: Molecular mechanisms and therapeutic
applications. Cell Mol Life Sci. 58:244–258. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Rea G, Bocedi A and Cervelli M: Question:
What is the biological function of the polyamines? IUBMB Life.
56:167–169. 2004.PubMed/NCBI
|
|
3
|
Wallace HM, Fraser AV and Hughes A: A
perspective of poly-amine metabolism. Biochem J. 376:1–14. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Li G, Regunathan S and Reis DJ: Agmatine
is synthesized by a mitochondrial arginine decarboxylase in rat
brain. Ann N Y Acad Sci. 763:325–329. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Sastre M, Regunathan S, Galea E and Reis
DJ: Agmatinase activity in rat brain: A metabolic pathway for the
degradation of agmatine. J Neurochem. 67:1761–1765. 1996.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Moretti M, Matheus FC, de Oliveira PA,
Neis VB, Ben J, Walz R, Rodrigues AL and Prediger RD: Role of
agmatine in neurode-generative diseases and epilepsy. Front Biosci
(Elite Ed). 6:341–359. 2014. View
Article : Google Scholar
|
|
7
|
Cervelli M, Angelucci E, Stano P, Leboffe
L, Federico R, Antonini G, Mariottini P and Polticelli F: The
Glu216/Ser218 pocket is a major determinant
of spermine oxidase substrate specificity. Biochem J. 461:453–459.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Cervelli M, Bellavia G, Fratini E,
Amendola R, Polticelli F, Barba M, Federico R, Signore F, Gucciardo
G, Grillo R, et al: Spermine oxidase (SMO) activity in breast tumor
tissues and biochemical analysis of the anticancer spermine
analogues BENSpm and CPENSpm. BMC Cancer. 10:5552010. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Cervelli M, Angelucci E, Germani F,
Amendola R and Mariottini P: Inflammation, carcinogenesis and
neurodegeneration studies in transgenic animal models for polyamine
research. Amino Acids. 46:521–530. 2014. View Article : Google Scholar
|
|
10
|
Casero RA Jr, Murray Stewart T and Pegg
AE: Polyamine metabolism and cancer: Treatments, challenges and
opportunities. Nat Rev Cancer. 18:681–695. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Amendola R, Cervelli M, Fratini E,
Polticelli F, Sallustio DE and Mariottini P: Spermine metabolism
and anticancer therapy. Curr Cancer Drug Targets. 9:118–130. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Polticelli F, Salvi D, Mariottini P,
Amendola R and Cervelli M: Molecular evolution of the polyamine
oxidase gene family in Metazoa. BMC Evol Biol. 12:902012.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Cervelli M, Salvi D, Polticelli F,
Amendola R and Mariottini P: Structure-function relationships in
the evolutionary framework of spermine oxidase. J Mol Evol.
76:365–370. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Tavladoraki P, Cervelli M, Antonangeli F,
Minervini G, Stano P, Federico R, Mariottini P and Polticelli F:
Probing mammalian spermine oxidase enzyme-substrate complex through
molecular modeling, site-directed mutagenesis and biochemical
characterization. Amino Acids. 40:1115–1126. 2011. View Article : Google Scholar
|
|
15
|
Cervelli M, Polticelli F, Federico R and
Mariottini P: Heterologous expression and characterization of mouse
spermine oxidase. J Biol Chem. 278:5271–5276. 2003. View Article : Google Scholar
|
|
16
|
Cervelli M, Amendola R, Polticelli F and
Mariottini P: Spermine oxidase: Ten years after. Amino Acids.
42:441–450. 2012. View Article : Google Scholar
|
|
17
|
Poulin R, Casero RA and Soulet D: Recent
advances in the molecular biology of metazoan polyamine transport.
Amino Acids. 42:711–723. 2012. View Article : Google Scholar
|
|
18
|
Abdulhussein AA and Wallace HM: Polyamines
and membrane transporters. Amino Acids. 46:655–660. 2014.
View Article : Google Scholar
|
|
19
|
Seiler N and Atanassov CL: The natural
polyamines and the immune system. Prog Drug Res. 43:87–141.
1994.PubMed/NCBI
|
|
20
|
Mastrantonio R, Cervelli M, Pietropaoli S,
Mariottini P, Colasanti M and Persichini T: HIV-Tat induces the
Nrf2/ARE pathway through NMDA receptor-elicited spermine oxidase
activation in human neuroblastoma cells. PLoS One. 11:e01498022016.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Igarashi K, Uemura T and Kashiwagi K:
Acrolein: An effective biomarker for tissue damage produced from
polyamines. Methods Mol Biol. 1694:459–468. 2018. View Article : Google Scholar
|
|
22
|
Pietropaoli S, Leonetti A, Cervetto C,
Venturini A, Mastrantonio R, Baroli G, Persichini T, Colasanti M,
Maura G, Marcoli M, et al: Glutamate excitotoxicity linked to
spermine oxidase overexpression. Mol Neurobiol. 55:7259–7270. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Leonetti A, Baroli G, Fratini E,
Pietropaoli S, Marcoli M, Mariottini P and Cervelli M: Epileptic
seizures and oxidative stress in a mouse model overexpressing
spermine oxidase. Amino Acids. Jun 13–2019.Epub ahead of print.
View Article : Google Scholar
|
|
24
|
Skatchkova SN, Antonovb SM and Eatona MJ:
Glia and glial polyamines. Role in brain function in health and
disease. Biochemistry (Moscow) Suppl Ser A Membr Cell Biol.
10:73–98. 2016. View Article : Google Scholar
|
|
25
|
Oliver D, Baukrowitz T and Fakler B:
Polyamines as gating molecules of inward-rectifier K+
channels. Eur J Biochem. 267:5824–5829. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Li J, Doyle KM and Tatlisumak T:
Polyamines in the brain: Distribution, biological interactions, and
their potential therapeutic role in brain ischaemia. Curr Med Chem.
14:1807–1813. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Williams K: Interactions of polyamines
with ion channels. Biochem J. 325:289–297. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Pegg AE: Functions of polyamines in
mammals. J Biol Chem. 291:14904–14912. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Williams K, Dawson VL, Romano C, Dichter
MA and Molinoff PB: Characterization of polyamines having agonist,
antagonist, and inverse agonist effects at the polyamine
recognition site of the NMDA receptor. Neuron. 5:199–208. 1990.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Elsayed M and Magistretti PJ: A new
outlook on mental illnesses: Glial involvement beyond the glue.
Front Cell Neurosci. 9:4682015. View Article : Google Scholar
|
|
31
|
Sayers J: The world health report
2001-Mental health: New understanding, new hope. Bull World Health
Organ. 79:10852001.
|
|
32
|
Merikangas KR, Nakamura EF and Kessler RC:
Epidemiology of mental disorders in children and adolescents.
Dialogues Clin Neurosci. 11:7–20. 2009.PubMed/NCBI
|
|
33
|
Kamal R, Cox C and Rousseau D: Kaiser
Family Foundation: Costs and outcomes of mental health and
substance use disorders in the US. JAMA. 318:4152017. View Article : Google Scholar
|
|
34
|
Mkrtchian A, Aylward J, Dayan P, Roiser JP
and Robinson OJ: Modeling avoidance in mood and anxiety disorders
using reinforcement learning. Biol Psychiatry. 82:532–539. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Benarous X, Consoli A, Cohen D, Renaud J,
Lahaye H and Guilé JM: Suicidal behaviors and irritability in
children and adolescents: A systematic review of the nature and
mechanisms of the association. Eur Child Adolesc Psychiatry.
28:667–683. 2019. View Article : Google Scholar
|
|
36
|
Ferrúa CP, Giorgi R, da Rosa LC, do Amaral
CC, Ghisleni GC, Pinheiro RT and Nedel F: MicroRNAs expressed in
depression and their associated pathways: A systematic review and a
bioin-formatics analysis. J Chem Neuroanat. 100:1016502019.
View Article : Google Scholar
|
|
37
|
Furuyashiki T and Kitaoka S: Neural
mechanisms underlying adaptive and maladaptive consequences of
stress: Roles of dopaminergic and inflammatory responses.
Psychiatry Clin Neurosci. Jun 19–2019.Epub ahead of print.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Jin Y, Sun LH, Yang W, Cui RJ and Xu SB:
The role of BDNF in the neuroimmune axis regulation of mood
disorders. Front Neurol. 10:5152019. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Peirce JM and Alviña K: The role of
inflammation and the gut microbiome in depression and anxiety. J
Neurosci Res. May 29–2019.Epub ahead of print. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Schildkraut JS: The catecholamine
hypothesis of affective disorders: A review of supporting evidence.
Am J Psychiatry. 122:509–522. 1965. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Whitaker-Azmitia PM: Serotonin and brain
development: Role in human developmental diseases. Brain Res Bull.
56:479–485. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Liu Y, Zhao J, Fan X and Guo W:
Dysfunction in serotonergic and noradrenergic systems and somatic
symptoms in psychiatric disorders. Front Psychiatry. 10:2862019.
View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Fiori LM, Wanner B, Jomphe V, Croteau J,
Vitaro F, Tremblay RE, Bureau A and Turecki G: Association of
polyaminergic loci with anxiety, mood disorders, and attempted
suicide. PLoS One. 5:e151462010. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Andrews RC: The side effects of
antimalarial drugs indicates a polyamine involvement in both
schizophrenia and depression. Med Hypotheses. 18:11–18. 1985.
View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Fiori LM and Turecki G: Implication of the
polyamine system in mental disorders. J Psychiatry Neurosci.
33:102–110. 2008.PubMed/NCBI
|
|
46
|
Das I, de Belleroche J, Essali MA,
Richardson-Andrews RC and Hirsch SR: Blood polyamine in
schizophrenia. Schizophr Res. 2:1461989. View Article : Google Scholar
|
|
47
|
Meltzer HY, Arora RC, Jackman H, Pscheidt
G and Smith MD: Platelet monoamine oxidase and plasma amine oxidase
in psychiatric patients. Schizophr Bull. 6:213–219. 1980.
View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Baron M, Asnis L, Gruen R and Levitt M:
Plasma amine oxidase and genetic vulnerability to schizophrenia.
Arch Gen Psychiatry. 40:275–279. 1983. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Dahel KA, Al-Saffar NM and Flayeh KA:
Polyamine oxidase activity in sera of depressed and schizophrenic
patients after ECT treatment. Neurochem Res. 26:415–418. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Bernstein HG, Grecksch G, Becker A, Höllt
V and Bogerts B: Cellular changes in rat brain areas associated
with neonatal hippocampal damage. Neuroreport. 10:2307–2311. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Middleton FA, Mirnics K, Pierri JN, Lewis
DA and Levitt P: Gene expression profiling reveals alterations of
specific metabolic pathways in schizophrenia. J Neurosci.
22:2718–2729. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
He Y, Yu Z, Giegling I, Xie L, Hartmann
AM, Prehn C, Adamski J, Kahn R, Li Y, Illig T, et al: Schizophrenia
shows a unique metabolomics signature in plasma. Trans Psychiatry.
2:e1492012. View Article : Google Scholar
|
|
53
|
Liu P, Jing Y, Collie ND, Dean B, Bilkey
DK and Zhang H: Altered brain arginine metabolism in schizophrenia.
Trans Psychiatry. 6:e8712016. View Article : Google Scholar
|
|
54
|
Genedani S, Saltini S, Benelli A,
Filaferro M and Bertolini A: Influence of SAMe on the modifications
of brain polyamine levels in an animal model of depression.
Neuroreport. 12:3939–3942. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Reis DJ and Regunathan S: Is agmatine a
novel neurotransmitter in brain? Trends Pharmacol Sci. 21:187–193.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Askalany AR, Yamakura T, Petrenko AB,
Kohno T, Sakimura K and Baba H: Effect of agmatine on heteromeric
N-methyl-D-aspartate receptor channels. Neurosci Res. 52:387–392.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Gross JA and Turecki G: Suicide and the
polyamine system. CNS Neurol Disord Drug Targets. 12:980–988. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Turecki G: Polyamines and suicide risk.
Mol Psychiatry. 18:1242–1243. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Naseer MI, Ullah I, Al-Qahtani MH, Karim
S, Ullah N, Ansari SA, Kim MO and Bibi F: Decreased GABABR
expression and increased neuronal cell death in developing rat
brain after PTZ-induced seizure. Neurol Sci. 34:497–503. 2013.
View Article : Google Scholar
|
|
60
|
Hauser WA and Kurland RT: The epidemiology
of epilepsy in Rochester, Minnesota, 1935 through 1967. Epilepsia.
16:1–66. 1975. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Genton P and Bureau M: Epilepsy with
myoclonic absences. CNS Drugs. 20:911–916. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Téllez-Zenteno JF and Hernández-Ronquillo
L: A review of the epidemiology of temporal lobe epilepsy. Epilepsy
Res Treat. 2012:6308532012.PubMed/NCBI
|
|
63
|
Halmekytö M, Alhonen L, Wahlfors J,
Sinervirta R, Eloranta T and Jänne J: Characterization of a
transgenic mouse line over-expressing the human ornithine
decarboxylase gene. Biochem J. 278:895–898. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Halonen T, Sivenius J, Miettinen R,
Halmekytö M, Kauppinen R, Sinervirta R, Alakuijala L, Alhonen L,
MacDonald E and Jänne J: Elevated seizure threshold and impaired
spatial learning in transgenic mice with putrescine overproduction
in the brain. Eur J Neurosci. 5:1233–1239. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Lukkarinen JA, Kauppinen RA, Gröhn OH, Oja
JM, Sinervirta R, Järvinen A, Alhonen LI and Jänne J:
Neuroprotective role of ornithine decarboxylase activation in
transient focal cerebral ischaemia: A study using ornithine
decarboxylase-overexpressing transgenic rats. Eur J Neurosci.
10:2046–2055. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Pietilä M, Alhonen L, Halmekytö M, Kanter
P, Jänne J and Porter CW: Activation of polyamine catabolism
profoundly alters tissue polyamine pools and affects hair growth
and female fertility in transgenic mice overexpressing
spermidine/spermine N1-acetyltransferase. J Biol Chem.
272:18746–18751. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Kaasinen K, Koistinaho J, Alhonen L and
Jänne J: Overexpression of spermidine/spermine N-acetyltransferase
in transgenic mice protects the animals from kainate-induced
toxicity. Eur J Neurosci. 12:540–548. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Kaasinen SK, Gröhn OH, Keinänen TA,
Alhonen L and Jänne J: Overexpression of spermidine/spermine
N1-acetyltransferase elevates the threshold to
pentylenetetrazol-induced seizure activity in transgenic mice. Exp
Neurol. 183:645–652. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Kaasinen SK, Oksman M, Alhonen L, Tanila H
and Jänne J: Spermidine/spermine N1-acetyltransferase
overexpression in mice induces hypoactivity and spatial learning
impairment. Pharmacol Biochem Behav. 78:35–45. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Cervelli M, Bellavia G, D'Amelio M,
Cavallucci V, Moreno S, Berger J, Nardacci R, Marcoli M, Maura G,
Piacentini M, et al: A new transgenic mouse model for studying the
neurotoxicity of spermine oxidase dosage in the response to
excitotoxic injury. PLoS One. 8:e648102013. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Cervetto C, Vergani L, Passalacqua M,
Ragazzoni M, Venturini A, Cecconi F, Berretta N, Mercuri N,
D'Amelio M, Maura G, et al: Astrocyte-dependent vulnerability to
excitotoxicity in spermine oxidase-overexpressing mouse.
Neuromolecular Med. 18:50–68. 2016. View Article : Google Scholar
|
|
72
|
Alhonen L, Uimari A, Pietilä M, Hyvönen
MT, Pirinen E and Keinänen TA: Transgenic animals modelling
polyamine metabolism-related diseases. Essays Biochem. 46:125–144.
2009. View Article : Google Scholar
|
|
73
|
Fleidervish IA, Libman L, Katz E and
Gutnick MJ: Endogenous polyamines regulate cortical neuronal
excitability by blocking voltage-gated Na+ channels.
Proc Natl Acad Sci USA. 105:18994–18999. 2008. View Article : Google Scholar
|
|
74
|
Traynelis SF, Wollmuth LP, McBain CJ,
Menniti FS, Vance KM, Ogden KK, Hansen KB, Yuan H, Myers SJ and
Dingledine R: Glutamate receptor ion channels: Structure,
regulation, and function. Pharmacol Rev. 62:405–496. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Jänne J, Alhonen L, Pietilä M and Keinänen
TA: Genetic approaches to the cellular functions of polyamines in
mammals. Eur J Biochem. 271:877–894. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Chapouthier G and Venault P: A
pharmacological link between epilepsy and anxiety? Trends Pharmacol
Sci. 22:491–493. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
77
|
Harden CL and Goldstein MA: Mood disorders
in patients with epilepsy: Epidemiology and management. CNS Drugs.
16:291–302. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Kanner AM: Epilepsy and mood disorders.
Epilepsia. 48:20–22. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
79
|
Stahl SM: Brainstorms: Symptoms and
circuits, part 2: Anxiety disorders. J Clin Psychiatry.
64:1408–1409. 2003. View Article : Google Scholar
|
|
80
|
Jackson MJ and Turkington D: Depression
and anxiety in epilepsy. J Neurol Neurosurg Psychiatry. 76(Suppl
1): i45–i47. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
81
|
Aroniadou-Anderjaska V, Qashu F and Braga
MF: Mechanisms regulating GABAergic inhibitory transmission in the
basolateral amygdala: Implications for epilepsy and anxiety
disorders. Amino Acids. 32:305–315. 2007. View Article : Google Scholar
|
|
82
|
Hamid H, Ettinger AB and Mula M: Anxiety
symptoms in epilepsy: Salient issues for future research. Epilepsy
Behav. 22:63–68. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
83
|
Gaitatzis A, Carroll K, Majeed A and W
Sander J: The epidemiology of the comorbidity of epilepsy in the
general population. Epilepsia. 45:1613–1622. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
84
|
Rogawski MA and Löscher W: The
neurobiology of antiepileptic drugs for the treatment of
nonepileptic conditions. Nat Med. 10:685–692. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
85
|
Pitkänen A and Sutula TP: Is epilepsy a
progressive disorder? Prospects for new therapeutic approaches in
temporal-lobe epilepsy. Lancet Neurol. 1:173–181. 2002. View Article : Google Scholar
|
|
86
|
Mula M, Pini S and Cassano GB: The role of
anticonvulsant drugs in anxiety disorders: A critical review of the
evidence. J Clin Psychopharmacol. 27:263–272. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
87
|
Hitiris N, Mohanraj R, Norrie J, Sills GJ
and Brodie MJ: Predictors of pharmacoresistant epilepsy. Epilepsy
Res. 75:192–196. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
88
|
Kanner AM: Psychiatric issues in epilepsy:
The complex relation of mood, anxiety disorders, and epilepsy.
Epilepsy Behav. 15:83–87. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
89
|
Theodore WH: Does serotonin play a role in
epilepsy? Epilepsy Curr. 3:173–177. 2003. View Article : Google Scholar
|
|
90
|
Richerson GB: Serotonin: The anti-sudden
death amine? Epilepsy Curr. 13:241–244. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
91
|
Maia GH, Soares JI, Almeida SG, Leite JM,
Baptista HX, Lukoyanova AN, Brazete CS and Lukoyanov NV: Altered
serotonin innervation in the rat epileptic brain. Brain Res Bull.
152:95–106. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
92
|
Jobe PC and Browning RA: The serotonergic
and noradrenergic effects of antidepressant drugs are
anticonvulsant, not proconvulsant. Epilepsy Behav. 7:602–619. 2005.
View Article : Google Scholar : PubMed/NCBI
|