Lactoferrin (Lf), which has a molecular weight of
77-80 kDa, is an iron-binding glycoprotein that has multiple
functions in the body; it is involved in the apoptosis of cancer
cells and can regulate various immune responses (1). Lf was first discovered in 1939 and
is a 'red protein' found in milk; it can be separated and purified
from human milk and cow's milk. The isolated protein structure of
Lf is similar to that of serum transferrin, with a 60% sequence
homology, and it reversibly binds to iron (Fe3+) ions
(2). Therefore, Lf is classified
as a member of the transferrin family, together with serum
transferrin, melanotransferrin and ovotransferrin (3). This multifunctional protein is
present in mucosal secretions, including tears, saliva, vaginal
secretions, semen, nasal secretions, bronchial secretions, bile,
gastrointestinal secretions, urine, cow's milk and human milk
(4). Lf is also present on the
mucosal surface and granules of white blood cells. Human milk and
cow's milk are the most abundant sources of Lf (5). Lf is very similar between different
species. In fact, the homology between the Lf of humans and cattle
is 77% (6).
Lf promotes the absorption of iron by the body; it
regulates cell growth, removes harmful free radicals and inhibits
the formation of toxic compounds. In the regulation of immune
responses, Lf exerts antibacterial, antiviral, anti-oxidation,
anticancer and anti-inflammatory activities (5). Accordingly, Lf is added to a number
of commercial products, including infant formula, fermented milk,
cosmetics, therapeutic drinks, toothpaste and other products used
in daily life (7).
Owing to the overexpression of a number of cell
surface receptors, Lf has a positive targeting effect; therefore,
it is considered to be an ideal nanocarrier for certain hydrophobic
therapeutic agents. In addition, as Lf can cross the blood-brain
barrier (BBB), it has proven to be a good candidate for
manufacturing nanocarriers to specifically deliver drugs for brain
tumors. Therefore, Lf appears as a promising molecule with multiple
applications in the fields of cancer treatment and nanomedicine. Lf
has numerous advantages in terms of its ability to actively
participate in the manufacture of nanocarriers. Furthermore, it is
one of the few proteins that have a net positive charge under
physiological conditions [isoelectric point (pI) 8.0-8.5]. Owing to
its high pI value, Lf is positively charged over a wide range of pH
values (15), is fairly stable in
the gastrointestinal tract and possesses a number of intestinal
receptors that facilitate the oral absorption and bioavailability
of Lf-based nanocarriers within the circulation.
Lf is a natural immune modulator that plays roles in
the innate and acquired immune systems, which regulate antibody
formation, T- and B-cell maturation, and increase the percentage of
natural killer cells in the lymphocyte population (16). The ability of Lf to regulate the
activity of the immune response may be due to its ability to bind
endotoxins [lipopolysaccharides (LPSs)] (17,18). In a previous study, Lf was found
to alleviate the cellular inflammation induced by LPS by
attenuating the nuclear factor-κB/mitogen-activated protein kinase
pathways, mitigating oxidative stress and maintaining cellular
barrier integrity. Such a finding implies that Lf plays an
important role in immune regulation (19). When gram-negative bacteria try to
invade the human host, the bacteria come into contact with various
proteins of the innate immune system. Part of the bacterial outer
membrane contains LPS. When this 'pathogen-related molecular
pattern' is recognized by Toll-like receptor 4, it triggers a
number of immune responses in various white blood cells and
platelets (20,21). The combination of Lf and
endotoxins released by bacteria can decrease the degree of
stimulation of the immune system. This process can prevent
overstimulation, which sometimes occurs in diseases such as sepsis.
The hLf1-11 peptide derived from human lactoferrin (hLf) can
inhibit myeloperoxidase, which is a major host defense enzyme found
in a variety of white blood cells, which may further decrease the
innate immune response (22).
Furthermore, hLf has been shown to stimulate the maturation of
dendritic cells and recruit various white blood cells (23). Therefore, Lf plays an activating
role in innate and adaptive immune responses.
Lactoferrin is an allosteric enhancer of the
proteolytic activity of cathepsin G, thereby affecting the function
of adaptive immune cells (24).
Lf has a positive charge that enables it to bind to the negatively
charged surface molecules of various immune system cells, and this
connection is believed to trigger signaling pathways that result in
cellular responses such as activation, differentiation and
proliferation. Lf can be transported to the nucleus in order to
bind DNA and activate various signaling pathways (25). Lactoferrin can bind to DNA, and
through its highly positively charged N-terminal region, which
remains associated with the extruded DNA in the neutrophil
extracellular traps, can still contribute to the bacterial killing
in this process. As the granules also secrete a variety of
proteolytic enzymes, Lf or other polypeptides may also be locally
released from intact Lf (26).
As well as the induction of systemic immunity, skin
immunity is promoted and allergic reactions are suppressed by Lf
(27). The immune system is
activated against skin allergens, resulting in the dose-dependent
inhibition of Langerhans cell migration and dendritic cell
accumulation in lymph nodes. The exposure of leukocyte Lf to
cytokines, pro-inflammatory cytokines, TNF-α, IL-6 and IL-1β may be
adjusted to increase and decrease. The production of these factors
depends on the type of signal that is recognized by the immune
system. At the cellular level, Lf can increase the number of
CD4+ and CD8+ cells in natural killer (NK)
cells and T cells (28), promote
the recruitment of leukocytes in the blood, induce phagocytosis and
regulate the process of bone marrow formation (29). Lf also increases the expression of
hyaluronic acid, which is required for the formation of granulation
tissue, upregulates platelet-derived growth factors and promotes
the proliferation and migration of keratinocytes; this is a
necessary condition for the re-epithelialization of wounds. Lf also
protects cells from apoptosis (30).
Lf is considered as a key component of the first
line of defense for the human body (15) and has a variety of biological
effects, including regulation of the immune response, iron
absorption, and anti-inflammatory and antioxidant activities. Lf
exerts antitumor effects through a variety of mechanisms. Oral bLf
can decrease chemically induced carcinogenesis in rodents and has
significant cytotoxicity and anti-metastatic activity in numerous
cancer cell lines, such as breast cancer and stomach cancer cell
lines (31,32). Table
I summarizes some of the anti-carcinogenic mechanisms of
Lf.
Lf is a survival factor of rheumatoid synovial
neutrophils, an iron-binding protein that is released from
activated neutrophils in inflammatory sites, and has
anti-inflammatory and antibacterial properties (33). Although the isolation of iron by
Lf and the direct effect on reactive oxygen intermediates are major
factors in decreasing excessive inflammatory response damage by
directly controlling the development of higher-order immune
functions, Lf can regulate injury and pathology caused by injury.
This ultimately leads to a decrease in the pathological damage
during inflammation (34). The
mechanism of action of Lf involves a component that differentially
regulates the cellular immune response in sepsis models in
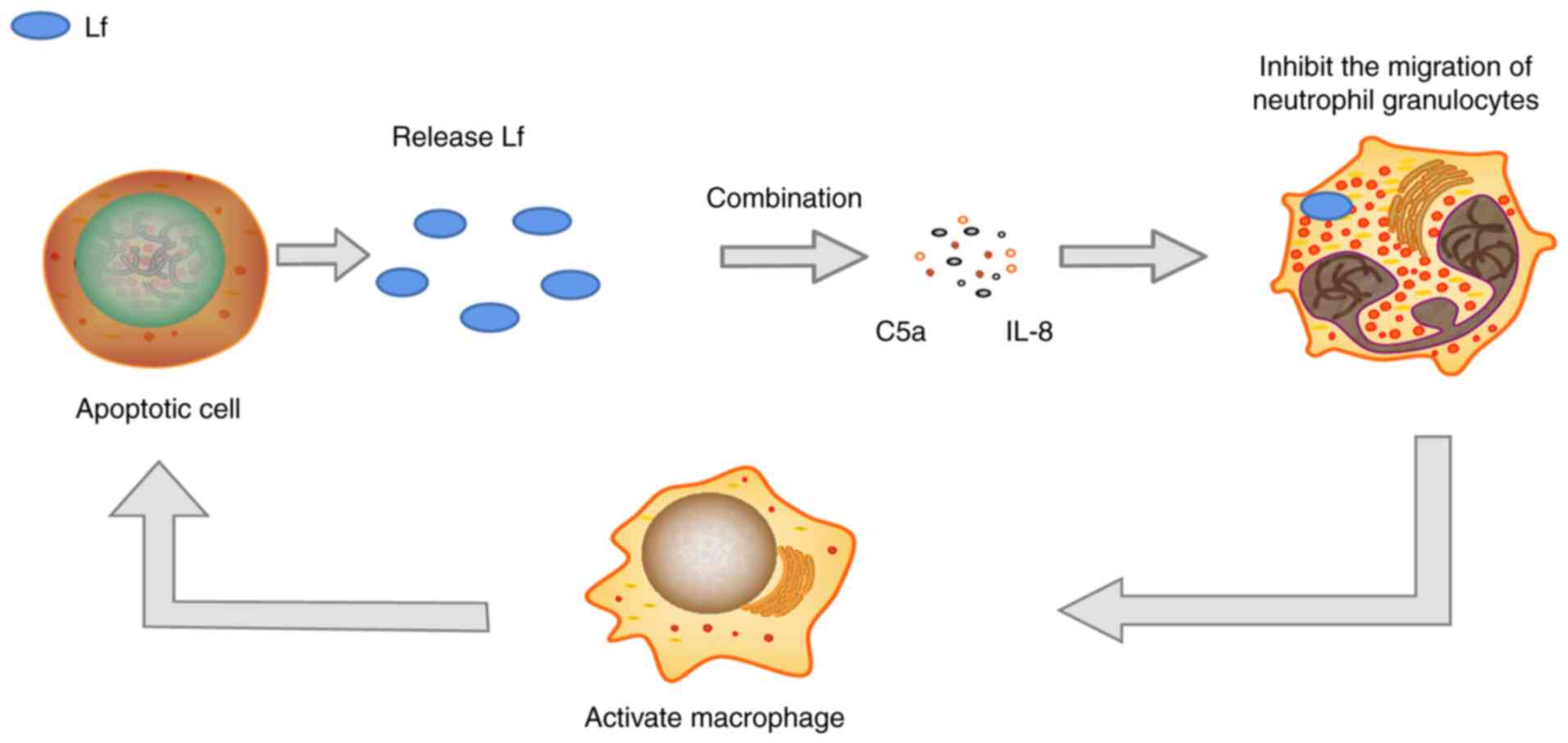
vivo. Apoptotic cells can release Lf and combine with
neutrophils to inhibit the chemotaxis of neutrophils, enabling
macrophages to swallow apoptotic cells, thereby exerting
anti-inflammatory effects (Fig.
1) (35).
Cell apoptosis induced by Lf has been described as
the pivotal pathway whereby peptides exert their cytotoxic effects
against various cancer cells. However, the apoptotic pathway that
they trigger depends on the cell type (56,57).
In a previous study, stomach cancer SGC-7901 cells
were treated with Lf, and phosphorylated Akt and numerous key
proteins involved in the Akt signaling pathway were decreased as a
result. However, the expression levels of phosphorylated caspase-9
and phosphorylated glycogen synthase kinase-3β were increased,
indicating that, in stomach cancer SGC-7901 cells, Lf-induced
apoptosis may be regulated via the Akt pathway (32). Lf was also found to induce a
stress-related mitogen-activated protein kinase pathway in Jurkat T
cells, where c-Jun N-terminal kinase (JNK) associated with Bcl-2
was hypothesized to be the pathway responsible for the apoptosis
induced by Lf (58). Lf treatment
induced caspase-9 and -3 activation and increased the level of
Bcl-2 phosphorylation. Following the abolition of JNK activation,
cell death did not occur in Lf-treated Jurkat cells. Additionally,
BLf was demonstrated to induce the apoptotic extrinsic pathway by
upregulating Fas signaling in the colon mucosa of
azoxymethane-treated rats (59).
Mammalian cell cycles are usually strictly
controlled by hormones and growth factors, and abnormal regulation
may lead to tumors. Cyclins, cyclin-dependent kinases (CdKs) and
their antagonists, CdK inhibitors, are key factors that regulate
cell cycle progression. In breast cancer MDA-MB-231 cells, hLF was
found to inhibit cell growth during the transition phase from
G1 to S of the cell cycle. At the molecular level, hLF
induces a significant decrease in the protein level and activity of
Cdk2 and Cdk4, activates cyclins D and E, and plays a key role in
the transition from the G1 to the S phase (60). Similar hLf effects have also been
reported in four head and neck cancer cell models, with blockage of
the G1 to S phase after hLf treatment (61). Lf was reported to induce cell
growth arrest by reducing phospho-Akt resulting in increased
expression and activity of p21Cip1 and p27Kip1 (62).
Lf has been confirmed to enhance the adaptive immune
response and is an effective anti-inflammatory drug (63). Although its molecular mechanism
still needs to be revealed, researchers have found that both hLf
and bLf can enter the host cell nucleus and bind to DNA to regulate
gene expression, thereby exerting their anti-inflammatory activity
(64). hLf has significantly
increased NK cell-mediated cytotoxicity in breast cancer and colon
cancer cell lines (65).
It is well know that tumor cells overexpress Lf
receptors in order to fulfill the increased nutritional demands of
these highly proliferative cells (67). Lf is an ideal nanocarrier for
certain hydrophobic therapeutics due to its active targeting
potential as a result of its receptor being overexpressed on the
surface of a number of cells. Moreover, Lf is good potential
candidate for fabricating nanocarriers that specifically deliver
drugs to brain tumors, as Lf can cross the BBB. Consequently, Lf
appears as a promising molecule with multiple applications in the
fields of cancer therapy and nanomedicine (68). Song et al (69) demonstrated the potential utility
of Lf-conjugated GO@Fe3O4 nanocomposites for therapeutic
applications in the treatment of gliomas (69). Lf-conjugated iron oxide
nanoparticles can be used as tracers for targeted brain glioma
imaging using magnetic particle imaging (70). Lf/phenylboronic
acid-functionalized hyaluronic acid (HA) nanogel crosslinked with a
disulfide bond crosslinker was generated as a reduction-sensitive
dual-targeting glioma therapeutic platform for doxorubicin
hydrochloride (DOX) delivery (71). Lf-HA-DOX significantly increased
drug delivery to the glioma and may thus serve as a promising
anti-glioma therapy (72).
Inflammatory bowel disease is a chronic inflammatory
and relapsing condition of the gastrointestinal tract (73). Normally, the gut microbiota is
composed of 90% Bacteroidetes and Firmicutes, with rare phyla, such
as Proteobacteria and Actinobacteria, as well as fungi, viruses,
and protists, composing the remaining 10%. Anti-microbial activity
has been described as the first Lf function linked to the ancestral
host defense-linked mechanisms to target pathogen infections. This
activity, evaluated in several in vitro (74,75) and in vivo (76) models, can be both independent and
dependent of the iron-binding ability of Lf. The anticancer
activity of Lf via host immunomodulation has been widely reported,
particularly in colorectal cancer (43).
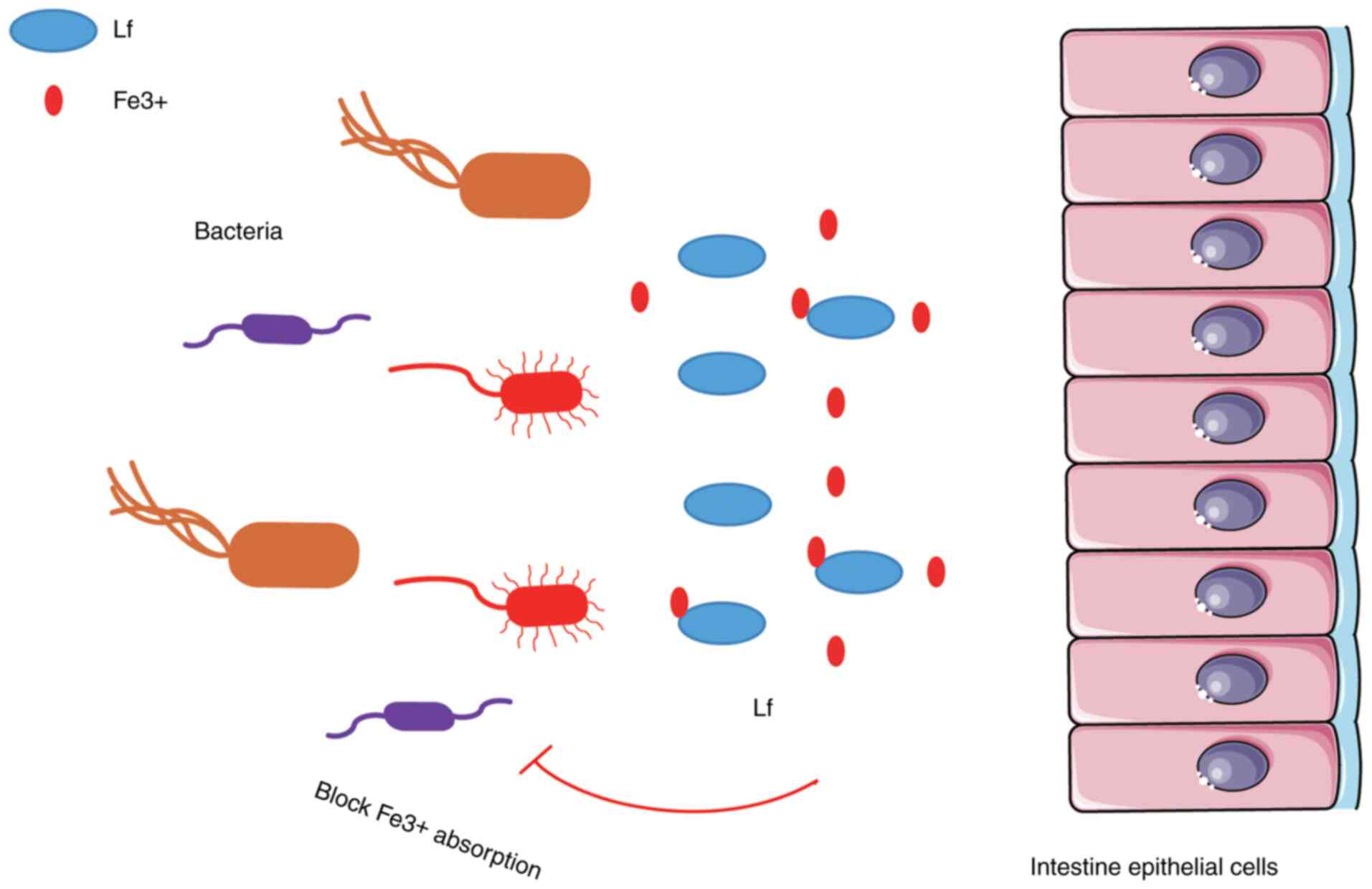
Lf is a multifunctional natural defense protein with
significant antibacterial activity (77); its function is mainly reflected in
the absorption of Fe3+, which limits the use of
Fe3+ by bacteria in the infected site, and inhibits the
growth and reproduction of these microorganisms and the expression
of their virulence factors. The bactericidal effect of Lf is mainly
mediated by its interaction with the bacterial surface. In
vivo (78) and in
vitro (79) studies have
shown that Lf prevents certain bacteria from adhering to host cells
(15) (Fig. 3).
Lf destroys the outer membrane of gram-negative
bacteria by interacting with LPS. The positively charged n-terminus
of Lf prevents LPS and bacterial cations (Ca2+ and
Mg2+) from interacting, resulting in the release of LPS
from the cell wall, increasing membrane permeability and
subsequently causing damage to the bacteria. The interaction
between Lf and LPS also enhances the effect of natural
antibacterial agents, such as lysozyme, which are secreted from the
mucosa at high concentrations together with Lf via the BBB
(15). Dialysis chamber research
indicates that bacterial killing requires direct contact with Lf,
and work with purified LPS suggests that this relates to direct
LPS-binding by the protein. As Lf and lysozyme are both present in
mucosal secretions and neutrophil granules, their interaction may
help the host defense (80).
Lf is an iron-binding glycoprotein found in some
mucosal secretions and with antiviral activity against DNA and RNA
viruses, such as HIV and rotavirus. The antiviral effect of LF
occurs in the early stages of infection. Lf prevents the virus from
entering the host cell by blocking cell receptors (84). In particular, Lf has antiviral and
immune responses, such as demonstrated by Lf against SARS-CoV,
which is closely related to SARS-CoV-2 that causes COVID-19
(85).
Lf can prevent the internalization of certain
viruses into host cells, such as poliovirus type 1, which causes
human polio, herpes simplex virus types I and II, and giant cell
viruses. For other viruses, such as hepatitis C virus (HCV) and
rotavirus, instead of preventing their entry, Lf inhibits virus
replication in the host cell. At present, the specific inhibitory
mechanism remains to be studied; however, a widely accepted
hypothesis is the binding and blockage of glycosaminoglycan virus
receptors, particularly heparan sulfate (HS), by Lf. LF and HS in
combination stops the first contact between the host and virus
cells, thereby preventing infection. In vitro studies have
shown that in human plasma and milk proteins, LF exerts a strong
activity on HIV, and this effect is due to the inhibition of virus
replication in the host cells (15). The HCV envelope protein binds to
Lf. hLf and bLf, a multifunctional immunomodulator, combines two
HCV envelope proteins. Based on western blotting with milk
separated by sodium lauryl sulfate-polyacrylamide gel
electrophoresis or immunopurification, bacteria expressing e1 and
e2 can bind to Lf (86-88).
The effect of Lf on gram-positive bacteria is mainly
based on the combination of cations on the surface of Lf and anions
on the surface of the bacteria, thereby neutralizing the negative
charges on the surface of the gram-positive bacteria. For example,
lipoteichoic acid decreases the negative charge on the cell wall,
thereby facilitating the contact between lysozyme and the
peptidoglycan under the cell wall, ultimately exerting an enzymatic
effect (15).
Lf functions as an enzyme in certain reactions. Lf
is the milk protein with the highest activity of DNase, RNase,
ATPase and amylase. However, these activities are not the only
enzymatic activities of Lf. bLf binds two HCV envelope proteins
(88). Lf has DNA-binding
properties (89) and can
participate in the transcriptional activation of specific DNA
sequences (90) or as a signal
transduction mediator (91).
Celiac disease has the highest amylase and ATP activities (92). The discovery of the properties of
the Lf enzyme helps to clarify a number of its physiological
functions.
Lf is used as a nanocarrier of DOX, as its receptor
is highly expressed on the surface of highly proliferating cells
(such as cancer cells). DOX is an effective cytotoxic anticancer
drug, but has been reported to exhibit extensive toxicity to the
heart and spleen, in addition to its limited oral absorption
(93,94). Drug-loaded preparations have shown
good physical stability, indicating that damage to the red blood
cell membrane is negligible. Drug delivery through
nano-formulations not only minimizes the cardiotoxicity of DOX, but
also improves the efficacy and bioavailability of the drug in a
targeted-specific manner (95).
A study has also shown that Lf nanoparticles are
used to encapsulate antiviral drugs. Zidovudine nano-encapsulation
into Lf nanoparticles has been achieved through sol-oil chemistry.
Zidovudine is an effective antiviral drug with good bioavailability
(50-75%); however, it can cause bone marrow suppression,
neutropenia and organ toxicity. In the study by Kumar et al
(96), the size of the prepared
nanoparticles was 50-60 nm, the drug encapsulation efficiency was
67% and good physical stability was observed at room temperature
and at 4°C. Furthermore, there was no significant change in
particle size or drug content. Oral administration of
efavirenz-loaded Lf nanoparticles resulted in anti-HIV-1 effects
comparable to those of the free drug. In addition, compared with
free efavirenz, drug-loaded nanoparticles showed improved
pharmacokinetic characteristics and lower organ toxicity,
indicating that this nanoformulation is a safe nanoplatform that
can enhance drug delivery (96).
The cationic nature of Lf can be used to form
complexes with negatively charged DNA through electrostatic
complexation. In this context, in a previous study, plasmid pGFPC1,
which encodes the green fluorescent protein, was used as a cargo
gene (97). Lf nanoparticles
loaded with plasmids were prepared using the sol-oil method. The
diameter of the prepared Lf nanoparticles was 60 nm and the PDI was
low, indicating the uniformity of the preparation. The prepared Lf
nanoparticles also showed enhanced physical stability at a
temperature of 4°C for ≤10 weeks without the particle size
exhibiting significant changes. Incubation in DMEM containing 10%
serum at 37°C for 8 h also did not result in changes to the
particle size, which would result in longer plasma levels. This
improved stability may be related to the strong electrostatic
interactions between the positively charged Lf and negatively
charged DNA. According to reports, Lf has a DNA-binding domain,
which may help to further promote DNA binding and the formation of
a tighter DNA-Lf nanocomplex (98).
The methods used to prepare Lf nanoparticles include
nanoparticle albumin-bound (NAB) and thermal denaturation methods.
The NAB technology is primarily dependent on the presence of an
oily phase, which is slowly added to the aqueous phase containing
Lf. As a nanoparticle, Lf forms a gel when exposed to heat
treatment, ionic strength or changes in pH. Generally, the thermal
gelation of a protein starts with a heating step to denature the
protein, followed by the addition of salt to induce protein
aggregation (87).
Lf can participate in the production of
polyelectrolyte complex nanocarriers. The synthesis of such
nanocarriers is based on the use of two oppositely charged
molecules, such as a positively charged protein and a negatively
charged natural polysaccharide. Nanocarriers based on
polyelectrolyte complexes are likely stabilized by strong
electrostatic interactions between cationic proteins and anionic
polysaccharides. This stabilization can also enhance the stability
of the encapsulated active ingredients (99). Table II summarizes representative
examples of Lf-based nanocarriers for drug delivery
applications.
This review provides an in-depth summary of the
biological characteristics of Lf, including the structure of Lf
biomolecules, its binding affinity to iron, and the interaction
between Lf and the host. Lf plays an important role in the
regulation of the immune response, and has immune stimulation,
immune activation, anti-inflammatory activity, antibacterial,
anti-viral effects and, in particular, anticancer activity. Based
on the results of several studies, it is known that Lf-based
nanocarriers can be easily prepared by simple methods and have
excellent active targeting potential for tumor tissues,
particularly brain tumors. The manufacture of Lf-based nanocarriers
can broadly enhance the therapeutic potential of encapsulating
active molecules. The present review discussed the latest
preparation methods for Lf-based nanocarriers prepared by the
sol-oil method, NAB technology and thermal denaturation method.
Not applicable.
HW, YG, XZ, GH, LL and SP collected the related
papers and completed the manuscript and figures. YG and XZ provided
constructive guidance and performed critical revisions. XY, SW, TZ,
HW and SP participated in the design of this review. All authors
read and approved the final manuscript. Data authentication is not
applicable.
Not applicable.
Not applicable.
The authors declare that they have no competing
interests.
Not applicable.
|
1
|
Rascon-Cruz Q, Espinoza-Sanchez EA,
Siqueiros-Cendon TS, Nakamura-Bencomo SI, Arévalo-Gallegos S and
Iglesias-Figueroa BF: Lactoferrin: A glycoprotein involved in
immunomodulation, anticancer, and antimicrobial processes.
Molecules. 26:2052021. View Article : Google Scholar
|
|
2
|
Baker EN, Baker HM and Kidd RD:
Lactoferrin and transferrin: Functional variations on a common
structural framework. Biochem Cell Biol. 80:27–34. 2002. View Article : Google Scholar
|
|
3
|
Lambert LA, Perri H and Meehan TJ:
Evolution of duplications in the transferrin family of proteins.
Comp Biochem Physiol B Biochem Mol Biol. 140:11–25. 2005.
View Article : Google Scholar
|
|
4
|
Pierce A, Legrand D and Mazurier J:
Lactoferrin: A multifunctional protein. Med Sci (Paris).
25:361–369. 2009.In French. View Article : Google Scholar
|
|
5
|
Hao L, Shan Q, Wei J, Ma F and Sun P:
Lactoferrin: Major physiological functions and applications. Curr
Protein Pept Sci. 20:139–144. 2019. View Article : Google Scholar
|
|
6
|
Garcia-Montoya IA, Cendon TS,
Arevalo-Gallegos S and Rascon-Cruz Q: Lactoferrin a multiple
bioactive protein: An overview. Biochim Biophys Acta. 1820:226–236.
2012. View Article : Google Scholar
|
|
7
|
Legrand D, Pierce A, Elass E, Carpentier
M, Mariller C and Mazurier J: Lactoferrin structure and functions.
Adv Exp Med Biol. 606:163–194. 2008. View Article : Google Scholar
|
|
8
|
Yanaihara A, Toma Y, Saito H and Yanaihara
T: Cell proliferation effect of lactoferrin in human endometrial
stroma cells. Mol Hum Reprod. 6:469–473. 2000. View Article : Google Scholar
|
|
9
|
Huang N, Bethell D, Card C, Cornish J,
Marchbank T, Wyatt D, Mabery K and Playford R: Bioactive
recombinant human lactoferrin, derived from rice, stimulates
mammalian cell growth. In Vitro Cell Dev Biol Anim. 44:464–471.
2008. View Article : Google Scholar
|
|
10
|
Siebert PD and Huang BC: Identification of
an alternative form of human lactoferrin mRNA that is expressed
differentially in normal tissues and tumor-derived cell lines. Proc
Natl Acad Sci USA. 94:2198–2203. 1997. View Article : Google Scholar
|
|
11
|
Klein G, Imreh S and Zabarovsky ER: Why do
we not all die of cancer at an early age? Adv Cancer Res. 98:1–16.
2007. View Article : Google Scholar
|
|
12
|
Mariller C, Hardiville S, Hoedt E, Huvent
I, Pina-Canseco S and Pierce A: Delta-lactoferrin, an intracellular
lactoferrin isoform that acts as a transcription factor. Biochem
Cell Biol. 90:307–319. 2012. View
Article : Google Scholar
|
|
13
|
Tsuda H, Ohshima Y, Nomoto H, Fujita K,
Matsuda E, Iigo M, Takasuka N and Moore MA: Cancer prevention by
natural compounds. Drug Metab Pharmacokinet. 19:245–263. 2004.
View Article : Google Scholar
|
|
14
|
Guedes JP, Pereira CS, Rodrigues LR and
Corte-Real M: Bovine milk lactoferrin selectively kills highly
metastatic prostate cancer PC-3 and osteosarcoma MG-63 cells in
vitro. Front Oncol. 8:2002018. View Article : Google Scholar
|
|
15
|
Gonzalez-Chavez SA, Arevalo-Gallegos S and
Rascon-Cruz Q: Lactoferrin: Structure, function and applications.
Int J Antimicrob Agents. 33:301.e1–e8. 2009. View Article : Google Scholar
|
|
16
|
Actor JK, Hwang SA and Kruzel ML:
Lactoferrin as a natural immune modulator. Curr Pharm Des.
15:1956–1973. 2009. View Article : Google Scholar
|
|
17
|
Appelmelk BJ, An YQ, Geerts M, Thijs BG,
de Boer HA, MacLaren DM, de Graaff J and Nuijens JH: Lactoferrin is
a lipid A-binding protein. Infect Immun. 62:2628–2632. 1994.
View Article : Google Scholar
|
|
18
|
Elass-Rochard E, Roseanu A, Legrand D,
Trif M, Salmon V, Motas C, Montreuil J and Spik G:
Lactoferrin-lipopolysaccharide interaction: Involvement of the
28-34 loop region of human lactoferrin in the high-affinity binding
to Escherichia coli 055B5 lipopolysaccharide. Biochem J.
312:839–845. 1995. View Article : Google Scholar
|
|
19
|
Hu P, Zhao F, Wang J and Zhu W:
Lactoferrin attenuates lipopolysaccharide-stimulated inflammatory
responses and barrier impairment through the modulation of
NF-kappaB/MAPK/Nrf2 pathways in IPEC-J2 cells. Food Funct.
11:8516–8526. 2020. View Article : Google Scholar
|
|
20
|
Baker HM and Baker EN: A structural
perspective on lactoferrin function. Biochem Cell Biol. 90:320–328.
2012. View Article : Google Scholar
|
|
21
|
Cools-Lartigue J, Spicer J, McDonald B,
Gowing S, Chow S, Giannias B, Bourdeau F, Kubes P and Ferri L:
Neutrophil extracellular traps sequester circulating tumor cells
and promote metastasis. J Clin Invest. 123:3446–3458. 2013.
View Article : Google Scholar
|
|
22
|
van der Does AM, Hensbergen PJ, Bogaards
SJ, Cansoy M, Deelder AM, van Leeuwen HC, Drijfhout JW, van Dissel
JT and Nibbering PH: The human lactoferrin-derived peptide hLF1-11
exerts immunomodulatory effects by specific inhibition of
myeloperoxidase activity. J Immunol. 188:5012–5019. 2012.
View Article : Google Scholar
|
|
23
|
Park HW, Park SH, Jo HJ, Kim TG, Lee JH,
Kang SG, Jang YS and Kim PH: Lactoferrin induces tolerogenic bone
marrow-derived dendritic cells. Immune Netw. 20:e382020. View Article : Google Scholar
|
|
24
|
Eipper S, Steiner R, Lesner A, Sienczyk M,
Palesch D, Halatsch ME, Zaczynska E, Heim C, Hartmann MD, Zimecki
M, et al: Lactoferrin is an allosteric enhancer of the proteolytic
activity of cathepsin G. PLoS One. 11:e1515092016. View Article : Google Scholar
|
|
25
|
Penco S, Scarfi S, Giovine M, Damonte G,
Millo E, Villaggio B, Passalacqua M, Pozzolini M, Garrè C and
Benatti U: Identification of an import signal for, and the nuclear
localization of, human lactoferrin. Biotechnol Appl Biochem.
34:151–159. 2001. View Article : Google Scholar
|
|
26
|
Vogel HJ: Lactoferrin, a bird's eye view.
Biochem Cell Biol. 90:233–244. 2012. View Article : Google Scholar
|
|
27
|
Takayama Y and Aoki R: Roles of
lactoferrin on skin wound healing. Biochem Cell Biol. 90:497–503.
2012. View Article : Google Scholar
|
|
28
|
Kabanov AV and Batrakova EV: New
technologies for drug delivery across the blood brain barrier. Curr
Pharm Des. 10:1355–1363. 2004. View Article : Google Scholar
|
|
29
|
Elzoghby AO, Abdelmoneem MA, Hassanin IA,
Abd Elwakil MM, Elnaggar MA, Mokhtar S, Fang JY and Elkhodairy KA:
Lactoferrin, a multi-functional glycoprotein: Active therapeutic,
drug nanocarrier & targeting ligand. Biomaterials.
263:1203552020. View Article : Google Scholar
|
|
30
|
Thangavel P, Ramachandran B, Chakraborty
S, Kannan R, Lonchin S and Muthuvijayan V: Accelerated healing of
diabetic wounds treated with L-Glutamic acid loaded hydrogels
through enhanced collagen deposition and angiogenesis: An in vivo
study. Sci Rep. 7:107012017. View Article : Google Scholar
|
|
31
|
Iigo M, Alexander DB, Long N, Xu J,
Fukamachi K, Futakuchi M, Takase M and Tsuda H: Anticarcinogenesis
pathways activated by bovine lactoferrin in the murine small
intestine. Biochimie. 91:86–101. 2009. View Article : Google Scholar
|
|
32
|
Xu XX, Jiang HR, Li HB, Zhang TN, Zhou Q
and Liu N: Apoptosis of stomach cancer cell SGC-7901 and regulation
of Akt signaling way induced by bovine lactoferrin. J Dairy Sci.
93:2344–2350. 2010. View Article : Google Scholar
|
|
33
|
Wong SH, Francis N, Chahal H, Raza K,
Salmon M, Scheel-Toellner D and Lord JM: Lactoferrin is a survival
factor for neutrophils in rheumatoid synovial fluid. Rheumatology
(Oxford). 48:39–44. 2009. View Article : Google Scholar
|
|
34
|
Kruzel ML, Zimecki M and Actor JK:
Lactoferrin in a context of inflammation-induced pathology. Front
Immunol. 8:14382017. View Article : Google Scholar
|
|
35
|
Bournazou I, Pound JD, Duffin R, Bournazos
S, Melville LA, Brown SB, Rossi AG and Gregory CD: Apoptotic human
cells inhibit migration of granulocytes via release of lactoferrin.
J Clin Invest. 119:20–32. 2009.
|
|
36
|
Bezault J, Bhimani R, Wiprovnick J and
Furmanski P: Human lactoferrin inhibits growth of solid tumors and
development of experimental metastases in mice. Cancer Res.
54:2310–2312. 1994.PubMed/NCBI
|
|
37
|
Damiens E, Mazurier J, el Yazidi I, Masson
M, Duthille I, Spik G and Boilly-Marer Y: Effects of human
lactoferrin on NK cell cytotoxicity against haematopoietic and
epithelial tumour cells. Biochim Biophys Acta. 1402:277–287. 1998.
View Article : Google Scholar
|
|
38
|
Shi H and Li W: Inhibitory effects of
human lactoferrin on U14 cervical carcinoma through upregulation of
the immune response. Oncol Lett. 7:820–826. 2014. View Article : Google Scholar
|
|
39
|
Iyer S, Yip TT, Hutchens TW and Lonnerdal
B: Lactoferrin-receptor interaction. Effect of surface exposed
histidine residues. Adv Exp Med Biol. 357:245–252. 1994. View Article : Google Scholar
|
|
40
|
Rastogi N, Singh A, Pandey SN, Sinha M,
Bhushan A, Kaur P, Sharma S and Singh TP: Structure of the
iron-free true C-terminal half of bovine lactoferrin produced by
tryptic digestion and its functional significance in the gut. FEBS
J. 281:2871–2882. 2014. View Article : Google Scholar
|
|
41
|
Teng CT: Factors regulating lactoferrin
gene expression. Biochem Cell Biol. 84:263–267. 2006. View Article : Google Scholar
|
|
42
|
Ieni A, Barresi V, Licata L, Cardia R,
Fazzari C, Nuciforo G, Caruso F, Caruso M, Adamo V and Tuccari G:
Immunoexpression of lactoferrin in triple-negative breast cancer
patients: A proposal to select a less aggressive subgroup. Oncol
Lett. 13:3205–3209. 2017. View Article : Google Scholar
|
|
43
|
Tsuda H, Kozu T, Iinuma G, Ohashi Y, Saito
Y, Saito D, Akasu T, Alexander DB, Futakuchi M, Fukamachi K, et al:
Cancer prevention by bovine lactoferrin: From animal studies to
human trial. Biometals. 23:399–409. 2010. View Article : Google Scholar
|
|
44
|
Gibbons JA, Kanwar RK and Kanwar JR:
Lactoferrin and cancer in different cancer models. Front Biosci
(Schol Ed). 3:1080–1088. 2011. View
Article : Google Scholar
|
|
45
|
Digumarti R, Wang Y, Raman G, Doval DC,
Advani SH, Julka PK, Parikh PM, Patil S, Nag S, Madhavan J, et al:
A randomized, double-blind, placebo-controlled, phase II study of
oral talactoferrin in combination with carboplatin and paclitaxel
in previously untreated locally advanced or metastatic non-small
cell lung cancer. J Thorac Oncol. 6:1098–1103. 2011. View Article : Google Scholar
|
|
46
|
Hoedt E, Hardiville S, Mariller C, Elass
E, Perraudin JP and Pierce A: Discrimination and evaluation of
lactoferrin and delta-lactoferrin gene expression levels in cancer
cells and under inflammatory stimuli using TaqMan real-time PCR.
Biometals. 23:441–452. 2010. View Article : Google Scholar
|
|
47
|
Zadvornyi TV, Lukianova NY, Borikun TV and
Chekhun VF: Effects of exogenous lactoferrin on phenotypic profile
and invasiveness of human prostate cancer cells (DU145 and LNCaP)
in vitro. Exp Oncol. 40:184–189. 2018. View Article : Google Scholar
|
|
48
|
Cutone A, Ianiro G, Lepanto MS, Rosa L,
Valenti P, Bonaccorsi di Patti MC and Musci G: Lactoferrin in the
prevention and treatment of intestinal inflammatory pathologies
associated with colorectal cancer development. Cancers (Basel).
12:38062020. View Article : Google Scholar
|
|
49
|
Duchardt F, Ruttekolk IR, Verdurmen WP,
Lortat-Jacob H, Bürck J, Hufnagel H, Fischer R, van den Heuvel M,
Löwik DW, Vuister GW, et al: A cell-penetrating peptide derived
from human lactoferrin with conformation-dependent uptake
efficiency. J biol Chem. 284:36099–36108. 2009. View Article : Google Scholar
|
|
50
|
Riedl S, Leber R, Rinner B, Schaider H,
Lohner K and Zweytick D: Human lactoferricin derived di-peptides
deploying loop structures induce apoptosis specifically in cancer
cells through targeting membranous phosphatidylserine. Biochim
Biophys Acta. 1848:2918–2931. 2015. View Article : Google Scholar
|
|
51
|
Kühnle A, Veelken R, Galuska CE,
Saftenberger M, Verleih M, Schuppe HC, Rudloff S, Kunz C and
Galuska SP: Polysialic acid interacts with lactoferrin and supports
its activity to inhibit the release of neutrophil extracellular
traps. Carbohydr Polym. 208:32–41. 2019. View Article : Google Scholar
|
|
52
|
El YI, Legrand D, Nuijens J, Slomianny MC,
van Berkel P and Spik G: The binding of lactoferrin to
glycosaminoglycans on enterocyte-like HT29-18-C1 cells is mediated
through basic residues located in the N-terminus. Biochim Biophys
Acta. 1568:197–204. 2001. View Article : Google Scholar
|
|
53
|
Jiang R and Lönnerdal B: Bovine
lactoferrin and lactoferricin exert antitumor activities on human
colorectal cancer cells (HT-29) by activating various signaling
pathways. Biochem Cell Biol. 95:99–109. 2017. View Article : Google Scholar
|
|
54
|
Sharma A, Shandilya UK, Sodhi M, Mohanty
AK, Jain P and Mukesh M: Evaluation of milk colostrum derived
lactoferrin of sahiwal (Bos indicus) and karan fries (Cross-Bred)
cows for its anti-cancerous potential. Int J Mol Sci. 20:63182019.
View Article : Google Scholar
|
|
55
|
Arias M, Hilchie AL, Haney EF, Bolscher
JG, Hyndman ME, Hancock RE and Vogel HJ: Anticancer activities of
bovine and human lactoferricin-derived peptides. Biochem Cell Biol.
95:91–98. 2017. View Article : Google Scholar
|
|
56
|
Zhou Y, Zeng Z, Zhang W, Xiong W, Wu M,
Tan Y, Yi W, Xiao L, Li X, Huang C, et al: Lactotransferrin: A
candidate tumor suppressor-Deficient expression in human
nasopharyngeal carcinoma and inhibition of NPC cell proliferation
by modulating the mitogen-activated protein kinase pathway. INT J
Cancer. 123:2065–2072. 2008. View Article : Google Scholar
|
|
57
|
Onishi J, Roy MK, Juneja LR, Watanabe Y
and Tamai Y: A lactoferrin-derived peptide with cationic residues
concentrated in a region of its helical structure induces necrotic
cell death in a leukemic cell line (HL-60). J Pept Sci.
14:1032–1038. 2008. View Article : Google Scholar
|
|
58
|
Lee SH, Park SW, Pyo CW, Yoo NK, Kim J and
Choi SY: Requirement of the JNK-associated Bcl-2 pathway for human
lactoferrin-induced apoptosis in the Jurkat leukemia T cell line.
Biochimie. 91:102–108. 2009. View Article : Google Scholar
|
|
59
|
Fujita K, Matsuda E, Sekine K, Iigo M and
Tsuda H: Lactoferrin enhances Fas expression and apoptosis in the
colon mucosa of Azoxymethane-treated rats. Carcinogenesis.
25:1961–1966. 2004. View Article : Google Scholar
|
|
60
|
Gopal SH and Das SK: Role of lactoferrin
in the carcinogenesis of triple-negative breast cancer. J Cancer
Clin Trials. 1:e1052016.
|
|
61
|
Velliyagounder K, Bahdila D, Pawar S and
Fine DH: Role of lactoferrin and lactoferrin-derived peptides in
oral and maxillofacial diseases. Oral Dis. 25:652–669. 2019.
View Article : Google Scholar
|
|
62
|
Damiens E, El YI, Mazurier J, Duthille I,
Spik G and Boilly-Marer Y: Lactoferrin inhibits G1 cyclin-dependent
kinases during growth arrest of human breast carcinoma cells. J
Cell Biochem. 74:486–498. 1999. View Article : Google Scholar
|
|
63
|
Ibrahim HM, Mohamed AH, Salem ML, Osman GY
and Morsi DS: Anti-neoplastic and immunomodulatory potency of
co-treatment based on bovine lactoferrin and/or muramyl dipeptide
in tumor-bearing mice. Toxicol Res (Camb). 9:137–147. 2020.
View Article : Google Scholar
|
|
64
|
Sangermano R, Pernarella S, Straker M,
Lepanto MS, Rosa L, Cutone A, Valenti P and Ottolenghi L: The
treatment of black stain associated with of iron metabolism
disorders with lactoferrin: A litterature search and two case
studies. Clin Ter. 170:e373–e381. 2019.PubMed/NCBI
|
|
65
|
Li HY, Li M, Luo CC, Wang JQ and Zheng N:
Lactoferrin exerts antitumor effects by inhibiting angiogenesis in
a HT29 human colon tumor model. J Agric Food Chem. 65:10464–10472.
2017. View Article : Google Scholar
|
|
66
|
Tammam SN, Azzazy H and Lamprecht A:
Nuclear and cytoplasmic delivery of lactoferrin in glioma using
chitosan nanoparticles: Cellular location dependent-action of
lactoferrin. Eur J Pharm Biopharm. 129:74–79. 2018. View Article : Google Scholar
|
|
67
|
Golla K, Bhaskar C, Ahmed F and Kondapi
AK: A target-specific oral formulation of Doxorubicin-protein
nanoparticles: Efficacy and safety in hepatocellular cancer. J
Cancer. 4:644–652. 2013. View Article : Google Scholar
|
|
68
|
Sabra S and Agwa MM: Lactoferrin, a unique
molecule with diverse therapeutical and nanotechnological
applications. Int J Biol Macromol. 164:1046–1060. 2020. View Article : Google Scholar
|
|
69
|
Song MM, Xu HL, Liang JX, Xiang HH, Liu R
and Shen YX: Lactoferrin modified graphene oxide iron oxide
nanocomposite for glioma-targeted drug delivery. Mater Sci Eng C
Mater Biol Appl. 77:904–911. 2017. View Article : Google Scholar
|
|
70
|
Tomitaka A, Arami H, Gandhi S and Krishnan
KM: Lactoferrin conjugated iron oxide nanoparticles for targeting
brain glioma cells in magnetic particle imaging. Nanoscale.
7:16890–16898. 2015. View Article : Google Scholar
|
|
71
|
Zhang M, Asghar S, Tian C, Hu Z, Ping Q,
Chen Z, Shao F and Xiao Y: Lactoferrin/phenylboronic
acid-functionalized hyaluronic acid nanogels loading doxorubicin
hydrochloride for targeting glioma. Carbohydr Polym.
253:1171942021. View Article : Google Scholar
|
|
72
|
Yin Y, Fu C, Li M, Li X, Wang M, He L,
Zhang LM and Peng Y: A pH-sensitive hyaluronic acid prodrug
modified with lactoferrin for glioma dual-targeted treatment. Mater
Sci Eng C Mater Biol Appl. 67:159–169. 2016. View Article : Google Scholar
|
|
73
|
Guan Q: A comprehensive review and update
on the pathogenesis of inflammatory bowel disease. J Immunol Res.
2019:72472382019. View Article : Google Scholar
|
|
74
|
Frioni A, Conte MP, Cutone A, Longhi C,
Musci G, di Patti MC, Natalizi T, Marazzato M, Lepanto MS, Puddu P,
et al: Lactoferrin differently modulates the inflammatory response
in epithelial models mimicking human inflammatory and infectious
diseases. Biometals. 27:843–856. 2014. View Article : Google Scholar
|
|
75
|
Sessa R, Di Pietro M, Filardo S, Bressan
A, Rosa L, Cutone A, Frioni A, Berlutti F, Paesano R and Valenti P:
Effect of bovine lactoferrin on chlamydia trachomatis infection and
inflammation. Biochem Cell Biol. 95:34–40. 2017. View Article : Google Scholar
|
|
76
|
Valenti P, Frioni A, Rossi A, Ranucci S,
De Fino I, Cutone A, Rosa L, Bragonzi A and Berlutti F: Aerosolized
bovine lactoferrin reduces neutrophils and pro-inflammatory
cytokines in mouse models of Pseudomonas aeruginosa lung
infections. Biochem Cell Biol. 95:41–47. 2017. View Article : Google Scholar
|
|
77
|
Svendsen J, Grant TM, Rennison D, Brimble
MA and Svenson J: Very short and stable lactoferricin-derived
antimicrobial peptides: Design principles and potential uses. Acc
Chem Res. 52:749–759. 2019. View Article : Google Scholar
|
|
78
|
Dial EJ, Romero JJ, Headon DR and
Lichtenberger LM: Recombinant human lactoferrin is effective in the
treatment of Helicobacter Felis-infected mice. J Pharm Pharmacol.
52:1541–1546. 2000. View Article : Google Scholar
|
|
79
|
Rodríguez-Franco DA, Vázquez-Moreno L and
Ramos-Clamont MG: Antimicrobial mechanisms and potential clinical
application of lactoferrin. Rev Latinoam Microbiol. 47:102–111.
2005.In Spanish.
|
|
80
|
Ellison RR and Giehl TJ: Killing of
gram-negative bacteria by lactoferrin and lysozyme. J Clin Invest.
88:1080–1091. 1991. View Article : Google Scholar
|
|
81
|
Dhaenens L, Szczebara F and Husson MO:
Identification, characterization, and immunogenicity of the
lactoferrin-binding protein from Helicobacter pylori. Infect Immun.
65:514–518. 1997. View Article : Google Scholar
|
|
82
|
Kuipers ME, de Vries HG, Eikelboom MC,
Meijer DK and Swart PJ: Synergistic fungistatic effects of
lactoferrin in combination with antifungal drugs against clinical
Candida isolates. Antimicrob Agents Chemother. 43:2635–2641. 1999.
View Article : Google Scholar
|
|
83
|
Hakansson A, Roche H, Mirza S, McDaniel
LS, Brooks-Walter A and Briles DE: Characterization of binding of
human lactoferrin to pneumococcal surface protein A. Infect Immun.
69:3372–3381. 2001. View Article : Google Scholar
|
|
84
|
van der Strate BW, Beljaars L, Molema G,
Harmsen MC and Meijer DK: Antiviral activities of lactoferrin.
Antiviral Res. 52:225–239. 2001. View Article : Google Scholar
|
|
85
|
Campione E, Cosio T, Rosa L, Lanna C, Di
Girolamo S, Gaziano R, Valenti P and Bianchi L: Lactoferrin as
protective natural barrier of respiratory and intestinal mucosa
against coronavirus infection and inflammation. Int J Mol Sci.
21:49032020. View Article : Google Scholar
|
|
86
|
Kell DB, Heyden EL and Pretorius E: The
biology of lactoferrin, an iron-binding protein that can help
defend against viruses and bacteria. Front Immunol. 11:12212020.
View Article : Google Scholar
|
|
87
|
Fu Q, Sun J, Zhang W, Sui X, Yan Z and He
Z: Nanoparticle albumin-bound (NAB) technology is a promising
method for anti-cancer drug delivery. Recent Pat Anticancer Drug
Discov. 4:262–272. 2009. View Article : Google Scholar
|
|
88
|
Yi M, Kaneko S, Yu DY and Murakami S:
Hepatitis C virus envelope proteins bind lactoferrin. J Virol.
71:5997–6002. 1997. View Article : Google Scholar
|
|
89
|
Bennett RM, Merritt MM and Gabor G:
Lactoferrin binds to neutrophilic membrane DNA. Br J Haematol.
63:105–117. 1986. View Article : Google Scholar
|
|
90
|
He J and Furmanski P: Sequence specificity
and transcriptional activation in the binding of lactoferrin to
DNA. Nature. 373:721–724. 1995. View Article : Google Scholar
|
|
91
|
Brandl N, Zemann A, Kaupe I, Marlovits S,
Huettinger P, Goldenberg H and Huettinger M: Signal transduction
and metabolism in chondrocytes is modulated by lactoferrin.
Osteoarthritis Cartilage. 18:117–125. 2010. View Article : Google Scholar
|
|
92
|
Kanyshkova TG, Babina SE, Semenov DV,
Isaeva N, Vlassov AV, Neustroev KN, Kul'minskaya AA, Buneva VN and
Nevinsky GA: Multiple enzymic activities of human milk lactoferrin.
Eur J Biochem. 270:3353–3361. 2003. View Article : Google Scholar
|
|
93
|
Fang JH, Lai YH, Chiu TL, Chen YY, Hu SH
and Chen SY: Magnetic core-shell nanocapsules with dual-targeting
capabilities and co-delivery of multiple drugs to treat brain
gliomas. Adv Healthc Mater. 3:1250–1260. 2014. View Article : Google Scholar
|
|
94
|
Wei M, Guo X, Tu L, Zou Q, Li Q, Tang C,
Chen B, Xu Y and Wu C: Lactoferrin-modified PEGylated liposomes
loaded with doxorubicin for targeting delivery to hepatocellular
carcinoma. Int J Nanomedicine. 10:5123–5137. 2015.PubMed/NCBI
|
|
95
|
Golla K, Cherukuvada B, Ahmed F and
Kondapi AK: Efficacy, safety and anticancer activity of protein
nanoparticle-based delivery of doxorubicin through intravenous
administration in rats. PLoS One. 7:e519602012. View Article : Google Scholar
|
|
96
|
Kumar P, Lakshmi YS, C B, Golla K and
Kondapi AK: Improved safety, bioavailability and pharmacokinetics
of zidovudine through lactoferrin nanoparticles during oral
administration in rats. PLoS One. 10:e1403992015. View Article : Google Scholar
|
|
97
|
Kumari S and Kondapi AK: Receptor-mediated
targeted delivery of DNA using Lactoferrin nanoparticles. Int J
Biol Macromol. 108:401–407. 2018. View Article : Google Scholar
|
|
98
|
Elfinger M, Maucksch C and Rudolph C:
Characterization of lactoferrin as a targeting ligand for nonviral
gene delivery to airway epithelial cells. Biomaterials.
28:3448–3455. 2007. View Article : Google Scholar
|
|
99
|
Yan JK, Qiu WY, Wang YY and Wu JY:
Biocompatible polyelectrolyte complex nanoparticles from
lactoferrin and pectin as potential vehicles for antioxidative
curcumin. J Agric Food Chem. 65:5720–5730. 2017. View Article : Google Scholar
|
|
100
|
Wang J, Li Q, Li K, Ou Y, Han Z, Gao D and
Li J: Effects of adenovirus vectors mediated human lactoferrin cDNA
on mice bearing EMT6 breast carcinoma. Pharmazie. 66:704–709.
2011.
|
|
101
|
Pereira CS, Guedes JP, Goncalves M,
Loureiro L, Castro L, Gerós H, Rodrigues LR and Côrte-Real M:
Lactoferrin selectively triggers apoptosis in highly metastatic
breast cancer cells through inhibition of plasmalemmal V-H+-ATPase.
Oncotarget. 7:62144–62158. 2016. View Article : Google Scholar
|
|
102
|
Lonnerdal B, Jiang R and Du X: Bovine
lactoferrin can be taken up by the human intestinal lactoferrin
receptor and exert bioactivities. J Pediatr Gastroenterol Nutr.
53:606–614. 2011. View Article : Google Scholar
|
|
103
|
Machado S, Alves R, Lima M, Leal I and
Massa A: Cutaneous necrobiotic xanthogranuloma (NXG)-successfully
treated with low dose chlorambucil. Eur J Dermatol. 11:458–462.
2001.PubMed/NCBI
|
|
104
|
Arcella A, Oliva MA, Staffieri S, Aalberti
S, Grillea G, Madonna M, Bartolo M, Pavone L, Giangaspero F,
Cantore G and Frati A: In vitro and in vivo effect of human
lactoferrin on glioblastoma growth. J Neurosurg. 123:1026–1035.
2015. View Article : Google Scholar
|
|
105
|
Son HJ, Lee SH and Choi SY: Human
lactoferrin controls the level of retinoblastoma protein and its
activity. Biochem Cell Biol. 84:345–350. 2006. View Article : Google Scholar
|
|
106
|
Tung YT, Chen HL, Yen CC, Lee PY, Tsai HC,
Lin MF and Chen CM: Bovine lactoferrin inhibits lung cancer growth
through suppression of both inflammation and expression of vascular
endothelial growth factor. J Dairy Sci. 96:2095–2106. 2013.
View Article : Google Scholar
|
|
107
|
Deng M, Zhang W, Tang H, Ye Q, Liao Q,
Zhou Y, Wu M, Xiong W, Zheng Y, Guo X, et al: Lactotransferrin acts
as a tumor suppressor in nasopharyngeal carcinoma by repressing AKT
through multiple mechanisms. Oncogene. 32:4273–4283. 2013.
View Article : Google Scholar
|
|
108
|
Chea C, Miyauchi M, Inubushi T, Febriyanti
Ayuningtyas N, Subarnbhesaj A, Nguyen PT, Shrestha M, Haing S, Ohta
K and Takata T: Molecular mechanism of inhibitory effects of bovine
lactoferrin on the growth of oral squamous cell carcinoma. PLoS
One. 13:e1916832018. View Article : Google Scholar
|
|
109
|
Sabra SA, Elzoghby AO, Sheweita SA, Haroun
M, Helmy MW, Eldemellawy MA, Xia Y, Goodale D, Allan AL and Rohani
S: Self-assembled amphiphilic zein-lactoferrin micelles for tumor
targeted co-delivery of rapamycin and wogonin to breast cancer. Eur
J Pharm Biopharm. 128:156–169. 2018. View Article : Google Scholar
|
|
110
|
Sabra SA, Sheweita SA, Haroun M, Ragab D,
Eldemellawy MA, Xia Y, Goodale D, Allan AL, Elzoghby AO and Rohani
S: Magnetically guided self-assembled protein micelles for enhanced
delivery of dasatinib to human triple-negative breast cancer cells.
J Pharm Sci. 108:1713–1725. 2019. View Article : Google Scholar
|
|
111
|
Abd Elwakil MM, Mabrouk MT, Helmy MW,
Abdelfattah EA, Khiste SK, Elkhodairy KA and Elzoghby AO: Inhalable
lactoferrin-chondroitin nanocomposites for combined delivery of
doxorubicin and ellagic acid to lung carcinoma. Nanomedicine
(Lond). 13:2015–2035. 2018. View Article : Google Scholar
|
|
112
|
Kumari S, Ahsan SM, Kumar JM, Kondapi AK
and Rao NM: Overcoming blood brain barrier with a dual purpose
Temozolomide loaded lactoferrin nanoparticles for combating glioma
(SERP-17-12433). Sci Rep. 7:66022017. View Article : Google Scholar
|
|
113
|
Kumari S and Kondapi AK: Lactoferrin
nanoparticle mediated targeted delivery of 5-fluorouracil for
enhanced therapeutic efficacy. Int J Biol Macromol. 95:232–237.
2017. View Article : Google Scholar
|
|
114
|
Wang H, Tang Y, Fang Y, Zhang M, Wang H,
He Z, Wang B, Xu Q and Huang Y: Reprogramming Tumor Immune
Microenvironment (TIME) and metabolism via biomimetic targeting
Codelivery of Shikonin/JQ1. Nano Lett. 19:2935–2944. 2019.
View Article : Google Scholar
|
|
115
|
Kumar P, Lakshmi YS and Kondapi AK: An
oral formulation of efavirenz-loaded lactoferrin nanoparticles with
improved biodistribution and pharmacokinetic profile. HIV Med.
18:452–462. 2017. View Article : Google Scholar
|
|
116
|
Zhang ZH, Wang XP, Ayman WY, Munyendo WL,
Lv HX and Zhou JP: Studies on lactoferrin nanoparticles of gambogic
acid for oral delivery. Drug Deliv. 20:86–93. 2013. View Article : Google Scholar
|
|
117
|
Shankaranarayanan JS, Kanwar JR,
Al-Juhaishi AJ and Kanwar RK: Doxorubicin conjugated to
immunomodulatory anticancer lactoferrin displays improved
cytotoxicity overcoming prostate cancer chemo resistance and
inhibits tumour development in TRAMP mice. Sci Rep. 6:320622016.
View Article : Google Scholar
|