Pancreatic ductal adenocarcinoma (PDAC) is a
high-mortality malignancy with a 5-year survival rate of 9% for all
stages and ~90% of patients are at advanced stages exhibiting a
5-year survival rate of 3% when diagnosed (1). PDAC is expected to become the second
leading cause of cancer-related mortality worldwide within the next
decade, due to its gradually increasing mortality rates (2). For patients who are not screened
early enough, late-stage PDAC remains difficult to treat. Systemic
chemotherapy, which includes neoadjuvant therapy, adjuvant therapy
and first-line or subsequent therapy, is imperative for metastatic
and locally advanced PDAC, as well as for other stages of PDAC.
Gemcitabine has been established as the main first-line drug for
PDAC chemotherapy. Other drugs, including albumin-bound paclitaxel,
5-fluorouracil (5-FU), capecitabine, cisplatin, irinotecan,
oxaliplatin and erlotinib, are used in various combinations or as
monotherapy, according to disease stage and patient status
(3). However, due to acquired
resistance or side effects during treatment, the efficacy for
patients with PDAC is not satisfactory and the 5-year survival rate
has not been significantly improved.
In recent years, an association between microbiomes
and the occurrence and development of PDAC have been identified.
Systems biology provides a more comprehensive and multiparametric
understanding of drug metabolism. The microbiome, which is the
comprehensive genomic information encoded by the microbiota and its
ecosystem, products and host environment, has therefore been
explored as a direction for therapy (4). Although the terms 'microbiota' and
'microbiome' are used interchangeably, the microbiota should be
studied more comprehensively from the perspective of omics, while
the functional microbiome is indispensable (5,6).
Therefore, the 'microbiome' has been fully summarized and its
function has been described. The composition of the microbiota is
primarily determined by host genes and affected by extrinsic
factors, including diet (7),
antibiotics (8), surgery
(9) and some drugs [e.g., proton
pump inhibitors (10)]. In
addition, pancreatic acini secrete peptides that can modulate the
gut microbiota, relying on the Ca2+ channel calcium
release-activated calcium channel protein 1 (11,12). The specific relationship between
the microbiome and cancer, including gastrointestinal (13), breast (14), liver (15), esophageal cancer (16) and PDAC (17), has attracted the attention of
researchers. Specifically, a large number of clinical studies have
shown that the microbiome mediates the response to systemic therapy
and that there are therapy-related changes in microbiota
composition (18,19).
The majority of studies examining the microbiome in
PDAC have focused on carcinogenicity, as the data on therapeutic
action are preliminary. However, although the complex and important
role of the microbiome in PDAC therapy requires in-depth study,
related research is limited and the mechanisms involved remain to
be fully elucidated. The aim of the present review was to outline
recent microbiome research-related developments and interesting
discoveries in systemic therapy drugs for PDAC and illustrate the
underlying mechanisms. Promising research directions with regards
to the involvement of the microbiome in PDAC treatment were also
discussed and proposed.
The majority of patients with PDAC require systemic
therapy, whether that consists of surgery followed by adjuvant
therapy, neoadjuvant therapy or palliative adjuvant therapy.
However, since the majority of patients with PDAC develop
resistance or toxicity to drugs, the treatment needs to be
delivered at a lower dose or suspended before the scheduled end
date, which leads to it being ineffective. Therefore, clarifying
the complex mechanisms to improve treatment response is important.
Functional research on the microbiota has suggested that it has the
potential to induce PDAC treatment resistance or toxicity. In this
section, the findings of previous studies with regards to the
relationship between the microbiome and several drugs for the
treatment of PDAC, including gemcitabine, 5-FU, capecitabine,
oxaliplatin, irinotecan, cisplatin and erlotinib (Tables II and III), were described in detail
(3).
The combination of gemcitabine and albumin-bound
paclitaxel has been upgraded as a category 1 recommendation
(3). Kesh et al (46) found that microbial dysbiosis
increases resistance to this combination. In a pancreatic
tumor-bearing mouse model of type II diabetes, Enterobacter
cloacae and carbohydrate- and lipid-metabolizing bacteria are
enriched. This enrichment of microbial metabolites prevents tumor
cells from chemotherapy-induced accumulation of reactive oxygen
species, leading to resistance (46); however, in that study, the
treatment regimen was a combination of gemcitabine and paclitaxel
and no study has yet focused on microbiome-induced paclitaxel
monotherapy resistance. Although lactic acid bacteria,
bifidobacteria and other bacteria of intestinal origin are not
susceptible to paclitaxel (47),
the bacterial populations are altered in paclitaxel-treated mice:
butyrate-producing bacteria, including Roseburia,
Eubacterium and Erysipelotrichaceae, are depleted
(48) and paclitaxel treatment
decreases the abundance of Akkermansia muciniphila and
alters that of other bacterial taxa, which are drivers of
chemotherapy-induced peripheral neuropathy (CIPN) (49). In addition, paclitaxel-containing
chemotherapeutic combinations are more likely to result in
Clostridioides difficile infection (50). Therefore, the antitumor effect of
paclitaxel could be improved by reversing paclitaxel-induced gut
microbiota dysbiosis (51).
Fluoropyrimidine is also a first-line
chemotherapeutic drug for patients with PDAC, including 5-FU,
capecitabine and TAS-1. 5-FU is frequently administered alongside
FOLFIRINOX/modified FOLFIRINOX and with or without leucovorin
(3). Capecitabine, a precursor of
5-FU, can be administered alone or co-administered with gemcitabine
to patients with PDAC (3). The
majority of studies on 5-FU focus on its effects on the abundance
of microbiota constituents and induction of mucositis (52–54). The main mechanisms of toxicity
have been demonstrated. First, 5-FU alters microbiota diversity by
decreasing Lactobacillus and Streptococcus abundance
and increasing Clostridium and Staphylococcus
abundance, leading to a decrease in the secretion of mucin, a
principal factor in the physiological defense of the
gastrointestinal mucosa (52,55). Therefore, supplementation with the
genera Lactobacillus and Bifidobacterium could
protect the human gastrointestinal tract from chemotherapy
(56). Secondly, the lack of a
detoxification enzyme of 5-FU (hepatic dihydropyridine
dehydrogenase) may lead to an increase in the systemic
concentrations of 5-FU in the blood and enhanced toxicity. The gut
microbiota serves a critical role in that process. Specific
bacteria, including Bacteroides species (B. vulgatus,
B. thetaiotaomicron, B. fragilis, B. uniformis
and B. eggerthii) can hydrolyze sorivudine to
(E)-5-(2-bromovinyl) uracil, which inactivates the detoxification
enzyme (57).
At present, the understanding of whether the gut
microbiota influences the antitumor efficacy of 5-FU treatment is
limited. A previous study proposes that 5-FU together with ABX, an
antibiotic cocktail, markedly reduces the antitumor effect of 5-FU
and the gut bacterial diversity and communities show significant
changes compared with those after 5-FU alone or 5-FU plus probiotic
treatments (58). This means that
the gut flora dysbiosis contributes to the induction of 5-FU
resistance. Mycoplasma hyorhinis, which was mentioned in the
gemcitabine and paclitaxel section, also degrades
fluoropyrimidines, including 5-FU, by TP to their inactive bases.
By contrast, capecitabine, which must be metabolized to
5-fluoro-5′-deoxyuridine (5′DFUR), can benefit from TP activity
(59). Fusobacterium
nucleatum, an anaerobic bacterium that is parasitic in the oral
cavity and highly abundant in the gut microbiota, may promote 5-FU
and oxaliplatin resistance by targeting TLR4 and myeloid
differentiation primary response 88 (MYD88) innate immune signaling
and then downregulating the expression of microRNA
(miR)-18a* and miR-4802, which activate the autophagy
pathway by increasing Unc-51 like autophagy activating kinase 1 and
autophagy related 7 expression (60). In addition, another study
demonstrated that F. nucleatum confers resistance to 5-FU by
upregulating the expression of baculoviral IAP repeat containing 3
via the TLR4/NF-κB pathway (61).
These two studies suggest that fully elucidating the mechanism of
the specific microbiota constituents inducing chemoresistance poses
a major challenge as the same bacteria may have two or several
regulatory pathways that alter drug response. García-González et
al (62) found that
Escherichia coli and Comamonas increase 5-FU efficacy
by bacterial nucleotide metabolism and lead to the sterility of
C. elegans, a powerful model system to study the effects of
the microbiota on chemotherapeutics. In addition to this mechanism,
another study reports that E. coli vitamin B6 and B9
metabolism are essential for 5-FU efficacy in the same C.
elegans model (63).
Cisplatin is a platinum-based potent antitumor agent
used for PDAC, along with gemcitabine, but only for patients with
known breast cancer type 1/2 or partner and localizer of BRCA2
mutations (3,64). Cisplatin causes tumor cytotoxicity
by forming platinum DNA adducts and intrastrand cross-links, as
well as through the modulation of the immune system (65,66). Few studies have focused on
microbiome-mediated cisplatin resistance (67,68). Gram-positive bacterial antibiotics
can weaken its antitumor effect (67), but the specific mechanism remains
to be elucidated. In addition, the majority of gastrointestinal
toxicities caused by cisplatin have been attributed to various
events, such as oxidative stress and inflammation (68). Although there is no direct
evidence that the microbiome induces toxicity, the combination of
cisplatin with the commensal microbiota or agents that balance it
could ameliorate cisplatin-induced gastrointestinal toxicity
(69–72), as well as other adverse effects
(73–75). These studies suggest the existence
of a crucial intrinsic link between the microbiome and cisplatin,
but additional research should focus on and clarify the
mechanism.
The efficacy of oxaliplatin is limited by peripheral
neuropathies, as well as gastrointestinal toxicity (78,79), but whether the microbiota induces
or mediates oxaliplatin toxicity has rarely been reported. Shen
et al (80) reports that
the gut microbiota may promote the development of
oxaliplatin-induced pain, which can be reduced in germ-free mice
and mice pretreated with antibiotics. Mechanistically, the dorsal
root ganglion (DRG) is a key anatomical site for CIPN pathogenesis
(81). Oxaliplatin may directly
alter the gut microbiota and increase LPS levels in the DRG
(80). LPS derived from the gut
microbiota targets TLR4, which is expressed on hematopoietic cells
and then stimulates primary macrophages, leading to the production
of inflammatory cytokines in the DRG, such as IL-6 and TNF-α
(80). Gastrointestinal injury is
one of the toxicities induced by oxaliplatin, an effect that may be
associated with alterations in the gut microbiota and activation of
inflammatory processes (82,83). Accordingly, fecal microbiota
transplantation (FMT) can alleviate the injury (84).
Although FOLFIRINOX causes marked improvements in
patients with metastatic PDAC compared with gemcitabine, the 3/4
toxicity rate is clearly greater (85), which always leads to a dose
reduction. Irinotecan (also known as CPT-11) is the main drug in
the FOLFIRINOX regimen that occasionally induces severe toxicities,
which limit its use and efficacy (86). Delayed-onset diarrhea is a common
clinical adverse effect. The most likely mechanism of the induction
of severe diarrhea is that the β-glucuronidase secreted by the gut
microbiota dissociates SN-38G to SN-38, which is responsible for
both antitumor activity and dose-limiting toxicity (87,88). This underlying mechanism reveals
that the gut microflora serves a critical role in the intestinal
toxicity of irinotecan (89),
even though the association appears to be controversial and
mechanisms other than this enzyme, such as TLR4-dependent
mechanisms (90), may be involved
in irinotecan treatment (91,92).
Erlotinib, an EGFR tyrosine kinase inhibitor,
increases overall survival when combined with gemcitabine (93) and this combination therapy is
another option for patients with locally advanced or metastatic
disease; it has a good performance status and is a category 1
recommendation for patients with metastatic disease in the National
Comprehensive Cancer Network guidelines (3). There is little research on the
relationship between the microbiome and erlotinib. Two studies
indicate that certain bacteria of intestinal origin had no
susceptibility to erlotinib and did not induce changes in
intestinal tissue morphology, but whether there were changes in the
abundance of the gut microbiome remain unknown (47,94). Heshiki et al (95) found that baseline microbiota
composition could predict treatment response and the responder
bacteria (Bacteroides ovatus and B. xylanisolvens)
increase the efficacy of erlotinib in mice more than the
non-responder bacteria (Cenarchaeum symbiosum and
Ruminococcus gnavus) when administered by oral gavage.
Mechanistically, the responder bacteria may synergistically
upregulate chemokines involved in T-cell recruitment and then
enhance erlotinib efficacy (95).
Pharmacomicrobiomics, a new discipline exploring the
interactions between drugs and microbes (96), has the potential to broaden our
understanding of the interplay between the microbiome and systemic
therapy for PDAC. In addition, clinical metagenomic next-generation
sequencing has provided a glimpse into the monitoring of
chemotherapy regimens (97). In
addition, the increased knowledge obtained in this field can
potentially generate novel chemotherapeutic or subsequent therapy
approaches to enhance efficacy and abrogate side effects by
manipulating the α- and β-diversity of the microbiota to
individualize treatment. The present review provided a detailed
overview of the association between the microbiome and systemic
therapy drugs for PDAC. However, since the majority of the studies'
objectives are not PDAC, the evidence obtained in the present study
remains limited. Therefore, carrying out research to further
elucidate the role of the microbiome in PDAC systemic therapy is
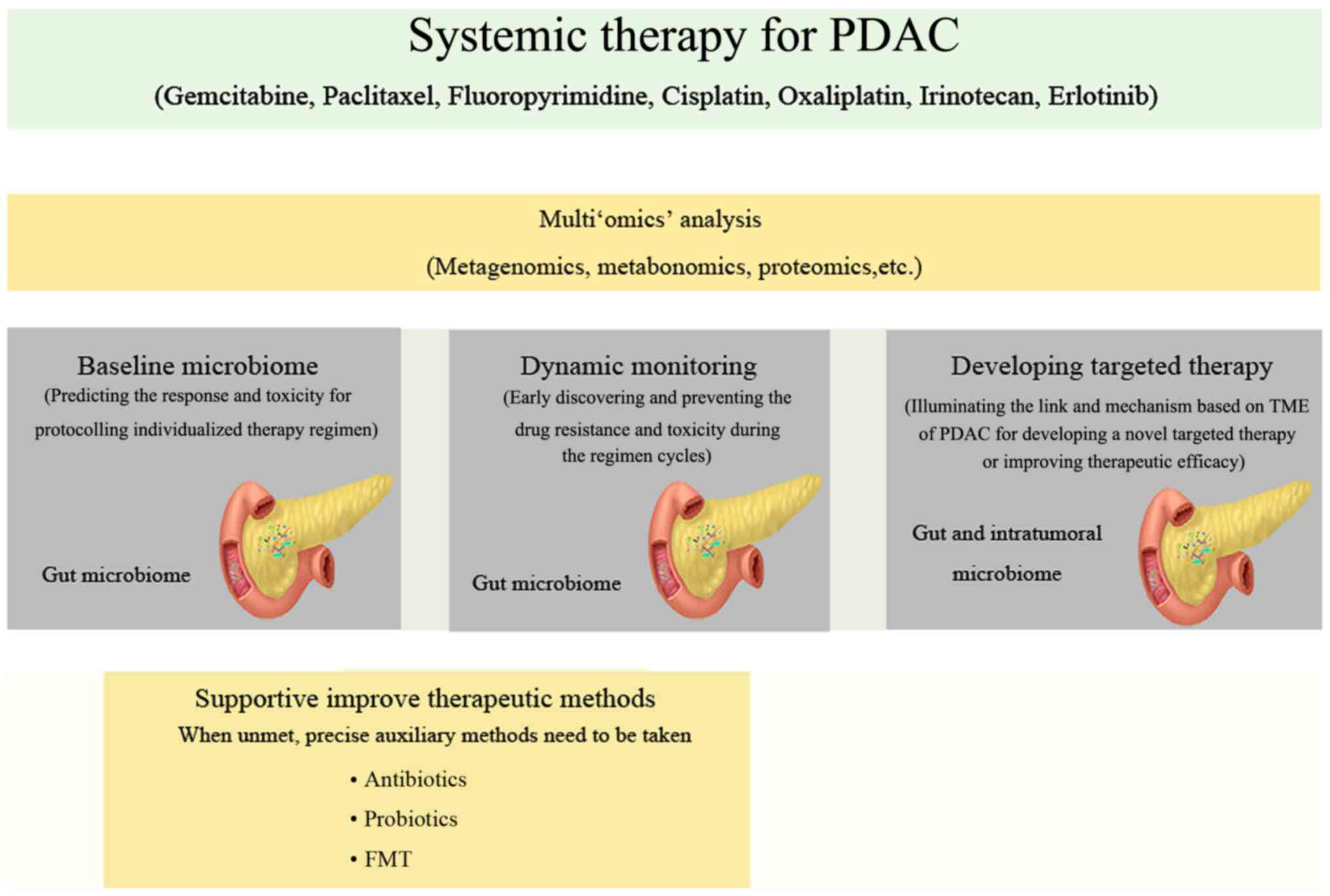
urgent. From the perspective of the present study, four main
aspects need additional attention in future research (Fig. 1).
Numerous clinical studies have investigated whether
the baseline gut microbiota predicts the clinical response to
systemic cancer therapy or bacterial infection (19,98–100). Aarnoutse et al (101) profile the microbiota composition
before, during and after three cycles of systemic treatment with
capecitabine or TAS-102 and attempt to detect a microbiota
composition that predicts chemotoxicity in patients with metastatic
and/or resectable colorectal cancer.
In addition, the baseline gut microbiome can also be
used to predict the toxicity of chemotherapy. Stringer et al
(102) analyze stool and serum
samples from 26 patients with cancer receiving chemotherapy. The
type of cancer and chemotherapy regimen both differed from patient
to patient; the latter included capecitabine, cisplatin/5-FU,
FOLFOX, 5-FU/folinic acid, COFF plus paclitaxel and carboplatin
plus gemcitabine. Specific bacteria were enriched (including E.
coli and Staphylococcus spp.) or depleted (including
Lactobacillus spp., Bifidobacterium spp.,
Bacteroides spp. and Enterococcus spp.) in the
majority of patients with chemotherapy-induced diarrhea (CD) and
alterations in inflammation and circulating matrix
metalloproteinases were observed (102). These changes may serve as
predictive biomarkers of chemotherapeutic toxicity. In addition,
the biomarkers based on the baseline gut microbiome could be
combined with additional biomarkers, including metabolites.
Relevant clinical studies of PDAC are not yet
available. However, ongoing or previous studies have suggested that
the baseline microbiota may be able to predict treatment response.
Related clinical research on PDAC exploring different regimens,
stages and performance statuses should therefore be performed. This
research can be more comprehensive, investigating not only the gut
or intratumor microbiota but also the related metabolites and other
small molecules, which could be generalized to the aforementioned
microbiome. That accumulated knowledge could help build a systemic
and comprehensive prediction model for the response to chemotherapy
regimens.
Systemic cancer therapies can affect the entire
body, as well as the human microbiota composition and abundance,
including that of gut and intratumor environment microbiota. The
majority of existing studies have focused on the link between
intestinal barrier dysfunction and toxicity (44,48,52,103–108) In addition, whether the drugs
induce changes in the microbiome and then lead to resistance is
unclear. This phenomenon is called function-mediated diversity and
certain studies have hinted at that possibility.
These limited studies suggest that multiple cycles
of chemotherapy cause changes in the gut microbiota and that
altered organisms may cause drug resistance or sensitivity. This
alteration in microbial diversity may be mediated and accelerated
by the functional response to chemotherapy, with the
microecosystems changing towards a more favorable environment
(109), which has been destroyed
by systemic chemotherapy. However, no related research has been
conducted for PDAC. Concentrating on dynamic changes and functional
response diversity will help clinicians deal with resistance or
impede infection quickly and effectively.
The microbiota is complex and certain studies have
illustrated that certain bacteria can mediate the resistance and
toxicity of certain chemotherapy drugs. However, the specific
mechanism may be multifactorial. For example, Fusobacterium,
an oral bacterium detected in PDAC tissue (20,27), mediates 5-FU resistance by
activating the autophagy and TLR4/NF-κB pathways (60,61). In addition, identifying specific
bacteria and mechanisms is conducive to developing a new targeted
therapy to improve resistance. For irinotecan, targeted gut
microbial enzyme inhibitors may be a new potential method to
alleviate gastrointestinal tract toxicity and enhance efficacy
(110,111). Due to the structural and
functional differences in β-glucuronidases from the human gut
microbes (112,113), β-glucuronidase inhibitors should
be selective and not affect the survival of the microbiota
(114,115); therefore, the molecular
mechanism should be specific. In addition, when analyzing the
species and functional composition of the gut microbiome, the focus
cannot only be placed on the abundant species, which do not always
equate to abundant molecular functions (116). Functional analysis needs to be
specific to a particular microbiome, including species with low
abundance.
In addition, an increased understanding of the
complex mechanisms underlying the role of the microbiome in the
systemic therapy of PDAC needs to include the role of the
microbiome in the TME, which is composed of a minority of malignant
cells, endothelial cells, immune cells, fibroblasts and
extracellular matrix (117), as
well as the microbiota. Therapeutic failures of chemotherapy,
particularly gemcitabine, have been attributed to the PDAC
microenvironment (118). Given
the complexity of the PDAC TME, the cause of resistance to
chemotherapy is multifactorial and the microbiome may serve a
moderate role. For example, intrapancreatic and gut-specific
microbes serve as helpers in the shaping of the immunosuppressive
PDAC TME, which leads to tumor-associated macrophages (TAMs)
becoming highly abundant in PDAC, ranging from M1-like TAMs to
immune-suppressive M2-like TAMs (33). In this process, deoxycytidine
macrophages release competitively inhibits gemcitabine uptake and
metabolism and leads to chemoresistance (119). Therefore, the microbiota may
induce gemcitabine resistance by shaping the PDAC intratumoral
immune microenvironment. The aforementioned data markedly indicate
that the microbiome can mediate therapeutic responses
systematically through numerous mechanisms and that these can also
be structured as the 'TIMER' mechanistic framework (120). Elucidating these mechanisms will
be conducive to developing a novel targeted therapy-based
microbiome.
As described aforementioned, the microbiota can
provide a novel way to enhance the efficacy and reduce the toxicity
of chemotherapeutic approaches. Several strategies can be used to
synergize with systemic therapy to improve efficacy, such as
antibiotics, probiotics, FMT, prebiotics, diet and physical
activity, by modulating the composition of the microbiome (121,122). However, future studies should
accurately improve our understanding of the value of these
synergistic methods.
The use of antibiotics to remodel the diversity and
constitution of the microbiota and alleviate toxicity have proven
to be effective (123–125); however, they may also impair the
response to chemotherapy. Iida et al (76) note that antibiotics impair
oxaliplatin therapy efficacy by decreasing ROS production, which is
the reason why anticancer drugs work (77) and are similarly regulated by
antibiotics. The overuse of antibiotics targeting anaerobes is
associated with a poor prognosis in patients with hepatocellular
carcinoma who have undergone chemotherapy (126). In addition, the elimination of
symbiotic bacteria increases the risk of pathogenic
bacteria-induced infection (127).
In addition to probiotics, other combinations
should be explored, such as metabolites or digestive enzymes.
Identifying an improved combination of probiotics can significantly
reduce the untoward effects of chemotherapy (136). Urolithin A, which is the main
metabolite produced by the human gut microbiota, can potentiate the
effects of both 5-FU and 5-dFUR on colon cancer cells (137). Probiotics supplemented with
digestive enzymes can restore the gut microbial community and
protect against 5-FU-induced gut dysbiosis (56).
There are few studies on FMT for improving systemic
anticancer therapy. Le Bastard et al (138) assess the efficacy of FMT in
5-FU-induced gut dysbiosis in a mouse model. FMT ameliorates the
disruption of the intestinal microbiota by significantly enriching
the species with anti-inflammatory properties in mice (138). The results show that FMT has the
potential to improve the resistance and toxicity induced by
systemic therapy for PDAC. However, due to its uncertainties, FMT
might increase the chance of infection and fecal donor selection
and screening are difficult. Therefore, selective microbiota
transplantation may be a better choice and additional studies
should be carried out to investigate that option.
Although few of these studies have focused on PDAC,
the mechanism underlying drug alterations by the microbiome may be
similar. Microbiome studies provide a novel direction for the
improvement of the response to systemic therapy for PDAC. A deep
exploration of the mechanism and the relationship between the
microbiome and systemic therapy drugs for PDAC is essential, due to
the low survival rate and chemotherapeutic resistance of PDAC. In
clinical practice, the combination of the microbiota and its
metabolites and metabolic pathways could be used to establish a
model for predicting the response to systemic chemotherapy
regimens, which can be conducted flexibly and individually. During
regimen cycles, the microbiota is destroyed, inducing resistance.
Therefore, dynamic monitoring of the gut microbiota and timely
adjustment of the regimen or restoration of the composition of the
microbiome through the use of cooperative strategies may prove
beneficial. Admittedly, the model and detection of the microbiome
composition of patients should be fast, robust and inexpensive. In
addition, mechanistic studies of the microbiome could provide novel
targeted therapies or synergetic schemes to establish personalized
medicine for each patient. In conclusion, the relationship and main
mechanism between the microbiome and drugs for PDAC treatment were
outlined in the present review and certain directions for future
research were proposed.
Data sharing is not applicable to this article, as
no data sets were generated or analyzed during the current
study.
BT conceived and supervised the work. XH, ML and SH
researched data and contributed equally to discussion of content,
XH and ML wrote the manuscript. All authors reviewed and approved
the final manuscript.
Not applicable.
Not applicable.
Not applicable.
This study was supported by Sichuan Science and Technology
Program (grant no. 2021YFS0234).
|
1
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2019. CA Cancer J Clin. 69:7–34. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Rahib L, Smith BD, Aizenberg R, Rosenzweig
AB, Fleshman JM and Matrisian LM: Projecting cancer incidence and
deaths to 2030: The unexpected burden of thyroid, liver, and
pancreas cancers in the United States. Cancer Res. 74:2913–2921.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Network (NCCN): NCC: Clinical Practice
Guidelines in Oncology. Pancreatic Adenocarcinoma, Version 1. NCCN;
Pennsylvania: 2020, https://www.nccn.org/professionals/physician_gls/pdf/pancreatic.pdf.
Accessed November 26, 2019.
|
|
4
|
Marchesi JR, Adams DH, Fava F, Hermes GDA,
Hirschfield GM, Hold G, Quraishi MN, Kinross J, Smidt H, Tuohy KM,
et al: The gut microbiota and host health: A new clinical frontier.
Gut. 65:330–339. 2016. View Article : Google Scholar
|
|
5
|
Heintz-Buschart A and Wilmes P: Human gut
microbiome: Function matters. Trends Microbiol. 26:563–574. 2018.
View Article : Google Scholar
|
|
6
|
Human Microbiome Project Consortium:
Structure, function and diversity of the healthy human microbiome.
Nature. 486:207–214. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Kolodziejczyk AA, Zheng D and Elinav E:
Diet-microbiota interactions and personalized nutrition. Nat Rev
Microbiol. 17:742–753. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Ianiro G, Tilg H and Gasbarrini A:
Antibiotics as deep modulators of gut microbiota: Between good and
evil. Gut. 65:1906–1915. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Guyton K and Alverdy JC: The gut
microbiota and gastrointestinal surgery. Nat Rev Gastroenterol
Hepatol. 14:43–54. 2017. View Article : Google Scholar
|
|
10
|
Imhann F, Bonder MJ, Vila AV, Fu J,
Mujagic Z, Vork L, Tigchelaar EF, Jankipersadsing SA, Cenit MC,
Harmsen HJ, et al: Proton pump inhibitors affect the gut
microbiome. Gut. 65:740–748. 2016. View Article : Google Scholar
|
|
11
|
Tilg H and Adolph TE: Beyond digestion:
The pancreas shapes intestinal microbiota and immunity. Cell Metab.
25:495–496. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Ahuja M, Schwartz DM, Tandon M, Son A,
Zeng M, Swaim W, Eckhaus M, Hoffman V, Cui Y, Xiao B, et al:
Orai1-mediated antimicrobial secretion from pancreatic acini shapes
the gut microbiome and regulates gut innate immunity. Cell Metab.
25:635–646. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Panebianco C, Potenza A, Andriulli A and
Pazienza V: Exploring the microbiota to better understand
gastrointestinal cancers physiology. Clin Chem Lab Med.
56:1400–1412. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Balhouse BN, Patterson L, Schmelz EM,
Slade DJ and Verbridge SS: N-(3-oxododecanoyl)-L-homoserine lactone
interactions in the breast tumor microenvironment: Implications for
breast cancer viability and proliferation in vitro. PLoS One.
12:e01803722017. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Grąt M, Wronka KM, Krasnodębski M, Masior
L, Lewandowski Z, Kosińska I, Grąt K, Stypułkowski J, Rejowski S,
Wasilewicz M, et al: Profile of gut microbiota associated with the
presence of hepatocellular cancer in patients with liver cirrhosis.
Transplant Proc. 48:1687–1691. 2016. View Article : Google Scholar
|
|
16
|
Zaidi AH, Kelly LA, Kreft RE, Barlek M,
Omstead AN, Matsui D, Boyd NH, Gazarik KE, Heit MI, Nistico L, et
al: Associations of microbiota and toll-like receptor signaling
pathway in esophageal adenocarcinoma. BMC Cancer. 16:522016.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Wei MY, Shi S, Liang C, Meng QC, Hua J,
Zhang YY, Liu J, Zhang B, Xu J and Yu XJ: The microbiota and
microbiome in pancreatic cancer: More influential than expected.
Mol Cancer. 18:972019. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Roy S and Trinchieri G: Microbiota: A key
orchestrator of cancer therapy. Nat Rev Cancer. 17:271–285. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Aarnoutse R, Ziemons J, Penders J, Rensen
SS, de Vos-Geelen J and Smidt ML: The clinical link between human
intestinal microbiota and systemic cancer therapy. Int J Mol Sci.
20:41452019. View Article : Google Scholar :
|
|
20
|
Fan X, Alekseyenko AV, Wu J, Peters BA,
Jacobs EJ, Gapstur SM, Purdue MP, Abnet CC, Stolzenberg-Solomon R,
Miller G, et al: Human oral microbiome and prospective risk for
pancreatic cancer: A population-based nested case-control study.
Gut. 67:120–127. 2018. View Article : Google Scholar
|
|
21
|
Torres PJ, Fletcher EM, Gibbons SM, Bouvet
M, Doran KS and Kelley ST: Characterization of the salivary
microbiome in patients with pancreatic cancer. PeerJ. 3:e13732015.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Olson SH, Satagopan J, Xu Y, Ling L, Leong
S, Orlow I, Saldia A, Li P, Nunes P, Madonia V, et al: The oral
microbiota in patients with pancreatic cancer, patients with IPMNs,
and controls: A pilot study. Cancer Causes Control. 28:959–969.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Michaud DS, Izard J, Wilhelm-Benartzi CS,
You DH, Grote VA, Tjønneland A, Dahm CC, Overvad K, Jenab M,
Fedirko V, et al: Plasma antibodies to oral bacteria and risk of
pancreatic cancer in a large European prospective cohort study.
Gut. 62:1764–1770. 2013. View Article : Google Scholar
|
|
24
|
Farrell JJ, Zhang L, Zhou H, Chia D,
Elashoff D, Akin D, Paster BJ, Joshipura K and Wong DT: Variations
of oral microbiota are associated with pancreatic diseases
including pancreatic cancer. Gut. 61:582–588. 2012. View Article : Google Scholar
|
|
25
|
Sun H, Zhao X, Zhou Y, Wang J, Ma R, Ren
X, Wang H and Zou L: Characterization of oral microbiome and
exploration of potential biomarkers in patients with pancreatic
cancer. Biomed Res Int. 2020:47124982020. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Vogtmann E, Han Y, Caporaso JG, Bokulich
N, Mohamadkhani A, Moayyedkazemi A, Hua X, Kamangar F, Wan Y, Suman
S, et al: Oral microbial community composition is associated with
pancreatic cancer: A case-control study in Iran. Cancer Med.
9:797–806. 2020. View Article : Google Scholar
|
|
27
|
Mitsuhashi K, Nosho K, Sukawa Y, Matsunaga
Y, Ito M, Kurihara H, Kanno S, Igarashi H, Naito T, Adachi Y, et
al: Association of Fusobacterium species in pancreatic cancer
tissues with molecular features and prognosis. Oncotarget.
6:7209–7220. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Half E, Keren N, Dorfman T, Reshef L,
Lachter I, Kluger Y, Konikoff F and Gphna U: Specific changes in
fecal microbiota may differentiate Pancreatic Cancer patients from
healthy individuals. Ann Oncol. 26:iv482015. View Article : Google Scholar
|
|
29
|
Ren Z: Gut microbial profile analysis by
MiSeq sequencing of pancreatic carcinoma patients in China.
Oncotarget. 8:95176–95191. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Sethi V, Kurtom S, Tarique M, Lavania S,
Malchiodi Z, Hellmund L, Zhang L, Sharma U, Giri B, Garg B, et al:
Gut microbiota promotes tumor growth in mice by modulating immune
response. Gastroenterology. 155:33–37.e36. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Half E, Keren N, Reshef L, Dorfman T,
Lachter I, Kluger Y, Reshef N, Knobler H, Maor Y, Stein A, et al:
Fecal microbiome signatures of pancreatic cancer patients. Sci Rep.
9:168012019. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Riquelme E, Zhang Y, Zhang L, Montiel M,
Zoltan M, Dong W, Quesada P, Sahin I, Chandra V, Lucas AS, et al:
Tumor microbiome diversity and composition influence pancreatic
cancer outcomes. Cell. 178:795–806.e712. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Pushalkar S, Hundeyin M, Daley D,
Zambirinis CP, Kurz E, Mishra A, Mohan N, Aykut B, Usyk M, Torres
LE, et al: The pancreatic cancer microbiome promotes oncogenesis by
induction of innate and adaptive immune suppression. Cancer Discov.
8:403–416. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Ikebe M, Kitaura Y, Nakamura M, Tanaka H,
Yamasaki A, Nagai S, Wada J, Yanai K, Koga K, Sato N, et al:
Lipopolysaccharide (LPS) increases the invasive ability of
pancreatic cancer cells through the TLR4/MyD88 signaling pathway. J
Surg Oncol. 100:725–731. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Eibl G and Rozengurt E: KRAS, YAP, and
obesity in pancreatic cancer: A signaling network with multiple
loops. Semin Cancer Biol. 54:50–62. 2019. View Article : Google Scholar
|
|
36
|
Aykut B, Pushalkar S, Chen R, Li Q,
Abengozar R, Kim JI, Shadaloey SA, Wu D, Preiss P, Verma N, et al:
The fungal mycobiome promotes pancreatic oncogenesis via activation
of MBL. Nature. 574:264–267. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Gaida MM, Mayer C, Dapunt U, Stegmaier S,
Schirmacher P, Wabnitz GH and Hänsch GM: Expression of the bitter
receptor T2R38 in pancreatic cancer: Localization in lipid droplets
and activation by a bacteria-derived quorum-sensing molecule.
Oncotarget. 7:12623–12632. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Mendez R, Kesh K, Arora N, Martino LD,
McAllister F, Merchant N and Banerjee S and Banerjee S: Microbial
dysbiosis and polyamine metabolism as predictive markers for early
detection of pancreatic cancer. Carcinogenesis. 41:561–570. 2020.
View Article : Google Scholar :
|
|
39
|
Burris HA III, Moore MJ, Andersen J, Green
MR, Rothenberg ML, Modiano MR, Cripps MC, Portenoy RK, Storniolo
AM, Tarassoff P, et al: Improvements in survival and clinical
benefit with gemcitabine as first-line therapy for patients with
advanced pancreas cancer: A randomized trial. J Clin Oncol.
15:2403–2413. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Von Hoff DD, Ramanathan RK, Borad MJ,
Laheru DA, Smith LS, Wood TE, Korn RL, Desai N, Trieu V, Iglesias
JL, et al: Gemcitabine plus nab-paclitaxel is an active regimen in
patients with advanced pancreatic cancer: A phase I/II trial. J
Clin Oncol. 29:4548–4554. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Gnanamony M and Gondi CS: Chemoresistance
in pancreatic cancer: Emerging concepts. Oncol Lett. 13:2507–2513.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Neale GA, Mitchell A and Finch LR: Enzymes
of pyrimidine deoxyribonucleotide metabolism in Mycoplasma mycoides
subsp. Mycoides J Bacteriol. 156:1001–1005. 1983. View Article : Google Scholar
|
|
43
|
Voorde JV, Sabuncuoğlu S, Noppen S, Hofer
A, Ranjbarian F, Fieuws S, Balzarini J and Liekens S:
Nucleoside-catabolizing enzymes in mycoplasma-infected tumor cell
cultures compromise the cytostatic activity of the anticancer drug
gemcitabine. J Biol Chem. 289:13054–13065. 2014. View Article : Google Scholar
|
|
44
|
Geller LT, Barzily-Rokni M, Danino T,
Jonas OH, Shental N, Nejman D, Gavert N, Zwang Y, Cooper ZA, Shee
K, et al: Potential role of intratumor bacteria in mediating tumor
resistance to the chemotherapeutic drug gemcitabine. Science.
357:1156–1160. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Lehouritis P, Cummins J, Stanton M, Murphy
CT, McCarthy FO, Reid G, Urbaniak C, Byrne WL and Tangney M: Local
bacteria affect the efficacy of chemotherapeutic drugs. Sci Rep.
5:145542015. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Kesh K, Mendez R, Abdelrahman L and
Banerjee S and Banerjee S: Type 2 diabetes induced microbiome
dysbiosis is associated with therapy resistance in pancreatic
adenocarcinoma. Microb Cell Fact. 19:752020. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Florez AB, Sierra M, Ruas-Madiedo P and
Mayo B: Susceptibility of lactic acid bacteria, bifidobacteria and
other bacteria of intestinal origin to chemotherapeutic agents. Int
J Antimicrob Agents. 48:547–550. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Loman BR, Jordan KR, Haynes B, Bailey MT
and Pyter LM: Chemotherapy-induced neuroinflammation is associated
with disrupted colonic and bacterial homeostasis in female mice.
Sci Rep. 9:16490. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Ramakrishna C, Corleto J, Ruegger PM,
Logan GD, Peacock BB, Mendonca S, Yamaki S, Adamson T, Ermel R,
McKemy D, et al: Dominant role of the gut microbiota in
chemotherapy induced neuropathic pain. Sci Rep. 9:20324. 2019.
View Article : Google Scholar
|
|
50
|
Peretz A, Shlomo IB, Nitzan O, Bonavina L,
Schaffer PM and Schaffer M: Clostridium difficile Infection:
Associations with chemotherapy, radiation therapy, and targeting
therapy treatments. Curr Med Chem. 23:4442–4449. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Su J, Li D, Chen Q, Li M, Su L, Luo T,
Liang D, Lai G, Shuai O, Jiao C, et al: Anti-breast cancer
enhancement of a polysaccharide from spore of ganoderma lucidum
with paclitaxel: Suppression on tumor metabolism with gut
microbiota reshaping. Front Microbiol. 9:30992018. View Article : Google Scholar
|
|
52
|
Stringer AM, Gibson RJ, Logan RM, Bowen
JM, Yeoh AS, Hamilton J and Keefe DM: Gastrointestinal microflora
and mucins may play a critical role in the development of
5-fluorouracil-induced gastrointestinal mucositis. Exp Biol Med
(Maywood). 234:430–441. 2009. View Article : Google Scholar
|
|
53
|
Yeung CY, Chiau JS, Cheng ML, Chan WT,
Chang SW, Chang YH, Jiang CB and Lee HC: Modulations of probiotics
on gut microbiota in a 5-fluorouracil-induced mouse model of
mucositis. J Gastroenterol Hepatol. 35:806–814. 2020. View Article : Google Scholar
|
|
54
|
Vanlancker E, Vanhoecke B, Smet R, Props R
and Van de Wiele T: 5-Fluorouracil sensitivity varies among oral
micro-organisms. J Med Microbiol. 65:775–783. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Saegusa Y, Ichikawa T, Iwai T, Goso Y,
Okayasu I, Ikezawa T, Shikama N, Saigenji K and Ishihara K: Changes
in the mucus barrier of the rat during 5-fluorouracil-induced
gastrointestinal mucositis. Scand J Gastroenterol. 43:59–65. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Ichim TE, Kesari S and Shafer K:
Protection from chemotherapy- and antibiotic-mediated dysbiosis of
the gut microbiota by a probiotic with digestive enzymes
supplement. Oncotarget. 9:30919–30935. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Nakayama H, Kinouchi T, Kataoka K, Akimoto
S, Matsuda Y and Ohnishi Y: Intestinal anaerobic bacteria hydrolyse
sorivudine, producing the high blood concentration of
5-(E)-(2-bromovinyl) uracil that increases the level and toxicity
of 5-fluorouracil. Pharmacogenetics. 7:35–43. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Yuan L, Zhang S, Li H, Yang F, Mushtaq N,
Ullah S, Shi Y, An C and Xu J: The influence of gut microbiota
dysbiosis to the efficacy of 5-Fluorouracil treatment on colorectal
cancer. Biomed Pharmacother. 108:184–193. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Bronckaers A, Balzarini J and Liekens S:
The cytostatic activity of pyrimidine nucleosides is strongly
modulated by Mycoplasma hyorhinis infection: Implications for
cancer therapy. Biochem Pharmacol. 76:188–197. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Yu T, Guo F, Yu Y, Sun T, Ma D, Han J,
Qian Y, Kryczek I, Sun D, Nagarsheth N, et al: Fusobacterium
nucleatum promotes chemoresistance to colorectal cancer by
modulating autophagy. Cell. 170:548–563.e516. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Zhang S, Yang Y, Weng W, Guo B, Cai G, Ma
Y and Cai S: Fusobacterium nucleatum promotes chemoresistance to
5-fluorouracil by upregulation of BIRC3 expression in colorectal
cancer. J Exp Clin Cancer Res. 38:142019. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
García-González AP, Ritter AD, Shrestha S,
Andersen EC, Yilmaz LS and Walhout AJM: Bacterial metabolism
affects the C. Elegans response to cancer chemotherapeutics. Cell.
169:431–441. 2017. View Article : Google Scholar :
|
|
63
|
Scott TA, Quintaneiro LM, Norvaisas P, Lui
PP, Wilson MP, Leung KY, Herrera-Dominguez L, Sudiwala S, Pessia A,
Clayton PT, et al: Host-microbe co-metabolism dictates cancer drug
efficacy in C. Elegans. Cell. 169:442–456.e418. 2017. View Article : Google Scholar
|
|
64
|
Fogelman D, Sugar EA, Oliver G, Shah N,
Klein A, Alewine C, Wang H, Javle M, Shroff R, Wolff RA, et al:
Family history as a marker of platinum sensitivity in pancreatic
adenocarcinoma. Cancer Chemother Pharmacol. 76:489–498. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Siddik ZH: Cisplatin: Mode of cytotoxic
action and molecular basis of resistance. Oncogene. 22:7265–7279.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Hato SV, Khong A, de Vries IJM and
Lesterhuis WJ: Molecular pathways: The immunogenic effects of
platinum-based chemotherapeutics. Clin Cancer Res. 20:2831–2837.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Pflug N, Kluth S, Vehreschild JJ, Bahlo J,
Tacke D, Biehl L, Eichhorst B, Fischer K, Cramer P, Fink AM, et al:
Efficacy of antineoplastic treatment is associated with the use of
antibiotics that modulate intestinal microbiota. Oncoimmunology.
5:e11503992016. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Shahid F, Farooqui Z and Khan F:
Cisplatin-induced gastrointestinal toxicity: An update on possible
mechanisms and on available gastroprotective strategies. Eur J
Pharmacol. 827:49–57. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Gui QF, Lu HF, Zhang CX, Xu ZR and Yang
YH: Well-balanced commensal microbiota contributes to anti-cancer
response in a lung cancer mouse model. Genet Mol Res. 14:5642–5651.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Wu CH, Ko JL, Liao JM, Huang SS, Lin MY,
Lee LH, Chang LY and Ou CC: D-methionine alleviates
cisplatin-induced mucositis by restoring the gut microbiota
structure and improving intestinal inflammation. Ther Adv Med
Oncol. 11:17588359188210212019. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Feng X, Cheng Q, Meng Q, Yang Y and Nie K:
Effects of ondansetron and [6]-gingerol on pica and gut microbiota
in rats treated with cisplatin. Drug Des Devel Ther. 13:2633–2641.
2019. View Article : Google Scholar :
|
|
72
|
Zhou P, Li Z, Xu D, Wang Y, Bai Q, Feng Y,
Su G, Chen P, Wang Y, Liu H, et al: Cepharanthine hydrochloride
improves cisplatin chemotherapy and enhances immunity by regulating
intestinal microbes in mice. Front Cell Infect Microbiol.
9:22510.3389. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Lee TH, Park D, Kim YJ, Lee I, Kim S, Oh
CT, Kim JY, Yang J and Jo SK: Lactobacillus salivarius BP121
prevents cisplatin-induced acute kidney injury by inhibition of
uremic toxins such as indoxyl sulfate and p-cresol sulfate via
alleviating dysbiosis. Int J Mol Med. 45:1130–1140. 2020.PubMed/NCBI
|
|
74
|
Lee YJ, Li KY, Wang PJ, Huang HW and Chen
MJ: Alleviating chronic kidney disease progression through
modulating the critical genus of gut microbiota in a
cisplatin-induced Lanyu pig model. J Food Drug Anal. 28:103–114.
2020. View Article : Google Scholar
|
|
75
|
Zhao L, Xing C, Sun W, Hou G, Yang G and
Yuan L: Lactobacillus supplementation prevents cisplatin-induced
cardiotoxicity possibly by inflammation inhibition. Cancer
Chemother Pharmacol. 82:999–1008. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Iida N, Dzutsev A, Stewart CA, Smith L,
Bouladoux N, Weingarten RA, Molina DA, Salcedo R, Back T, Cramer S,
et al: Commensal bacteria control cancer response to therapy by
modulating the tumor microenvironment. Science. 342:967–970. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
77
|
Ozben T: Oxidative stress and apoptosis:
Impact on cancer therapy. J Pharm Sci. 96:2181–2196. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Ewertz M, Qvortrup C and Eckhoff L:
Chemotherapy-induced peripheral neuropathy in patients treated with
taxanes and platinum derivatives. Acta Oncol. 54:587–591. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
79
|
Stojanovska V, Sakkal S and Nurgali K:
Platinum-based chemotherapy: Gastrointestinal immunomodulation and
enteric nervous system toxicity. Am J Physiol Gastrointest Liver
Physiol. 308:G223–G232. 2015. View Article : Google Scholar
|
|
80
|
Shen S, Lim G, You Z, Ding W, Huang P, Ran
C, Doheny J, Caravan J, Tate S, Hu K, et al: Gut microbiota is
critical for the induction of chemotherapy-induced pain. Nat
Neurosci. 20:1213–1216. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
81
|
Sprowl JA, Ciarimboli G, Lancaster CS,
Giovinazzo H, Gibson AA, Du G, Janke LJ, Cavaletti G, Shields AF
and Sparreboom A: Oxaliplatin-induced neurotoxicity is dependent on
the organic cation transporter OCT2. Proc Natl Acad Sci USA.
110:11199–11204. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
82
|
Forsgård RA, Marrachelli VG, Korpela K,
Frias R, Collado MC, Korpela R, Monleon D, Spillmann T and
Österlund P: Chemotherapy-induced gastrointestinal toxicity is
associated with changes in serum and urine metabolome and fecal
microbiota in male Sprague-Dawley rats. Cancer Chemother Pharmacol.
80:317–332. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
83
|
Chang CW, Liu CY, Lee HC, Huang YH, Li LH,
Chiau JS, Wang TE, Chu CH, Shih SC, Tsai TH and Chen YJ: Variety
probiotic preventively attenuates
5-fluorouracil/oxaliplatin-induced intestinal injury in a syngeneic
colorectal cancer model. Front Microbiol. 9:9832018. View Article : Google Scholar
|
|
84
|
Chang CW, Lee HC, Li LH, Chiau JS, Wang
TE, Chuang WH, Chen MJ, Wang HY, Shih SC, Liu CY, et al: Fecal
microbiota transplantation prevents intestinal injury, upregulation
of toll-like receptors, and 5-fluorouracil/oxaliplatin-induced
toxicity in colorectal cancer. Int J Mol Sci. 21:3862020.
View Article : Google Scholar :
|
|
85
|
Conroy T, Desseigne F, Ychou M, Bouché O,
Guimbaud R, Bécouarn Y, Adenis A, Raoul JL, Gourgou-Bourgade S, de
la Fouchardière C, et al: FOLFIRINOX versus gemcitabine for
metastatic pancreatic cancer. N Engl J Med. 364:1817–1825. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
86
|
Vanhoefer U, Harstrick A, Achterrath W,
Cao S, Seeber S and Rustum YM: Irinotecan in the treatment of
colorectal cancer: Clinical overview. J Clin Oncol. 19:1501–1518.
2001. View Article : Google Scholar : PubMed/NCBI
|
|
87
|
Sparreboom A, de Jonge MJ, de Bruijn P,
Brouwer E, Nooter K, Loos WJ, van Alphen RJ, Mathijssen RH, Stoter
G and Verweij J: Irinotecan (CPT-11) metabolism and disposition in
cancer patients. Clin Cancer Res. 4:2747–2754. 1998.PubMed/NCBI
|
|
88
|
Takasuna K, Hagiwara T, Hirohashi M, Kato
M, Nomura M, Nagai E, Yokoi T and Kamataki T: Involvement of
beta-glucuronidase in intestinal microflora in the intestinal
toxicity of the antitumor camptothecin derivative irinotecan
hydrochloride (CPT-11) in rats. Cancer Res. 56:3752–3757.
1996.PubMed/NCBI
|
|
89
|
Brandi G, Dabard J, Raibaud P, Battista
MD, Bridonneau C, Pisi AM, Labate AM, Pantaleo MA, Vivo AD and
Biasco G: Intestinal microflora and digestive toxicity of
irinotecan in mice. Clin Cancer Res. 12:1299–1307. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
90
|
Wardill HR, Gibson RJ, Van Sebille YZA,
Secombe KR, Coller JK, White IA, Manavis J, Hutchinson MR,
Staikopoulos V, Logan RM and Bowen JM: Irinotecan-induced
gastrointestinal dysfunction and pain are mediated by common
TLR4-dependent mechanisms. Mol Cancer Ther. 15:1376–1386. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
91
|
Pedroso SHSP, Vieira AT, Bastos RW,
Oliveira JS, Cartelle CT, Arantes RM, Soares PM, Generoso SV,
Cardoso VN, Teixeira MM, et al: Evaluation of mucositis induced by
irinotecan after microbial colonization in germ-free mice.
Microbiology. 161:1950–1960. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
92
|
Ribeiro RA, Wanderley CWS, Wong DVT, Mota
JM, Leite CA, Souza MH, Cunha FQ and Lima-Júnior RC: Irinotecan-
and 5-fluorouracil-induced intestinal mucositis: Insights into
pathogenesis and therapeutic perspectives. Cancer Chemother
Pharmacol. 78:881–893. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
93
|
Moore MJ, Goldstein D, Hamm J, Figer A,
Hecht JR, Gallinger S, Au HJ, Murawa P, Walde D, Wolff RA, et al:
Erlotinib plus gemcitabine compared with gemcitabine alone in
patients with advanced pancreatic cancer: A phase III trial of the
national cancer institute of Canada clinical trials group. J Clin
Oncol. 25:1960–1966. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
94
|
Forsgård RA, Marrachelli VG, Lindén J,
Frias R, Collado MC, Korpela R, Monleon D, Spillmann T and
Österlund P: Two-week aflibercept or erlotinib administration does
not induce changes in intestinal morphology in male sprague-dawley
rats but aflibercept affects serum and urine metabolic profiles.
Transl Oncol. 12:1122–1130. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
95
|
Heshiki Y, Vazquez-Uribe R, Li J, Ni Y,
Quainoo S, Imamovic L, Li J, Sørensen M, Chow BK, Weiss GJ, et al:
Predictable modulation of cancer treatment outcomes by the gut
microbiota. Microbiome. 8:282020. View Article : Google Scholar : PubMed/NCBI
|
|
96
|
Panebianco C, Andriulli A and Pazienza V:
Pharmacomicrobiomics: Exploiting the drug-microbiota interactions
in anticancer therapies. Microbiome. 6:922018. View Article : Google Scholar : PubMed/NCBI
|
|
97
|
Chiu CY and Miller SA: Clinical
metagenomics. Net Rev Genet. 20:341–355. 2019. View Article : Google Scholar
|
|
98
|
Chaput N, Lepage P, Coutzac C, Soularue E,
Roux KL, Monot C, Boselli L, Routier E, Cassard L, Collins M, et
al: Baseline gut microbiota predicts clinical response and colitis
in metastatic melanoma patients treated with ipilimumab. Ann Oncol.
28:1368–1379. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
99
|
Farowski F, Solbach P, Tsakmaklis A,
Brodesser S, Aguilar MR, Cornely OA, Dettmer K, Higgins PG,
Suerbaum S, Jazmati N, et al: Potential biomarkers to predict
outcome of faecal microbiota transfer for recurrent Clostridioides
difficile infection. Dig Liver Dis. 51:944–951. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
100
|
Khanna S, Montassier E, Schmidt B, Patel
R, Knights D, Pardi DS and Kashyap P: Gut microbiome predictors of
treatment response and recurrence in primary clostridium difficile
infection. Aliment Pharmacol Ther. 44:715–727. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
101
|
Aarnoutse R, de Vos-Geelen JMPGM, Penders
J, Boerma EG, Warmerdam FA, Goorts B, Damink SWM, Soons Z, Rensen
SS and Smidt ML: Study protocol on the role of intestinal
microbiota in colorectal cancer treatment: A pathway to
personalized medicine 2.0. Int J Colorectal Dis. 32:1077–1084.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
102
|
Stringer AM, Al-Dasooqi N, Bowen JM, Tan
TH, Radzuan M, Logan RM, Mayo B, Keefe DM and Gibson RJ: Biomarkers
of chemotherapy-induced diarrhoea: A clinical study of intestinal
microbiome alterations, inflammation and circulating matrix
metalloproteinases. Support Care Cancer. 21:1843–1852. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
103
|
Lin XB, Dieleman LA, Ketabi A, Bibova I,
Sawyer MB, Xue H, Field CJ, Baracos VE and Gänzle MG: Irinotecan
(CPT-11) chemotherapy alters intestinal microbiota in tumour
bearing rats. PLoS One. 7:e397642012. View Article : Google Scholar : PubMed/NCBI
|
|
104
|
Panebianco C, Adamberg K, Jaagura M,
Copetti M, Fontana A, Adamberg S, Kolk K, Vilu R, Andriulli A and
Pazienza V: Influence of gemcitabine chemotherapy on the microbiota
of pancreatic cancer xenografted mice. Cancer Chemother Pharmacol.
81:773–782. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
105
|
Meunier A, Nerich V, Fagnoni-Legat C,
Richard M, Mazel D, Adotevi O, Bertrand X and Hocquet D: Enhanced
emergence of antibiotic-resistant pathogenic bacteria after in
vitro induction with cancer chemotherapy drugs. J Antimicrob
Chemother. 74:1572–1577. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
106
|
Zwielehner J, Lassl C, Hippe B, Pointner
A, Switzeny OJ, Remely M, Kitzweger E, Ruckser R and Haslberger AG:
Changes in human fecal microbiota due to chemotherapy analyzed by
TaqMan-PCR, 454 sequencing and PCR-DGGE fingerprinting. PLoS One.
6:e286542011. View Article : Google Scholar : PubMed/NCBI
|
|
107
|
Montassier E, Gastinne T, Vangay P,
Al-Ghalith GA, des Varannes SB, Massart S, Moreau P, Potel G, de La
Cochetière MF, Batard E and Knights D: Chemotherapy- driven
dysbiosis in the intestinal microbiome. Aliment Pharmacol Ther.
42:515–528. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
108
|
Kong C, Gao R, Yan X, Huang L, He J, Li H,
You J and Qin H: Alterations in intestinal microbiota of colorectal
cancer patients receiving radical surgery combined with adjuvant
CapeOx therapy. Sci China Life Sci. 62:1178–1193. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
109
|
Lozupone CA, Stombaugh JI, Gordon JI,
Jansson JK and Knight R: Diversity, stability and resilience of the
human gut microbiota. Nature. 489:220–230. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
110
|
Bhatt AP, Pellock SJ, Biernat KA, Walton
WG, Wallace BD, Creekmore BC, Letertre MM, Swann JR, Wilson ID,
Roques JR, et al: Targeted inhibition of gut bacterial
β-glucuronidase activity enhances anticancer drug efficacy. Proc
Natl Acad Sci USA. 117:7374–7381. 2020. View Article : Google Scholar
|
|
111
|
Roberts AB, Wallace BD, Venkatesh MK, Mani
S and Redinbo MR: Molecular insights into microbial β-glucuronidase
inhibition to abrogate CPT-11 toxicity. Mol Pharmacol. 84:208–217.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
112
|
Pellock SJ, Walton WG, Biernat KA,
Torres-Rivera D, Creekmore BC, Xu Y, Liu J, Tripathy A, Stewart LJ
and Redinbo MR: Three structurally and functionally distinct
β-glucuronidases from the human gut microbe. J Biol Chem.
293:18559–18573. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
113
|
Wallace BD, Roberts AB, Pollet RM, Ingle
JD, Biernat KA, Pellock SJ, Venkatesh MK, Guthrie L, O'Neal SK,
Robinson SJ, et al: Structure and inhibition of microbiome
β-glucuronidases essential to the alleviation of cancer drug
toxicity. Chem Biol. 22:1238–1249. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
114
|
Wallace BD, Wang H, Lane KT, Scott JE,
Orans J, Koo JS, Venkatesh M, Jobin C, Yeh LA, Mani S and Redinbo
MR: Alleviating cancer drug toxicity by inhibiting a bacterial
enzyme. Science. 330:831–835. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
115
|
Chamseddine AN, Ducreux M, Armand JP,
Paoletti X, Satar T, Paci A and Mir O: Intestinal bacterial
β-glucuronidase as a possible predictive biomarker of
irinotecan-induced diarrhea severity. Pharmacol Ther. 199:1–15.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
116
|
Arumugam M, Raes J, Pelletier E, Paslier
DL, Yamada T, Mende DR, Fernandes GR, Tap J, Bruls T, Batto JM, et
al: Enterotypes of the human gut microbiome. Nature. 473:174–180.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
117
|
Dougan SK: The pancreatic cancer
microenvironment. Cancer J. 23:321–325. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
118
|
Binenbaum Y, Na'ara S and Gil Z:
Gemcitabine resistance in pancreatic ductal adenocarcinoma. Drug
Resist Updat. 23:55–68. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
119
|
Halbrook CJ, Pontious C, Kovalenko I,
Lapienyte L, Dreyer S, Lee HJ, Thurston G, Zhang Y, Lazarus J,
Sajjakulnukit P, et al: Macrophage-released pyrimidines inhibit
gemcitabine therapy in pancreatic cancer. Cell Metab. 29:1390–1399.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
120
|
Alexander JL, Wilson ID, Teare J, Marchesi
JR, Nicholson JK and Kinross JM: Gut microbiota modulation of
chemotherapy efficacy and toxicity. Nat Rev Gastroenterol Hepatol.
14:356–365. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
121
|
Villéger R, Lopès A, Carrier G, Veziant J,
Billard E, Barnich N, Gagnière J, Vazeille E and Bonnet M:
Intestinal microbiota: A novel target to improve anti-tumor
treatment? Int J Mol Sci. 20:45842019. View Article : Google Scholar :
|
|
122
|
McQuade JL, Daniel CR, Helmink BA and
Wargo JA: Modulating the microbiome to improve therapeutic response
in cancer. Lancet Oncol. 20:e77–e91. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
123
|
Becattini S, Taur Y and Pamer EG:
Antibiotic-induced changes in the intestinal microbiota and
disease. Trends Mol Med. 22:458–478. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
124
|
Flieger D, Klassert C, Hainke S, Keller R,
Kleinschmidt R and Fischbach W: Phase II clinical trial for
prevention of delayed diarrhea with cholestyramine/levofloxacin in
the second-line treatment with irinotecan biweekly in patients with
metastatic colorectal carcinoma. Oncology. 72:10–16. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
125
|
Kurita A, Kado S, Matsumoto T, Asakawa N,
Kaneda N, Kato I, Uchida K, Onoue M and Yokokura T: Streptomycin
alleviates irinotecan-induced delayed-onset diarrhea in rats by a
mechanism other than inhibition of β-glucuronidase activity in
intestinal lumen. Cancer Chemother Pharmacol. 67:201–213. 2011.
View Article : Google Scholar
|
|
126
|
Iida N, Mizukoshi E, Yamashita T,
Terashima T, Arai K, Seishima J and Kaneko S: Overuse of
antianaerobic drug is associated with poor postchemotherapy
prognosis of patients with hepatocellular carcinoma. Int J Cancer.
145:2701–2711. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
127
|
Levy SB and Marshall B: Antibacterial
resistance worldwide: Causes, challenges and responses. Nat Med.
10(12 Suppl): S122–S129. 2004. View
Article : Google Scholar : PubMed/NCBI
|
|
128
|
George Kerry R, Patra JK, Gouda S, Park Y,
Shin HS and Das G: Benefaction of probiotics for human health: A
review. J Food Drug Anal. 26:927–939. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
129
|
Yu AQ and Li L: The potential role of
probiotics in cancer prevention and treatment. Nutr Cancer.
68:535–544. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
130
|
Vivarelli S, Salemi R, Candido S, Falzone
L, Santagati M, Stefani S, Torino F, Banna GL, Tonini G and Libra
M: Gut microbiota and cancer: From pathogenesis to therapy. Cancers
(Basel). 11:382019. View Article : Google Scholar
|
|
131
|
An J and Ha EM: Combination therapy of
lactobacillus plantarum supernatant and 5-fluouracil increases
chemosensitivity in colorectal cancer cells. J Microbiol
Biotechnol. 26:1490–1503. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
132
|
Chang CW, Liu CY, Lee HC, Huang YH, Li LH,
Chiau JS, Wang TE, Chu CH, Shih SC, Tsai TH and Chen YJ:
Lactobacillus casei variety rhamnosus probiotic preventively
attenuates 5-fluorouracil/oxaliplatin-induced intestinal injury in
a syngeneic colorectal cancer model. Front Microbiol. 9:9832018.
View Article : Google Scholar :
|
|
133
|
Wang Y, Sun L, Chen S, Guo S, Yue T, Hou
Q, Feng M, Xu H, Liu Y, Wang P and Pan Y: The administration of
Escherichia coli Nissle 1917 ameliorates irinotecan-induced
intestinal barrier dysfunction and gut microbial dysbiosis in mice.
Life Sci. 231:1165292019. View Article : Google Scholar : PubMed/NCBI
|
|
134
|
Serkova MI, Urtenova MA, Tkachenko EI,
Avalueva EB, Orlov SV, Ivanov SV, Orishak EA and Skazyvaeva EV: On
the possibilities of correction of changes of the gastrointestinal
tract microbiota in patients with lung cancer treated receiving
chemotherapy. Eksp Klin Gastroenterol. 15–20. 2013.
|
|
135
|
Mego M, Koncekova R, Mikuskova E, Drgona
L, Ebringer L, Demitrovicova L, Nemova I, Trupl J, Mardiak J, Koza
I and Zajac V: Prevention of febrile neutropenia in cancer patients
by probiotic strain Enterococcus faecium M-74. Phase II study
Support Care Cancer. 14:285–290. 2006. View Article : Google Scholar
|
|
136
|
Picó-Monllor JA and Mingot-Ascencao JM:
Search and selection of probiotics that improve mucositis symptoms
in oncologic patients. A systematic review. Nutrients. 11:23222019.
View Article : Google Scholar :
|
|
137
|
González-Sarrías A, Tome-Carneiro J,
Bellesia A, Tomás-Barberán FA and Espin JC: The ellagic
acid-derived gut microbiota metabolite, urolithin A, potentiates
the anticancer effects of 5-fluorouracil chemotherapy on human
colon cancer cells. Food Funct. 6:1460–1469. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
138
|
Le Bastard Q, Ward T, Sidiropoulos D,
Hillmann BM, Chun CL, Sadowsky MJ, Knights D and Montassier E:
Fecal microbiota transplantation reverses antibiotic and
chemotherapy-induced gut dysbiosis in mice. Sci Rep. 8:62192018.
View Article : Google Scholar : PubMed/NCBI
|