Biogenesis and communication of
exosomes
Biogenesis and composition
Exosomes are nanometric vesicles (30–150 nm in
diameter) enclosed by a lipid bilayer membrane containing lipids,
proteins, DNA [mitochondrial DNA (mtDNA), single-stranded DNA
(ssDNA) and double stranded DNA (dsDNA)] and RNA [mRNAs, microRNAs
(miRNA/miRs), long non-coding RNAs (lncRNAs) and circular RNAs
(circRNAs)] (1). Exosomes can be
produced and secreted by almost all cell types, including
hematopoietic, neuronal, fibroblastic and cancer cells, and are
commonly found in physical body fluids. The small size and special
biofilm composition allow exosomes to cross main biological
barriers, and even the blood-brain barrier. Carrying a variety of
biomolecules, exosomes can mediate local and distant cellular
material transport and information transfer by delivering specific
‘cargoes’ (2,3).
Exosome formation begins when the plasma membrane
invaginates to generate early endosomes, and the endosome membrane
then further inverts to form late endosomes containing intraluminal
vesicles (ILVs), which are termed multivesicular bodies (MVBs).
During this process, signaling molecules derived from the parental
plasma membrane, cytoplasmic lysates, as well as common proteins
which play a role in membrane fusion and cytoskeletal regulation
(Rab family, Alix, GTPase, endosomal sorting complexes required for
transport, etc.) are doped into the vesicles, and when the MVBs
fuse with the plasma membrane, the ILVs can be released from the
parental cell into the extracellular environment as exosomes by
cytosolic ejection (4) (Fig. 1).
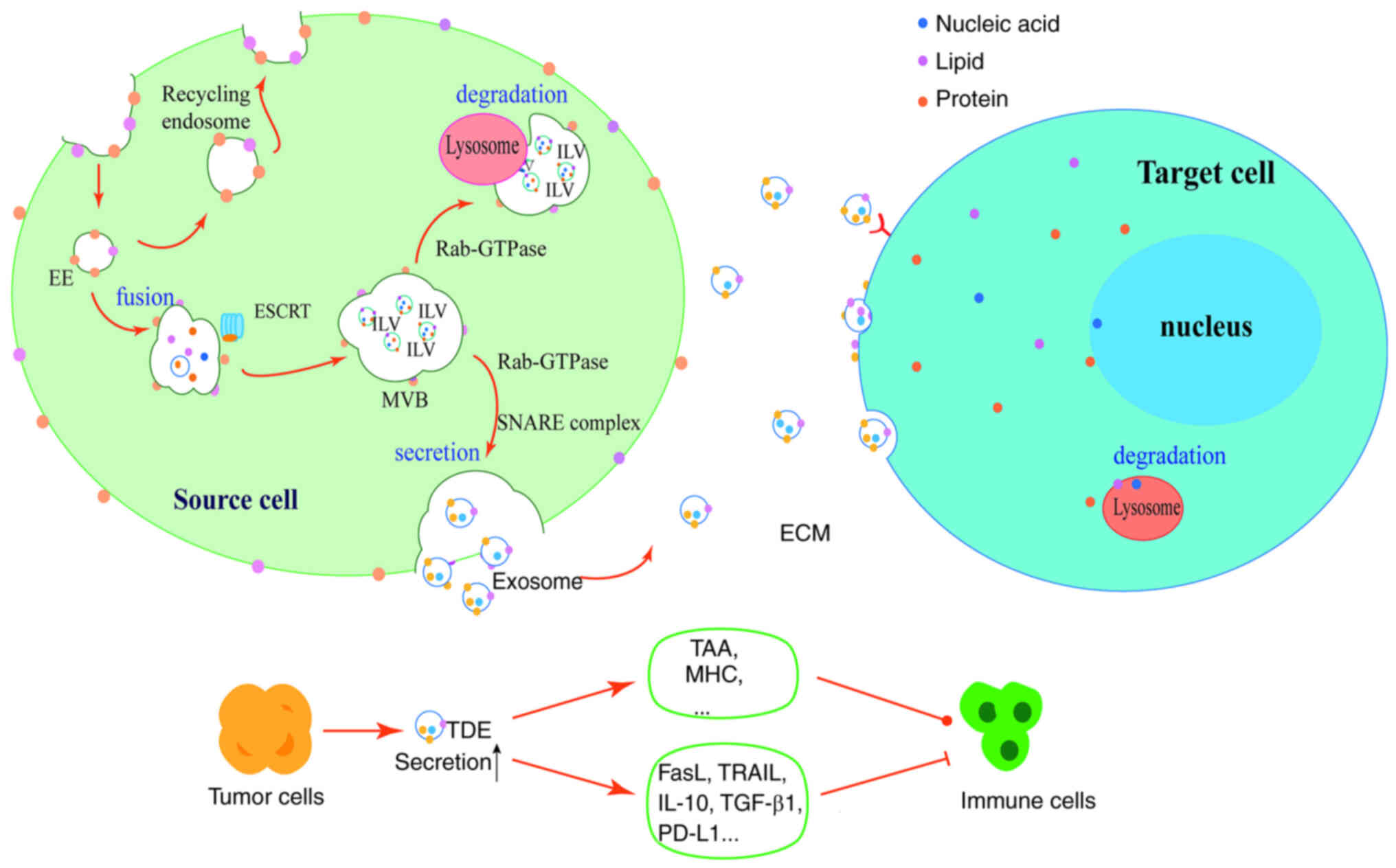 | Figure 1.Biogenesis of exosomes and their
communications with cells. The production of exosomes begins with
the invagination of the cell plasma membrane to form early
endosomes, which fuse. As the plasma membrane continues to
invaginate and closes to form numerous ILVs, the endosomes are
transformed into MVBs containing numerous ILVs, each containing
proteins, lipids and nucleic acids, with the assistance of the
endosomal sorting complex required for translocation. Under the
regulation of Rab-GTPase, MVBs can fuse with lysosomes for
degradation, or release ILVs extracellularly to form exosomes via
cytosolic exocytosis under the action of the soluble
N-ethylmaleimide-sensitive factor attachment protein receptor
complex. Exosomes can bind to receptor cells via ligand-receptor
form, or fuse directly with target cell membranes, or interact with
target cells via endocytosis. Tumor cells can produce more exosomes
and carry tumor-associated antigens, major histocompatibility
complex molecules or immunosuppressive proteins that activate or
inhibit the immune cells. ILVs, intraluminal vesicles; MVBs,
multivesicular bodies; EE, early endosomes; ESCRT, endosomal
sorting complex required for translocation; ECM, extracellular
matrix; TAA, tumor-associated antigen; MHC, major
histocompatibility complex; TRAIL, TNF-related apoptosis-inducing
ligand; PD-L1, programmed death-ligand 1; TDE, tumor-derived
exosome. |
The endosomal sorting complex required for
translocation (ESCRT) is generally considered to be the driving
factor for MVB formation (5).
ESCRT is composed of five soluble protein complexes [ESCRT-0,
ESCRT-I, ESCRT-II, ESCRT-III and auxiliary proteins (Vps4, Vta1 and
Alix)]. First, the interaction between the ESCRT-0 subunits and
endosomal-enriched phosphatidylinositol 3-phosphate [PtdIns(3)P]
enriches ESCRT-0 toward the endosome. The ESCRT-0 complex then
recruits ESCRT-I subunits to recognize and aggregate ubiquitinated
‘cargo’ on endosome-restricted membrane microdomains. The ESCRT-II
subunits then provide a platform that initiates the stepwise
assembly of the ESCRT-III complex by coordinating the multiple
interactions of ESCRT-I, PtdIns(3)P and ubiquitin to transfer and
wrap the ubiquitinated ‘cargo’, resulting in the formation of
‘cargo’-loaded MVBs (6). Alix is
an auxiliary protein that binds with ESCRT-III subunits and
contributes to the budding and shedding process of ILV generation,
which plays a prominent role in exosome formation (3). MVBs can also be generated in an
ESCRT-independent manner; however, they require the involvement of
neutral sphingomyelinase-dependent ceramide, cholesterol, or
tetraspanins (e.g., CD9, CD63 and CD81) (7).
MVB fusion with the plasma membrane is mediated by
the soluble N-ethylmaleimide-sensitive factor attachment protein
receptor (SNARE), which localizes in or is recruited into the
docking MVBs and the plasma membrane, and then initiates membrane
fusion by forming SNARE complexes, where vesicles within the MVB
are released to form exosomes outside the cell. In addition, MVBs
are able to fuse with lysosomes and then degrade. The Rab family of
GTPases are essential for the regulation of the secretion and
degradation of MVBs (Fig. 1).
Rab27A and Rab27B are necessary for vesicle transport and fusion
during exosome secretion (8);
Rab7 is present on late endosomes and lysosomes, and functions by
translocating MVBs to lysosomes for degradation (9), whereas active Rab31 can recruit TBC1
structural domain family member 2B to inactivate Rab7, preventing
the fusion of MVBs with lysosomes and promoting the secretion of
exosomes. Other members of the Rab family, including Rab11, Rab35
and Rab27, are also critical for exosome secretion by regulating
the transport of MVBs to the plasma membrane (6,10).
Exosomes are loaded with various proteins, including
chaperone proteins, heat shock proteins (HSPs), cytoskeletal
proteins (i.e., actin and microtubulin), fusion proteins,
mitochondrial proteins and tetraspanins, etc. HSPs and cytoskeletal
proteins are present in almost all types of exosomes, while some
proteins are cell-specific, such as major histocompatibility
complex (MHC) II and MHC I, transferrin receptor and CD3 (11). Exosomes also contain nucleic acid
molecules, such as mRNAs and lncRNAs, and exosome RNAs delivered to
target cells can alter the epigenetic characteristics of cells and
thereby affect their functionality (12).
Although exosomes can be produced by almost all
cells, there is evidence to indicate that tumor cells produce
significantly more exosomes than other cells, and the level of
exosomes in the blood of cancer patients is always higher than that
in the blood of healthy individuals (13). It has been proposed that the
prevalent hypoxic conditions in the tumor microenvironment (TME)
may be responsible for the high secretion of exosomes from tumor
cells. Hypoxia may affect key steps of exosome release, such as
cargo sorting, MVBs transport and membrane fusion (14). In ovarian cancer cells, hypoxia
can stimulate exosome release via the upregulation of Rab27A and
the downregulation of Rab7 (15).
In addition, the expression of certain four-transmembrane proteins
involved in MVB formation, such as CD63 and CD9, is also
upregulated due to hypoxia (16).
The expression of SNAP-25, a SNARE protein participates in the
fusion of MVB with plasma membrane, is also increased under hypoxic
conditions (17). However, the
exact mechanisms through which hypoxia leads to greater exosome
release from tumor cells remain unclear (14).
Tumor-derived exosomes (TDEs) are known to transport
nucleic acids, proteins and metabolites from the maternal tumor
cells to other sites to act on recipient cells, and play a role in
tumor development and immune regulation (18). Nucleic acids, membrane surface
material and other cellular contents derived from maternal tumor
cells confer specific molecular characteristics on exosomes, thus
leading to the distinction between TDEs and exosomes of
non-malignant cell origin, and also between exosomes of different
tumor cell origins. As tumor cells are more likely to express and
activate a variety of immunosuppressive molecules to block
antitumor immune responses, TDEs are often rich in
immunosuppressive proteins, including FasL, TNF-related
apoptosis-inducing ligand (TRAIL), programmed death-ligand 1
(PD-L1), IL-10, TGF-β1, etc. (4).
On the other hand, TDEs also carry tumor-associated antigens
(TAAs), MHC molecules and co-stimulatory molecules, enabling them
to activate immune system and promote anti-tumor responses
(19). Such a complex molecular
profile of TDEs confers the dual ability on them to mediate either
immunosuppression or immunostimulation under various
conditions.
Cell-to-cell communications via
exosomes
First discovered in 1981, exosomes were initially
considered to be a cellular waste product (20). However, it was later demonstrated
that exosomes released extracellularly can be transferred to
recipient cells to perform various functions in recipient cells
through their membrane molecules and loaded substances, effectively
altering the biological responses of recipient cells and therefore
enabling intercellular communication (21).
Typically, exosomes can interact with target cells
through one or more of the following mechanisms: i) The activation
of intracellular signaling pathways by signaling through surface
receptor-ligand interactions; ii) fusion with the target cell
plasma membrane, and incorporation of its membrane contents into
the plasma membrane or the release of cytoplasmic contents into the
target cell; iii) the internalization of exosomes by lipid rafts,
lattice-protein-dependent giant cell ‘drinking’ action or
phagocytosis; iv) receptor-mediated endocytosis. Exosomes enter
target cells by phagocytosis or endocytosis, and the ‘cargo’
carried within them can be incorporated directly into the cellular
machinery to initiate reprogramming, or directed to the lysosome
for degradation and removal (22)
(Fig. 1).
Studies have demonstrated that different integrins
expressed on exosomes determine their specific cell fusion or
adhesion to extracellular matrix molecules in selected organs.
However, the main components that dictate the targeting specificity
of exosomes are currently unknown and warrant further investigation
(1,23).
Effects of tumor-derived exosomes on the
tumor microenvironment
The TME consists of peritumor blood vessels,
extracellular matrix and surrounding cells including fibroblasts,
adipocytes, immune cells, etc., and plays a key role in the
occurrence and progression of tumors (24). TDEs have been shown to be released
autonomously by tumor cells and affect the TME via a variety of
processes, such as promoting cellular epithelial mesenchymal
transition, inducing angiogenesis and inducing various functional
changes in immune cells, thus mediating tumor metastasis to
proximal or distant tissues and organs (25) (Fig.
2).
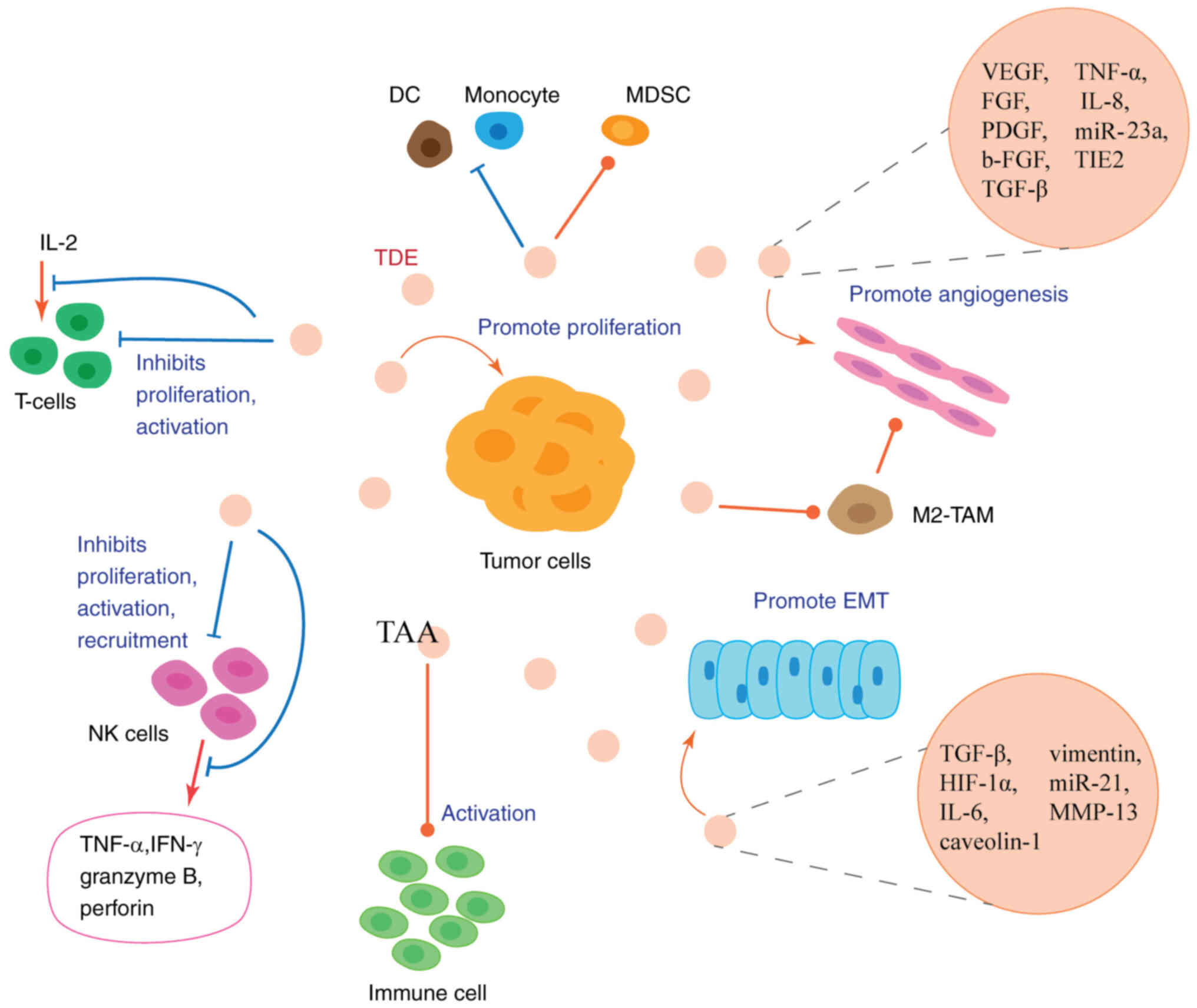 | Figure 2.TDEs in the tumor microenvironment.
TDEs can be released autonomously by tumor cells and carry
EMT-promoting factors and pro-angiogenic factors to promote EMT and
angiogenesis. TDEs can also promote the production of M2 phenotype
tumor-associated macrophages, and M2-type tumor-associated
macrophages secrete pro-angiogenic factors and cytokines to promote
angiogenesis. TDEs can carry tumor-associated antigens to activate
immune cell functions; however, they mainly exert inhibitory
functions on immune cells, including the functions of natural
killer cells, T-cells, dendritic cells and monocytes. Ultimately,
TDEs can suppress antitumor immunity and induce a microenvironment
which promotes tumor proliferation, activation and metastasis.
TDEs, tumor-derived exosomes; EMT, epithelial-mesenchymal
transition; NK, natural killer; DC, dendritic cell; MDSC,
myeloid-derived suppressor cell; TAA, tumor-associated antigen. |
TDEs promote epithelial-mesenchymal
transition (EMT)
Malignant tumors are often aggressive, and tumor
epithelial cells need to undergo the EMT process in order to
acquire the ability to migrate. Study has confirmed that TDEs are
able to enhance the migratory capacity of tumor cells by promoting
the conversion of tumor epithelial cells to a mesenchymal
phenotype. TGF-β, HIF-1α, IL-6, caveolin-1, vimentin and miRNAs
have all been proven to be EMT-promoting factors carried by TDEs
(10) (Fig. 2).
EMT usually occurs in the initial stages of tumor
metastasis, where tumor cells become invasive by reducing the
expression of E-cadherin, the loss of cell polarity, and the
increased expression of N-cadherin, imentin and Twist (10). It has been demonstrated that TDEs
can transfer EMT-associated RNAs and proteins, such as miR-21 and
MMP-13 to recipient cells, mediating calmodulin instability and
subsequent EMT. In turn, the exosomal miRNA profile produced
following cellular EMT may be altered, which further promotes EMT
and tumor cell migration and invasion (26,27).
TGF-β plays a critical role in the maintenance of
the tumor stroma and the induction of EMT (28). TGF-β triggers signaling through
TGF-βRI and TGF-βRII. Upon binding to TGF-β, TGF-βRII functions as
a high-affinity TGF-β receptor to recruit and phosphorylate
TGF-βRI, which subsequently activates downstream SMAD proteins,
SMAD2 and SMAD3 phosphorylation, and forms a complex with SMAD4
that transfers to the nucleus to bind transcription factors and
chromatin proteins to regulate gene transcription (29,30). TDEs have been shown to increase
the expression of TGF-βRI and TGF-βRII in recipient cells, and to
promote the TGF-β-induced transcription of EMT-related
transcription factors (29). For
example, an in vitro study using cell lines verified that
breast cancer-derived exosomes activated TGF-β receptor-mediated
signaling pathways in mesenchymal stem cells and promoted their
differentiation into fibroblasts, which is a key constituent of the
tumor stroma (31).
TDEs promote angiogenesis
Tumor growth is dependent on the construction of the
host vascular network, and for this reason, tumor cells can induce
a pro-angiogenic environment to build a vascular network, and
promote tumor growth and spread. TDEs are one of the main
mechanisms through which tumor cells induce angiogenesis. VEGF,
FGF, PDGF, b-FGF, TGF-β, TNF-α and IL-8 are the key angiogenic
stimulating factors carried by TDEs, and TDEs can effectively
induce angiogenesis through the transfer and release of these
factors (10) (Fig. 2).
It has been reported that exosomal miR-23a derived
from lung cancer cells directly inhibits its targets, prolyl
hydroxylase (PHD)1 and PHD2, leading to the accumulation of HIF-1α
in endothelial cells, thereby enhancing angiogenesis. miR-23a also
increases vascular permeability and promotes the migration of tumor
cells to the epithelium by inhibiting the tight junction protein,
Zonula occludens-1 (32). There
is also evidence to indicate that lncRNA-carrying TDEs released
from hepatocellular carcinoma (HCC) cells enhance the expression of
VEGFR1 in endothelial cells, thereby promoting angiogenesis
(33).
Endothelial-specific receptor tyrosine kinase
(TIE2), a receptor for angiopoietin, is mainly expressed in
endothelial cells and regulates vascular development, thus playing
a key role in vascular remodeling for tumor angiogenesis (34). In vitro and in vivo
analyses have demonstrated that TDEs can transfer high levels of
TIE2 produced by tumor cells to tumor-associated macrophages (TAMs)
and promote angiogenesis by facilitating the conversion of TAMs to
TIE2-expressing macrophages (TEMs), which possess a pro-angiogenic
phenotype. Exosomes carrying TIE2 can also enter the peripheral
blood circulation, induce the conversion of monocytes to TEMs, and
eventually interact with endothelial cells to promote angiogenesis
(35).
TDEs promote the proliferation and
metastasis of tumor cells
The ‘cargo’ loaded in TDEs can function as critical
regulators to induce signal transduction and gene expression, and
thus establish the tumor metastastic niche to promote tumor cell
proliferation and migration.
Toll-like receptors (TLRs) are a main class of
proteins involved in non-specific immunity, with the ability to
monitor and recognize a variety of disease-related molecular
patterns. Research using mouse models has demonstrated that TLR3 in
lung epithelial cells can be activated by TDEs-RNA, leading to the
production of chemokines and the recruitment of neutrophils to the
lungs, which induces the formation of the pre-metastatic niche in
lung tumors by suppressing antitumor immunity (36). Another study demonstrated that
exosomal miRNAs secreted by mouse lung cancer cells could be
transferred to macrophages and could bind to TLR7/8, resulting in
NF-κB activation and the secretion of the pro-metastatic
inflammatory factor, TNF-α (37).
Except nucleic acids, proteins such as HSP72 on the surface of
TDEs, can also activate NF-κB signaling and induce IL-6 and TNF-α
production in myeloid suppressor cells in a TLR2/MYD88-dependent
manner. These results suggest that TDEs play a crucial role in the
induction of tumor pre-metastatic niches, and that the TLR
signaling pathway may be an essential mediator of the action of
TDEs by mediating NF-κB activation and pro-inflammatory factor
production (38).
TDEs can also transfer oncogenic mutated genes to
recipient cells. For example, colon cancer cells expressing mutant
K-ras can promote tumor growth and metastasis by transferring the
oncogene via exosomes to adjacent normal cells (28).
Activation of immune cells by
TDEs
As TDEs carry a number of TAAs and can metastasize
to almost any area, they can be uptaken by antigen-presenting cells
and can active an effective antitumor immune response in the body.
Moreover, exosomes in the TME can also be produced by activated
immune cells, including dendritic cells (DCs), natural killer (NK)
cells and etc., and can subsequently stimulate the antitumor immune
response of the immune cells (4,39).
In a previous study on non-small cell lung cancer
(NSCLC), exosomes derived from Rab27a-overexpressing tumor cells
were found to significantly promote DC maturation by upregulating
the MHC I, CD80 and CD86 levels, and enhancing the proliferation
and immune response of CD4+ T-cells (40). It has been demonstrated that
radiation therapy at certain doses leads to the accumulation of
cytoplasmic dsDNA in tumor cells; TDEs produced by irradiated mouse
breast cancer cells can deliver dsDNA to DCs, and DCs can then
sense the DNA via cyclic GMP-AMP synthase, stimulate IFN-β
production and induce the expression of several
interferon-stimulated genes, ultimately leading to DC activation
and the antitumor T-cell response (41). There is evidence to indicate that
exosomes isolated from patients with non-metastatic primary
melanoma have the ability to inhibit tumor lung metastasis.
Mechanistically, ‘non-metastatic’ TDEs can stimulate an intrinsic
immune response through the expansion of Ly6Clow patrolling
monocytes in bone marrow, and that melanoma cells in pre-metastatic
ecological sites are then cleared through NK cell recruitment and
TRAIL-dependent macrophage killing (42).
Suppression of immune cells by
TDEs
Although TDEs have a certain activating effect on
immune cells, they also have an inordinate inhibitory effect, and
the majority of the current studies focus on the inhibitory effects
of TDEs on immune cells. As tumors grow and develop, tumor cells
express a large number of immunosuppressive molecules in order to
escape from immune surveillance. They also produce numerous
exosomes to carry immunosuppressive ‘cargo’, which can suppress the
body's antitumor immunity by inhibiting the activation of NK cells,
suppressing the proliferation and immune response of T-cells,
promoting the differentiation of bone marrow progenitor cells and
maintaining T-lymphocytes in a quiescent state (43). Since exosomes exist in all types
of human bodily fluids, exosomes not only affect immune cells in
the TME, but also affect immune cells in the systematic circulation
and a variety of lymphoid organs (4).
NK cells
As NK cells do not require prior exposure to
antigens to recognize tumor cells, they are considered to be the
first responders to the malignant transformation of cells (44). It has been suggested that TDEs can
deliver their own loaded ‘cargo’ to NK cells, thus blocking their
antitumor effect (45).
A previous study demonstrated that exosomes isolated
from patients with acute myeloid leukemia significantly decreased
the migration of NK-92 cells towards tumor cells (46). In a pioneering study, it was also
found that pre-treatment with exosomes derived from TS/A cells, a
murine breast cancer cell line, significantly reduced the number of
NK cells in vitro, which indicated that TS/A-exosomes
inhibited the proliferation and survival of NK cells (47). Moreover, TDEs can also block NK
cell activation. NKG2D and NKp30 are both critical activation
receptors on NK cells, and the expression levels of activation
receptors determine the antitumor capacity of NK cells (48). TDEs can carry ligands of NKG2D and
high levels of membrane-associated TGF-β, which reduces the surface
expression of key activation receptors (NKG2D) on NK cells, thus
profoundly diminishing the activation of NK cells (48,49).
Apart from reducing the number of NK cells and
interfering with NK cell recruitment and activation, TDEs have also
been demonstrated to suppress activated NK cells by inhibiting the
production of TNF-α and IFN-γ, the two main cytokines involved in
the cytotoxic effects of NK cells (24). In addition, the cytotoxic effects
of activated NK cells are also largely dependent on the release of
perforin and granzyme B, and it has been shown that TDEs can
significantly reduce the expression of perforin and granzyme B at
the protein level in a concentration-dependent manner (50). The aforementioned effects of TDEs
result in the attenuation of the toxic effects of NK cells from
various aspects.
Macrophages
Exosomes have been proven to alter the phenotype of
macrophages. They can convert the macrophage phenotype to either
the M1- or M2-type. M1 macrophages are in a pro-inflammatory
phenotype and increase the secretion of pro-inflammatory cytokines
and chemotactic molecules, inducing immune stimulation and the
effective clearance of pathogens and infections; M2 macrophages are
anti-inflammatory and play a central role in stimulating
angiogenesis, as well as promoting tumor progression (1).
Exosome-activated macrophages are considered to play
a key role in tumor progression, as macrophages activate paracrine
signaling pathways to promote tumor growth, angiogenesis and
invasion, while blocking inflammation and immune remodeling. In
co-culture studies, tumor cells have been shown to secrete
chemokines, such as CCL2 and CSF1, which recruit monocytes from the
peripheral circulating blood to the TME and induce them to
differentiate into TAMs, which exhibit the M2 phenotype, and can
secrete pro-angiogenic factors and cytokines that promote
angiogenesis, tumor growth and metastasis (51,52).
It has also been proposed that HCC cell-derived
exosomes containing high levels of lncRNA TUC339 can regulate the
development of neighboring macrophages toward the M2 phenotype,
ultimately creating an immunosuppressive environment conducive to
tumor proliferation and progression (53). Under tumor characteristic hypoxic
conditions, lactate-rich exosomes secreted by prostate cancer cells
can induce VEGF and Arginase 1 (Arg1) expression in macrophages via
HIF1α. VEGF mediates tumor neovascularization; Arg1 is involved in
regulating cell proliferation and promoting tumor growth. In
addition, an increased Arg1 expression leads to a diminished
cytotoxic response of macrophages to growing tumor cells (54).
While M1 macrophages are pro-inflammatory and
anti-tumor phenotypes, TDEs have also been shown to have the
potential to polarize macrophages to the M1 phenotype. Therefore,
promoting the conversion of macrophages to the M1 phenotype through
exosomes may be a developable direction for tumor immunotherapy
(52).
T-lymphocytes
T lymphocytes play a central role in the immune
response. CD8+ T-cells are cytotoxic and can directly
kill pathogen-infected cells or cancer cells, while CD4+
T-cells mainly play regulatory and paracrine roles. It has been
demonstrated that TDEs can induce immunosuppression by promoting
the apoptosis of CD8+ T-cells and enhancing the
suppressive activity of CD4+ regulatory T-cells, thereby
facilitating tumor immune escape (55).
TDEs carry a variety of surface ligands which can be
delivered to T-cell surface receptors to regulate T-cell gene
expression and function. It has also been shown that exosomes from
tumor cells carry higher levels of PD-L1, which can directly
interact with PD-1 on the surface of CD8+ T-cells, and inhibit
their proliferation and activation, thereby promoting tumor immune
escape (56), and that TDEs can
transfer PD-L1 to PD-L1-negative tumor cells and confer them the
ability to inhibit T-cell function (57). In addition, TDEs also induce
immunosuppressive monocytes, thus indirectly inhibiting T-cell
proliferation and activation (58). Since exosomes can be released into
bodily fluids, TDEs carrying PD-L1 can also enter lymph nodes with
lymphatic drainage, as well as other remote tissues and organs,
such as the spleen, via the blood circulation to inhibit the
proliferation and function of local T-cells (59).
Additionally, exosomes can be internalized into
cells and release proteins, miRNAs and other inclusions that
interfere with T-cell function, as demonstrated by previous
studies: Exosomes derived from several head and neck cancers have
been found to be loaded with an immunomodulatory protein
galectin-1, inducing CD8+ T-cells to differentiate
towards a suppressive phenotype (60); TDEs deliver high levels of TGF-β
secreted by tumor cells to T-cells to inhibit their proliferation
and differentiation, and TGF-β1-rich exosomes also exert
immunosuppressive effects by suppressing lymphocyte responses to
IL-2 (28); in addition, it has
been reported that TDEs can function as long-distance transport
carriers of Arg1 and deliver it to peripheral T-cells to inhibit
their proliferation, thus attenuating the body's antitumor immune
response (61).
Furthermore, TDEs have been shown to carry
CD39/CD73, the most critical extracellular enzyme for adenosine
(ADO) production, contributing to the upregulation of ADO in
recipient cells (62).
Subsequently, ADO inhibits the function and proliferation of both
CD4+ T-helper cells and CD8+ cytotoxic
T-cells via G protein-coupled receptors, facilitating tumor cell
escape from the host immune system (63).
Other immune cells
DCs are antigen-presenting cells that function as
messengers between intrinsic and adaptive immunity. Exosomes from
breast cancer or Lewis lung cancer cells have been shown to inhibit
bone marrow progenitor cell differentiation into DC and induce
apoptosis (64). TDEs also
transfer miRNAs to DCs, leading to the downregulation of TLR4
expression, and subsequently resulting in the decreased production
of cytokines involved in the immune response in DCs (1). In addition, TDEs can block the
migration of DCs to secondary lymphoid organs by inhibiting the
expression of chemokine receptors, such as CCR6, CCR7 and CXCR3
(65).
Monocytes are a critical part of the body's defense
system and can differentiate into macrophages and DCs in the
peripheral area, while TDEs suppress the immune response by
affecting the maturation and differentiation of monocytes (66). For example, TDEs have been
reported to inhibit the differentiation of monocyte precursor cells
into DCs or promote the monocytes to differentiate into
TGF-β-expressing DCs interfering with T-lymphocyte proliferation;
TDEs can also drive monocytes to differentiate towards a pro-tumor
phenotype that releases tumor-supporting cytokines and leads them
to express the immunosuppressive molecule, PD-L1 (1).
Myeloid-derived suppressor cells (MDSCs) are
immature bone marrow cells with immunosuppressive activity that
produce numerous immunosuppressive factors and suppress antitumor
immune responses; TDEs have been shown to play a critical role in
MDSC expansion and activation (24). A previous in vitro study
demonstrated that miR-10a and miR-21 expression in exosomes
produced by glioblastoma enhanced MDSC expansion and
immunosuppressive factor levels (67). TDEs can also induce the conversion
of bone marrow cells into MDSCs through the delivery of
prostaglandin E2 (PGE2) and TGF-β; the blocking of exosomal PGE2
and TGF-β can eliminate the induction of MDSCs and diminish the
MDSC-mediated immunosuppressive effects (68).
Effect of exosomes on the TME during
microbial infections
An increasing number of studies have demonstrated
that exosomes can not only be produced by a variety of mammalian
cells, but are also found in bacteria, viruses and fungi (69). Since the infection of certain
microorganisms is closely related to tumor development, exosomes
may also have a potential effect on their interaction.
Helicobacter pylori (H. pylori)
infection
Studies have proposed that Gram-negative bacteria
can release ‘cargo’-loaded exosomes that can spread infection by
activating the transcription of genes which promote bacterial
infection (70). Exosomes
produced by infected gastric epithelial cells are capable of
transferring virulence factors to modulate host immune responses
and assist bacterial invasion (71). H. pylori is a Gram-negative
bacterium that colonizes the gastric epithelium and damages the
gastric mucosa; it has a high infection rate and is strongly
associated with various gastric mucosal diseases, including gastric
cancer (GC) (72). Previous
research has revealed the impact of H. pylori infection on
the treatment outcomes of tumors in sites beyond the
gastrointestinal tract (73).
However, due to the particular survival and growth environment of
H. pylori and the gastric tissue barrier, H. pylori
itself does not enter the blood circulation or reach other tissues
and organs (72); presumably
there may be an exosomal influence during this action.
A previous study demonstrated that the protein
levels of mesenchymal-epithelial transition (MET) factors were
significantly decreased in GC cells following H. pylori
infection, while their mRNA levels remained unchanged. However, the
phosphorylated active form of MET factors was increased in exosomes
released from H. pylori-infected GC cells, which were
delivered to tumor-infiltrating macrophages and internalized. This
induced macrophages to convert into a pro-tumorigenic phenotype and
stimulating the expression of the pro-inflammatory factor, IL-1β,
and the activation of its downstream signaling pathways, thereby
promoting tumor growth and progression (74). Cytotoxin-associated gene A (cagA)
positive strains of H. pylori is one of the most virulent
strains. CagA-positive H. pylori-infected GC cells secrete
CagA-containing exosomes that induce morphological changes in
gastric epithelial cells and GC cells, and transmit functional CagA
into cells that may be involved in the development of extragastric
diseases associated with CagA-positive H. pylori infection
(2).
Fusobacterium nucleatum (Fn)
infection
Infection of Fn, a specialized anaerobic bacterium,
is the most important microbial-related risk factor in the
development of colorectal cancer (CRC) (75). Fn chronic infection involved in
the development of CRC is achieved by interacting with immune cells
and stromal cells by the surface molecules of Fn and then inducing
immune escape and immunosuppression (76). Fn infection has been shown to
increase the secretion of exosomes from CRC cells, and that
proteins and miRNAs involved in cell migration are enriched in the
secreted exosomes, which are delivered from Fn-infected CRC cells
to uninfected cells to increase their migration capacity and
promote tumor metastasis. The levels of miRNAs and proteins in
circulating exosomes associated with CRC cell migration are also
closely related to Fn abundance and the tumor stage of patients
with CRC (77).
Hepatitis B virus (HBV) infection
HBV is considered an important pathogenic factor in
the development of HCC. HBV-infected hepatocytes secrete exosomes
(HBx) containing numerous HBV DNA, RNA and protein components, that
can be delivered to uninfected hepatocytes to regulate cell
function and phenotype, thereby shaping a microenvironment
conducive to viral replication (78). HBx contain significantly higher
levels of immunosuppressive miRNAs, and these miRNAs can inhibit
the body's antiviral immune response by downregulating IL-12
expression to inhibit NK cell activity (79). In addition, exosomes with HBV
replication secreted from HBV-induced HepAD38 cells can
significantly upregulate PD-L1 expression in monocytes, which can
bind to PD-1 on T-cells to induce T-cell failure, thereby
inhibiting T-cell function to promote HBV infection (80). TDEs from HCC can also transport
interferon induced transmembrane protein 2 to DCs, thereby
inhibiting IFN-α secretion and blocking the antiviral effects
(81). All these effects of
exosomes contribute to the development of HBV infection and
HCC.
Impact of exosomes on immunotherapy
Since the suppressive effects of TDEs on the body's
immunity far exceeds the activating effect, and immunotherapy
mostly promotes antitumor immunity by regulating the body's immune
system, the presence of TDEs inevitably has several adverse effects
on immunotherapy. TDEs carry TAAs, which effectively bind to
tumor-reactive antibodies used in immunotherapy, and significantly
inhibit the binding of these antibodies to tumor cells, as well as
the antibody-dependent cell-mediated cytotoxic effect, leading to
the reduced effectiveness of tumor immunotherapy drugs (66).
Anti-PD-1/PD-L1 immunotherapy is currently one of
the most commonly used types of immunotherapy in the clinical
practice; however, its effectiveness remains limited. Exosomal
PD-L1 from tumor cells is considered to play a key role in
mediating resistance to anti-PD-1/PD-L1 immunotherapy. A possible
mechanism involved in drug resistance has been suggested to be the
high expression level of exosomal PD-L1 and its competitive binding
to anti-PD-L1 antibodies results in the failure to inhibit PD-L1
function on the surface of tumor cells (82). It has also been shown that miR-21
overexpression is associated with acquired resistance to epidermal
growth factor receptor-tyrosine kinase inhibitors (EGFR-TKI) in
NSCLC. In EGFR-TKI-resistant NSCLC cells, miR-21 is overexpressed
and exosomes can transfer miR-21 into EGFR-TKI-sensitive NSCLC
cells, reducing the sensitivity of cancer cells to EGFR-TKI, and
thus inducing drug resistance (83,84).
Cancer stem cells (CSCs) are a small subpopulation
of malignant tumor cells with an extremely potent capacity for
self-renewal and metastatic spread; they are crucial for tumor
progression and treatment resistance (85). Under the stimulation of a hypoxic
environment, the expression levels of survival and growth factors
in CSCs within the TME increase, and CSC-derived exosomes can act
on recipient cells by transferring these factors. This induces
genetic instability and malignant transformation, thus promoting
CSC survival, differentiation, self-renewal and enhances their
resistance to various cancer therapies (86). Currently, targeting CSCs by
exosomes is also considered a highly promising novel tumor
immunotherapy (87).
Clinical applications of exosomes in tumor
therapy
Diagnostic and monitoring
potential
Exosomes carry genetic materials, proteins and other
molecules that can reflect their cellular origin and disease state,
and these ‘cargoes’ are encapsulated by the plasma membrane, and
can be relatively stable in blood and other body fluids. For these
reasons, exosome-based liquid biopsies have the potential to
function as biomarkers for cancer diagnosis and prognosis, as well
as for the monitoring of disease progression and the
immunotherapeutic response (4).
Exosomal miRNAs are key regulators of gene
expression in recipient cells and are involved in intercellular
communications in the TME, which affects tumor development.
Therefore, exosomal miRNAs have been shown promise as biomarkers
for various tumors and have significant clinical value in the early
diagnosis of tumor metastasis (26,38). It has also been shown that
exosomal mRNAs have potential clinical applications. TDEs have been
reported to contain ~10,000 different mRNA species (88). In previous a retrospective
vaccination study, the mRNA levels of four immunomodulatory genes
(IL-8, ZAP70, TGFB and TIMP1) were significantly
decreased in TDEs isolated from the plasma of patients with
recurrent gliomas, and this change only occurred in patients who
exhibited a clinical response to the vaccine (89). That study indicated that the
analysis of mRNAs in TDEs from plasma of cancer patients receiving
immunotherapy may be helpful in predicting reactivity and
determining prognosis (89).
Exosomes also contain various proteins that reflect
the characteristics of parental cells. Proteins that are aberrantly
expressed in tumor cells can be expressed in exosomes; thus,
exosomal proteins may also serve as potential biomarkers for tumor
diagnosis, as well as for the monitoring of disease progression and
the immunotherapeutic response.
An example of this is the extensively studied
PD-1/PD-L1. It has been demonstrated that higher levels of exosomal
PD-L1 (ExoPD-L1) are present in the circulation of cancer patients
compared to healthy individuals (59), and appear to be consistent with
the expression levels of PD-L1 in their parental tumor cells
(90). However, in another study
on prostate cancer, it was found that cancer cells expressed high
mRNA levels of PD-L1 and produced high levels of ExoPD-L1, whereas
no PD-L1 was present on the surface of the cancer cells (91); this suggests that exosome-based
tumor marker detection can be used to screen some patients with a
normal histology. Furthermore, high levels of circulating ExoPD-L1
have been shown to be associated with the poor prognosis of
patients with various tumors, including metastatic melanoma, and
pancreatic and metastatic gastric cancer (59,92). It also has been shown that a
quantitative increase in circulating ExoPD-L1 levels in patients
with melanoma during early treatment can distinguish the clinical
responders from non-responders (93). A previous prospective study on
melanoma also suggested that monitoring circulating ExoPD-L1 levels
may help to predict treatment efficacy and clinical outcomes
(94). From the aforementioned
findings, it can be concluded that circulating ExoPD-L1 has
potential for use as an effective biomarker for tumor diagnosis,
and for the determination of tumor progression, prognosis and
immunotherapy efficacy. It can also be used as a complement to
tumor tissue PD-L1 expression levels to improve accuracy.
Therapeutic potential
Exosomes as delivery vehicles
The lipid bilayer membrane of exosomes protects the
delivered substances from biodegradation, and exosomes carrying
‘cargo’ also have a low immunogenicity and a long half-life in the
body, which endow them with the high stability in the circulation
and the ability to cross biological barriers. Thus, exosomes can be
modified to deliver a wide range of substances, including
therapeutic RNAs, antitumor drugs, functional proteins,
immunomodulators, etc., in a specific manner to the target cells of
the body to exert their effects (10,83). For example, siRNA targeting RAD51
(a DNA repair protein) can be delivered by exosomes to HeLa cells,
leading to the death of a large proportion of proliferating cells
(95). A previous study on lung
cancer also reported that exosomes loaded with paclitaxel could be
targeted to lung cancer cells and exerted superior antitumor
effects compared to direct treatment with paclitaxel (96).
Exosomes can facilitate drugs or antigenic
substances to specific cells by modifying their own lipids, and
glycosylation has been shown to protect exosomes from protein
hydrolysis and to direct exosomes to specific destinations in
vivo (97). The modification
of exosome membranes with certain markers also allows them to
target specific recipient cells; for example, loading exosome
membranes with unique markers for CSCs (e.g., CD44, CD24, CD133 and
CD200) allows exosomes to target CSCs (10), exerting an inhibitory effect on
tumor progression by targeting CSCs.
Exosomes as cancer vaccines
As exosomes can induce tumor-specific immunity,
their use in cancer treatments has received increasing attention,
and several studies have revealed the potential of exosomes as a
cancer vaccine (98). A previous
study on melanoma found that NK cell-derived exosomes enriched in
perforin and FasL exerted cytotoxic effects on melanoma cells, and
that NK cell-derived exosomes secreted TNF-α, which affected cell
proliferation signaling pathways; the results of that study
demonstrated that NK cell-derived exosomes exerted antitumor
effects against aggressive melanoma both in vitro and in
vivo (99). Another study
demonstrated that exosomes from class II trans-activator
gene-modified mouse melanoma cells expressed MHC II and the tumor
antigen TRP2, and when injected into mice as a vaccine, they were
found to induce a Th1 polarized immune response and stimulated the
production of Th1 IgG2a antibody and IFN-γ, as well as the
upregulation of TRP2-specific CD8+ T-cells, therefore
attenuating tumor growth (100).
DC-derived exosomes (DEX) are enriched in MHC I, MHC
II and CD86, which can stimulate the activation of CD4+
T and CD8+ T-cells, induce CD8+ T-cell
proliferation, and exert potent antitumor effects in vivo
(101). There have been two
phase I clinical trials and one phase II clinical trial analyzing
the feasibility of DEX as a cancer vaccine (102–104). In these clinical trials, some
patients achieved disease stabilization, and a small number of
patients observed a mild response, indicating that DEX as a vaccine
was well-tolerated and safe. However, it is worth noting that in
all three trials, the results of low clinical efficacy and
insufficient antigen-specific T-cell response are presented; thus,
further research is required in order to identify strategies with
which to enhance the DEX-induced T-cell function the antitumor
effect (105).
Targeted inhibition of TDEs to improve
immunotherapy efficacy
As TDEs are involved in several immunomodulatory
processes related to tumorigenesis and tumor development, the
targeted inhibition of the biosynthesis or secretion of TDEs may
prove to be beneficial for the treatment of tumors (59).
Current immunosuppressive therapies and immune
checkpoint blockade mainly focus on the PD-1/PD-L1 pathway;
however, there are certain disadvantages, such as individual
variations in the drug response, differences in efficacy against
various tumors and drug resistance. Targeting exosomal PD-L1 may be
a potentially effective approach to address these issues (82). Previous studies have demonstrated
that anti-PD-L1 antibody therapy has a higher therapeutic efficacy
when combined with inhibitors targeting exosome biosynthesis
(57,91). In addition, researchers have found
that blocking ExoPD-L1 exerts synergistic antitumor effects by
inhibiting not only local tumor growth, but also distant tumor
growth. Furthermore, as previously demonstrated, when tumor cells
previously treated with ExoPD-L1 blockade therapy were re-injected
into the tumor site, the cells did not grow and were rapidly
destroyed, which suggested that the T-cells developed an antitumor
immune memory following ExoPD-L1 blockade (91). Therefore, anti-PD-L1 combined with
targeted TDE inhibition may prove to be a novel approach with which
to improve the efficacy of conventional immunotherapy.
Frontier research advances in exosomes
Due to the wide distribution of exosomes in
vivo and their special biological properties, exosomes have
become a new research hotspot, particularly in the fields of cancer
diagnosis and treatment, targeted drug delivery. In recent years, a
number of studies have also elaborated on the role and mechanisms
of exosomes in neoplastic, neurodegenerative and cardiovascular
diseases, providing several new avenues for the diagnosis and
treatment of these diseases (106). For example, previous multicenter
studies using transmission electron microscopy to analyze exosomes
confirmed the value of exosomes in the diagnosis of pancreatic
cancer and subjective cognitive decline (107,108). Other studies using mouse models
and human breast cancer organoid models have used transmission
electron microscopy and nanoscale flow cytometry to analyze
exosomes, revealing that engineered exosomes can enhance the
antitumor immunity of the body by promoting the activation of type
one conventional DCs in situ (109). The findings of recent studies on
exosomes are presented in Table I
(72,108–117).
 | Table I.Frontier research advances in
exosomes. |
Table I.
Frontier research advances in
exosomes.
| Authors | Date of
publication | Type of study | Exosome research
methods | Conclusions of the
study | Exosome potential
applications | (Refs.) |
|---|
| Li et
al | July, 2022 | Clinical study | Transmission
electron microscopy, Western blot analysis and ELISA | The plasma
NCAM/ABCA1 dual-labeled exosomal Aβ42/40 and miR-384 had potential
advantages in the diagnosis of subjective cognitive decline | Diagnosis of
subjective cognitive decline | (108) |
| Wang et
al | May, 2022 | In vitro
cell lines and mouse model | BCA protein assay,
dynamic light scattering, transmission electron microscopy,
nanosight tracking analysis and nanoparticle tracking analysis | Engineering a 293T
exosome-based delivery platform for efficient tumor-targeting
chemotherapy and internal irradiation combination therapy | Medication
delivery | (110) |
| Huang et
al | February, 2022 | Mouse and model
organoid model | Transmission
electron microscopy, nanoscale flow cytometry and western blot
analysis | HeLa-Exos exhibit
potent antitumor activity by promoting the activation of cDC1s
in situ and thus improving the subsequent tumor-reactive
CD8+ T-cell responses | Tumor
treatment | (109) |
| Bai et
al | February, 2022 | In vitro
cell lines | Transmission
electron microscopy and nanoparticle tracking analysis | Human placental
exosomes suppress the adaptive immune system by promoting the
expression of PD-L1, down-regulating phosphatase and tension
protein homologs in monocytes |
Immunomodulatory | (111) |
| Song et
al | January, 2022 | Clinical study and
mouse model | BCA protein assay,
nanoparticle tracking analysis, transmission electron microscopy
and western blot analysis | miR-155-3p-loaded
M2 macrophages-derived exosomes enhances the growth of
medulloblastoma cells by downregulating WDR82 | Tumor
immunomodulation | (112) |
| Gao et
al | January, 2022 | In vitro
cell lines and mouse model | Nanoparticle
tracking analysis, BCA protein assay, transmission electron
microscopy and western blot analysis | Hepatocellular
carcinoma cells with high expression levels of GOLPH3 can promote
angiogenesis and sorafenib resistance by enhancing exosomal
miR-494-3p secretion | Tumor
microenvironment modulates and mediates drug resistance | (113) |
| Zhang et
al | January, 2022 | Mouse model | Nanosight tracking
analysis, transmission electron microscopy and western blot
analysis | Delivery of
platelet-rich plasma-derived exosomes incorporated in
thermosensitive hydrogel provides a novel approach of cell-free
therapy and has therapeutic effect on subtalar osteoarthritis | Arthritis
treatment | (114) |
| Qin et
al | August, 2021 | Clinical study | Nanoscale liquid
chromatography coupled to tandem mass spectrometry | Patients with RA
have different plasma exosome protein profiles. These proteins not
only take important parts in the vicious circle in the pathogenic
process of RA, but also serve as novel biomarkers in RA diagnosis
and prognosis | Diagnosis and
prognostic determination of RA | (115) |
| Du et
al | April, 2021 | Clinical study | Ultra-performance
liquid chromatography-tandem mass spectrometry | Exosomal
metabolites have the potential to function as biomarkers for
schizophrenia diagnostics | Diagnosis of
schizophrenia | (116) |
| Kohaar et
al | February, 2021 | Clinical study | Nanoparticle
tracking analysis and transmission electron microscopy | Expression levels
of urine exosome-specific genes (PCA3 and PCGEM1) contribute to the
prediction of high-grade prostate cancer | Adjunctive
diagnosis of prostate cancer | (117) |
| Xia et
al | March, 2020 | Clinical study and
mouse model | BCA protein assay,
transmission electron microscopy, particle and molecular size
analyzer and western blot analysis | Helicobacter
pylori infection impaired endothelial function in patients and
mice through exosome-medicated mechanisms | Involved in the
endothelial damage caused by Helicobacter pylori
infection | (72) |
Conclusions and future perspectives
Exosomes, as intercellular message carriers, are
critical mediators of cellular communications, both locally and
systematically. Exosomes produced by tumor cells have been shown to
play an essential connective role among various cells in the TME,
and are closely associated with tumorigenesis, tumor progression
and therapeutic resistance (118). TDEs construct a pro-tumor
metastatic microenvironment by promoting EMT and angiogenesis, as
well as by inducing a pre-metastatic ecological niche. More
importantly, TDEs can inhibit the function of immune cells via
multiple mechanisms, facilitate tumor immune escape, and even
impair the efficacy of immunotherapy. In cancers associated with
microbial infections, exosomes are also involved in infection
factor-associated immune responses, and play a role in promoting
infection and tumor development (119). In addition, due to the
biological properties of exosomes, their potential application in
clinical practice is also highly noteworthy. Compared to
conventional biopsy methods, exosomes based on bodily fluid sample
detection may prove to be superior biomarkers for tumor diagnosis
and monitoring (120).
Furthermore, due to their low immunogenicity and molecule delivery
function, exosomes exhibit immense potential for use in tumor
immunotherapy, particularly as cancer vaccines and targeted
carriers for antigens and drugs (106). As nanoscale particles of lipid
components, exosomes are also able to across biological barriers,
even the blood-brain barrier, an effect which is difficult to
achieve with several drugs (121). Nevertheless, the complex
crosstalk between TDEs and the TME is not yet fully understood and
thus further investigations into this matter are warranted. Another
unresolved issue is the identification of mechanisms with which to
obtain intact and stable exosomes, and to modify and package them
for clinical treatment (122).
The current research progress regarding the u se of exosomes in
tumor therapy also appears to be limited to animal studies and
in vitro experiments; thus further clinical trials are
required for verification (123). Providing a more in-depth
understanding of the characteristics of exosomes and their
functional role in tumorigenesis may provide novel insight and may
further improve antitumor therapeutics and cancer diagnostics in
the future.
Acknowledgements
Not applicable.
Funding
The present study was funded by the National Natural Science
Foundation of China (grant nos. 81700496 and 32000923), the Peking
University Medicine Fund of Fostering Young Scholars' Scientific
and Technological Innovation (grant no. BMU2021PY002), and the
Beijing Municipal Natural Science Foundation (grant no.
7214304).
Availability of data and materials
Not applicable.
Authors' contributions
ML searched the literature and wrote the manuscript.
HC and RD re-examined the literature. YS designed the study. YS and
JC revised the manuscript. All authors contributed to the article
and have read and approved the final manuscript. Data
authentication is not applicable.
Ethical approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Hao Q, Wu Y, Wu Y, Wang P and Vadgama JV:
Tumor-Derived exosomes in tumor-induced immune suppression. Int J
Mol Sci. 23:14612022. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Fu M, Gu J, Jiang P, Qian H, Xu W and
Zhang X: Exosomes in gastric cancer: Roles, mechanisms, and
applications. Mol Cancer. 18:412019. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Gurung S, Perocheau D, Touramanidou L and
Baruteau J: The exosome journey: From biogenesis to uptake and
intracellular signalling. Cell Commun Signal. 19:472021. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Whiteside TL: Tumor-Derived exosomes and
their role in cancer progression. Adv Clin Chem. 74:103–141. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Hurley JH and Ren X: The circuitry of
cargo flux in the ESCRT pathway. J Cell Biol. 185:185–187. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Hou PP and Chen HZ: Extracellular vesicles
in the tumor immune microenvironment. Cancer Lett. 516:48–56. 2021.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Wei D, Zhan W, Gao Y, Huang L, Gong R,
Wang W, Zhang R, Wu Y, Gao S and Kang T: RAB31 marks and controls
an ESCRT-independent exosome pathway. Cell Res. 31:157–177. 2021.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Koh HM, Jang BG and Kim DC: Prognostic
significance of Rab27 expression in solid cancer: A systematic
review and meta-analysis. Sci Rep. 10:141362020. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Borchers AC, Langemeyer L and Ungermann C:
Who's in control? Principles of Rab GTPase activation in
endolysosomal membrane trafficking and beyond. J Cell Biol.
220:e2021051202021. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Mashouri L, Yousefi H, Aref AR, Ahadi AM,
Molaei F and Alahari SK: Exosomes: Composition, biogenesis, and
mechanisms in cancer metastasis and drug resistance. Mol Cancer.
18:752019. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
van Niel G, Porto-Carreiro I, Simoes S and
Raposo G: Exosomes: A common pathway for a specialized function. J
Biochem. 140:13–21. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Wei Z, Batagov AO, Schinelli S, Wang J,
Wang Y, El Fatimy R, Rabinovsky R, Balaj L, Chen CC, Hochberg F, et
al: Coding and noncoding landscape of extracellular RNA released by
human glioma stem cells. Nat Commun. 8:11452017. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Kalluri R: The biology and function of
exosomes in cancer. J Clin Invest. 126:1208–1215. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
He G, Peng X, Wei S, Yang S, Li X, Huang
M, Tang S, Jin H, Liu J, Zhang S, et al: Exosomes in the hypoxic
TME: From release, uptake and biofunctions to clinical
applications. Mol Cancer. 21:192022. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Dorayappan KDP, Wanner R, Wallbillich JJ,
Saini U, Zingarelli R, Suarez AA, Cohn DE and Selvendiran K:
Hypoxia-induced exosomes contribute to a more aggressive and
chemoresistant ovarian cancer phenotype: A novel mechanism linking
STAT3/Rab proteins. Oncogene. 37:3806–3821. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Lewitowicz P, Matykiewicz J, Koziel D,
Chrapek M, Horecka-Lewitowicz A and Gluszek S: CD63 and GLUT-1
overexpression could predict a poor clinical outcome in GIST: A
study of 54 cases with follow-up. Gastroenterol Res Pract.
2016:64783742016. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Valdez SR, Patterson SI, Ezquer ME,
Torrecilla M, Lama MC and Seltzer AM: Acute sublethal global
hypoxia induces transient increase of GAP-43 immunoreactivity in
the striatum of neonatal rats. Synapse. 61:124–137. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Liu J, Ren L, Li S, Li W, Zheng X, Yang Y,
Fu W, Yi J, Wang J and Du G: The biology, function, and
applications of exosomes in cancer. Acta Pharm Sin B. 11:2783–2797.
2021. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Bobrie A and Théry C: Exosomes and
communication between tumours and the immune system: Are all
exosomes equal? Biochem Soc Trans. 41:263–267. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Trams EG, Lauter CJ, Salem N Jr and Heine
U: Exfoliation of membrane ecto-enzymes in the form of
micro-vesicles. Biochim Biophys Acta. 645:63–70. 1981. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Wan Z, Gao X, Dong Y, Zhao Y, Chen X, Yang
G and Liu L: Exosome-mediated cell-cell communication in tumor
progression. Am J Cancer Res. 8:1661–1673. 2018.PubMed/NCBI
|
|
22
|
Mulcahy LA, Pink RC and Carter DR: Routes
and mechanisms of extracellular vesicle uptake. J Extracell
Vesicles. 3:246412014. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Hoshino A, Costa-Silva B, Shen TL,
Rodrigues G, Hashimoto A, Tesic Mark M, Molina H, Kohsaka S, Di
Giannatale A, Ceder S, et al: Tumour exosome integrins determine
organotropic metastasis. Nature. 527:329–335. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Tian X, Shen H, Li Z, Wang T and Wang S:
Tumor-derived exosomes, myeloid-derived suppressor cells, and tumor
microenvironment. J Hematol Oncol. 12:842019. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Whiteside TL, Diergaarde B and Hong CS:
Tumor-Derived Exosomes (TEX) and their role in immuno-oncology. Int
J Mol Sci. 22:62342021. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Wang B, Tan Z and Guan F: Tumor-Derived
exosomes mediate the instability of cadherins and promote tumor
progression. Int J Mol Sci. 20:36522019. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Jiang C, Zhang N, Hu X and Wang H:
Tumor-associated exosomes promote lung cancer metastasis through
multiple mechanisms. Mol Cancer. 20:1172021. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Saleem SN and Abdel-Mageed AB:
Tumor-derived exosomes in oncogenic reprogramming and cancer
progression. Cell Mol Life Sci. 72:1–10. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Rodrigues-Junior DM, Tsirigoti C, Lim SK,
Heldin CH and Moustakas A: Extracellular vesicles and transforming
growth factor β signaling in cancer. Front Cell Dev Biol.
10:8499382022. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Hao Y, Baker D and Ten Dijke P:
TGF-β-Mediated epithelial-mesenchymal transition and cancer
metastasis. Int J Mol Sci. 20:27672019. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Cho JA, Park H, Lim EH and Lee KW:
Exosomes from breast cancer cells can convert adipose
tissue-derived mesenchymal stem cells into myofibroblast-like
cells. Int J Oncol. 40:130–138. 2012.PubMed/NCBI
|
|
32
|
Hsu YL, Hung JY, Chang WA, Lin YS, Pan YC,
Tsai PH, Wu CY and Kuo PL: Hypoxic lung cancer-secreted exosomal
miR-23a increased angiogenesis and vascular permeability by
targeting prolyl hydroxylase and tight junction protein ZO-1.
Oncogene. 36:4929–4942. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Alzahrani FA, El-Magd MA,
Abdelfattah-Hassan A, Saleh AA, Saadeldin IM, El-Shetry ES, Badawy
AA and Alkarim S: Potential effect of exosomes derived from cancer
stem cells and MSCs on progression of DEN-Induced HCC in rats. Stem
Cells Int. 2018:80589792018. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Karabid NM, Wiedemann T, Gulde S, Mohr H,
Segaran RC, Geppert J, Rohm M, Vitale G, Gaudenzi G, Dicitore A, et
al: Angpt2/Tie2 autostimulatory loop controls tumorigenesis. EMBO
Mol Med. 14:e143642022. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Du S, Qian J, Tan S, Li W, Liu P, Zhao J,
Zeng Y, Xu L, Wang Z and Cai J: Tumor cell-derived exosomes deliver
TIE2 protein to macrophages to promote angiogenesis in cervical
cancer. Cancer Lett. 529:168–179. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Liu Y, Gu Y, Han Y, Zhang Q, Jiang Z,
Zhang X, Huang B, Xu X, Zheng J and Cao X: Tumor Exosomal RNAs
promote lung pre-metastatic niche formation by activating alveolar
epithelial TLR3 to recruit neutrophils. Cancer Cell. 30:243–256.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Fabbri M, Paone A, Calore F, Galli R,
Gaudio E, Santhanam R, Lovat F, Fadda P, Mao C, Nuovo GJ, et al:
MicroRNAs bind to Toll-like receptors to induce prometastatic
inflammatory response. Proc Natl Acad Sci USA. 109:E2110–E2116.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Zhou X, Xie F, Wang L, Zhang L, Zhang S,
Fang M and Zhou F: The function and clinical application of
extracellular vesicles in innate immune regulation. Cell Mol
Immunol. 17:323–334. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Naseri M, Bozorgmehr M, Zöller M, Ranaei
Pirmardan E and Madjd Z: Tumor-derived exosomes: The next
generation of promising cell-free vaccines in cancer immunotherapy.
Oncoimmunology. 9:17799912020. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Li W, Mu D, Tian F, Hu Y, Jiang T, Han Y,
Chen J, Han G and Li X: Exosomes derived from Rab27a-overexpressing
tumor cells elicit efficient induction of antitumor immunity. Mol
Med Rep. 8:1876–1882. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Diamond JM, Vanpouille-Box C, Spada S,
Rudqvist NP, Chapman JR, Ueberheide BM, Pilones KA, Sarfraz Y,
Formenti SC and Demaria S: Exosomes Shuttle TREX1-Sensitive
IFN-Stimulatory dsDNA from irradiated cancer cells to DCs. Cancer
Immunol Res. 6:910–920. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Plebanek MP, Angeloni NL, Vinokour E, Li
J, Henkin A, Martinez-Marin D, Filleur S, Bhowmick R, Henkin J,
Miller SD, et al: Pre-metastatic cancer exosomes induce immune
surveillance by patrolling monocytes at the metastatic niche. Nat
Commun. 8:13192017. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Wang M and Zhang B: The immunomodulation
potential of exosomes in tumor microenvironment. J Immunol Res.
2021:37103722021. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Huntington ND, Cursons J and Rautela J:
The cancer-natural killer cell immunity cycle. Nat Rev Cancer.
20:437–454. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Batista IA, Quintas ST and Melo SA: The
interplay of exosomes and NK cells in cancer biology. Cancers
(Basel). 13:4732021. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Hong CS, Sharma P, Yerneni SS, Simms P,
Jackson EK, Whiteside TL and Boyiadzis M: Circulating exosomes
carrying an immunosuppressive cargo interfere with cellular
immunotherapy in acute myeloid leukemia. Sci Rep. 7:146842017.
View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Liu C, Yu S, Zinn K, Wang J, Zhang L, Jia
Y, Kappes JC, Barnes S, Kimberly RP, Grizzle WE and Zhang HG:
Murine mammary carcinoma exosomes promote tumor growth by
suppression of NK cell function. J Immunol. 176:1375–1385. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Paul S and Lal G: The molecular mechanism
of natural killer cells function and its importance in cancer
immunotherapy. Front Immunol. 8:11242017. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Hosseini R, Sarvnaz H, Arabpour M, Ramshe
SM, Asef-Kabiri L, Yousefi H, Akbari ME and Eskandari N: Cancer
exosomes and natural killer cells dysfunction: Biological roles,
clinical significance and implications for immunotherapy. Mol
Cancer. 21:152022. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Liu S, Galat V, Galat Y, Lee YKA,
Wainwright D and Wu J: NK cell-based cancer immunotherapy: From
basic biology to clinical development. J Hematol Oncol. 14:1–17.
2021. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Pritchard A, Tousif S, Wang Y, Hough K,
Khan S, Strenkowski J, Chacko BK, Darley-Usmar VM and Deshane JS:
Lung tumor cell-derived exosomes promote M2 macrophage
polarization. Cells. 9:13032020. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Baig MS, Roy A, Rajpoot S, Liu D, Savai R,
Banerjee S, Kawada M, Faisal SM, Saluja R, Saqib U, et al:
Tumor-derived exosomes in the regulation of macrophage
polarization. Inflamm Res. 69:435–451. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Li X, Lei Y, Wu M and Li N: Regulation of
macrophage activation and polarization by HCC-Derived Exosomal
lncRNA TUC339. Int J Mol Sci. 19:29582018. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Panigrahi GK, Praharaj PP, Peak TC, Long
J, Singh R, Rhim JS, Abd Elmageed ZY and Deep G: Hypoxia-induced
exosome secretion promotes survival of African-American and
Caucasian prostate cancer cells. Sci Rep. 8:38532018. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Wieckowski EU, Visus C, Szajnik M,
Szczepanski MJ, Storkus WJ and Whiteside TL: Tumor-derived
microvesicles promote regulatory T cell expansion and induce
apoptosis in tumor-reactive activated CD8+ T lymphocytes. J
Immunol. 183:3720–3730. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Rasihashemi SZ, Rezazadeh Gavgani E,
Majidazar R, Seraji P, Oladghaffari M, Kazemi T and Lotfinejad P:
Tumor-derived exosomal PD-L1 in progression of cancer and
immunotherapy. J Cell Physiol. 237:1648–1660. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Yang Y, Li CW, Chan LC, Wei Y, Hsu JM, Xia
W, Cha JH, Hou J, Hsu JL, Sun L and Hung MC: Exosomal PD-L1 harbors
active defense function to suppress T cell killing of breast cancer
cells and promote tumor growth. Cell Res. 28:862–864. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Himes BT, Peterson TE, de Mooij T, Garcia
LMC, Jung MY, Uhm S, Yan D, Tyson J, Jin-Lee HJ, Parney D, et al:
The role of extracellular vesicles and PD-L1 in
glioblastoma-mediated immunosuppressive monocyte induction. Neuro
Oncol. 22:967–978. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Zhou K, Guo S, Li F, Sun Q and Liang G:
Exosomal PD-L1: New insights into tumor immune Escape mechanisms
and therapeutic strategies. Front Cell Dev Biol. 8:5692192020.
View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Maybruck BT, Pfannenstiel LW, Diaz-Montero
M and Gastman BR: Tumor-derived exosomes induce CD8+ T cell
suppressors. J Immunother Cancer. 5:652017. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Czystowska-Kuzmicz M, Sosnowska A, Nowis
D, Ramji K, Szajnik M, Chlebowska-Tuz J, Wolinska E, Gaj P, Grazul
M, Pilch Z, et al: Small extracellular vesicles containing
arginase-1 suppress T-cell responses and promote tumor growth in
ovarian carcinoma. Nat Commun. 10:30002019. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Whiteside TL: The role of tumor-derived
exosomes (TEX) in shaping anti-tumor immune competence. Cells.
10:30542021. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Azambuja JH, Ludwig N, Braganhol E and
Whiteside TL: Inhibition of the adenosinergic pathway in cancer
rejuvenates innate and adaptive immunity. Int J Mol Sci.
20:56982019. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Hosseini R, Asef-Kabiri L, Yousefi H,
Sarvnaz H, Salehi M, Akbari ME and Eskandari N: The roles of
tumor-derived exosomes in altered differentiation, maturation and
function of dendritic cells. Mol Cancer. 20:832021. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Ning Y, Shen K, Wu Q, Sun X, Bai Y, Xie Y,
Pan J and Qi C: Tumor exosomes block dendritic cells maturation to
decrease the T cell immune response. Immunol Lett. 199:36–43. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Olejarz W, Dominiak A, Zolnierzak A,
Kubiak-Tomaszewska G and Lorenc T: Tumor-Derived exosomes in
immunosuppression and immunotherapy. J Immunol Res.
2020:62724982020. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Guo X, Qiu W, Liu Q, Qian M, Wang S, Zhang
Z, Gao X, Chen Z, Xue H and Li G: Immunosuppressive effects of
hypoxia-induced glioma exosomes through myeloid-derived suppressor
cells via the miR-10a/Rora and miR-21/Pten Pathways. Oncogene.
37:4239–4259. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Xiang X, Poliakov A, Liu C, Liu Y, Deng
ZB, Wang J, Cheng Z, Shah SV, Wang GJ, Zhang L, et al: Induction of
myeloid-derived suppressor cells by tumor exosomes. Int J Cancer.
124:2621–2633. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Jones LB, Bell CR, Bibb KE, Gu L, Coats MT
and Matthews QL: Pathogens and their effect on exosome biogenesis
and composition. Biomedicines. 6:792018. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Toyofuku M, Nomura N and Eberl L: Types
and origins of bacterial membrane vesicles. Nat Rev Microbiol.
17:13–24. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Shimoda A, Ueda K, Nishiumi S,
Murata-Kamiya N, Mukai SA, Sawada S, Azuma T, Hatakeyama M and
Akiyoshi K: Exosomes as nanocarriers for systemic delivery of the
Helicobacter pylori virulence factor CagA. Sci Rep. 6:183462016.
View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Xia X, Zhang L, Chi J, Li H, Liu X, Hu T,
Li R, Guo Y, Zhang X, Wang H, et al: Helicobacter pylori infection
impairs endothelial function through an exosome-mediated mechanism.
J Am Heart Assoc. 9:e0141202020. View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Oster P, Vaillant L, Riva E, McMillan B,
Begka C, Truntzer C, Richard C, Leblond MM, Messaoudene M, Machremi
E, et al: Helicobacter pylori infection has a detrimental impact on
the efficacy of cancer immunotherapies. Gut. 71:457–466. 2022.
View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Che Y, Geng B, Xu Y, Miao X, Chen L, Mu X,
Pan J, Zhang C, Zhao T, Wang C, et al: Helicobacter pylori-induced
exosomal MET educates tumour-associated macrophages to promote
gastric cancer progression. J Cell Mol Med. 22:5708–5719. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Brennan CA and Garrett WS: Fusobacterium
nucleatum-symbiont, opportunist and oncobacterium. Nat Rev
Microbiol. 17:156–166. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Gholizadeh P, Eslami H and Kafil HS:
Carcinogenesis mechanisms of Fusobacterium nucleatum. Biomed
Pharmacother. 89:918–925. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
77
|
Guo S, Chen J, Chen F, Zeng Q, Liu WL and
Zhang G: Exosomes derived from Fusobacterium nucleatum-infected
colorectal cancer cells facilitate tumour metastasis by selectively
carrying miR-1246/92b-3p/27a-3p and CXCL16. Gut. Nov 10–2020.(Epub
ahead of print).
|
|
78
|
Kapoor NR, Chadha R, Kumar S, Choedon T,
Reddy VS and Kumar V: The HBx gene of hepatitis B virus can
influence hepatic microenvironment via exosomes by transferring its
mRNA and protein. Virus Res. 240:166–174. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
79
|
Jiang Y, Han Q, Zhao H and Zhang J: The
Mechanisms of HBV–Induced hepatocellular carcinoma. J Hepatocell
Carcinoma. 8:435–450. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
80
|
Li S, Li S, Wu S and Chen L: Exosomes
modulate the viral replication and host immune responses in HBV
infection. Biomed Res Int. 2019:21039432019.PubMed/NCBI
|
|
81
|
Shi Y, Du L, Lv D, Li H, Shang J, Lu J,
Zhou L, Bai L and Tang H: Exosomal interferon-induced transmembrane
protein 2 transmitted to dendritic cells inhibits interferon alpha
pathway activation and blocks anti-hepatitis B virus efficacy of
exogenous interferon alpha. Hepatology. 69:2396–2413.
2019.PubMed/NCBI
|
|
82
|
Ye L, Zhu Z, Chen X, Zhang H, Huang J, Gu
S and Zhao X: The importance of exosomal PD-L1 in cancer
progression and its potential as a therapeutic target. Cells.
10:32472021. View Article : Google Scholar : PubMed/NCBI
|
|
83
|
Rizwan MN, Ma Y, Nenkov M, Jin L, Schröder
DC, Westermann M, Gaßler N and Chen Y: Tumor-derived exosomes: Key
players in non-small cell lung cancer metastasis and their
implication for targeted therapy. Mol Carcinog. 61:269–280. 2022.
View Article : Google Scholar : PubMed/NCBI
|
|
84
|
Jing C, Cao H, Qin X, Yu S, Wu J, Wang Z,
Ma R and Feng J: Exosome-mediated gefitinib resistance in lung
cancer HCC827 cells via delivery of miR-21. Oncol Lett.
15:9811–9817. 2018.PubMed/NCBI
|
|
85
|
Lin S, Xu Y, Gan Z, Han K, Hu H, Yao Y,
Huang M and Min D: Monitoring cancer stem cells: Insights into
clinical oncology. Onco Targets Ther. 9:731–740. 2016.PubMed/NCBI
|
|
86
|
Fatima F and Nawaz M: Stem cell-derived
exosomes: Roles in stromal remodeling, tumor progression, and
cancer immunotherapy. Chin J Cancer. 34:541–553. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
87
|
Clara JA, Monge C, Yang Y and Takebe N:
Targeting signalling pathways and the immune microenvironment of
cancer stem cells-a clinical update. Nat Rev Clin Oncol.
17:204–232. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
88
|
Skog J, Würdinger T, van Rijn S, Meijer
DH, Gainche L, Sena-Esteves M, Curry WT Jr, Carter BS, Krichevsky
AM and Breakefield XO: Glioblastoma microvesicles transport RNA and
proteins that promote tumour growth and provide diagnostic
biomarkers. Nat Cell Biol. 10:1470–1476. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
89
|
Muller L, Muller-Haegele S, Mitsuhashi M,
Gooding W, Okada H and Whiteside TL: Exosomes isolated from plasma
of glioma patients enrolled in a vaccination trial reflect
antitumor immune activity and might predict survival.
Oncoimmunology. 4:e10083472015. View Article : Google Scholar : PubMed/NCBI
|
|
90
|
Kim DH, Kim H, Choi YJ, Kim SY, Lee JE,
Sung KJ, Sung YH, Pack CG, Jung MK, Han B, et al: Exosomal PD-L1
promotes tumor growth through immune escape in non-small cell lung
cancer. Exp Mol Med. 51:1–13. 2019. View Article : Google Scholar
|
|
91
|
Poggio M, Hu T, Pai CC, Chu B, Belair CD,
Chang A, Montabana E, Lang UE, Fu Q, Fong L and Blelloch R:
Suppression of exosomal PD-L1 induces systemic anti-tumor immunity
and memory. Cell. 177:414–427.e13. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
92
|
Fan Y, Che X, Qu J, Hou K, Wen T, Li Z, Li
C, Wang S, Xu L, Liu Y and Qu X: Exosomal PD-L1 retains
immunosuppressive activity and is associated with gastric cancer
prognosis. Ann Surg Oncol. 26:3745–3755. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
93
|
Chen G, Huang AC, Zhang W, Zhang G, Wu M,
Xu W, Yu Z, Yang J, Wang B, Sun H, et al: Exosomal PD-L1
contributes to immunosuppression and is associated with anti-PD-1
response. Nature. 560:382–386. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
94
|
Cordonnier M, Nardin C, Chanteloup G,
Derangere V, Algros MP, Arnould L, Garrido C, Aubin F and Gobbo J:
Tracking the evolution of circulating exosomal-PD-L1 to monitor
melanoma patients. J Extracell Vesicles. 9:17108992020. View Article : Google Scholar : PubMed/NCBI
|
|
95
|
Shtam TA, Kovalev RA, Varfolomeeva EY,
Makarov EM, Kil YV and Filatov MV: Exosomes are natural carriers of
exogenous siRNA to human cells in vitro. Cell Commun Signal.
11:882013. View Article : Google Scholar : PubMed/NCBI
|
|
96
|
Kim MS, Haney MJ, Zhao Y, Yuan D, Deygen
I, Klyachko NL, Kabanov AV and Batrakova EV: Engineering
macrophage-derived exosomes for targeted paclitaxel delivery to
pulmonary metastases: In vitro and in vivo evaluations.
Nanomedicine. 14:195–204. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
97
|
Taghikhani A, Farzaneh F, Sharifzad F,
Mardpour S, Ebrahimi M and Hassan ZM: Engineered tumor-derived
extracellular vesicles: Potentials in cancer immunotherapy. Front
Immunol. 11:2212020. View Article : Google Scholar : PubMed/NCBI
|
|
98
|
Xu Z, Zeng S, Gong Z and Yan Y:
Exosome-based immunotherapy: A promising approach for cancer
treatment. Mol Cancer. 19:1602020. View Article : Google Scholar : PubMed/NCBI
|
|
99
|
Zhu L, Kalimuthu S, Gangadaran P, Oh JM,
Lee HW, Baek SH, Jeong SY, Lee SW, Lee J and Ahn BC: Exosomes
derived from natural killer cells exert therapeutic effect in
melanoma. Theranostics. 7:2732–2745. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
100
|
Lee YS, Kim SH, Cho JA and Kim CW:
Introduction of the CIITA gene into tumor cells produces exosomes
with enhanced anti-tumor effects. Exp Mol Med. 43:281–290. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
101
|
André F, Chaput N, Schartz NE, Flament C,
Aubert N, Bernard J, Lemonnier F, Raposo G, Escudier B, Hsu DH, et
al: Exosomes as potent cell-free peptide-based vaccine. I.
Dendritic cell-derived exosomes transfer functional MHC class
I/peptide complexes to dendritic cells. J Immunol. 172:2126–2136.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
102
|
Morse MA, Garst J, Osada T, Khan S,
Hobeika A, Clay TM, Valente N, Shreeniwas R, Sutton MA, Delcayre A,
et al: A phase I study of dexosome immunotherapy in patients with
advanced non-small cell lung cancer. J Transl Med. 3:92005.
View Article : Google Scholar : PubMed/NCBI
|
|
103
|
Escudier B, Dorval T, Chaput N, André F,
Caby MP, Novault S, Flament C, Leboulaire C, Borg C, Amigorena S,
et al: Vaccination of metastatic melanoma patients with autologous
dendritic cell (DC) derived-exosomes: Results of thefirst phase I
clinical trial. J Transl Med. 3:102005. View Article : Google Scholar : PubMed/NCBI
|
|
104
|
Besse B, Charrier M, Lapierre V, Dansin E,
Lantz O, Planchard D, Le Chevalier T, Livartoski A, Barlesi F,
Laplanche A, et al: Dendritic cell-derived exosomes as maintenance
immunotherapy after first line chemotherapy in NSCLC.
Oncoimmunology. 5:e10710082015. View Article : Google Scholar : PubMed/NCBI
|
|
105
|
Yao Y, Fu C, Zhou L, Mi QS and Jiang A:
DC-Derived exosomes for cancer immunotherapy. Cancers (Basel).
13:36672021. View Article : Google Scholar : PubMed/NCBI
|
|
106
|
Kalluri R and LeBleu VS: The biology,
function, and biomedical applications of exosomes. Science.
367:eaau69772020. View Article : Google Scholar : PubMed/NCBI
|
|
107
|
Xiao D, Dong Z, Zhen L, Xia G, Huang X,
Wang T, Guo H, Yang B, Xu C, Wu W, et al: Combined Exosomal GPC1,
CD82, and Serum CA19-9 as multiplex targets: A specific, sensitive,
and reproducible detection panel for the diagnosis of pancreatic
cancer. Mol Cancer Res. 18:300–310. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
108
|
Li Y, Meng S, Di W, Xia M, Dong L, Zhao Y,
Ling S, He J, Xue X, Chen X and Liu C: Amyloid-β protein and
MicroRNA-384 in NCAM-Labeled exosomes from peripheral blood are
potential diagnostic markers for Alzheimer's disease. CNS Neurosci
Ther. 28:1093–1107. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
109
|
Huang L, Rong Y, Tang X, Yi K, Qi P, Hou
J, Liu W, He Y, Gao X, Yuan C and Wang F: Engineered exosomes as an
in situ DC-primed vaccine to boost antitumor immunity in breast
cancer. Mol Cancer. 21:452022. View Article : Google Scholar : PubMed/NCBI
|
|
110
|
Wang C, Li N, Li Y, Hou S, Zhang W, Meng
Z, Wang S, Jia Q, Tan J, Wang R and Zhang R: Engineering a HEK-293T
exosome-based delivery platform for efficient tumor-targeting
chemotherapy/internal irradiation combination therapy. J
Nanobiotechnology. 20:2472022. View Article : Google Scholar : PubMed/NCBI
|
|
111
|
Bai K, Lee CL, Liu X, Li J, Cao D, Zhang
L, Hu D, Li H, Hou Y, Xu Y, et al: Human placental exosomes induce
maternal systemic immune tolerance by reprogramming circulating
monocytes. J Nanobiotechnology. 20:862022. View Article : Google Scholar : PubMed/NCBI
|
|
112
|
Song L, Luan B, Xu Q, Shi R and Wang X:
microRNA-155-3p delivered by M2 macrophages-derived exosomes
enhances the progression of medulloblastoma through regulation of
WDR82. J Transl Med. 20:132022. View Article : Google Scholar : PubMed/NCBI
|
|
113
|
Gao Y, Yin Z, Qi Y, Peng H, Ma W, Wang R
and Li W: Golgi phosphoprotein 3 promotes angiogenesis and
sorafenib resistance in hepatocellular carcinoma via upregulating
exosomal miR-494-3p. Cancer Cell Int. 22:352022. View Article : Google Scholar : PubMed/NCBI
|
|
114
|
Zhang Y, Wang X, Chen J, Qian D, Gao P,
Qin T, Jiang T, Yi J, Xu T, Huang Y, et al: Exosomes derived from
platelet-rich plasma administration in site mediate cartilage
protection in subtalar osteoarthritis. J Nanobiotechnology.
20:562022. View Article : Google Scholar : PubMed/NCBI
|
|
115
|
Qin Q, Song R, Du P, Gao C, Yao Q and
Zhang JA: Systemic proteomic analysis reveals distinct exosomal
protein profiles in rheumatoid arthritis. J Immunol Res.
2021:94217202021. View Article : Google Scholar : PubMed/NCBI
|
|
116
|
Du Y, Chen L, Li XS, Li XL, Xu XD, Tai SB,
Yang GL, Tang Q, Liu H, Liu SH, et al: Metabolomic identification
of exosome-derived biomarkers for Schizophrenia: A large
multicenter study. Schizophr Bull. 47:615–623. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
117
|
Kohaar I, Chen Y, Banerjee S, Borbiev T,
Kuo HC, Ali A, Ravindranath L, Kagan J, Srivastava S, Dobi A, et
al: A urine exosome gene expression panel distinguishes between
indolent and aggressive prostate cancers at biopsy. J Urol.
205:420–425. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
118
|
Jahan S, Mukherjee S, Ali S, Bhardwaj U,
Choudhary RK, Balakrishnan S, Naseem A, Mir SA, Banawas S,
Alaidarous M, et al: Pioneer role of extracellular vesicles as
modulators of cancer initiation in progression, drug therapy, and
vaccine prospects. Cells. 11:4902022. View Article : Google Scholar : PubMed/NCBI
|
|
119
|
Wang J, Yao Y, Chen X, Wu J, Gu T and Tang
X: Host derived exosomes-pathogens interactions: Potential
functions of exosomes in pathogen infection. Biomed Pharmacother.
108:1451–1459. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
120
|
Yu W, Hurley J, Roberts D, Chakrabortty
SK, Enderle D, Noerholm M, Breakefield XO and Skog JK:
Exosome-based liquid biopsies in cancer: Opportunities and
challenges. Ann Oncol. 32:466–477. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
121
|
Zagrean AM, Hermann DM, Opris I, Zagrean L
and Popa-Wagner A: Multicellular crosstalk between exosomes and the
neurovascular unit after cerebral ischemia. Therapeutic
Implications. Front Neurosci. 12:8112018. View Article : Google Scholar : PubMed/NCBI
|
|
122
|
Luo L, Wu Z, Wang Y and Li H: Regulating
the production and biological function of small extracellular
vesicles: Current strategies, applications and prospects. J
Nanobiotechnology. 19:4222021. View Article : Google Scholar : PubMed/NCBI
|
|
123
|
Rajagopal C and Harikumar KB: The origin
and functions of exosomes in cancer. Front Oncol. 8:662018.
View Article : Google Scholar : PubMed/NCBI
|














