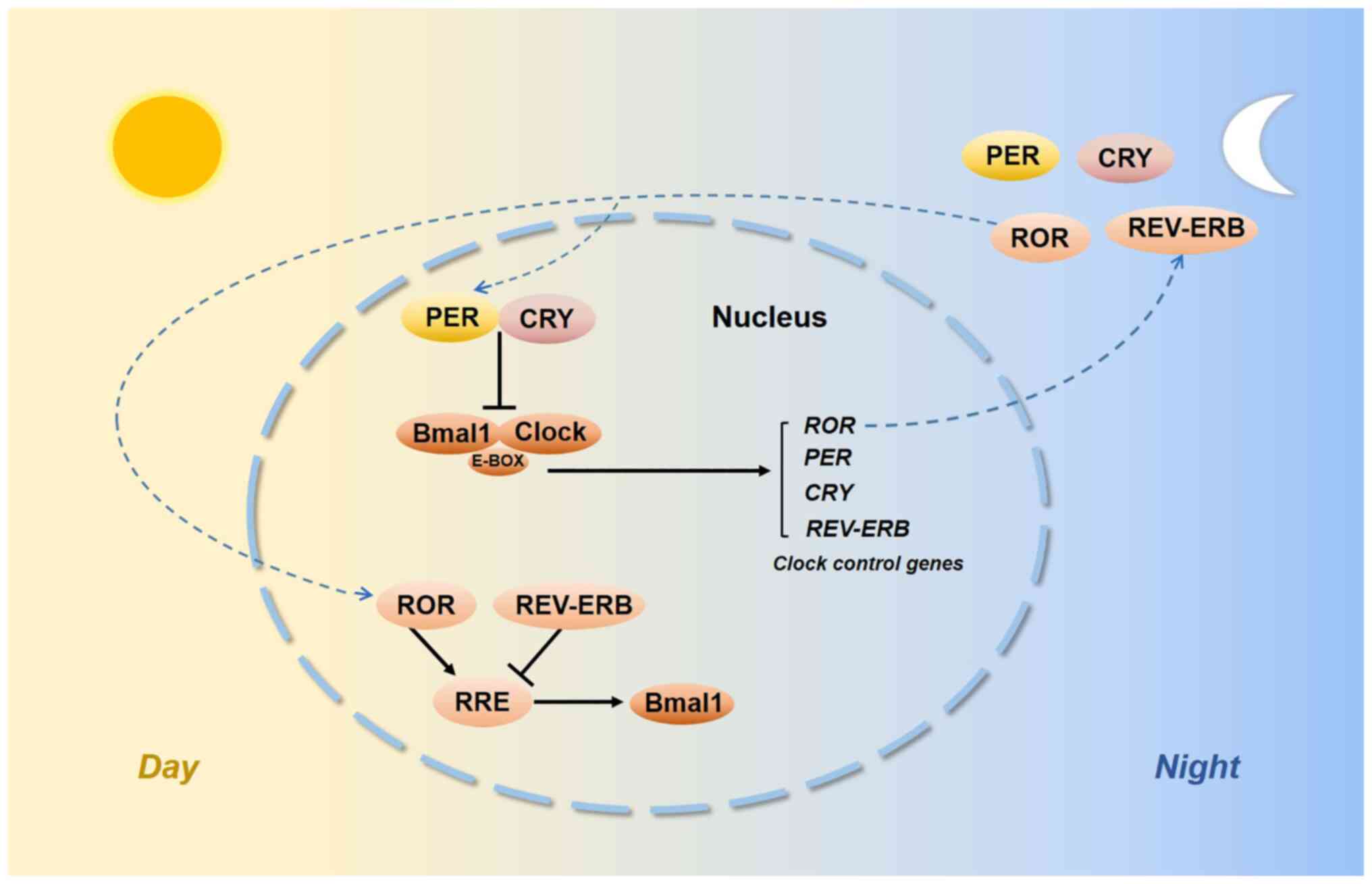
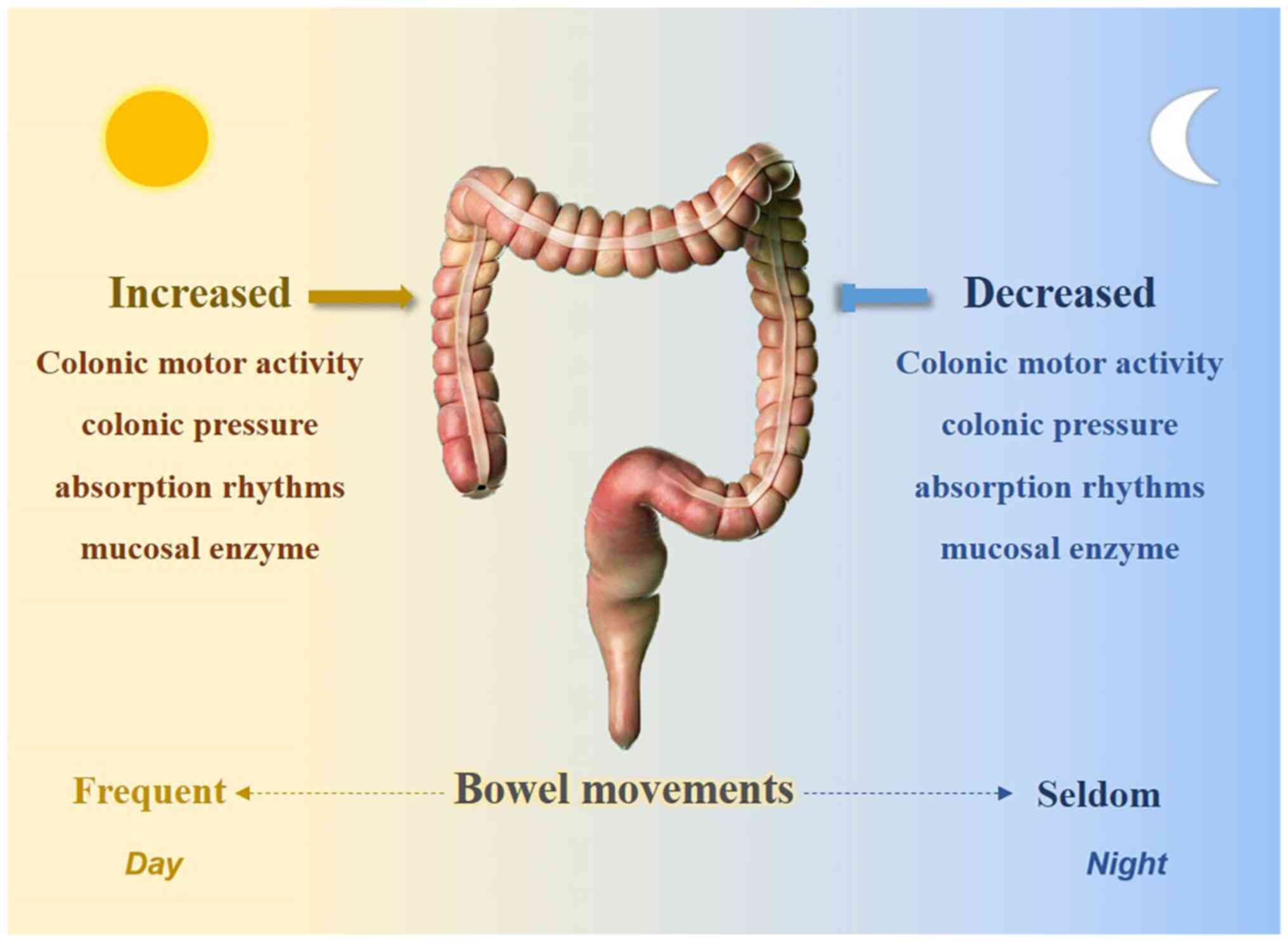
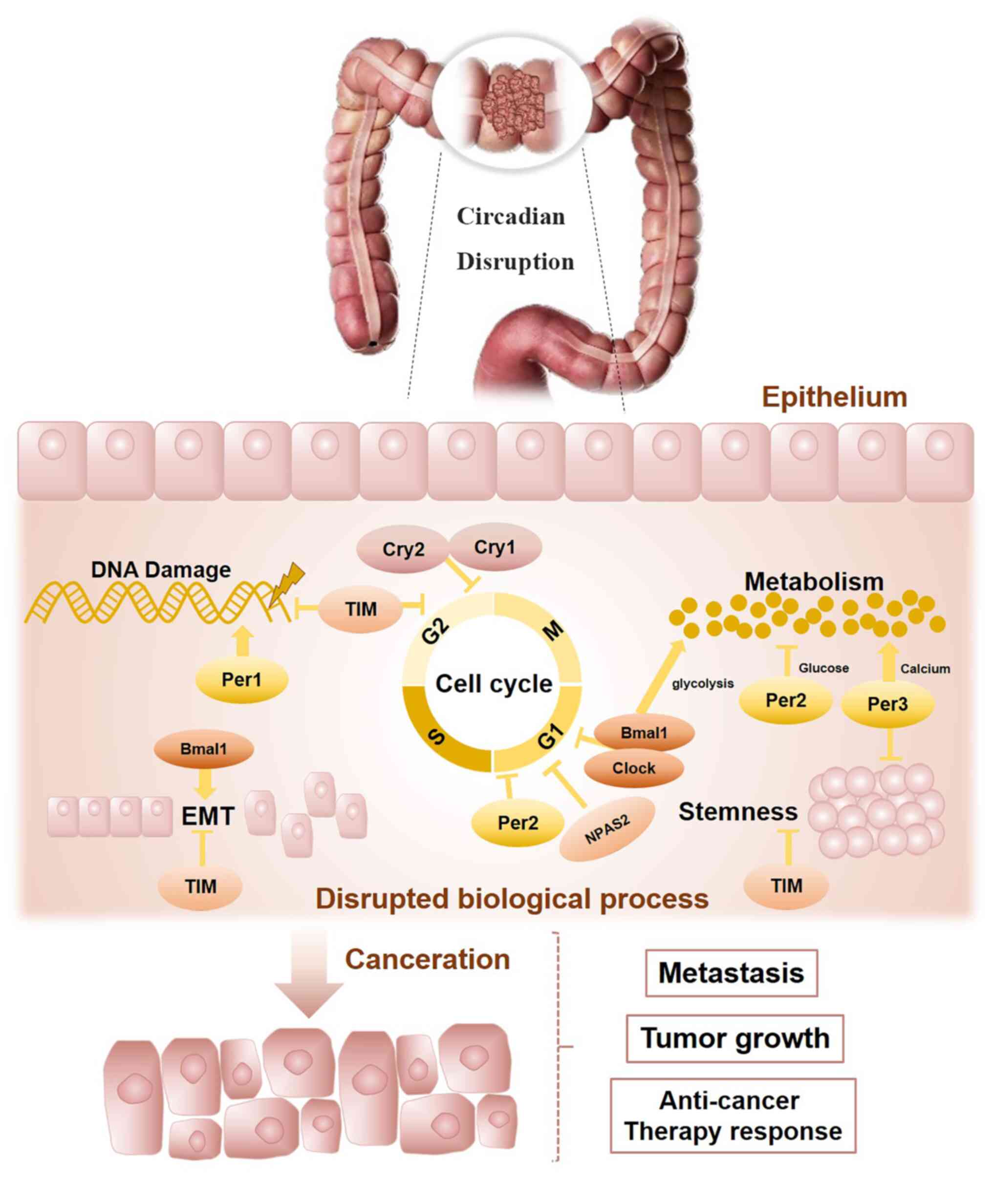
Biological rhythms exist in our internal bodies and
their functions vary according to oscillations with a 24-h
light/dark cycle (1). The
circadian time-keeping system is controlled by both central and
peripheral oscillators. The suprachiasmatic nuclei of the
hypothalamus are the central pacemaker (2,3) and
certain interconnected specific clock genes control the peripheral
circadian rhythm. The discovery of the circadian locomotor output
cycles kaput (CLOCK) and its heterodimeric partner brain and muscle
ARNT-like-1 (BMAL1) led to the first site of circadian oscillations
(4,5). The transactivation of
E-box-containing genes by the CLOCK:BMAL1 complex was able to
activate Period (Per) genes, i.e. Per1-3 and Cryptochrome (Cry)
genes, i.e. Cry1 and Cry2, which then negatively repressed
CLOCK:BMAL1 transcription. As auxiliary feedback loops, REV-ERB and
retinoic acid receptor-related orphan receptors (RORs) bind to a
common response element known as the REV-ERB/ROR response element
(RRE) and their intrinsic repressive and inductive activities have
been proposed to contribute to CLOCK function and BMAL1
transcription (6,7) (Fig.
1). Histone acetylation induces an open chromatin conformation
that is thought to activate gene expression (8). Deacetylation would shift back to
condensing chromatin and silencing gene expression. The enzymes
that participate in these transitions are histone
acetyltransferases (HATs) and histone deacetylases (HDACs)
(9). CLOCK is an enzyme with HAT
activity (10) and CLOCK
specifically acetylates nonhistone targets, such as BMAL1 (11). Sirtuin1 (SIRT1) is an
NAD+-dependent HDAC, and SIRT1 and CLOCK converge in a coordinated
manner. SIRT1 activity depends on nicotinamide phosphoribosyl
transferase, which is the rate-limiting enzyme involved in NAD+
synthesis, which is regulated by CLOCK:BMAL1 (12,13).
The functions of other clock genes, such as timeless (TIM) and
timeless-interacting protein, in the circadian system remain to be
fully elucidated.
Shifts in wake and sleep schedules or sleep
deprivation desynchronize the circadian rhythm. Emerging evidence
suggests that impaired circadian rhythms may be linked to causal
webs of cancer initiation (14,15).
Colorectal cancer (CRC) is one of the cancers that is closely
associated with circadian disruption (16-18).
CRC is the world's fourth most deadly cancer and approximately one
in four CRC cases will be diagnosed with metastatic CRC (mCRC)
(19). Multiple genetic factors
contribute to the development of carcinogenesis of the large bowel,
such as amplification of human epidermal growth factor receptor 2
(HER2), as well as KRAS, NRAS and BRAF mutations (20,21),
chromosomal instability such as microsatellite instability (MSI) or
mismatch repair (MMR) (22), which
all provide a basis for optimal therapeutic intervention for
individuals. Surgical excision is the first choice of treatment for
CRC and chemotherapy that is given in combination with different
molecular therapies (such as bevacizumab, panitumumab, cetuximab,
regorafenib and aflibercept) significantly extends overall survival
(OS) (23). Experimental data
further indicated how the clock gene network exerts an influence on
CRC progression, offers potential biomarkers for prognosis, serves
as a guide for treatment decisions and provides molecular targets
for treating CRC.
Rhythmic processes were already characterized within
the intestine prior to the identification of the molecular clock.
Bowel movements usually occur during the day and certain studies
have suggested that a potential biological clock exists in the
bowel. Research has indicated that colonic motor activity is
minimal during sleep but increases significantly at morning
awakening (24). Rao et al
(25) determined a significant
increase in colonic pressure activity after awakening. Other
physiologically relevant factors, such as absorption rhythms
(26) and mucosal enzymes
(27), also exhibited their lowest
activities at night (Fig. 2).
Furthermore, the rhythmicity of circular muscle contractility and
intracolonic pressure were absent in Per1/2 double-knockout mice
(28). In addition, the gut
microbiota exhibits circadian oscillations in both mice and humans;
Per1/2 double-knockout mice lose rhythmic fluctuations in
microbiota compositions and time shift-induced jet lag also
disrupts diurnal oscillations in commensal bacterial abundances
(29).
Studies have determined that clock genes have an
important role in intestinal physiological activities. Per1-2,
Cry1, CLOCK, BMAL1 and REV-ERBα are expressed in the epithelial
cells of the colon and exhibit circadian rhythmicity (30). Per2 and BMAL1 in the myenteric
plexus and epithelial cells have an important role in coordinating
gastrointestinal functions, such as cell proliferation and
migration (31), and rectal cell
proliferation fluctuates during the day in colon cancer (32). BMAL1 was also reported to regulate
intestinal drug disposition function (33).
These results clearly support that the circadian
system is strongly connected with the gastrointestinal tract.
Clinical research indicated that disruption of the
circadian rhythm accelerated CRC progression (34,35).
Loss of circadian rhythms was also observed to potentiate CRC
initiation in an Apcmin model (36). Abnormal expression levels of
circadian clock genes were observed in CRC tissues and current
studies suggested that clock genes, mainly including Per, Cry,
ARNT1 (BMAL1) and CLOCK, may influence colon cancer
progression.
Period genes are identified most frequently in CRC.
Per1-3 were reported to have tumour suppressor properties, but
certain studies reached different conclusions (Table I). A total of seven studies
determined that the expression of Per1 was significantly decreased
in tumour tissue compared to healthy mucosa (37-44).
Furthermore, one study indicated a decrease in Per1 in
undifferentiated tumours, while no differences were obtained in
differentiated colon carcinomas (45), but two studies found no significant
differences in Per1 between colon tumours and healthy mucosa
(46,47). Mostafaie et al (40) demonstrated a correlated decrease in
Per1 and estrogen receptor-β in colorectal tumours. Furthermore,
six studies reported that Per2 expression was significantly
decreased in tumour tissues (39,48-52).
Hasakova et al (50,53)
found that Per2 was downregulated in tumour tissues of male
patients with CRC, but Krugluger et al (45) determined that Per2 expression
exhibited no differences in colon tumour tissues; the following six
studies also confirmed this (38,40-42,47).
Another six studies examined the gene expression of Per3 and all
found significant decreases (37-39,41,42,54).
In addition, Alexander et al (55) found a Per3 gene length
polymorphism, and the 5-repeat Per3 VNTR sequence may increase the
chance of colorectal adenoma formation.
Studies have also explored the relationships among
period genes and clinical-pathological features of patients with
CRC. Compared to tumours that did not express period genes, Momma
et al (56) indicated that
positive Per1 and Per2 staining was associated with larger tumour
sizes. Tumours with positivity for Per2 expression tended to have a
greater depth of invasion and were generally more advanced, and
patients tended to exhibit poorer OS. He et al (37) reported that Per2 was correlated
with immune cell infiltration and was associated with unfavorable
prognosis in colon adenocarcinoma. Hasakova et al (50,53)
determined that Per2 was significantly downregulated in tumour
tissues of male patients with CRC, and a negative correlation
between Per2 and microRNA (miR)-34a was found. Low expression of
Per2 and high expression of miR-34a were associated with
significantly better outcomes in male patients. However, certain
studies obtained the opposite result, claiming that high Per2
expression was associated with better survival and that low
expression of the Per1 gene was related to liver metastasis
(42). Low expression of Per2 was
significantly associated with CRC metastasis (49). Mazzoccoli et al (39) indicated that lower expression of
Per1 and Per3 in tumour tissues may be suggestive of a poorer
survival prognosis. Wu et al (43) next determined that low expression
levels of Per1 and miR-192 were correlated with unfavourable
survival in patients with stage III/IV CRC but with better OS rates
in patients with stage II CRC. In addition, Štorcelová et al
(57) identified certain cell
cycle regulatory genes related to the Per2 expression in human CRC
tissues.
Substantial evidence indicates that the cell cycle
occurs with a daily rhythm. Healthy mice exhibited rhythmicity of
Per1, Per2, Wee1 and p21 in the intestine, but the circadian
rhythmicity was significantly reduced in tumours (58). Knocking down Per2 subsequently
inhibited p53 and caused CRC cells to acquire malignant biological
features (52). Studies have
explained how periodic genes regulate CRC progression by
interfering with the cell cycle. DNA double-strand breaks activate
the DNA damage response (DDR). The DDR is initiated by kinases such
as ataxia telangiectasia mutated (ATM) (59) and the G1-S checkpoint is regulated
by serine/threonine-protein kinase checkpoint kinase 2 (CHK2),
whereas the G2/M checkpoint is regulated by CHK1 (60). Gery et al (61) reported that Per1 promoted DNA
damage-induced apoptosis by interacting with ATM and CHK2. In
addition, overexpression of c-Myc is associated with CRC (62); in Per1-knockout mice and
Per2−mutant mice, c-Myc expression levels are elevated
without restriction, inducing excess cyclin D1 expression that
increases proliferation (63). Of
note, Per2 has been indicated to be suppressed in colon cancer cell
lines, which increased cyclin D and cell proliferation (64). Another study also supported that
Per2 functions in tumour suppression by regulating the cell cycle
(65).
Crosstalk between the circadian clock and
Wnt/β-catenin signalling has been reported to be involved in CRC.
β-catenin is a main signal transducer of the Wnt pathway (66) and activation of the Wnt/β-catenin
pathway promotes cell proliferation and invasion (67). Wood et al (64) demonstrated that downregulation of
Per2 in colon cells increased β-catenin expression. In in
vivo experiments, when compared to Apc(Min/+) mice, Per2-mutant
Apc(Min/+) mice developed smaller colonic polyps. A study from the
same group, by Yang et al (68), discovered that increased levels of
β-catenin in Apc(Min/+) mice could destabilize the Per2 protein. In
addition, period genes were reported to inhibit drug resistance in
CRC cells. Per2 was significantly upregulated when glucose
metabolism was restricted and thus weakened the ability of cancer
cells to survive chemotherapy (69). Per3 was reported to be
downregulated in drug-resistant CRC cells and cancer stem-like
cells (CSCs). Overexpression of Per3 strengthened the
5-fluorouracil-induced inhibitory effects on colorectal CSCs, while
knockdown of Per3 decreased its inhibitory effects. In addition,
Per3 overexpression decreased stemness markers, such as CD44, CD133
and SOX2. It was confirmed that Per3 reduces the chemoresistance
and self-renewal capability of CRC by inhibiting both Notch and
Wnt/β-catenin signalling (70).
Calcium significantly prevents tumorigenesis of CRC and Per3
significantly upregulates calcium to prevent CRC tumorigenesis
(71).
The Cry1 and Cry2 genes were also associated with
adverse clinical-pathological features in patients with CRC. It was
determined that high Cry1 expression was correlated with poor OS
rates in patients with CRC (73).
Another study also indicated that high expression of Cry1 was
associated with lower OS and disease-free survival at five years in
patients with CRC (49). Hasakova
et al (50,72) indicated that the expression of Cry1
and Cry2 was correlated differently with sex or tumour location in
colon carcinoma tissues. Cry1 expression increased significantly in
females with distant metastases, while females without distant
metastases frequently exhibited downregulation of Cry2 expression.
Better survival rates were associated with low expression of Cry2,
high expression of Cry1 in tumour tissue was associated with an
unfavorable survival prognosis, but this was not observed in males.
In addition, higher Cry1 expression in right-sided colon tumour
tissues was associated with worse survival rates in females, and
the expression of Cry1 in the left-sided colon tumour tissues was
higher than in the adjacent tissue in males. Furthermore, Cry2
expression was associated with tumour location in males with grade
2 cancer. However, Mazzoccoli et al (74) indicated that patients with CRC with
elevated Cry1 or Cry2 expression levels had poorer survival rates.
The Cry1 gene was particularly decreased in elderly female patients
with CRC and tumours located in the transverse colon tended to
exhibit lower Cry expression levels.
CLOCK and its heterodimeric partner, ARNTL1 (BMAL1),
are the core components of the circadian clock system, and modified
expression of these two genes in CRC tumour tissues is related to
clinico-pathological features (Table
III).
BMAL1 may be a biomarker for CRC treatment
prognoses. Studies indicated that the expression levels of BMAL1
decreased by half in mCRC tumour tissues, high expression levels of
BMAL1 were associated with reduced efficacy of the anti-angiogenic
drug bevacizumab (Beva) and patients who had low expression levels
of BMAL1 tended to have good responses to Beva therapy (78); furthermore, upregulation of the
BMAL1 gene was correlated with liver metastasis in patients with
CRC (42). Patients with CRC with
high BMAL1 expression exhibited a significant enrichment of
epithelial-mesenchymal transition (EMT) and invasion-associated
gene signatures (79). Another
study suggested that BMAL1 expression decreased in CRC (37), and low BMAL1 expression was
significantly associated with metastasis (49). However, Karantanos et al
(38) determined that the BMAL1
gene expression levels were higher in cancerous tissues, but no
correlation was found with the clinical significance.
Although the BMAL1 and CLOCK expression levels were
observed to be related to advanced pathological clinical features,
certain experiments have indicated that these two genes may inhibit
CRC cell growth. This inconsistency requires to be clarified by
further research.
Altered expression levels of other circadian clock
genes were also detected in CRC tissue samples (Table IV). Compared to adjacent normal
colon tissues, the SIRT1 and RORA expression levels decreased in
CRC tumour tissues, while TIM was reported to be increased
(37,39,93,94).
High TIM levels were an unfavourable prognostic factor
significantly associated with microsatellite instability and
proximal lymph nodes and were prevalent in TNM stages III-IV
(39). However, the most recent
study indicated that loss of TIM expression is associated with
advanced tumour stage, metastasis and microsatellite stability
status, and TIM expression levels are inversely correlated with a
set of gene signatures of EMT markers (95). In addition, NPAS2 expression was
significantly downregulated in CRC tumour tissues. Low NPAS2
expression was associated with tumour size, TNM stage and
metastases (96).
TIM has no apparent circadian clock function in
mammals, but research has identified it as being important for CRC
progression (97). Recent data
revealed that TIM depletion increases γH2AX, a marker of DNA
damage, and increases CHK1 and CDK1 phosphorylation (98). A report also demonstrated that
overexpression of TIM in colon cancer cells suppressed G2/M arrest,
while TIM depletion increased CHK1 and CDK1 phosphorylation and
triggered G2/M arrest (98).
Another study suggested that TIM inhibited EMT, and its ectopic
silencing promoted invasion, migration and stemness in CRC cells
(95). Overexpression of SIRT1 in
Apc(Min/+) mice resulted in reduced neoplasia, while SIRT1-knockout
mice had an increased tumour incidence (99). NPAS2 increased the G0/G1 phase
population in CRC cells and inhibited proliferation, invasion and
migration in vitro (96).
Circadian scheduling produced advantages in
chemotherapy treatments of patients with CRC. A 24-h programmable
time ambulatory pump was used to administer oxaliplatin by venous
infusion against mCRC. This type of treatment increased the maximum
tolerated doses and decreased the toxicities of oxaliplatin
(100). Another previous phase II
trial also proved that if chemo-drug delivery was chronomodulated
rather than constant over time, chemotherapy was more effective and
less toxic (101). The addition
of cetuximab to chronotherapy in patients with mCRC increased the
chance of complete resection compared to conventional chemotherapy
(102). However, an international
randomized trial reported that chronomodulated FLO4 provided no
survival benefit compared with ordinary FOLFOX2 in patients with
mCRC (103). Of note, Innominato
et al (104)
chronomodulated irinotecan, oxaliplatin, 5-fluorouracil and
leucovorin dosing times against mCRC and found encouraging activity
in second-line treatments, with limited haematological toxicity.
Another study indicated sex differences in the advantages of
chronomodulated irinotecan delivery to minimize adverse events; it
is better to administer irinotecan in the morning for males and in
the afternoon for females (105).
Fluoropyrimidine-related toxicity is strongly
affected by the activity of dihydropyrimidine dehydrogenase (DPD),
a 5-fluorouracil-metabolizing enzyme (106). Krugluger et al (45) revealed that disturbed transcription
of Per1 may be a cause of disrupted daily DPD oscillation in CRC
cells. Fang et al (107)
demonstrated that the expression levels of Cry2 are elevated in
chemoresistant CRC samples and knockdown of Cry2 increased
oxaliplatin sensitivity in CRC cells. Circadian delivery of
irinotecan to Caco-2 colon cancer cells indicated that
chronomodulated chemotherapy may be an optimized option (108). Hesse et al (109) established a mathematical model to
predict the impact of various parameters (e.g., BMAL1 degradation
rates, cytosolic BMAL1 degradation rates and CLOCK activation
rates) on irinotecan toxicity, but clinical evidence is required to
prove this hypothesis.
These findings indicated that the circadian clock
system may guide oncological treatments of CRC.
The present review outlines the connections between
the circadian clock and CRC progression (Fig. 3). Bowel physiology indicates a
circadian rhythm and molecular clockwork dysregulation may host the
colorectal carcinogenesis process. According to previously
published reports, circadian clock genes are related to CRC
progression, patient survival and response to chemotherapy.
Although the findings are not all consistent, it may be
hypothesized that this largely resulted from individual
differences, such as different ethnicities, gender and age.
Furthermore, multiple detection methods may also lead to different
findings. Based on the circadian rhythm system, chronotherapy may
be an optimal method for effective treatment, particularly to
increase the maximum tolerated doses and alleviate the toxicity of
chemotherapy agents. In addition, the oscillating circadian clock
regulates multiple cellular activities related to colorectal
carcinogenesis, such as metabolism, the cell cycle, DNA damage
response, EMT processes and stemness. Finally, it may be concluded
that the circadian clock is tightly correlated with CRC prognosis
and allows further advances in modern therapeutic approaches. The
relationship between the circadian clock and CRC warrants further
investigation.
Not applicable.
LL was involved in the conception and design of the
structure of the manuscript. XR then performed the literature
search, generated the figures and wrote the manuscript.
Not applicable.
Not applicable.
The authors declare that they have no competing
interests.
Not applicable.
This work was supported by grants from the National Natural
Science Foundation of China (grant no. 82104647), the Chinese
Postdoctoral Science Foundation (grant no. 2021M700964) and
Guangzhou Science and Technology Bureau (grant no.
201904010396).
|
1
|
Shafer OT, Levine JD, Truman JW and Hall
JC: Flies by night: Effects of changing day length on Drosophila's
circadian clock. Curr Biol. 14:424–432. 2004.PubMed/NCBI
|
|
2
|
Rusak B and Zucker I: Neural regulation of
circadian rhythms. Physiol Rev. 59:449–526. 1979. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Dibner C, Schibler U and Albrecht U: The
mammalian circadian timing system: Organization and coordination of
central and peripheral clocks. Annu Rev Physiol. 72:517–549. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Gekakis N, Staknis D, Nguyen HB, Davis FC,
Wilsbacher LD, King DP, Takahashi JS and Weitz CJ: Role of the
CLOCK protein in the mammalian circadian mechanism. Science.
280:1564–1569. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Antoch MP, Song EJ, Chang AM, Vitaterna
MH, Zhao Y, Wilsbacher LD, Sangoram AM, King DP, Pinto LH and
Takahashi JS: Functional identification of the mouse circadian
Clock gene by transgenic BAC rescue. Cell. 89:655–667. 1997.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Preitner N, Damiola F, Lopez-Molina L,
Zakany J, Duboule D, Albrecht U and Schibler U: The orphan nuclear
receptor REV-ERBalpha controls circadian transcription within the
positive limb of the mammalian circadian oscillator. Cell.
110:251–260. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Cho H, Zhao X, Hatori M, Yu RT, Barish GD,
Lam MT, Chong LW, DiTacchio L, Atkins AR, Glass CK, et al:
Regulation of circadian behaviour and metabolism by REV-ERB-α and
REV-ERB-β. Nature. 485:123–127. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Strahl BD and Allis CD: The language of
covalent histone modifications. Nature. 403:41–45. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Grunstein M: Histone acetylation in
chromatin structure and transcription. Nature. 389:349–352. 1997.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Doi M, Hirayama J and Sassone-Corsi P:
Circadian regulator CLOCK is a histone acetyltransferase. Cell.
125:497–508. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Hirayama J, Sahar S, Grimaldi B, Tamaru T,
Takamatsu K, Nakahata Y and Sassone-Corsi P: CLOCK-mediated
acetylation of BMAL1 controls circadian function. Nature.
450:1086–1090. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Nakahata Y, Kaluzova M, Grimaldi B, Sahar
S, Hirayama J, Chen D, Guarente LP and Sassone-Corsi P: The
NAD+-dependent deacetylase SIRT1 modulates CLOCK-mediated chromatin
remodeling and circadian control. Cell. 134:329–340. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Asher G, Gatfield D, Stratmann M, Reinke
H, Dibner C, Kreppel F, Mostoslavsky R, Alt FW and Schibler U:
SIRT1 regulates circadian clock gene expression through PER2
deacetylation. Cell. 134:317–328. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Sulli G, Lam MTY and Panda S: Interplay
between circadian clock and cancer: New frontiers for cancer
treatment. Trends Cancer. 5:475–494. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Erren TC, Morfeld P, Foster RG, Reiter RJ,
Groß JV and Westermann IK: Sleep and cancer: Synthesis of
experimental data and meta-analyses of cancer incidence among some
1,500,000 study individuals in 13 countries. Chronobiol Int.
33:325–350. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Papantoniou K, Devore EE, Massa J,
Strohmaier S, Vetter C, Yang L, Shi Y, Giovannucci E, Speizer F and
Schernhammer ES: Rotating night shift work and colorectal cancer
risk in the nurses' health studies. Int J Cancer. 143:2709–2717.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Shi Y, Liu L, Hamada T, Nowak JA,
Giannakis M, Ma Y, Song M, Nevo D, Kosumi K, Gu M, et al:
Night-shift work duration and risk of colorectal cancer according
to IRS1 and IRS2 expression. Cancer Epidemiol Biomarkers Prev.
29:133–140. 2020. View Article : Google Scholar :
|
|
18
|
Bishehsari F, Engen PA, Voigt RM, Swanson
G, Shaikh M, Wilber S, Naqib A, Green SJ, Shetuni B, Forsyth CB, et
al: Abnormal eating patterns cause circadian disruption and promote
alcohol-associated colon carcinogenesis. Cell Mol Gastroenterol
Hepatol. 9:219–237. 2020. View Article : Google Scholar :
|
|
19
|
Bray F, Ferlay J, Soerjomataram I, Siegel
RL, Torre LA and Jemal A: Global cancer statistics 2018: GLOBOCAN
estimates of incidence and mortality worldwide for 36 cancers in
185 countries. CA Cancer J Clin. 68:394–424. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Pelullo M, Nardozza F, Zema S, Quaranta R,
Nicoletti C, Besharat ZM, Felli MP, Cerbelli B, d'Amati G, Palermo
R, et al: Kras/ADAM17-dependent Jag1-ICD reverse signaling sustains
colorectal cancer progression and chemoresistance. Cancer Res.
79:5575–5586. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Afrăsânie VA, Marinca MV, Alexa-Stratulat
T, Gafton B, Păduraru M, Adavidoaiei AM, Miron L and Rusu C: KRAS,
NRAS, BRAF, HER2 and microsatellite instability in metastatic
colorectal cancer-practical implications for the clinician. Radiol
Oncol. 53:265–274. 2019. View Article : Google Scholar
|
|
22
|
Slik K, Turkki R, Carpén O, Kurki S,
Korkeila E, Sundström J and Pellinen T: CDX2 loss with
microsatellite stable phenotype predicts poor clinical outcome in
stage II colorectal carcinoma. Am J Surg Pathol. 43:1473–1482.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Wolpin BM and Mayer RJ: Systemic treatment
of colorectal cancer. Gastroenterology. 134:1296–1310. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Narducci F, Bassotti G, Gaburri M and
Morelli A: Twenty four hour manometric recording of colonic motor
activity in healthy man. Gut. 28:17–25. 1987. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Rao SS, Sadeghi P, Beaty J, Kavlock R and
Ackerson K: Ambulatory 24-h colonic manometry in healthy humans. Am
J Physiol Gastrointest Liver Physiol. 280:G629–G639. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Clench J, Reinberg A, Dziewanowska Z,
Ghata J and Smolensky M: Circadian changes in the bioavailability
and effects of indomethacin in healthy subjects. Eur J Clin
Pharmacol. 20:359–369. 1981. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Markiewicz A, Kamiński M, Chocilowski W,
Gomoluch T, Bołdys H and Skrzypek B: Circadian rhythms of four
marker enzymes activity of the jejunal villi in man. Acta
Histochem. 72:91–99. 1983. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Hoogerwerf WA, Shahinian VB, Cornélissen
G, Halberg F, Bostwick J, Timm J, Bartell PA and Cassone VM:
Rhythmic changes in colonic motility are regulated by period genes.
Am J Physiol Gastrointest Liver Physiol. 298:G143–G150. 2010.
View Article : Google Scholar :
|
|
29
|
Thaiss CA, Zeevi D, Levy M,
Zilberman-Schapira G, Suez J, Tengeler AC, Abramson L, Katz MN,
Korem T, Zmora N, et al: Transkingdom control of microbiota diurnal
oscillations promotes metabolic homeostasis. Cell. 159:514–529.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Sládek M, Rybová M, Jindráková Z, Zemanová
Z, Polidarová L, Mrnka L, O'Neill J, Pácha J and Sumová A: Insight
into the circadian clock within rat colonic epithelial cells.
Gastroenterology. 133:1240–1249. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Hoogerwerf WA, Hellmich HL, Cornélissen G,
Halberg F, Shahinian VB, Bostwick J, Savidge TC and Cassone VM:
Clock gene expression in the murine gastrointestinal tract:
Endogenous rhythmicity and effects of a feeding regimen.
Gastroenterology. 133:1250–1260. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Brandi G, Calabrese C, Pantaleo MA,
Morselli Labate A, Di Febo G, Hakim R, De Vivo A, Di Marco MC and
Biasco G: Circadian variations of rectal cell proliferation in
patients affected by advanced colorectal cancer. Cancer Lett.
208:193–196. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Yu F, Zhang T, Zhou C, Xu H, Guo L, Chen M
and Wu B: The circadian clock gene Bmal1 controls intestinal
exporter MRP2 and drug disposition. Theranostics. 9:2754–2767.
2019. View Article : Google Scholar :
|
|
34
|
Lévi F, Dugué PA, Innominato P, Karaboué
A, Dispersyn G, Parganiha A, Giacchetti S, Moreau T, Focan C,
Waterhouse J, et al: Wrist actimetry circadian rhythm as a robust
predictor of colorectal cancer patients survival. Chronobiol Int.
31:891–900. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Innominato PF, Focan C, Gorlia T, Moreau
T, Garufi C, Waterhouse J, Giacchetti S, Coudert B, Iacobelli S,
Genet D, et al: Circadian rhythm in rest and activity: A biological
correlate of quality of life and a predictor of survival in
patients with metastatic colorectal cancer. Cancer Res.
69:4700–4707. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Stokes K, Nunes M, Trombley C, Flôres
DEFL, Wu G, Taleb Z, Alkhateeb A, Banskota S, Harris C, Love OP, et
al: The circadian clock gene, Bmal1, regulates intestinal stem cell
signaling and represses tumor initiation. Cell Mol Gastroenterol
Hepatol. 12:1847–1872.e0. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
He A, Huang Z, Zhang R, Lu H, Wang J, Cao
J and Feng Q: Circadian clock genes are correlated with prognosis
and immune cell infiltration in colon adenocarcinoma. Comput Math
Methods Med. 2022:17099182022. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Karantanos T, Theodoropoulos G, Gazouli M,
Vaiopoulou A, Karantanou C, Lymberi M and Pektasides D: Expression
of clock genes in patients with colorectal cancer. Int J Biol
Markers. 28:280–285. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Mazzoccoli G, Panza A, Valvano MR, Palumbo
O, Carella M, Pazienza V, Biscaglia G, Tavano F, Di Sebastiano P,
Andriulli A and Piepoli A: Clock gene expression levels and
relationship with clinical and pathological features in colorectal
cancer patients. Chronobiol Int. 28:841–851. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Mostafaie N, Kállay E, Sauerzapf E, Bonner
E, Kriwanek S, Cross HS, Huber KR and Krugluger W: Correlated
downregulation of estrogen receptor beta and the circadian clock
gene Per1 in human colorectal cancer. Mol Carcinog. 48:642–647.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Orhan T, Nielsen PB, Hviid TVF, Rosen AW
and Gögenür I: Expression of circadian clock genes in human
colorectal cancer tissues using droplet digital PCR. Cancer Invest.
37:90–98. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Oshima T, Takenoshita S, Akaike M,
Kunisaki C, Fujii S, Nozaki A, Numata K, Shiozawa M, Rino Y, Tanaka
K, et al: Expression of circadian genes correlates with liver
metastasis and outcomes in colorectal cancer. Oncol Rep.
25:1439–1446. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Wu S, Fesler A and Ju J: Implications of
circadian rhythm regulation by microRNAs in colorectal cancer.
Cancer Transl Med. 2:1–6. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
No authors listed. Expression of PER, CRY,
and TIM genes for the pathological features of colorectal cancer
patients [Retraction]. Onco Targets Ther. 9:56992016. View Article : Google Scholar
|
|
45
|
Krugluger W, Brandstaetter A, Kállay E,
Schueller J, Krexner E, Kriwanek S, Bonner E and Cross HS:
Regulation of genes of the circadian clock in human colon cancer:
Reduced period-1 and dihydropyrimidine dehydrogenase transcription
correlates in high-grade tumors. Cancer Res. 67:7917–7922. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Lu H, Chu Q, Xie G, Han H, Chen Z, Xu B
and Yue Z: Circadian gene expression predicts patient response to
neoadjuvant chemo-radiation therapy for rectal cancer. Int J Clin
Exp Pathol. 8:10985–10994. 2015.
|
|
47
|
Nemeth C, Humpeler S, Kallay E, Mesteri I,
Svoboda M, Rögelsperger O, Klammer N, Thalhammer T and Ekmekcioglu
C: Decreased expression of the melatonin receptor 1 in human
colorectal adenocarcinomas. J Biol Regul Homeost Agents.
25:531–542. 2011.
|
|
48
|
Wang Y, Hua L, Lu C and Chen Z: Expression
of circadian clock gene human Period2 (hPer2) in human colorectal
carcinoma. World J Surg Oncol. 9:1662011. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Aroca-Siendones MI, Moreno-SanJuan S,
Puentes-Pardo JD, Verbeni M, Arnedo J, Escudero-Feliu J,
García-Costela M, García-Robles A, Carazo Á and León J: Core
circadian clock proteins as biomarkers of progression in colorectal
cancer. Biomedicines. 9:9672021. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Hasakova K, Vician M, Reis R, Zeman M and
Herichova I: Sex-dependent correlation between survival and
expression of genes related to the circadian oscillator in patients
with colorectal cancer. Chronobiol Int. 35:1423–1434. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Wang Y, Cheng Y, Yu G, Jia B, Hu Z and
Zhang L: Expression of PER, CRY, and TIM genes for the pathological
features of colorectal cancer patients. Onco Targets Ther.
9:1997–2005. 2016.PubMed/NCBI
|
|
52
|
Xiong Y, Zhuang Y, Zhong M, Qin W, Huang
B, Zhao J, Gao Z, Ma J, Wu Z, Hong X, et al: Period 2 suppresses
the malignant cellular behaviors of colorectal cancer through the
epithelial-mesenchymal transformation process. Cancer Control.
29:107327482210813692022. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Hasakova K, Reis R, Vician M, Zeman M and
Herichova I: Expression of miR-34a-5p is up-regulated in human
colorectal cancer and correlates with survival and clock gene PER2
expression. PLoS One. 14:e02243962019. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Wang X, Yan D, Teng M, Fan J, Zhou C, Li
D, Qiu G, Sun X, Li T, Xing T, et al: Reduced expression of PER3 is
associated with incidence and development of colon cancer. Ann Surg
Oncol. 19:3081–3088. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Alexander M, Burch JB, Steck SE, Chen CF,
Hurley TG, Cavicchia P, Ray M, Shivappa N, Guess J, Zhang H, et al:
Case-control study of the PERIOD3 clock gene length polymorphism
and colorectal adenoma formation. Oncol Rep. 33:935–941. 2015.
View Article : Google Scholar :
|
|
56
|
Momma T, Okayama H, Saitou M, Sugeno H,
Yoshimoto N, Takebayashi Y, Ohki S and Takenoshita S: Expression of
circadian clock genes in human colorectal adenoma and carcinoma.
Oncol Lett. 14:5319–5325. 2017.PubMed/NCBI
|
|
57
|
Štorcelová M, Vicián M, Reis R, Zeman M
and Herichová I: Expression of cell cycle regulatory factors hus1,
gadd45a, rb1, cdkn2a and mre11a correlates with expression of clock
gene per2 in human colorectal carcinoma tissue. Mol Biol Rep.
40:6351–6361. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Soták M, Polidarová L, Ergang P, Sumová A
and Pácha J: An association between clock genes and
clock-controlled cell cycle genes in murine colorectal tumors. Int
J Cancer. 132:1032–1041. 2013. View Article : Google Scholar
|
|
59
|
Bednarski JJ and Sleckman BP: At the
intersection of DNA damage and immune responses. Nat Rev Immunol.
19:231–242. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Ciccia A and Elledge SJ: The DNA damage
response: Making it safe to play with knives. Mol Cell. 40:179–204.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Gery S, Komatsu N, Baldjyan L, Yu A, Koo D
and Koeffler HP: The circadian gene per1 plays an important role in
cell growth and DNA damage control in human cancer cells. Mol Cell.
22:375–382. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Arango D, Mariadason JM, Wilson AJ, Yang
W, Corner GA, Nicholas C, Aranes MJ and Augenlicht LH: c-Myc
overexpression sensitises colon cancer cells to
camptothecin-induced apoptosis. Br J Cancer. 89:1757–1765. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Borgs L, Beukelaers P, Vandenbosch R,
Belachew S, Nguyen L and Malgrange B: Cell 'circadian' cycle: New
role for mammalian core clock genes. Cell Cycle. 8:832–837. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Wood PA, Yang X, Taber A, Oh EY, Ansell C,
Ayers SE, Al-Assaad Z, Carnevale K, Berger FG, Peña MM and
Hrushesky WJ: Period 2 mutation accelerates ApcMin/+ tumorigenesis.
Mol Cancer Res. 6:1786–1793. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Fu L, Pelicano H, Liu J, Huang P and Lee
C: The circadian gene Period2 plays an important role in tumor
suppression and DNA damage response in vivo. Cell. 111:41–50. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Shen P, Pichler M, Chen M, Calin GA and
Ling H: To Wnt or lose: The missing non-coding linc in colorectal
cancer. Int J Mol Sci. 18:20032017. View Article : Google Scholar :
|
|
67
|
Filipovich A, Gehrke I, Poll-Wolbeck SJ
and Kreuzer KA: Physiological inhibitors of Wnt signaling. Eur J
Haematol. 86:453–465. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Yang X, Wood PA, Ansell CM, Ohmori M, Oh
EY, Xiong Y, Berger FG, Peña MM and Hrushesky WJ: Beta-catenin
induces beta-TrCP-mediated PER2 degradation altering circadian
clock gene expression in intestinal mucosa of ApcMin/+ mice. J
Biochem. 145:289–297. 2009. View Article : Google Scholar
|
|
69
|
Schroll MM, LaBonia GJ, Ludwig KR and
Hummon AB: Glucose restriction combined with autophagy inhibition
and chemotherapy in HCT 116 spheroids decreases cell clonogenicity
and viability regulated by tumor suppressor genes. J Proteome Res.
16:3009–3018. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Zhang F, Sun H, Zhang S, Yang X, Zhang G
and Su T: Overexpression of PER3 inhibits self-renewal capability
and chemoresistance of colorectal cancer stem-like cells via
inhibition of notch and β-catenin signaling. Oncol Res. 25:709–719.
2017. View Article : Google Scholar
|
|
71
|
Wang JL, Lin YW, Chen HM, Kong X, Xiong H,
Shen N, Hong J and Fang JY: Calcium prevents tumorigenesis in a
mouse model of colorectal cancer. PLoS One. 6:e225662011.
View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Hasakova K, Vician M, Reis R, Zeman M and
Herichova I: The expression of clock genes cry1 and cry2 in human
colorectal cancer and tumor adjacent tissues correlates differently
dependent on tumor location. Neoplasma. 65:986–992. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Yu H, Meng X, Wu J, Pan C, Ying X, Zhou Y,
Liu R and Huang W: Cryptochrome 1 overexpression correlates with
tumor progression and poor prognosis in patients with colorectal
cancer. PLoS One. 8:e616792013. View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Mazzoccoli G, Colangelo T, Panza A, Rubino
R, De Cata A, Tiberio C, Valvano MR, Pazienza V, Merla G, Augello
B, et al: Deregulated expression of cryptochrome genes in human
colorectal cancer. Mol Cancer. 15:62016. View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Heald R, McLoughlin M and McKeon F: Human
wee1 maintains mitotic timing by protecting the nucleus from
cytoplasmically activated Cdc2 kinase. Cell. 74:463–474. 1993.
View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Backert S, Gelos M, Kobalz U, Hanski ML,
Böhm C, Mann B, Lövin N, Gratchev A, Mansmann U, Moyer MP, et al:
Differential gene expression in colon carcinoma cells and tissues
detected with a cDNA array. Int J Cancer. 82:868–874. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
77
|
van der Horst GT, Muijtjens M, Kobayashi
K, Takano R, Kanno S, Takao M, de Wit J, Verkerk A, Eker AP, van
Leenen D, et al: Mammalian Cry1 and Cry2 are essential for
maintenance of circadian rhythms. Nature. 398:627–630. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Burgermeister E, Battaglin F, Eladly F, Wu
W, Herweck F, Schulte N, Betge J, Härtel N, Kather JN, Weis CA, et
al: Aryl hydrocarbon receptor nuclear translocator-like
(ARNTL/BMAL1) is associated with bevacizumab resistance in
colorectal cancer via regulation of vascular endothelial growth
factor A. EBioMedicine. 45:139–154. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
79
|
Zhang Y, Devocelle A, Desterke C, de Souza
LEB, Hadadi É, Acloque H, Foudi A, Xiang Y, Ballesta A, Chang Y and
Giron-Michel J: BMAL1 knockdown leans epithelial-mesenchymal
balance toward epithelial properties and decreases the
chemoresistance of colon carcinoma cells. Int J Mol Sci.
22:52472021. View Article : Google Scholar : PubMed/NCBI
|
|
80
|
Wang L, Chen B, Wang Y, Sun N, Lu C, Qian
R and Hua L: hClock gene expression in human colorectal carcinoma.
Mol Med Rep. 8:1017–1022. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
81
|
Wang Y, Sun N, Lu C, Bei Y, Qian R and Hua
L: Upregulation of circadian gene 'hClock' contribution to
metastasis of colorectal cancer. Int J Oncol. 50:2191–2199. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
82
|
Karantanos T, Theodoropoulos G, Gazouli M,
Vaiopoulou A, Karantanou C, Stravopodis DJ, Bramis K, Lymperi M and
Pektasidis D: Association of the clock genes polymorphisms with
colorectal cancer susceptibility. J Surg Oncol. 108:563–567. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
83
|
Kurzawski G, Suchy J, Debniak T, Kładny J
and Lubiński J: Importance of microsatellite instability (MSI) in
colorectal cancer: MSI as a diagnostic tool. Ann Oncol. 15(Suppl
4): iv283–iv284. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
84
|
Alhopuro P, Björklund M, Sammalkorpi H,
Turunen M, Tuupanen S, Biström M, Niittymäki I, Lehtonen HJ,
Kivioja T, Launonen V, et al: Mutations in the circadian gene CLOCK
in colorectal cancer. Mol Cancer Res. 8:952–960. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
85
|
Fuhr L, El-Athman R, Scrima R, Cela O,
Carbone A, Knoop H, Li Y, Hoffmann K, Laukkanen MO, Corcione F, et
al: The circadian clock regulates metabolic phenotype rewiring via
HKDC1 and modulates tumor progression and drug response in
colorectal cancer. EBioMedicine. 33:105–121. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
86
|
Nelson RL: Iron and colorectal cancer
risk: Human studies. Nutr Rev. 59:140–148. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
87
|
Osborne NJ, Gurrin LC, Allen KJ,
Constantine CC, Delatycki MB, McLaren CE, Gertig DM, Anderson GJ,
Southey MC, Olynyk JK, et al: HFE C282Y homozygotes are at
increased risk of breast and colorectal cancer. Hepatology.
51:1311–1318. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
88
|
Okazaki F, Matsunaga N, Okazaki H, Azuma
H, Hamamura K, Tsuruta A, Tsurudome Y, Ogino T, Hara Y, Suzuki T,
et al: Circadian clock in a mouse colon tumor regulates
intracellular iron levels to promote tumor progression. J Biol
Chem. 291:7017–7028. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
89
|
Sakamoto W and Takenoshita S:
Overexpression of both clock and BMAL1 inhibits entry to S phase in
human colon cancer cells. Fukushima J Med Sci. 61:111–124. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
90
|
Zeng ZL, Wu MW, Sun J, Sun YL, Cai YC,
Huang YJ and Xian LJ: Effects of the biological clock gene Bmal1 on
tumour growth and anti-cancer drug activity. J Biochem.
148:319–326. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
91
|
Zhang Y, Devocelle A, Souza L, Foudi A,
Tenreira Bento S, Desterke C, Sherrard R, Ballesta A, Adam R,
Giron-Michel J and Chang Y: BMAL1 knockdown triggers different
colon carcinoma cell fates by altering the delicate equilibrium
between AKT/mTOR and P53/P21 pathways. Aging (Albany NY).
12:8067–8083. 2020. View Article : Google Scholar
|
|
92
|
Dong P, Wang Y, Liu Y, Zhu C, Lin J, Qian
R, Hua L and Lu C: BMAL1 induces colorectal cancer metastasis by
stimulating exosome secretion. Mol Biol Rep. 49:373–384. 2022.
View Article : Google Scholar
|
|
93
|
Gu D, Li S, Ben S, Du M, Chu H, Zhang Z,
Wang M, Zhang ZF and Chen J: Circadian clock pathway genes
associated with colorectal cancer risk and prognosis. Arch Toxicol.
92:2681–2689. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
94
|
Pazienza V, Piepoli A, Panza A, Valvano
MR, Benegiamo G, Vinciguerra M, Andriulli A and Mazzoccoli G: SIRT1
and the clock gene machinery in colorectal cancer. Cancer Invest.
30:98–105. 2012. View Article : Google Scholar
|
|
95
|
Colangelo T, Carbone A, Mazzarelli F,
Cuttano R, Dama E, Nittoli T, Albanesi J, Barisciano G, Forte N,
Palumbo O, et al: Loss of circadian gene timeless induces EMT and
tumor progression in colorectal cancer via Zeb1-dependent
mechanism. Cell Death Differ. 29:1552–1568. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
96
|
Xue X, Liu F, Han Y, Li P, Yuan B, Wang X,
Chen Y, Kuang Y, Zhi Q and Zhao H: Silencing NPAS2 promotes cell
growth and invasion in DLD-1 cells and correlated with poor
prognosis of colorectal cancer. Biochem Biophys Res Commun.
450:1058–1062. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
97
|
Yang X, Wood PA and Hrushesky WJ:
Mammalian TIMELESS is required for ATM-dependent CHK2 activation
and G2/M checkpoint control. J Biol Chem. 285:3030–3034. 2010.
View Article : Google Scholar :
|
|
98
|
Neilsen BK, Frodyma DE, McCall JL, Fisher
KW and Lewis RE: ERK-mediated TIMELESS expression suppresses G2/M
arrest in colon cancer cells. PLoS One. 14. pp. e2092242019,
View Article : Google Scholar
|
|
99
|
Pruitt K, Zinn RL, Ohm JE, McGarvey KM,
Kang SH, Watkins DN, Herman JG and Baylin SB: Inhibition of SIRT1
reactivates silenced cancer genes without loss of promoter DNA
hypermethylation. PLoS Genet. 2:e402006. View Article : Google Scholar : PubMed/NCBI
|
|
100
|
Levi F, Perpoint B, Garufi C, Focan C,
Chollet P, Depres-Brummer P, Zidani R, ienza S, Itzhaki M,
Iacobelli S, et al: Oxaliplatin activity against metastatic
colorectal cancer. A phase II study of 5-day continuous venous
infusion at circadian rhythm modulated rate. Eur J Cancer.
29A:1280–1284. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
101
|
Lévi FA, Zidani R, Vannetzel JM, Perpoint
B, Focan C, Faggiuolo R, Chollet P, Garufi C, Itzhaki M, Dogliotti
L, et al: Chronomodulated versus fixed-infusion-rate delivery of
ambulatory chemotherapy with oxaliplatin, fluorouracil, and folinic
acid (leucovorin) in patients with colorectal cancer metastases: A
randomized multi-institutional trial. J Natl Cancer Inst.
86:1608–1617. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
102
|
Lévi F, Karaboué A, Gorden L, Innominato
PF, Saffroy R, Giacchetti S, Hauteville D, Guettier C, Adam R and
Bouchahda M: Cetuximab and circadian chronomodulated chemotherapy
as salvage treatment for metastatic colorectal cancer (mCRC):
Safety, efficacy and improved secondary surgical resectability.
Cancer Chemother Pharmacol. 67:339–348. 2011. View Article : Google Scholar
|
|
103
|
Innominato PF, Giacchetti S, Moreau T,
Smaaland R, Focan C, Bjarnason GA, Garufi C, Iacobelli S,
Tampellini M, Tumolo S, et al: Prediction of survival by
neutropenia according to delivery schedule of
oxaliplatin-5-fluorouracil-leucovorin for metastatic colorectal
cancer in a randomized international trial (EORTC 05963).
Chronobiol Int. 28:586–600. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
104
|
Innominato PF, Karaboué A, Focan C,
Chollet P, Giacchetti S, Bouchahda M, Ulusakarya A, Torsello A,
Adam R, Lévi FA and Garufi C: Efficacy and safety of
chronomodulated irinotecan, oxaliplatin, 5-fluorouracil and
leucovorin combination as first- or second-line treatment against
metastatic colorectal cancer: Results from the international EORTC
05011 trial. Int J Cancer. 148:2512–2521. 2020.Epub ahead of print.
View Article : Google Scholar
|
|
105
|
Innominato PF, Ballesta A, Huang Q, Focan
C, Chollet P, Karaboué A, Giacchetti S, Bouchahda M, Adam R, Garufi
C and Lévi FA: Sex-dependent least toxic timing of irinotecan
combined with chronomodulated chemotherapy for metastatic
colorectal cancer: Randomized multicenter EORTC 05011 trial. Cancer
Med. 9:4148–4159. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
106
|
Henricks LM, Opdam FL, Beijnen JH, Cats A
and Schellens JHM: DPYD genotype-guided dose individualization to
improve patient safety of fluoropyrimidine therapy: Call for a drug
label update. Ann Oncol. 28:2915–2922. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
107
|
Fang L, Yang Z, Zhou J, Tung JY, Hsiao CD,
Wang L, Deng Y, Wang P, Wang J and Lee MH: Circadian clock gene
CRY2 degradation is involved in chemoresistance of colorectal
cancer. Mol Cancer Ther. 14:1476–1487. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
108
|
Ballesta A, Dulong S, Abbara C, Cohen B,
Okyar A, Clairambault J and Levi F: A combined experimental and
mathematical approach for molecular-based optimization of
irinotecan circadian delivery. PLoS Comput Biol. 7:e10021432011.
View Article : Google Scholar : PubMed/NCBI
|
|
109
|
Hesse J, Martinelli J, Aboumanify O,
Ballesta A and Relógio A: A mathematical model of the circadian
clock and drug pharmacology to optimize irinotecan administration
timing in colorectal cancer. Comput Struct Biotechnol J.
19:5170–5183. 2021. View Article : Google Scholar : PubMed/NCBI
|