Osteosarcoma (OS) is a frequently occurring primary
bone tumor in children, adolescents and young adults with a
worldwide incidence rate of 3.4 cases per 10,000 individuals per
year (1,2). OS commonly occurs in the metaphysis
of long bones, mostly in the humerus, distal femur and proximal
tibia, and is characterized by swelling and persistent pain in the
affected bone (3). Although the
combination of radical surgery and systemic chemotherapy improves
the prognosis of patients with primary OS, its side effects
significantly affect patients' quality of life. Moreover, the
prognosis of metastatic or recurrent OS remains unsatisfactory, as
the recurrence and/or metastasis rate of OS is >30%.
Additionally, in certain sarcomas such as early stage uterine
leiomyosarcoma, adjuvant chemotherapy (with or without
radiotherapy) has shown no significant decrease in recurrence rates
(4). The resistance of tumor
cells to both radiotherapy and chemotherapy leads to a 5-year
overall survival rate of <25% in patients with metastatic or
recurrent OS (5). Rizzo et
al (6) suggested that a more
in-depth biological characterization was essential to comprehend
the molecular biology of sarcomas and to improve identification of
patients who could benefit from adjuvant therapy. Immunotherapies
such as immune checkpoint inhibitors have been developed as
groundbreaking treatments for patients with cancer (7). More recently, chimeric antigen
receptor T (CAR-T) cell therapy, which is known as an innovative
form of immunotherapy, has garnered significant attention.
CAR-T cell therapy has become a groundbreaking
treatment for hematological malignancies and multiple myeloma.
However, recent research has focused on extending the use of CAR-T
therapy to treat OS, which is a solid tumor and different from
blood tumors. Although CAR-T cells have excellent effectiveness in
hematological tumors and have been approved for use in
hematological tumors, their use in the treatment of OS remains in
its infancy, and there are still problems such as difficult homing
and colonization of CAR-T cells, off-target effects, immune escape,
immunosuppressive tumor microenvironment (TME) and cytokine release
syndrome (CRS) (18). To address
these issues, previous studies have tried a variety of CAR-T cells
to treat OS and have obtained favorable results, for example,
increased elimination of OS and reduced incidence of adverse
effects (19-23).
The present study introduced in detail the structure
of CAR-T cells and the influence of changing their structure on the
therapeutic effect of CAR-T cells, which should help physicians to
design CAR-T cells for optimal therapeutic effect (19,20). In addition, the current study
provided an overview of all target antigens that can be used to
treat OS and the results of the corresponding CAR-T cells (21-31). Compared with other reviews on
CAR-T cell therapy for OS published in the literature, the present
review described in detail the challenges that CAR-T cell therapy
faces (32,33), the ways to solve these problems
(34-41), and the advantages of combining
radiotherapy (42), chemotherapy
(43) and chemoradiotherapy
(44), so that bone oncologists
can have a more comprehensive understanding of CAR-T cell for
OS.
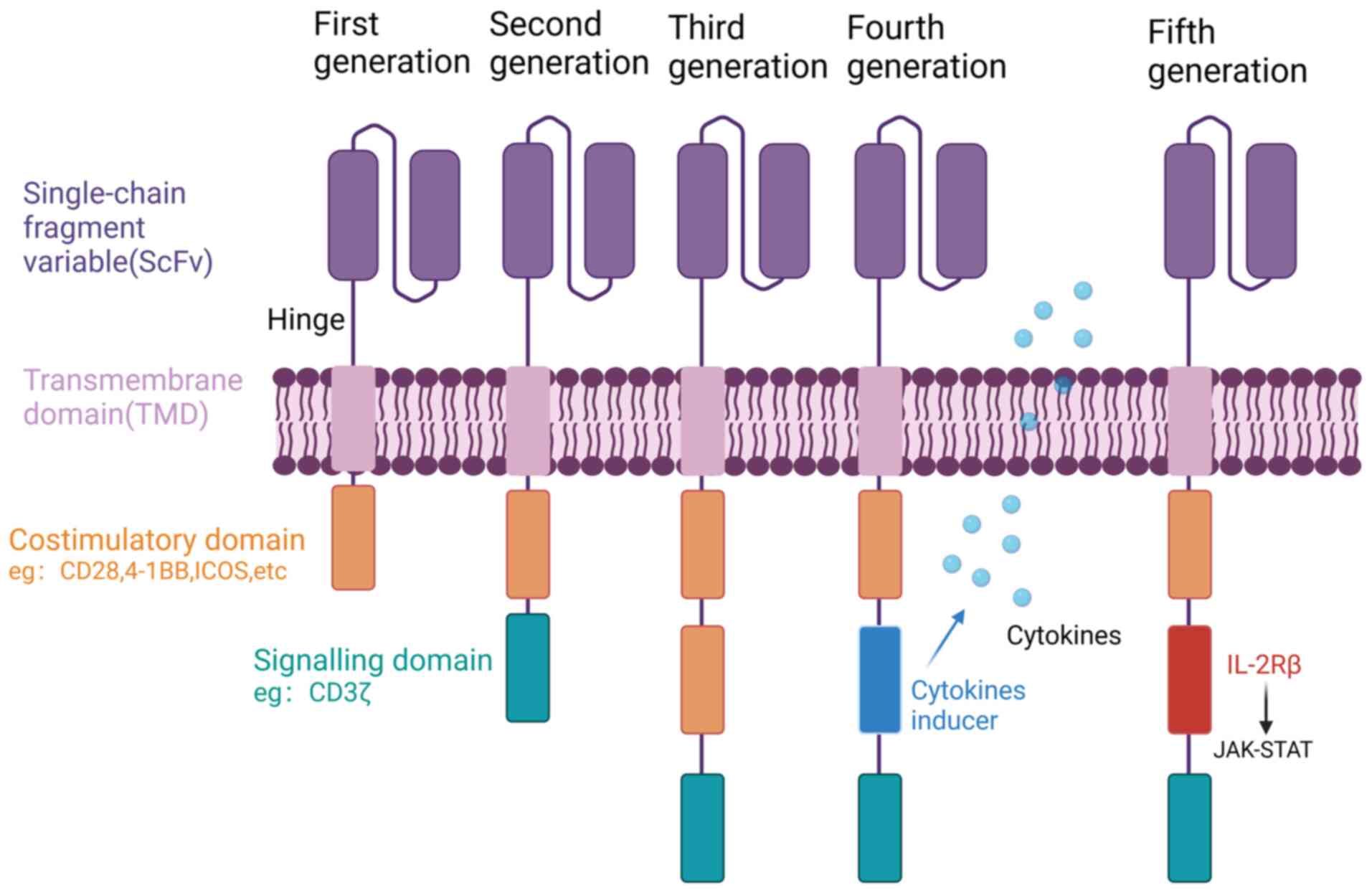
CAR-T cell therapy is a novel form of immunotherapy
that utilizes genetic engineering techniques to modify and boost a
patient's own immune cells outside the body (45). The therapy involves the use of a
genetically engineered chimeric receptor called CAR, which is
composed of various components, including an extracellular
single-chain fragment variable (scFv), a hinge domain (HD), a
transmembrane domain (TMD) and an intracellular domain (Fig. 1). The scFv is responsible for
recognizing specific antigens on the surface of tumor cells, while
the HD serves as a bridge between the scFv and TMD. The TMD anchors
CAR to the membranes of T cells and promotes signal transduction
into the cells, while the intracellular domain is responsible for T
cell activation (19).
First-generation CARs comprise a scFv and an intracellular CD3ζ
activation domain. However, they have inadequate antitumor activity
and proliferative capacity due to the absence of costimulatory
signaling. To address this limitation, second-generation CARs have
been developed. These incorporate an additional costimulatory
domain, for example CD28, 4-1BB, OX40 or inducible costimulator
(ICOS), which enhances their ability to proliferate and release a
greater number of cytokines. At present, the marketable CAR-T cell
products predominantly employ second-generation CAR structures.
However, there have been advancements in the development of
third-generation CARs, which incorporate two different
costimulatory molecules, including CD28 and 4-1BB, to enhance their
efficacy (46). Furthermore,
fourth-generation CARs have emerged, which besides costimulatory
domains, contain domains that regulate cytokine release. These
cytokines, including interleukin (IL)-12, IL-18, IL-21 and IL-23,
have the function of improving the TME and promoting the production
of memory T cells, leading to increased persistence of CAR-T cells
in vivo (47).
Fifth-generation CARs incorporate IL-2Rβ membrane receptors, which
offer a docking site for signal transducer and activator of
transcription 3 (STAT3) and trigger the activation of a Janus
kinase (JAK)-STAT signaling domain. Additionally, these advanced
CARs exhibit a combined activation of triple signals originating
from the CD3ζ, costimulatory domain and the cytokine-inducing
JAK-STAT3/5 pathway (48). CARs
and T cells are assembled by lentiviral or other vectors to form
CAR-T cells. When CAR-T cells specifically recognize the target
antigen, the T cells are activated via the intracellular STD and
play a tumor cell-eliminating role (45).
Traditional immunotherapies include monoclonal
antibodies, tumor vaccines, oncolytic viruses, immune checkpoint
inhibitors, and adoptive transfer of activated T cells and natural
killer (NK) cells in vitro (49). Upon contact with tumor antigens, B
cells produce highly homogeneous monoclonal antibodies that target
only specific antigenic epitopes. Binding of the monoclonal
antibodies to their target antigen can activate the complement
system to promote the triggering of antibody-dependent cytotoxicity
(50,51). Compared with monoclonal
antibodies, CAR-T cells possess numerous advantages: Firstly, CAR-T
cells can recognize tumor cells even when the expression of the
targeted antigen is low, whereas monoclonal antibodies cannot do
so; secondly, CAR-T cells are not only able to eliminate tumor
cells directly, but also secrete cytokines such as IFN-γ, which
enhance their antitumor effect; and thirdly, CAR-T cells are able
to proliferate and maintain a long-lasting antitumor effect after
infusion (52,53).
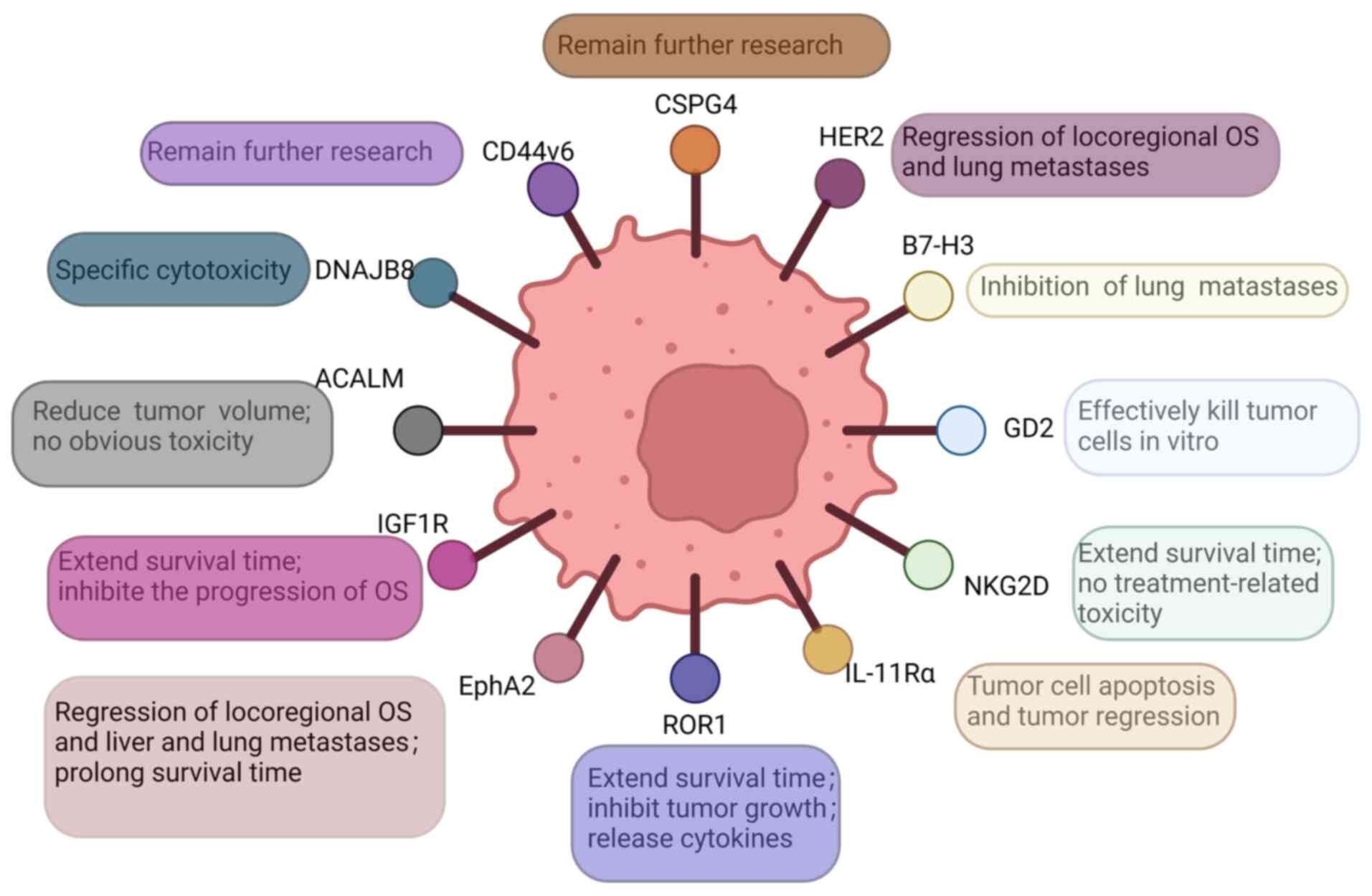
Several target antigens for the treatment of OS have
been reported. In the following sections, some of these antigens
are presented, including DnaJ homolog subfamily B member 8
(DNAJB8), human epidermal growth factor receptor 2 (HER2), CD276,
activated leukocyte cell adhesion molecule (ALCAM, also known as
CD166), EphA2, GD2, IL-11 receptor α chain (IL-11Rα), insulin-like
growth factor type I receptor (IGF-1R), receptor tyrosine
kinase-like orphan receptor 1 (ROR1), NK group 2D (NKG2D),
chondroitin sulfate proteoglycan 4 (CSPG4) and CD44v6 (Fig. 2).
DNAJB8 belongs to the heat shock protein 40 family.
In health tissues, DNAJB8 is expressed only in the testis. In tumor
tissues, DNAJB8 is preferentially expressed in cancer stem cells
and has the function of cancer stem cell maintenance. Cancer stem
cells can promote tumorigenesis, self-renewal and differentiation.
Previous studies found that the overexpression of DNAJB8 increased
the percentage of cells in the population representing cancer stem
cells in renal cell carcinoma, and enhanced their ability to
initiate tumors (21,54,55). Morita et al (21) proposed that DNAJB8-specific
cytotoxic T lymphocytes had higher eliminating activity against
colorectal cancer stem cells and exhibited antitumor effects.
Watanabe et al (56)
successfully developed second-generation CARs targeting a peptide
complex derived from DNAJB8, and confirmed that DNAJB8-CAR-T cells
showed a target eliminating effect on OS cells, indicating that
DNAJB8 could serve as a potential and promising target antigen of
CAR-T cell for OS.
HER 2 is a member of the epidermal growth factor
receptor family, and plays an important role in cell proliferation
(57). HER 2 is unique among the
epidermal growth factor receptor family members due to its role in
regulating cell proliferation and differentiation. Abnormalities in
HER2 expression or function have been implicated in various
malignancies, such as OS, where HER2 overexpression is associated
with aggressive tumor growth. In the majority of OS cases, HER2
positivity is observed regardless of histological grade, and Abdou
et al (58) indicated that
positive expression of HER2 was significantly correlated with
high-grade OS. Ahmed et al (52) demonstrated that HER2-CAR-T cells
could eliminate HER2+ OS cells. A phase I/II clinical
trial by Ahmed et al (22)
revealed that patients with cancer could be transfused with safe
doses of HER2-CAR-T cells that could reach the tumor and maintain
it at low levels for >6 weeks in a dose-dependent manner. Thus,
according to the aforementioned results, HER2 has been demonstrated
to be an effective target antigen in CAR T-cell therapy for OS.
CD276 is an important immune checkpoint molecule
belonging to the B7 family. It plays an essential role in
modulating immune responses by exerting inhibitory effects on the
activation, proliferation and cytokine production of T cells.
Overexpression of CD276 is commonly observed in various cancer
types, including OS, melanoma, Ewing sarcoma, non-small cell lung
cancer and rhabdomyosarcoma (23). Picarda et al (23) reported that 91.8% of tumor cells
in OS tissues expressed CD276, whereas the expression of CD276 in
normal tissues was low. Accordingly, CD276 may be a potential
target for the treatment of OS. CD276-CAR-T cells induced the
regression of esophageal cancer and prolonged the survival time of
mice, in addition to showing significant antitumor effects in
radiotherapy-resistant prostate adenocarcinoma, ovarian cancer,
pancreatic adenocarcinoma, atypical teratoid/rhabdoid tumor and
neuroblastoma models (59-62).
Previous studies have reported that CD276-CAR-T cells exhibit a
dose-dependent antitumor activity and inhibit the development of
spontaneous lung metastases in a mouse model of OS (63,64). Since CD276 has T-cell inhibitory
effects, anti-CD276 CAR-T cells are a promising immunotherapy for
the treatment of OS.
CD166 belongs to the immunoglobulin superfamily and
plays a role in a variety of biological activities, including
neuronal growth, hematopoiesis and inflammatory responses (65). CD166 has been found in primary OS
biopsy specimens, where its expression level is high (66). Flow cytometric analysis indicated
that the expression of CD166 on OS cell lines varied and ranged
from 36.9 to 96.7%, which suggested that CD166-CAR-T cells could be
a promising treatment approach for OS (24). CD166-CAR-T cells exhibit complete
activation upon encountering CD166+ OS cells,
effectively eliminating OS cell lines in vitro.
Encouragingly, when CD166-CAR-T cells were infused into mice, there
was a notable regression of tumors without causing significant
toxicity (24). This promising
outcome highlights the potential of CD166-CAR-T cell therapy as a
safe and effective method for treating OS. In addition, CD166-CAR-T
cells showed strong cytotoxicity against colorectal cancer stem
cells in vitro, thus they may be an effective method for the
clinical treatment of colorectal cancer (67). According to the aforementioned
studies, CD166 appears to be an essential target antigen in the
treatment of OS.
During embryonic development, EphA2 serves as a
tyrosine kinase receptor for Eph signaling. However, in the later
stages, EphA2 expression becomes predominantly limited to specific
epithelial cells (68).
Overexpression of EphA2 has been frequently detected in various
cancer types, including OS, breast cancer, ovarian cancer and
prostate adenocarcinoma (69,70). Compared with those in normal bone
tissue, EphA2 protein levels were significantly elevated in OS bone
tissue. Fritsche-Guenther et al (71) conducted a study where a
correlation between the overexpression of EphA2 and the development
of OS was observed. In an animal model of implanted OS, EphA2-CAR-T
cells successfully eliminated EPHA2-overexpressing OS cells in
immunodeficient mice and significantly prolonged mouse survival.
Moreover, in animal models of metastatic OS, intravenously injected
EphA2-CAR-T cells could target the tumors, and effectively
eliminated OS liver and lung metastases (25). Based on the findings from the
aforementioned studies, it can be concluded that EphA2 holds great
potential as an effective and crucial target antigen for OS.
Gangliosides are glycated lipid molecules belonging
to the sphingolipid group, which have the function of mediating
tumor cell adhesion to extracellular matrix proteins and in
directing signaling during cell death. However, their role in
tumorigenesis remains unknown (72). GD2 expression has been observed to
vary in tumor tissues among patients with sarcoma, central nervous
system tumors and lung cancer (73). GD2-CAR-T cells have been revealed
to exhibit antigen-dependent cytotoxicity against GD2-expressing
lung cancer models both in vitro and in vivo
(26). Long et al
(74) found that third-generation
GD2-CAR-T cells exhibited significant effectiveness in eliminating
GD2+ OS cell lines in laboratory conditions, but showed
limited efficacy in eliminating OS in vivo. Nevertheless,
the phase I clinical trial (NCT02107963) has demonstrated the
safety of increasing doses of third-generation GD2-CAR-T cells in
patients with GD2+ OS (75), and bispecific antibodies targeting
GD2 and HER2 have potent antitumor effects against OS in
vitro and in vivo (76). According to the aforementioned
studies, it can be concluded that GD2 holds promise as a valuable
and effective target antigen for OS.
Previous studies have demonstrated that the
expression of IL-11Rα on the cell surface is linked to unfavorable
patient prognosis. IL-11Rα is activated by its ligand IL-11, which
can activate multiple signaling cascades in its target cells and is
considered an important regulator of bone homeostasis. IL-11 and
IL-11R are both expressed in OS in situ and in lung
metastases, and are markers of high-risk OS, while the expression
of IL-11Rα is absent in neighboring health lung tissue (77,78). IL-11Rα-CAR-T cells are able to
effectively eliminate OS cells in vitro, and have been
observed to specifically target and accumulate in OS lung
metastases following intravenous administration, while
demonstrating no accumulation in the surrounding health lung
tissue. The utilization of IL-11Rα-CAR-T cells for the treatment of
OS led to notable apoptosis and regression of tumors in mice with
lung metastases (27). These
findings suggested that IL-11Rα serves as a promising, effective
and crucial target antigen for OS.
IGF-1 is a hormone with an insulin-like molecular
structure, and overexpression of IGF-1 and its receptor (IGF-1R)
has been associated with cancer development. At OS, elevated levels
of IGF-1 and IGF-1R contribute to cancer development by promoting
transformation, proliferation and metastasis, and by reducing
susceptibility to apoptosis. Overexpression of IGF-1/IGF-1R also
contributes to tumor cell survival and resistance to chemotherapy
(79). It has been demonstrated
that chemotherapy-resistant OS cell lines and primary OS tissues
exhibit high expression of IGF-1R (80). Moreover, the application of
IGF1R-CAR-T cells has been found to effectively impede the
advancement of both systemic and localized OS in immunodeficient
mice, leading to a significant extension in animal survival time
(28). MicroRNA (miRNA or
miR)-26a, miR-100 and miRNA-133a inhibit OS cell proliferation,
migration and invasion, and increase tumor cell sensitivity to
chemotherapy by targeting IGF1-R (81-83). In accordance with the results of
the aforementioned studies, IGF1-R is a potential, effective and
important target antigen for OS.
ROR1, a transmembrane tyrosine kinase receptor,
holds great importance in facilitating cancer cell migration,
invasion and metastasis. Notably, elevated levels of ROR1 have been
detected in both hematological malignancies and solid tumors, such
as OS, Ewing sarcoma and rhabdomyosarcoma. By contrast, ROR1 is not
expressed in normal adult tissues (28,84). Previous studies have shown that
anti-ROR1 monoclonal antibodies significantly block the metastasis
of OS cells, and ROR1-CAR-T cells exhibit specific cytotoxicity
against ROR1+ OS tissues and release sarcoma-specific
cytokines such as interferon (IFN)-γ, tumor necrosis factor (TNF)-α
and IL-13 in vitro, while significantly inhibiting local and
disseminated sarcoma growth. Furthermore, the administration of
ROR1-CAR-T cells has demonstrated promising results in extending
the survival time of local sarcoma models, suggesting that
ROR1-CAR-T cell therapy holds potential as a management option for
high-risk sarcomas (28,85). Consequently, CAR-T cells
engineered by anti-ROR1 antigens are an effective measure for the
treatment of OS.
NKG2D is a receptor responsible for activating NK
cells. NKG2D ligand (NKG2DL) is expressed in various
immunosuppressive cells present in the TME. This expression of
NKG2DL enables tumor cells to evade immune surveillance by evading
detection and elimination by NK and T cells. It has been reported
that the binding of NKG2DL on tumor cells to NKG2D in NK cells
could lead to cell-mediated cytotoxicity and destruction of target
cells (29,86,87). NKG2D-CAR-T cell therapy has been
investigated in phase II clinical trials (NCT02203825) for the
treatment of hematological malignancies, and has shown promising
results with no major safety concerns raised among the participants
(88). Moreover, NKG2D-CAR-T
cells exhibited an NKG2D-dependent mechanism to target solid
tumors, including OS, and stomach, liver and cervical cancer, and
demonstrated a potent antitumor activity (89-91). Fernández et al (29) indicated that NKG2D-CAR-T cells had
a strong anti-OS activity. Thus, NKG2D holds great potential as an
effective target antigen for OS.
CSPG4 is a cell surface proteoglycan that is
under-expressed in healthy tissues but overexpressed in tumor cells
and TME. It is involved in tumor progression by regulating
intracellular signaling pathways related to tumor cell adhesion and
migration. Several studies have reported that CSPG4 mediates
multidrug resistance by stimulating the PI3K/AKT signaling pathway,
which plays a critical role in promoting tumor cell proliferation
and survival (30,92,93). The results of those studies
identified that overexpression of CSPG4 was associated with shorter
survival in patients with OS. Anti-CSPG4 monoclonal antibodies
significantly inhibited the proliferation and migration of
CSPG4+ OS cells and improved the efficacy of doxorubicin
(30). Considering the results of
the aforementioned studies, CSPG4 appears to be a potential,
effective and important target antigen of CAR-T cells in the
treatment of OS. However, clinical trials should be carried out to
further assess their effectiveness.
CD44 is a transmembrane glycoprotein that can be
expressed in its standard form (CD44s) and a variety of variants
(CD44v). One of these variants is CD44v6, which regulates the
extracellular matrix and inhibits tumor cell apoptosis (94,95). CD44v6 is expressed in several
malignancies, including colorectal and breast cancer (96,97). A previous meta-analysis showed
that overexpression of CD44v6 was associated with lower survival
and metastasis in patients with OS (31). Nakajima et al (98) found that CD44v6 could be expressed
in metastatic OS and was a protein of bone tumors. Although there
are no studies in the literature focused on treating OS by
targeting CD44v6, based on the outcomes of the aforementioned
studies, CD44v6 appears to be a promising potential OS target
antigen.
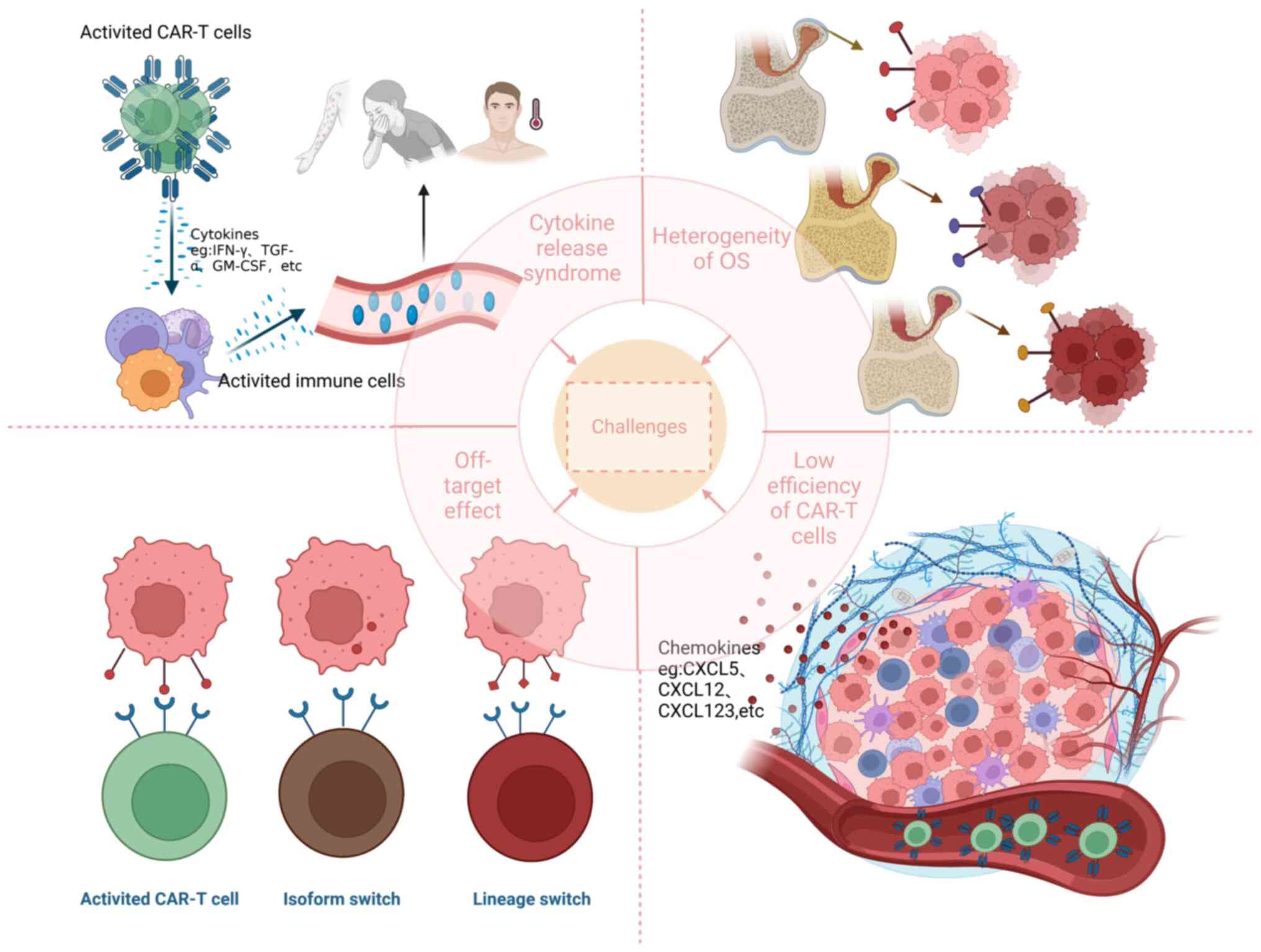
At present, CAR-T cell therapy for OS encounters
several challenges, including off-target effect, inhibitory TME,
CRS, low efficiency of CAR-T cells and heterogeneity of OS
(Fig. 3).
Tumor antigens can be classified into two types: i)
Tumor-specific antigens (TSAs) and ii) tumor-associated antigens
(TAAs) (99). TSAs are unique to
cancer cells and are not found in normal healthy cells, which are
ideal targets for immunotherapy, as they can be specifically
targeted by the immune system without affecting normal cells.
However, the expression of TSAs on the surface of OS cells is quite
rare, which poses challenges in developing CAR-T cells that target
these specific antigens. On the other hand, TAAs are present on
both cancer cells and some healthy cells, but they are
overexpressed in tumors. While they may not be as specific as TSAs,
they still provide viable targets for immunotherapy. Nevertheless,
the disadvantage of such CAR-T cells is that they can attack normal
tissue cells and therefore have certain toxic side effects
(100).
For instance, CD166 is not exclusively present in OS
tissues, but is also found in certain normal tissues, including
epithelial cells, smooth muscle cells and neurons (101,102). A patient with colorectal cancer
who received numerous third-generation CAR-T cells targeting HER2
developed respiratory distress and cardiac arrest shortly after the
infusion of the T cells, and succumbed to multiple organ failure 5
days later. It was therefore hypothesized that CAR-T cells
recognize HER2 in the lung epithelium despite being expressed at
low levels, causing pulmonary edema and cytokine storms that lead
to adverse outcomes. This was the first fatal adverse event caused
by CARs recognition of target antigens outside tumor tissues
(57). Thus, it is essential to
carefully evaluate the safety and effectiveness of CAR-T cell
therapy.
Target antigens such as DNAJB8, HER2 and CD276 have
been found on the surface of OS cells, and CAR-T cells targeting
these antigens have been developed to specifically eliminate tumors
(22,56,103). If the antigen of the tumor cell
is lost or the density of the tumor antigen is reduced, the tumor
cells cannot be targeted by CAR-T cells, and therefore the tumor
cannot be effectively eradicated. Based on extensive clinical
experience with CD19-CAR-T cells in acute B-lymphoblastic leukemia,
the loss of CD19 antigen was postulated to occur through two
distinct mechanisms: i) Isoform switching and ii) lineage
switching. Isoform switching refers to the fact that mutated
leukemia cells lack the epitope recognized by CD19-CAR-T cells,
and/or CD19 is preferentially retained in the cell and therefore
not recognized by T cells, while lineage switching refers to a
phenomenon in the development of myeloid leukemia where tumor cells
do not express CD19 in the presence of CD19-CARs (104). The same occurs with OS. Hsu
et al (25) reported an
increase in tumor cells lacking EphA2 expression following
EphA2-CAR-T cell therapy for OS. Consequently, immune escape may
seriously affect the antitumor effect of CAR-T cells.
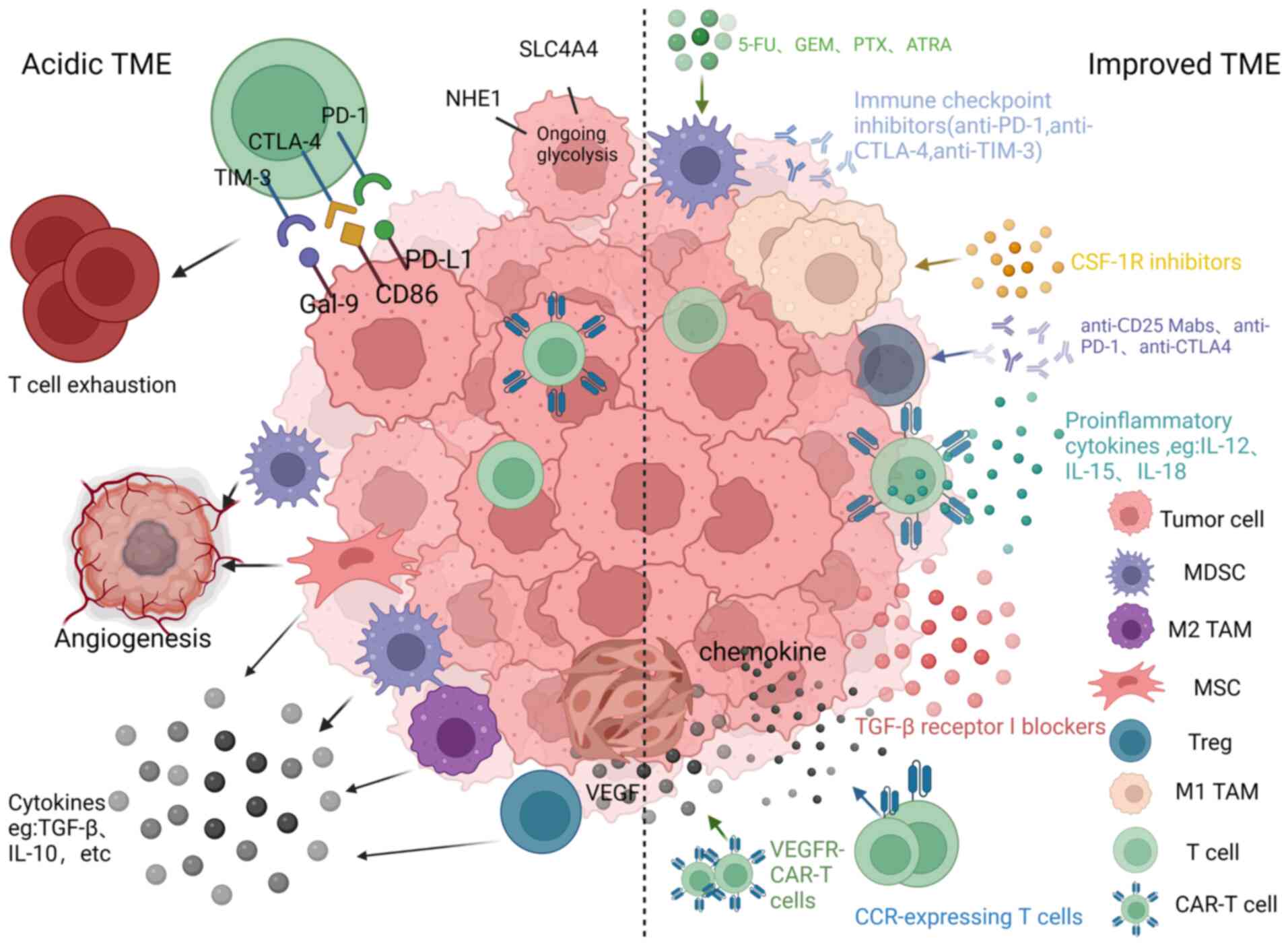
CAR-T cells often encounter an inhibitory TME when
infused into patients. This microenvironment can support
angiogenesis, promote tumor progression and immune escape, and
reduce the ability of CAR-T cells to proliferate and persist in
vivo (Fig. 4) (105,106). The following subsection
introduces specific immunosuppressive mechanisms.
Immune checkpoints serve as immunosuppressive
molecules that help to regulate the intensity and magnitude of the
immune response. Immune checkpoints, including cytotoxic T
lymphocyte antigen 4 (CTLA-4), TIM-3, Gal-9, programmed death
receptor 1 (PD-1) and PD ligand 1 (PD-L1), play a crucial role in
promoting immune tolerance during tumor formation and development
(107). PD-1 is expressed on
activated effector T cells and other tumor-infiltrating lymphocytes
(TILs), including NK cells. Its presence inhibits immune responses
in activated T cells, resulting in a suppressed immune response.
This phenomenon, namely the fact that PD-1+ T cells are
no longer cytotoxic and therefore cannot fight tumors, is called T
cell exhaustion (108).
Hashimoto et al (109)
studied 16 samples of OS, and suggested that PD-1 and PD-L1 may be
associated with tumor recurrence, metastasis and patient mortality.
CTLA-4 was found to be expressed in OS cells. Administration of
anti-CTLA-4 antibodies has been observed to augment the antitumor
response of cytotoxic T cells (110,111). In addition, TIM-3 and Gal-9 are
also expressed in OS tissues, and significantly promote the
apoptosis of CD4+ and CD8+ T cells in the TME
of OS, and lead to poor prognosis in patients with OS (112,113). Thus, blocking immune checkpoints
could further enhance the function of CAR-T cells.
Within the TME, there are diverse cell populations
that contribute to immune suppression, including myeloid-derived
suppressor cells (MDSCs), tumor-associated macrophages (TAMs) and
regulatory T cells (Tregs). Together with tumor cells, these cells
orchestrate the control of the tumor and facilitate the production
of inhibitory cytokines, chemokines and growth factors (114).
MDSCs are a prominent type of immune-suppressive
cells that can hinder the antitumor function of CAR-T cells and can
facilitate tumor angiogenesis. Additionally, MDSCs impede T cell
proliferation by employing arginase, transforming growth factor
(TGF)-β, inducible nitric oxide synthase and IL-10 (115). Moreover, MDSCs can phagocytose
antigens, degrade them and present them to CD8+ T cells
that are specialized in recognizing such antigens. When MDSCs
interact directly with T cells, MDSCs produce a substance called
peroxynitrite. This substance modifies certain amino acids in the
T-cell receptor and CD8+ molecules on the surface of T
cells, which in turn impairs the response of T cells to specific
antigen stimulation (115). The
activity of MDSCs reduces the effectiveness of the immune response
within the TME.
TAMs can be classified into two distinct categories:
i) M1 TAMs, which inhibit tumor growth, and ii) M2 TAMs, which
promote tumor growth (116). In
the majority of cases, tumor cells have the ability to drive the
differentiation of TAMs towards the immunosuppressive M2 subtype
rather than the immuno-stimulatory M1 subtype. These M2 TAMs play
an important role in promoting tumor formation and facilitating the
spread of high-grade OS metastasis. This is achieved through the
release of immunosuppressive soluble factors such as IL-10, TGF-β2
and C-C motif chemokine ligand (CCL) 22 (117,118).
Tregs play a crucial role in inducing
immunosuppression through various mechanisms. Firstly, Tregs
express CTLA-4, which inhibits the maturation of antigen-presenting
cells by reducing the expression of CD80/86, and hinders the
activation of immune responses. Secondly, Tregs consume IL-2, a
molecule necessary for the activation of effector T cells,
including CD8+ T cells. As Tregs have high-affinity IL-2
receptors but produce minimal IL-2 themselves, this leads to a lack
of IL-2 for effector T cells, resulting in their dysfunction.
Lastly, Tregs secrete immunosuppressive cytokines like IL-10 and
IL-35, directly inhibiting the activation of T cells (119).
Immunosuppressive cytokines such as IL-10, IL-6 and
TGF-β play an essential role in reducing the effectiveness of T
cells in fighting against the tumor (120-122). Among them, the most critical
inhibitory cytokine is TGF-β, which can act in tumors to stimulate
tumor cells to spread further, metastasize and produce cytokines
(123). In OS tissue, OS cells
can directly release TGF-β1, and improved expression of TGF-β1 is
associated with metastasis of high-grade OS (124). Moreover, elevated levels of
IL-10 in tumor tissue hinder the production of IL-12 by dendritic
cells, which inhibits the cytotoxic T cell response and the
activation of NK cells (125).
Furthermore, IL-10 hinders the antigen presentation process and
suppresses the release of pro-inflammatory cytokines by
antigen-presenting cells, which promotes immune tolerance. Elevated
levels of IL-10 not only exhibit strong immunosuppressive activity
but also contribute to tumor cell proliferation and increased
resistance to chemotherapy (126). IL-6 acts not only internally in
tumor cells to promote tumor cell proliferation, survival and
metastasis, but also externally in other cells present in the TME
to promote tumor angiogenesis and evade immune surveillance
(127).
MSCs are considered the utmost important factor in
promoting OS migration in the TME (128). On the one hand, the interaction
between MSCs and tumor cells facilitates angiogenesis and
ultimately leads to the formation of new blood vessel networks in
tumor tissues (129,130). However, the newly formed blood
vessels are often structurally and morphologically abnormal,
leading to persistent hypoxia, acidic TME and invasion of tumor
cells into blood vessels, which facilitates tumor cell metastasis,
and impairs CAR-T cell survival (131). For example, in a rat model of
OS, intravenous administration of MSCs had no effect on tumor
growth but significantly promoted metastasis to the lung (132). MSCs could stimulate the
proliferation of CD4+ T cells and Tregs, while
simultaneously causing a decrease in CD8+ T cells
(133). Moreover, certain
cytokines secreted by MSCs, such as TGF-β, may promote tumor immune
escape and debilitate the antitumor immune response (129). Furthermore, MSCs induce the
upregulation of indoleamine 2,3-dioxygenase and PD-L1, which are
both involved in the immunosuppression of tumors, leading to
inhibition of the efficacy of CAR-T cells (133).
Tissue hypoxia is a characteristic of the OS
microenvironment. Hypoxia, or low oxygen conditions, within the TME
plays an important role in chemotherapy resistance, tumor
advancement and metastasis in OS. Firstly, hypoxia could regulate
OS by primarily activating the hypoxia-inducible factor (HIF)
(134). HIF is a heterodimer
composed of α and β subunits. HIF expression is increased in OS and
has the function of regulating the cellular adaptive response to
hypoxia by promoting the transcription of various hypoxia-inducible
genes. There are three subtypes of HIF-α, namely HIF-1α, HIF-2α and
HIF-3α. Among these HIFs, the most important is HIF-1α (135), which promotes the distant
metastasis of OS, and the invasion and proliferation of the OS MG63
and U2OS cell lines, which are inhibited when the expression of
HIF-1α is reduced (136,137).
Extracellular vesicles are membrane-bound vesicles
that cells release into the surrounding extracellular matrix. They
contain various essential signaling molecules, including proteins,
nucleic acids, enzymes and other soluble factors. These vesicles
play a crucial role in cell communication, angiogenesis and tumor
cell proliferation. They achieve this by exchanging information
with target cells. Yang et al (145) demonstrated that extracellular
vesicles were involved in the progression and spread of OS.
Tumor-derived exosomes have been shown in multiple studies to
hinder the function of T and NK cells through diverse mechanisms,
including induction of T cell apoptosis, enabling OS cells to evade
immune surveillance (146).
Prudowsky and Yustein (147)
demonstrated that extracellular vesicles facilitated the transfer
of multidrug resistance 1 mRNA between OS cells, thereby enhancing
resistance to Adriamycin.
In the TME, CAR-T cells require sufficient nutrients
and energy substances, including glucose, amino acids and
glutamine, which are also necessary for tumor cell metabolism, for
exerting their antitumor effect. However, tumor cells consume
energy more rapidly than T cells, which inhibits T cell
proliferation and function due to insufficient nutrient
requirements (148).
First, OS cells promote c-Myc expression through
platelet-derived growth factor (PDGF)/PDGF receptor β signaling and
increase aerobic glycolysis under aerobic conditions (149). Second, TME may also be deprived
of essential amino acids. In mouse models of colon and lung cancer,
MDSCs with high arginase activity were locally present in tumors
and spleens, consuming arginine, and inhibiting T-cell
proliferation and cytokine production (150). Arginase-1 is an amino
acid-degrading enzyme commonly expressed in the TME, which promotes
L-arginine metabolism and inhibits T-lymphocyte responses (151). Third, glutamine is another
important factor affecting T-cell function. Since glutamine is
required for T cell proliferation, decreased glutamine metabolism
in T cells reduces the expression of metabolic regulators and
impedes T cell activation (152,153). Thus, it is of utmost importance
to maintain a metabolic equilibrium between T and tumor cells,
which is vital for the successful management of tumors.
CRS, a severe adverse effect linked to CAR-T cell
therapy, is an inflammatory syndrome triggered by the production of
several cytokines by CAR-T cells. This syndrome is considered to be
associated with high antitumor activity and tumor burden. The
cytokines implicated in CRS consist of IFN-γ, TNF-α, IL-8, IL-10
and macrophage colony-stimulating factor (154,155). The initial indication of CRS
typically manifests as fever, which can emerge within hours to days
following the infusion of cells. After the initial fever, patients
may experience nausea, vomiting, diarrhea, skin rash, delirium, low
blood pressure and even severe multi-organ failure (99). A previous study reported serious
adverse events, even fatal, caused by CRS (57).
Clinical evidence indicates that the expandability
and persistence of T cells after infusion are crucial factors for
achieving effective cancer therapy (156). However, during CAR-T cell
therapy for OS, T cells may become depleted due to various reasons,
rendering them unable to fulfill their intended function. A
previous study has indicated that a transcription factor known as
nuclear receptor subfamily 4 group A number 1 (NR4A1) plays an
important role in T cell fatigue. In mouse models with
tumor-bearing mice, the injection of NR4A1-CAR-T cells has shown a
remarkable ability to impede tumor growth (157). Furthermore, when antigens
stimulate naive T cells, they undergo proliferation and
differentiate into memory T cells. This process involves several
stages, starting with naive T cells and progressing to stem cell
memory T cells, central memory T cells, effector memory T cells and
finally effector T cells. In mice, it has been observed that the
less differentiated subtypes, including naive T cells, stem cell
memory T cells and central memory T cells, consistently exhibit
higher expansion, longer persistence and stronger antitumor
abilities compared with the more differentiated effector memory T
cells and finally effector T cells (158). Thus, naive T cells, stem cell
memory T cells, central memory T cells, can be employed to augment
the antitumor efficacy of CAR-T cells during CAR-T cell
production.
CAR-T cells targeting solid tumors must penetrate
the extracellular matrix and immunosuppressive TME to reach the
tumor (159). Some chemokines
secreted by solid tumors, including CXCL123, CXCL12 and CXCL5,
impede the movement and entry of T cells into tumor sites; however,
the lack of appropriate chemokine receptors on T cells hampers
their ability to infiltrate the tumor region (160-162). The presence of tumor cells
diminishes the quantity and effectiveness of CAR-T cells,
significantly compromising their ability to eliminate tumor cells.
Hence, enhancing the penetration capacity of CAR-T cells is
crucial.
In contrast to hematological tumors, OS is a type
of solid tumor. The antigens targeted by immunotherapy on solid
tumors typically exhibit heterogeneity, varying not only between
different types of solid tumors but also between the primary and
metastatic stages of the same tumor (163). Thus, careful selection of
appropriate TAAs plays a vital role in achieving an effective
anti-OS response.
To increase the eliminating efficiency of CAR-T
cells and reduce their side effects, a number of measures have been
taken based on the shortcomings of CAR-T cells. These measures are
mainly aimed at promoting the effect of CAR-T cells, including
optimizing CAR-T cell structure, improving the immunological
microenvironment, increasing T cell efficiency and preventing
CRS.
The utilization of CAR-T cell therapy comes with
the inherent risk of off-target effects due to the fact that the
TAAs, which CAR-T cells recognize, are not restricted to tumor
cells, but can also be detected in normal tissue cells.
Additionally, the efficacy of CAR-T cells can be directly reduced
by the escape, depletion or loss of tumor antigens. To address
these challenges, clinical trials are currently exploring the use
of CAR-T cells with two or three target antigens to improve their
effectiveness. Therefore, enhancing the anti-OS effect of CAR-T
cells could be achieved by optimizing their structure.
The main function of the extracellular domain of
CAR-T cells is to recognize tumor cell antigens. Researchers have
improved the extracellular domain of CAR-T cells to tackle the
issues of tumor antigen escape, reduction, loss and off-target
effects (164,165). As a result, numerous CAR-T cells
have been constructed, including dual-targeted, triple-targeted and
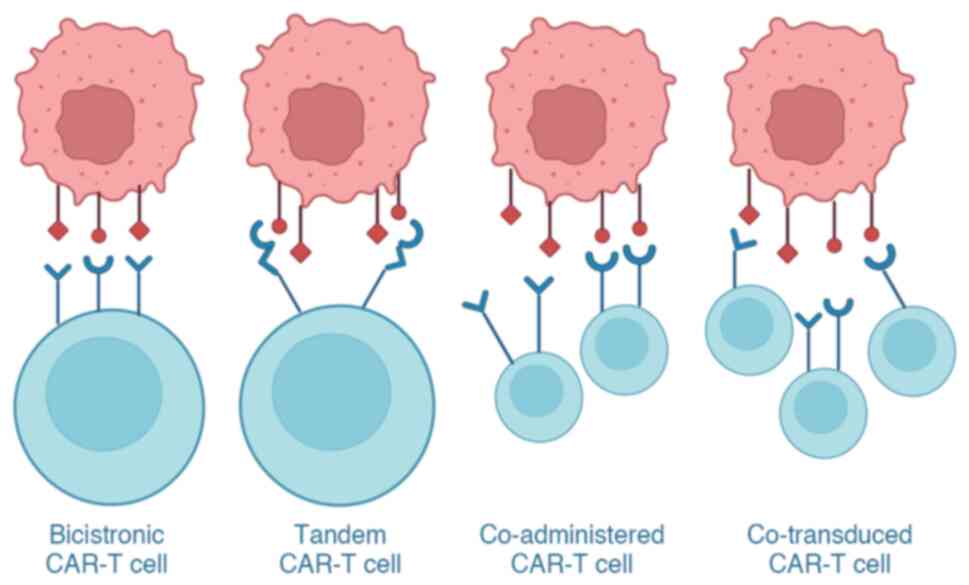
tandem CAR-T cells (Fig. 5).
Dual- and triple-targeted CAR-T cells refer to the design of two or
three separate CARs on a T lymphocyte, where each CAR is specific
for different antigens and thus has dual or triple targeting
ability. Tandem CAR-T cells are defined as those with two different
scFv regions in a single CAR, targeting different antigens.
The HD or spacer region is located in the
extracellular region of CARs. The HD mainly plays the following
roles: i) It increases the flexibility of the ARD, which is
conducive to target antigen recognition; and ii) both the hinge
length and type affect the strength of the activation signal of the
CARs and the formation of immune synapses (167,168). To a certain extent, the size of
the HD in CAR can be adjusted, reducing the obstacle created by the
spatial distance between target and CAR-T cells (169). A long hinge has the advantage of
favorable flexibility, which favors the capture of target epitopes
close to the membrane with the ARD (170,171). Hudecek et al (172) demonstrated that enhancing the
length of the HD could enhance the ability of CAR-T cells to
recognize tumors, but a short hinge has the advantage of
facilitating the binding of target epitopes far from the membrane
(173). The main sources of
hinge mentioned in the literature are CD8, CD28, immunoglobulin
(Ig)G1, IgG4 and IgD (172,174). Previous studies have revealed
that variations in the HD can impact the elimination efficiency,
longevity and cytotoxicity of CAR-T cells towards healthy cells
(167,175-178). Therefore, optimizing the HD
holds great importance for improving the antitumor effect of CAR-T
cells.
The TMD serves to anchor CARs to the cell membrane
of T lymphocytes. The TMD is normally formed by CD28, CD8, CD4 and
ICOS (179). The TMD has been
identified to increase the stability and mobility of CARs (180,181). In first-generation CARs, TMD
derived from CD247 promotes T lymphocyte activation (180). The commonly used second- and
third-generation CD8- and CD28-derived TMD further improves the
stability, mobility and efficacy of CARs compared with the
CD247-derived TMD (181-183). Fujiwara et al (184) reported that altering the
structure of the TMD affected the expression levels and activity of
CARs. The authors also found that modifying the TMD could modulate
the function of CAR-T cells without impacting the antigen-binding
properties of the ARD or the signal transduction properties of the
signal transduction domain (STD). Although previous studies have
improved the persistence of T cells, inhibited tumor growth and
reduced the incidence of complications such as CRS by optimizing
the structure of the TMD of CAR, the role of the TMD needs further
investigation (181,182,185). Thus, ongoing optimization of the
TMD structure is important for improving the efficacy of CAR-T
cells.
The intracellular domain of CAR-T cells comprises
both costimulatory molecules and a STD. Huang et al
(186) observed that the
costimulation domains significantly enhanced the expansion and
long-term persistence of CAR-T cells for therapeutic purposes. The
costimulatory domains significantly contributed to the
proliferation and persistence of CAR-T cells for therapy. The TNF
receptor (CD137, CD27 and OX40) (53,187,188) and CD28 (189,190) families are the most commonly
used costimulatory molecules. Exploring the activation mechanism of
costimulatory molecules is a crucial approach to identify
optimization strategies for CAR-T cells. Currently, CD28 and CD137
are FDA-approved costimulatory molecules for CAR-T cells (191). Although the effectiveness of
CAR-T cells with these two costimulatory molecules in treating
hematological tumors is similar, there are differences in the
eliminating effect, duration and mechanism of CAR-T cells.
Specifically, CAR-T cells using CD28 as a costimulatory molecule
are characterized by rapid eliminating and high efficacy but low
durability (192,193), while CAR-T cells using CD137 as
a costimulatory molecule are characterized by moderate and weak
eliminating but high durability (193). Several studies have attempted to
optimize costimulatory molecules to enhance T lymphocyte activity.
Guedan et al (194)
identified an amino acid residue in CD28 that promoted T lymphocyte
exhaustion. The authors replaced phenylalanine with asparagine in
CD28, which delayed T cell exhaustion, increased durability and
improved antitumor efficacy. Further studies should combine the
advantages and disadvantages of the costimulatory molecules CD28
and CD137, and redesign them to increase antitumor activity.
Overall, the signals provided by costimulatory molecules during
CAR-T cell activation are critical for T lymphocyte metabolism,
survival and efficacy. Therefore, a detailed study of the mechanism
of costimulatory molecules would be helpful to construct CAR-T
cells with high efficacy, long endurance and low side effects to
improve their effect on tumor treatment.
The STD primarily consists of CD247, which is
mainly derived from the T cell receptor complex. CD247 contains
three immunoreceptor tyrosine-based activation motifs (ITAMs) and
is present in almost all CAR structures. Van der Merwe and Dushek
(195) reported that the ITAM
was an immune receptor activation domain consisting of two Yxx
(I/L) and 6-8 amino acids. Gaud et al (196) found that Src family
lymphocyte-specific protein tyrosine kinase phosphorylates the
tyrosine in the ITAM, which then recruits ZAP70 kinase and further
activates signaling molecules such as T lymphocyte linker protein
and phospholipase C-γ. In addition, Feucht et al (197) found that altering the second and
third ITAMs in CD247 increased the efficacy and durability of CAR-T
cells. James (198) noticed that
augmenting the number of ITAMs in a second-generation CAR led to an
enhancement in the activity of CAR-T cells. Thus, the localization
and quantity of ITAMs can influence the functionality of CAR-T
cells. A comprehensive investigation of the STD and its associated
functions in CAR should therefore help to develop new CAR-T cells
and improve their effectiveness against tumors.
To improve the efficacy of tumor cell eradication,
various studies have experimented with co-transducing CAR-T cells
and employing a combination of two distinct single-targeted CAR-T
cell therapies (199-201) (Fig. 5). Co-transduced CAR-T cells are T
cells that have been co-transduced with two different lentiviral
vectors, with some of the resulting T cells expressing both CARs
and some expressing only one CAR (199). Ghorashian et al (200) employed co-transduced CAR-T cells
for treating patients with relapsed or refractory ALL, and found
that co-transduced CAR-T cells exhibited favorable safety profiles,
robust expansion capacity and promising early efficacy. Dual CAR-T
cell therapy refers to the administration of two single-targeted
CAR-T cells, either simultaneously or sequentially, with each CAR-T
cell targeting a distinct antigen (Fig. 5). It has been demonstrated that
the sequential infusion of distinct CAR-T cells, each designed to
target specific tumor antigens, exhibited promising antitumor
effects in clinical trials (201). These findings suggested that the
sequential injection of different CAR-T cells may be an effective
strategy to combat tumors.
There are numerous ways of improving the TME, and
the mechanisms involved are described below (Fig. 4).
Checkpoint inhibitors increase the immune activity
of CAR-T cells, reducing immunosuppression caused by increased
cytokines and loss of target antigens, leading to activation of
inhibited T cells (202). Zheng
et al (35) reported that
the PD-1 inhibitor nivolumab effectively stopped the spread of OS
in mice by boosting the population of CD8+ cells and
enhancing the cytotoxic function of CD8+ T cells.
Furthermore, PD-1 treatment not only decreased the number of tumor
cells and promoted apoptosis in lung metastases of OS, but also
made macrophages change from an M2 to an M1 phenotype, resulting in
the regression of lung metastases in OS (203). Moreover, Rafiq et al
(204) have engineered CAR-T
cells capable of producing scFv to block PD-1, and preclinical
studies have indicated that these CAR-T cells demonstrate
equivalent or improved efficacy compared with combination therapy
involving CAR-T cells and PD-1 inhibitors. Kenderian et al
(205) reported that immune
checkpoint blockers such as TIM-3 combined with CAR T-cell therapy
could have synergistic antitumor effects.
Immunosuppressive cells present in the TME, such as
MDSCs, Tregs and TAMs, hinder the antitumor effectiveness of CAR-T
cells.
MDSCs play a critical role as immunosuppressive
cells and can be effectively targeted through the following
approaches: i) Depletion of MDSCs in the bloodstream and at tumor
infiltration sites by using low-dose chemotherapy. Drugs such as
5-fluorouracil, paclitaxel and gemcitabine have been shown to
effectively reduce MDSC populations. Combining this low-dose
chemotherapy with CAR-T-cell therapy can enhance antitumor
responses. By targeting and depleting MDSCs, this combination
therapy holds great potential in improving the outcomes of cancer
treatment (206-209); ii) preventing the aggregation of
MDSCs. A previous study revealed a positive association between
MDSC levels and IL-18, indicating that IL-18 promoted the migration
of MDSCs into tumor tissue. By administering anti-IL-18 treatment,
a significant reduction in MDSCs in both tumors and peripheral
blood was observed. This finding suggested that targeting IL-18
could be a favorable approach to improve the effectiveness of
immunotherapy in patients with OS (34); iii) counteracting the
immunosuppressive function of MDSCs by the use of
all-trans-retinoic acid (ATRA). Administering this compound to mice
with OS was found to effectively eliminate MDSCs and reduce their
inhibitory impact. Additionally, combining ATRA with GD2-CAR-T cell
therapy in the management of OS demonstrated significant
improvements in antitumor efficacy. This combination approach holds
great potential in enhancing the immune response against OS and
improving the overall outcome of treatment (74); and iv) promoting the
differentiation of MDSCs into a non-suppressive immune state. ATRA
was found to have a direct effect on MDSCs, inducing their
differentiation into mature myeloid cells such as macrophages and
dendritic cells. This process led to a significant decrease in the
inhibitory effect of MDSCs on T cells, ultimately promoting an
improved immune response (210).
Tregs are another type of immunosuppressive cells.
One of the primary approaches in Tregs-based immunotherapy is
decreasing the population of Tregs, for example, by applying
anti-CD25 monoclonal antibodies to eliminate CD4+ and
CD25+ Tregs, and to facilitate the specific eliminating
effect of CD8+ T cells in OS. The second approach
involves preventing the immunosuppressive effects of Tregs, which
express certain proteins that suppress the immune response. These
proteins, such as CTLA-4 and PD-L1, can be blocked using specific
inhibitors. The third option aims to enhance the activity of
effector T cells, which is responsible for suppressing the
suppressive effects of Tregs. In immune surveillance, checkpoints
inhibit the activation of T cells, leading to the inability of TILs
to eliminate cancer cells in OS. Yoshida et al (211) found that checkpoint inhibitors
could restore T cell-mediated antitumor responses by releasing the
brake and engaging major histocompatibility complex molecules.
TAMs are also immunosuppressive cells in the TME,
and targeting TAMs is mainly achieved by switching subtype and
reducing the number of TAMs. Blocking the receptor of
colony-stimulating factor 1, an important regulator of TAMs
recruitment, not only decreases TAMs recruitment to tumors but also
enhances the differentiation of TAMs into a proinflammatory M1
phenotype, leading to an increased intratumoral M1/M2 ratio in mice
(212). Furthermore, it was
revealed that low-dose radiotherapy could reprogram TAMs into M1
TAMs (213,214). Moreover, certain antigens
expressed on immunosuppressive cells within the TME could be
targeted to eradicate both tumor and immunosuppressive cells, as
demonstrated by the specific action of CD123-CAR-T cells against
tumor cells and TAMs in Hodgkin lymphoma (215).
Neutralizing TGF-β, a critical tumor suppressive
cytokine, has the potential to amplify the antitumor immune
response mediated by CD8+ T cells. Several strategies
have been explored based on this concept. Inhibition of TGF-β1
signaling using vactosertib, a TGF-β receptor (TGF-βR)1 inhibitor,
has demonstrated a significant reduction in OS cell proliferation
both in laboratory settings and in animal models (216). Wallace et al (36) observed that inhibiting TGF-βR led
to a significant improvement in the efficacy of adoptive T-cell
therapy in animal models of solid tumors. Tang et al
(217) demonstrated that
removing TGF-βR 2 by using genome editing techniques reduced
exhaustion in CAR-T cells and enhanced the effectiveness of
mesothelin-CAR-T cells against ovarian cancer cells.
Another approach to increase the efficiency of
CAR-T cells is to induce them to produce proinflammatory cytokines.
Armored CAR-T cells have been hereditarily engineered to produce
proinflammatory cytokines, aiming to shield them from the
inhibitory effects of the TME. IL-18-secreting CAR-T cells, for
example, exhibited increased expansion, persistence and antitumor
cytotoxicity compared with CAR-T cells that target only tumor
antigens. Additionally, these enhanced CAR-T cells not only altered
the TME in mouse lymphomas but also administered IL-18 directly to
the tumors. This approach facilitated the recruitment and
activation of native antitumor immune effector cells, leading to a
robust and comprehensive endogenous antitumor immune response
(39). In addition, CAR-T cells
that could secrete cytokines such as IL-15 (218) and IL-12 (219) demonstrated amplified
proinflammatory capabilities. Thus, CAR-T cells secreting
proinflammatory cytokines would be an ideal solution for treating
OS.
CAR-T cells produce abundant lactate through
glycolysis, which increases the acidity of the TME and inhibits T
lymphocyte function. Small molecules and metabolites such as
cytokines, the glucose analogue 2-deoxy-D-glucose, L-arginine and
carnosine have been revealed to affect T-cell metabolism and
differentiation. Numerous studies have investigated and confirmed
the ability of the cytokines IL-7, IL-15 and IL-21 to limit or
interfere with glycolysis (220,221). 2-Deoxy-D-glucose is a glycolytic
inhibitor. During in vitro expansion, CD8+ T
cells change their differentiation direction in the presence of
2-deoxy-D-glucose to form more memory cells, resulting in
long-lasting antitumor function (222). Intracellular L-arginine is
significantly reduced after activation of naive T cells, and the
additional incorporation of L-arginine into the culture medium
causes a shift in metabolism from glycolysis to oxidative
phosphorylation. It also increases the level of stem cell memory T
cells, resulting in greater antitumor activity (223). In another study, carnosine was
found to limit the acidification of extracellular fluid, changing
the metabolism of activated T cells from glycolytic to aerobic,
ultimately leading to an improved antitumor function of T cells. In
addition, transiently elevating the level of carnosine in the
culture medium increased lentivirus gene expression (224). Therefore, these results provided
ideas to increase the therapeutic efficacy of CAR-T-cell
therapy.
According to the criteria used to evaluate adverse
reactions, CRS has been classified into five levels, ranging from
mild reactions/grade 1 to mortality/grade 5. The extent of CRS
triggered by CAR-T cells is directly associated with the expansion
of CAR-T cells and the concentration of IL-6 in the bloodstream
(225). Turtle et al
(226) conducted a study that
revealed that tocilizumab, an anti-human IL-6 receptor monoclonal
antibody, was the primary management option for patients
experiencing moderate to severe CRS. Li et al (10) emphasized the importance of timely
tocilizumab treatment for elderly patients with grade 3 or 2 CRS
and comorbidities to prevent severe CRS (grade 4 and 5), with the
majority of patients achieving rapid remission post-tocilizumab.
Davila et al (227)
observed that administering high-dose corticosteroids could
effectively impede the proliferation and persistence of CAR-T cells
in vivo. As a result, high-dose steroids could also be
employed as a management alternative for patients experiencing CRS.
However, it is essential to highlight that high-dose steroids are
typically reserved for patients with severe and life-threatening
CRS who have not responded to tocilizumab treatment.
A previous study constructed inhibitory CARs
(iCARs) based on the inhibitory molecules CTLA-4 or PD-1, and
showed that these iCARs could block T cell responses activated by
their endogenous T cell receptors or activated CARs. This
inhibitory effect was transient, and, in the absence of an
inhibitory signal, suppressed T cells could be reactivated when
exposed to an activating signal. Thus, iCARs may allow T cells to
target tumor cells while avoiding attacking normal tissue (37). Moreover, an interesting and
feasible approach is the use the tyrosine kinase inhibitor
dasatinib, which, in preclinical models, efficiently and
temporarily hindered the activation of CAR-T cells. Brief treatment
with dasatinib early after infusion of CAR-T cells could prevent
lethal CRS in mice (228).
It could therefore be inferred that high-grade CRS
may be reversed through early and aggressive treatment. Notably,
resolution of CRS could result in an anti-CRS syndrome
characterized by hypothermia and bradycardia, accompanied by
alterations in cytokine levels (229). Therefore, patients should be
monitored for vital signs even after successful CRS treatment.
Tumor blood vessels, by being highly perfused, have
a crucial role in the progression and metastasis of OS. Thus, CAR-T
cells targeting OS blood vessels have attracted the attention of
researchers. Previous studies have revealed the existence of
pro-angiogenic growth factors in the TME, such as vascular
endothelial growth factor (VEGF), with tumor cells exhibiting
overexpression of receptors for these factors, which is strongly
correlated with adverse prognosis and tumor metastasis (38). Fukumura et al (230) found that VEGFR-2 was
overexpressed in some tumor stromal cells. CARs targeting VEGFR-1
and VEGFR-2 have demonstrated promising efficacy in disrupting
tumor vasculature and reducing tumor cell proliferation by limiting
nutrient and oxygen delivery, and preserving normal tissue
(231,232).
The TME in OS tissue is characterized by a high
density of newly formed blood vessels, immunosuppressive cells and
inhibitory cytokines. These factors work together to shield tumor
cells and hinder the infiltration of CAR-T cells (233). Local drug delivery is considered
as one of the most effective measures to tackle this issue, and it
can be accomplished by directly administering drugs into the tumor
site in various tumor models. Local delivery via
intracranial/intravenous drug administration has yielded positive
results in models of head and neck squamous cell carcinoma,
glioblastoma and colorectal cancer with liver metastases (234,235). Moreover, an alternative method
for delivering CAR-T cells into solid tumors involves the use of
implantable biopolymer devices. This approach eliminates the need
for direct injection into the tumor, allowing for prolonged
exposure of tumor cells to high concentrations of immune cells. In
mouse models with normal immune function but afflicted with
pancreatic cancer and melanoma, the use of a bioactive vector
substantially enhanced T cell expansion and function (40).
Runt-related transcription factor 3 (Runx3) protein
has been found to promote T cell re-accumulation in tumor tissue.
In melanoma mouse models, the expression of the Runx3 gene has been
found to effectively promote the accumulation of T cells in tumor
tissues and suppress tumor growth (236). In addition, accumulation of
collagen and heparan sulfate proteoglycans in tumor tissue hampers
the penetration of T cells into the extracellular matrix, but this
obstacle can be overcome by overexpressing heparanase. Heparanase,
an enzyme naturally present in T cells but absent in CAR-T cells,
degrades heparan sulfate proteoglycans. Research conducted by
Caruana et al (237)
confirmed that CAR-T cells engineered to produce heparanase could
essentially reduce tumor volume. In solid tumors, chemokines
secreted by the tumor can hinder the penetration and migration of T
cells. However, certain chemokine receptors expressed on T cells
can recognize these chemokines present in the TME. Therefore, it is
possible to enhance the homing and infiltration of CAR-T cells by
engineering them to express these specific chemokine receptors.
This approach has shown promise in increasing the significance of
CAR-T cell therapy (238).
A major factor contributing to the inefficacy of
CAR-T cell therapy is the notable decline in the persistence of
CAR-T cells. This decline is primarily a result of the immune
system's impaired ability to identify foreign peptides derived from
CAR, which subsequently leads to modifications in T lymphocytes
through immune-mediated mechanisms (239,240). Methods to improve the
persistence of CAR-T cells mainly include binding to oncolytic
viruses and engineering chimeric inverted receptors. Xia et
al (41) suggested that
combining oncolytic viruses with CAR-T cells improved the activity
of CAR-T cells. In addition, it was suggested that being infected
with oncolytic viruses could potentially boost the capacity of
CAR-T cells to infiltrate tumors, overcome immune system
suppression and ultimately enhance their ability to persist
(241-243). In addition, engineered chimeric
inversion receptors represent a practical way to improve the
persistence of CAR-T cells by transforming the immunosuppressive
surroundings into an immune-stimulating environment. IL-4 secreted
by the tumor exerts an inhibitory impact on T lymphocytes. A novel
approach called reverse CAR has been engineered on T lymphocytes,
where it incorporates the extracellular domain of IL-4 and the
intracellular domain of IL-7. Mohammed et al (105) reported that reverse CAR had the
ability to transform the inhibitory signal of T lymphocytes into a
stimulatory signal, thereby enhancing the persistence of CAR-T
cells. Liu et al (244)
engineered a chimeric PD-1 receptor by combining a truncated PD-1
extracellular domain with the transmembrane and internal domains of
CD28, transforming the PD-1's inhibitory signal into a stimulating
signal to improve the persistence of CAR-T cells. Furthermore,
secretion of cytokines by CAR-T cells, such as IL-2, IL-15 and
IL-18, has been demonstrated to selectively amplify memory CAR-T
cells and enhance their persistence (245). Feucht et al (197) reported that modulating the ITAM
of CD247 could enhance CAR signaling, leading to a reduction in T
cell exhaustion. Continuous activation signal generated by
spontaneous clustering of CAR molecules during in vitro
expansion of CAR-T cells resulted in T cell exhaustion, which
hampered the therapeutic effectiveness and the persistence of CAR-T
cells in vivo. Thus, it would be beneficial to regulate the
density of CAR molecules on the cell surface to prevent the
aggregation of CARs and subsequent sustained cell activation
(246).
CAR-T cell therapy for OS continues to face several
obstacles. Thus, researchers have made several efforts to boost the
effectiveness of CAR-T cell therapy in OS by exploring potential
approaches such as integrating it with radiotherapy or chemotherapy
(Fig. 6) (18).
The efficacy of local control of CAR-T cells or
chemotherapy alone in the treatment of high-grade OS remains
suboptimal (247). As a result,
certain researchers have proposed that CAR-T cells applied together
with chemotherapy could potentially enhance the treatment
outcome.
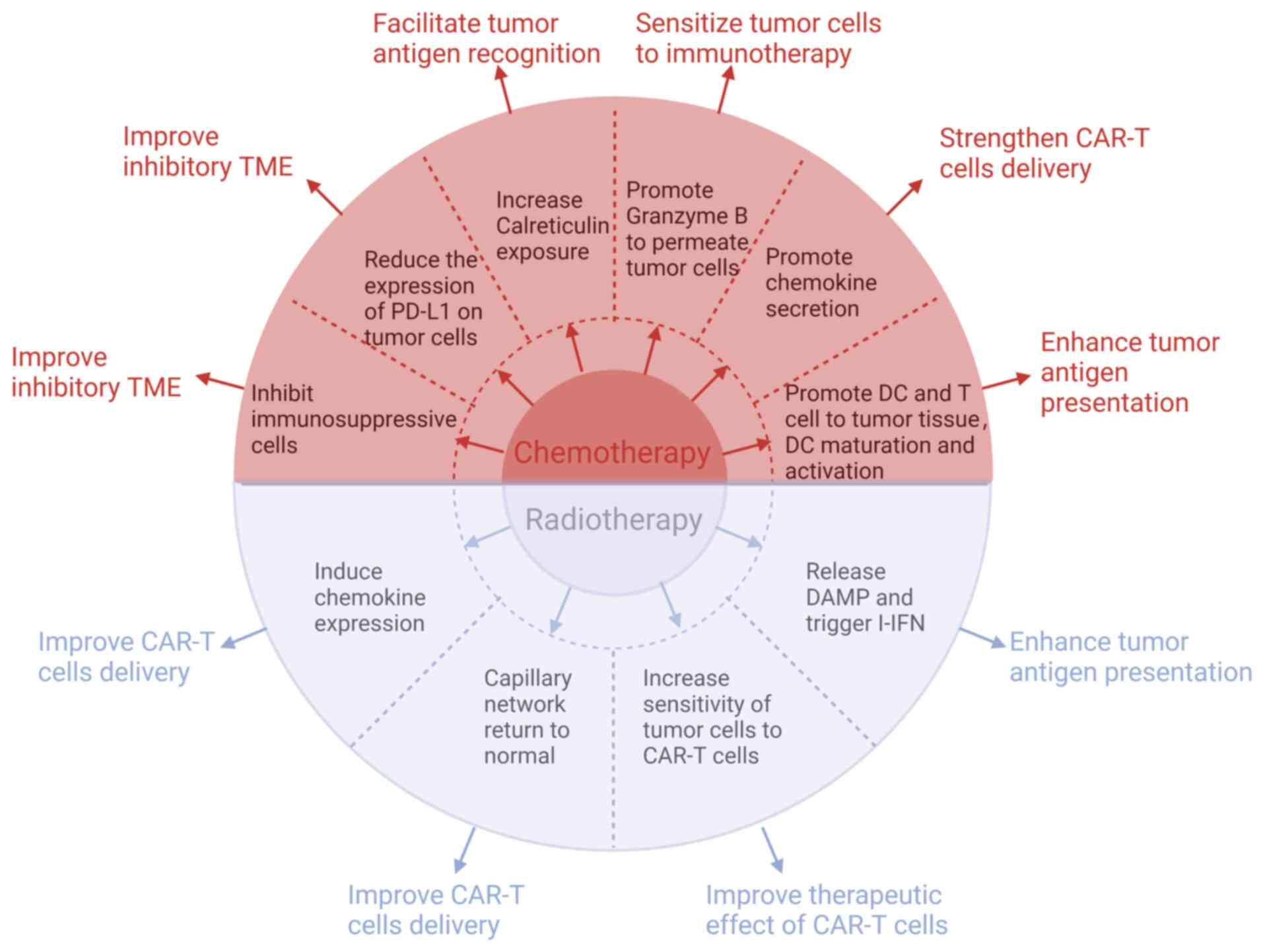
Chemotherapeutic agents such as adriamycin,
methotrexate, cisplatin and ifosfamide, possess not only
tumor-reducing properties but also notable immunomodulatory effects
(43). Regular administration of
low-dose cyclophosphamide can effectively decrease the quantity and
suppressive function of Tregs. Furthermore, it has been observed to
augment the presence of CD8+ T cells, which have a
pivotal function in triggering immune responses against malignant
tumors (248,249). Adriamycin, when combined with
CAR-T cells, boosts their antitumor efficacy by suppressing Tregs
and MDSCs, leading to a significant inhibition of tumor growth in
both mice and humans (250,251). In the context of solid tumors,
Seliger and Quandt (252) noted
that tumor cells possessed the capability to enhance the levels of
immune checkpoint molecules, namely PD-L1 and PD-L2. This mechanism
effectively hindered the activity of T and CAR-T cells attempting
to penetrate and combat the tumor. Chulanetra et al
(43) observed that adriamycin
was able to increase the response of OS to GD2-CAR-T cells. This
effect was achieved by diminishing the expression of PD-L1, an
immune checkpoint, in OS cells. In murine models of pancreatic and
prostate cancer, a pretreatment strategy involving cyclophosphamide
followed by administration of CAR-T cells led to a significant
change in the TME. It effectively shifted the TME from an
anti-inflammatory state to a proinflammatory state, thereby
stimulating the production of chemokines. These chemokines play a
crucial role in attracting CAR-T cells and ultimately revitalize
immunologically 'cold' tumors, transforming them into 'hot' tumors
that are highly responsive to treatment (253).
Overcoming the obstacle of efficiently transporting
T cells to the tumor site remains a critical concern in enhancing
the potency of CAR-T cell treatment. Research on mice at
preclinical stage indicated that there was a link between limited
migration of T cells and diminished levels of chemokine expression
(254). CXCL9 and CXCL10 induced
by adriamycin could attract NKG2D-CD8+ T cells to OS
cells and promote NKG2D-CD8+ T cell homing (250). The study by Srivastava et
al (255) revealed that,
when mice were pre-treated with oxaliplatin and cyclophosphamide, a
notable alteration in the TME was observed. The occurrence
triggered an inflammatory reaction, leading to an increased release
of CCL5, CXCL9, CXCL10 and CXCL16 by tumor macrophages. This
facilitated ROR1-CAR-T cells to enter and effectively target lung
tumor cells, leading to a significant improvement in their ability
to eliminate them (255). The
study by Srivastava et al (255) was not on OS but on lung tumors.
However, both lung tumors and OS are solid tumors and both have a
similar TME. Hence, further exploration into the combination of
chemotherapy and CAR T-cell therapy for enhancing the therapeutic
outcomes of OS is a valuable endeavor.
Following targeted chemotherapy, there was a
notable increase in the presence of mannose-6-phosphate receptor on
cancer cells, which, in turn, allowed granzyme B to enter these
cells more effectively. This process not only aided in
self-regulation but also heightened the responsiveness of tumor
cells to immunotherapy through autophagy (256-258). Parente-Pereira et al
(259) conducted a preclinical
investigation revealing that combining HER2-CAR-T cells with a low
dose of carboplatin not only amplified the sensitivity of tumor
cells to HER2-CAR-T cell-induced cytotoxicity but also augmented
the overall effectiveness of antitumor treatment. While the
mechanism behind increased tumor tissue susceptibility to
immunotherapy after chemotherapy remains unclear, Proietti et
al (260) have observed
improved efficacy in tumor treatment when combining immunotherapy
with chemotherapy. Therefore, it could be hypothesized that
chemotherapy may improve the ability of CAR T-cell therapy in the
treatment of OS and may be a promising direction for future
insightful research.
Radiotherapy has the dual benefit of not only
eliminating tumor cells, but also triggering a targeted immune
response against the tumor, offering a potential treatment for both
local and distant metastatic tumors (266). DeSelm et al (42) found that administering low-dose
total body irradiation can enhance pancreatic adenocarcinoma cells'
responsiveness to CAR-T cells, enabling the elimination of tumor
cells with a reduced dose of CAR-T cells, consequently reducing CRS
incidence and enhancing treatment effectiveness. Thus, it is
plausible to consider that radiotherapy may enhance the effect of
CAR-T cells in the treatment of OS.
Effective infiltration of CAR-T cells into the
tumor are critical steps in inhibiting OS. Weiss et al
(273) demonstrated that the
concurrent use of local radiotherapy and NKG2D-CAR-T cells enhanced
the trafficking of CAR-T cells to the tumor site, leading to an
augmented antitumor response of T cells in glioblastoma. Moreover,
previous studies have reported that local radiotherapy can
stimulate the production of chemokines such as CXCL9, CXCL10 and
CXCL16, which in turn increases the recruitment of T cells into the
TME, thereby facilitating tumor-suppressive effects (274,275). In addition, low-dose
radiotherapy promoted the secretion of IFN-γ, leading to the
normalization of the capillary network, which promoted the
infiltration of cytotoxic T lymphocytes into the tumor (276). As a result, radiotherapy appears
to have the potential to enhance the trafficking of CAR-T cells to
OS, leading to improved treatment outcomes for overall
survival.
Given the crucial role of chemoradiotherapy in
treating OS, it represents a novel and innovative approach to
amplify the antitumor impact of CAR-T cell therapy through this
method. However, CAR-T cell therapy combined with chemoradiotherapy
for treating OS still faces numerous problems. Firstly,
chemoradiotherapy results in treatment-related toxicities,
including effects on the immune system of the host such as
lymphopenia (277).
Additionally, the utilization of CAR-T cells for the treatment of
OS is at an early stage, and the exploration of combining
chemoradiotherapy with CAR-T cell therapy for OS requires further
investigation. While the mechanism of CAR-T cells in conjunction
with chemoradiotherapy for treating OS is not yet fully understood,
this approach remains a promising treatment option. In summary,
CAR-T cell therapy combined with chemoradiotherapy presents a new
and innovative approach to treat OS. However, additional research
is required to improve understanding of its clinical efficacy and
mechanisms of action.
CAR-T cell immunotherapy represents a new
alternative strategy for the treatment of OS. A variety of target
antigens, including DNAJB8, HER2, CD276, CD166, EphA2, GD2,
IL-11Rα, IGF-1R, ROR1, NKG2D, CSPG4 and CD44v6, have been explored
for CAR-T cell establishment and OS cell elimination, and have
produced encouraging results. However, CAR-T cell therapy in OS
still encounters several challenges, including the absence of
specific tumor antigens, tumor antigen evasion, inhibitory TME,
off-target effects and CRS. Consequently, numerous measures have
been undertaken to enhance the effectiveness of CAR-T cells, which
encompass structural modifications of CAR-T cells, TME enhancement,
improved persistence and homing capabilities, engineering CAR-T
cells to target the OS vasculature, and combining CAR-T cells with
radiotherapy, chemotherapy or checkpoint inhibitors. In conclusion,
CAR-T cell therapy represents a novel method for the treatment of
OS, offering an alternative therapeutic option for patients with
OS.
Not applicable.
TY, YW and YZ wrote parts of the initial draft of
this review. WJ modified the initial draft of the manuscript. JJ
contributed to the figures. WJ and MW edited the initial draft of
this paper and made final changes to the final version before
submission. All authors read and approved the final manuscript.
Data authentication is not applicable.
Not applicable.
Not applicable.
The authors declare that they have no competing
interests.
The authors would like to thank Miss Ran Meng
(student; Chengde Medical University) and Miss Zhijia Ma
(postgraduate; Tianjin Medical University) for identifying relevant
literature in the manuscript and for editing the manuscript.
The present study was supported by the Jilin Provincial Science
and Technology Department Project (grant no. YDZJ202301ZYTS032),
the Qinhuangdao Science & Technology Bureau (grant no.
201902A119), the Jilin Health Science and Technology Capability
Enhancement Project (grant no. 2023JC012) and the Jilin University
Bethune Plan Project and Jilin Higher Education Research Project
(grant no. JGJX2021D28).
|
1
|
Misaghi A, Goldin A, Awad M and Kulidjian
AA: Osteosarcoma: A comprehensive review. SICOT J. 4:122018.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Dorfman HD and Czerniak B: Bone cancers.
Cancer. 75(1 Suppl): S203–S210. 1995. View Article : Google Scholar
|
|
3
|
Isakoff MS, Bielack SS, Meltzer P and
Gorlick R: Osteosarcoma: Current treatment and a collaborative
pathway to success. J Clin Oncol. 33:3029–3035. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Rizzo A, Nannini M, Astolfi A, Indio V, De
Iaco P, Perrone AM, De Leo A, Incorvaia L, Di Scioscio V and
Pantaleo MA: Impact of chemotherapy in the adjuvant setting of
early stage uterine leiomyosarcoma: A systematic review and updated
meta-analysis. Cancers (Basel). 12:18992020. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Zhou Y, Yang D, Yang Q, Lv X, Huang W,
Zhou Z, Wang Y, Zhang Z, Yuan T, Ding X, et al: Single-cell RNA
landscape of intratumoral heterogeneity and immunosuppressive
microenvironment in advanced osteosarcoma. Nat Commun. 11:63222020.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Rizzo A, Pantaleo MA, Saponara M and
Nannini M: Current status of the adjuvant therapy in uterine
sarcoma: A literature review. World J Clin Cases. 7:1753–1763.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Santoni M, Rizzo A, Mollica V, Matrana MR,
Rosellini M, Faloppi L, Marchetti A, Battelli N and Massari F: The
impact of gender on the efficacy of immune checkpoint inhibitors in
cancer patients: The MOUSEION-01 study. Crit Rev Oncol Hematol.
170:1035962022. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Stancovski I, Schindler DG, Waks T, Yarden
Y, Sela M and Eshhar Z: Targeting of T lymphocytes to
Neu/HER2-expressing cells using chimeric single chain Fv receptors.
J Immunol. 151:6577–6582. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Grupp SA, Kalos M, Barrett D, Aplenc R,
Porter DL, Rheingold SR, Teachey DT, Chew A, Hauck B, Wright JF, et
al: Chimeric antigen receptor-modified T cells for acute lymphoid
leukemia. N Engl J Med. 368:1509–1518. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Li W, Wu L, Huang C, Liu R, Li Z, Liu L
and Shan B: Challenges and strategies of clinical application of
CAR-T therapy in the treatment of tumors-a narrative review. Ann
Transl Med. 8:10932020. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Abbas MZ: Strategic use of patent
opposition safeguard to improve equitable access to innovative
health technologies: A case study of CAR T-cell therapy Kymriah.
Glob Public Health. 17:3255–3265. 2022. View Article : Google Scholar
|
|
12
|
Astolfi A, Nannini M, Indio V, Schipani A,
Rizzo A, Perrone AM, De Iaco P, Pirini MG, De Leo A, Urbini M, et
al: Genomic database analysis of uterine leiomyosarcoma mutational
profile. Cancers (Basel). 12:21262020. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Berdeja JG, Madduri D, Usmani SZ,
Jakubowiak A, Agha M, Cohen AD, Stewart AK, Hari P, Htut M,
Lesokhin A, et al: Ciltacabtagene autoleucel, a B-cell maturation
antigen-directed chimeric antigen receptor T-cell therapy in
patients with relapsed or refractory multiple myeloma
(CARTITUDE-1): A phase 1b/2 open-label study. Lancet. 398:314–324.
2021. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Van Oekelen O, Aleman A, Upadhyaya B,
Schnakenberg S, Madduri D, Gavane S, Teruya-Feldstein J, Crary JF,
Fowkes ME, Stacy CB, et al: Neurocognitive and hypokinetic movement
disorder with features of parkinsonism after BCMA-targeting CAR-T
cell therapy. Nat Med. 27:2099–2103. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Raje N, Berdeja J, Lin Y, Siegel D,
Jagannath S, Madduri D, Liedtke M, Rosenblatt J, Maus MV, Turka A,
et al: Anti-BCMA CAR T-cell therapy bb2121 in relapsed or
refractory multiple myeloma. N Engl J Med. 380:1726–1737. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Garfall AL, Stadtmauer EA, Hwang WT, Lacey
SF, Melenhorst JJ, Krevvata M, Carroll MP, Matsui WH, Wang Q,
Dhodapkar MV, et al: Anti-CD19 CAR T cells with high-dose melphalan
and autologous stem cell transplantation for refractory multiple
myeloma. JCI Insight. 4:e1276842018. View Article : Google Scholar
|
|
17
|
Guo B, Chen M, Han Q, Hui F, Dai H, Zhang
W, Zhang Y, Wang Y, Zhu H and Han W: CD138-directed adoptive
immunotherapy of chimeric antigen receptor (CAR)-modified T cells
for multiple myeloma. J Cell Immunother. 2:28–35. 2016. View Article : Google Scholar
|
|
18
|
Zhu J, Simayi N, Wan R and Huang W: CAR T
targets and microenvironmental barriers of osteosarcoma.
Cytotherapy. 24:567–576. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Boettcher M, Joechner A, Li Z, Yang SF and
Schlegel P: Development of CAR T cell therapy in children-A
comprehensive overview. J Clin Med. 11:21582022. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Sadelain M, Brentjens R and Rivière I: The
basic principles of chimeric antigen receptor design. Cancer Discv.
3:388–398. 2013. View Article : Google Scholar
|
|
21
|
Morita R, Nishizawa S, Torigoe T,
Takahashi A, Tamura Y, Tsukahara T, Kanaseki T, Sokolovskaya A,
Kochin V, Kondo T, et al: Heat shock protein DNAJB8 is a novel
target for immunotherapy of colon cancer-initiating cells. Cancer
Sci. 105:389–395. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Ahmed N, Brawley VS, Hegde M, Robertson C,
Ghazi A, Gerken C, Liu E, Dakhova O, Ashoori A, Corder A, et al:
Human epidermal growth factor receptor 2 (HER2)-specific chimeric
antigen receptor-modified T cells for the immunotherapy of
HER2-positive sarcoma. J Clin Oncol. 33:1688–1696. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Picarda E, Ohaegbulam KC and Zang X:
Molecular pathways: Targeting B7-H3 (CD276) for human cancer
immunotherapy. Clin Cancer Res. 22:3425–3431. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Wang Y, Yu W, Zhu J, Wang J, Xia K, Liang
C and Tao H: Anti-CD166/4-1BB chimeric antigen receptor T cell
therapy for the treatment of osteosarcoma. J Exp Clin Cancer Res.
38:1682019. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Hsu K, Middlemiss S, Saletta F, Gottschalk
S, McCowage GB and Kramer B: Chimeric antigen receptor-modified T
cells targeting EphA2 for the immunotherapy of paediatric bone
tumours. Cancer Gene Ther. 28:321–334. 2021. View Article : Google Scholar :
|
|
26
|
Reppel L, Tsahouridis O, Akulian J, Davis
IJ, Lee H, Fucà G, Weiss J, Dotti G, Pecot CV and Savoldo B:
Targeting disialoganglioside GD2 with chimeric antigen
receptor-redirected T cells in lung cancer. J Immunother Cancer.
10:e0038972022. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Huang G, Yu L, Cooper LJN, Hollomon M,
Huls H and Kleinerman ES: Genetically modified T cells targeting
interleukin-11 receptor α-chain kill human osteosarcoma cells and
induce the regression of established osteosarcoma lung metastases.
Cancer Res. 72:271–281. 2012. View Article : Google Scholar
|
|
28
|
Huang X, Park H, Greene J, Pao J, Mulvey
E, Zhou SX, Albert CM, Moy F, Sachdev D, Yee D, et al: IGF1R- and
ROR1-specific CAR T cells as a potential therapy for high risk
sarcomas. PLoS One. 10:e01331522015. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Fernández L, Metais JY, Escudero A, Vela
M, Valentín J, Vallcorba I, Leivas A, Torres J, Valeri A,
Patiño-García A, et al: Memory T cells expressing an NKG2D-CAR
efficiently target osteosarcoma cells. Clin Cancer Res.
23:5824–5835. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Riccardo F, Tarone L, Iussich S, Giacobino
D, Arigoni M, Sammartano F, Morello E, Martano M, Gattino F, Maria
R, et al: Identification of CSPG4 as a promising target for
translational combinatorial approaches in osteosarcoma. Ther Adv
Med Oncol. 11:17588359198554912019. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Zhang Y, Ding C, Wang J, Sun G, Cao Y, Xu
L, Zhou L and Chen X: Prognostic significance of CD44V6 expression
in osteosarcoma: A meta-analysis. J Orthop Surg Res. 10:1872015.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Lin Z, Wu Z and Luo W: Chimeric antigen
receptor T-cell therapy: The light of day for osteosarcoma. Cancers
(Basel). 13:44692021. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Shah NN and Fry TJ: Mechanisms of
resistance to CAR T cell therapy. Nat Rev Clin Oncol. 16:372–385.
2019.PubMed/NCBI
|
|
34
|
Guan Y, Zhang R, Peng Z, Dong D, Wei G and
Wang Y: Inhibition of IL-18-mediated myeloid derived suppressor
cell accumulation enhances anti-PD1 efficacy against osteosarcoma
cancer. J Bone Oncol. 9:59–64. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Zheng B, Ren T, Huang Y, Sun K, Wang S,
Bao X, Liu K and Guo W: PD-1 axis expression in musculoskeletal
tumors and antitumor effect of nivolumab in osteosarcoma model of
humanized mouse. J Hematol Oncol. 11:162018. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Wallace A, Kapoor V, Sun J, Mrass P,
Weninger W, Heitjan DF, June C, Kaiser LR, Ling LE and Albelda SM:
Transforming growth factor-beta receptor blockade augments the
effectiveness of adoptive T-cell therapy of established solid
cancers. Clin Cancer Res. 14:3966–3974. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Fedorov VD, Themeli M and Sadelain M:
PD-1- and CTLA-4-based inhibitory chimeric antigen receptors
(iCARs) divert off-target immunotherapy responses. Sci Transl Med.
5:215ra1722013. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Chen JC, Chang YW, Hong CC, Yu YH and Su
JL: The role of the VEGF-C/VEGFRs axis in tumor progression and
therapy. Int J Mol Sci. 14:88–107. 2012. View Article : Google Scholar
|
|
39
|
Avanzi MP, Yeku O, Li X, Wijewarnasuriya
DP, van Leeuwen DG, Cheung K, Park H, Purdon TJ, Daniyan AF,
Spitzer MH and Brentjens RJ: Engineered tumor-targeted T cells
mediate enhanced anti-tumor efficacy both directly and through
activation of the endogenous immune system. Cell Rep. 23:2130–2141.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Smith TT, Moffett HF, Stephan SB, Opel CF,
Dumigan AG, Jiang X, Pillarisetty VG, Pillai SPS, Wittrup KD and
Stephan MT: Biopolymers codelivering engineered T cells and STING
agonists can eliminate heterogeneous tumors. J Clin Invest.
127:2176–2191. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Xia T, Konno H and Barber GN: Recurrent
loss of STING signaling in melanoma correlates with susceptibility
to viral oncolysis. Cancer Res. 76:6747–6759. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
DeSelm C, Palomba ML, Yahalom J, Hamieh M,
Eyquem J, Rajasekhar VK and Sadelain M: Low-dose radiation
conditioning enables CAR T cells to mitigate antigen escape. Mol
Ther. 26:2542–2552. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Chulanetra M, Morchang A, Sayour E,
Eldjerou L, Milner R, Lagmay J, Cascio M, Stover B, Slayton W,
Chaicumpa W, et al: GD2 chimeric antigen receptor modified T cells
in synergy with sub-toxic level of doxorubicin targeting
osteosarcomas. Am J Cancer Res. 10:674–687. 2020.PubMed/NCBI
|
|
44
|
Buka D, Dvořák J, Sitorová V, Hátlová J,
Richter I and Sirák I: Changes in the CD8+ density of tumor
infiltrating lymphocytes after neoadjuvant radiochemotherapy in
patients with rectal adenocarcinom. Klin Onkol. 29:204–209. 2016.In
Czech. View Article : Google Scholar
|
|
45
|
Makita S, Imaizumi K, Kurosawa S and
Tobinai K: Chimeric antigen receptor T-cell therapy for B-cell
non-Hodgkin lymphoma: Opportunities and challenges. Drugs Context.
8:2125672019. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Zhang X, Zhu L, Zhang H, Chen S and Xiao
Y: CAR-T cell therapy in hematological malignancies: Current
opportunities and challenges. Front Immunol. 13:9271532022.
View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Kong Y, Tang L, You Y, Li Q and Zhu X:
Analysis of causes for poor persistence of CAR-T cell therapy in
vivo. Front Immunol. 14:10634542023. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Asmamaw Dejenie T, Tiruneh G/Medhin M,
Dessie Terefe G, Tadele Admasu F, Wale Tesega W and Chekol Abebe E:
Current updates on generations, approvals, and clinical trials of
CAR T-cell therapy. Hum Vaccin Immunother. 18:21142542022.
View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Farkona S, Diamandis EP and Blasutig IM:
Cancer immunotherapy: The beginning of the end of cancer? BMC Med.
14:732016. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
García Merino A: Anticuerpos monoclonales.
Aspectos básicos. Neurología. 26:301–306. 2011. View Article : Google Scholar
|
|
51
|
Lu J, Ding J, Liu Z and Chen T:
Retrospective analysis of the preparation and application of
immunotherapy in cancer treatment (review). Int J Oncol. 60:122022.
View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Ahmed N, Salsman VS, Yvon E, Louis CU,
Perlaky L, Wels WS, Dishop MK, Kleinerman EE, Pule M, Rooney CM, et
al: Immunotherapy for osteosarcoma: Genetic modification of T cells
overcomes low levels of tumor antigen expression. Mol Ther.
17:1779–1787. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Pulè MA, Straathof KC, Dotti G, Heslop HE,
Rooney CM and Brenner MK: A chimeric T cell antigen receptor that
augments cytokine release and supports clonal expansion of primary
human T cells. Mol Ther. 12:933–941. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Yamada R, Takahashi A, Torigoe T, Morita
R, Tamura Y, Tsukahara T, Kanaseki T, Kubo T, Watarai K, Kondo T,
et al: Preferential expression of cancer/testis genes in cancer
stem-like cells: Proposal of a novel sub-category,
cancer/testis/stem gene. Tissue Antigens. 81:428–434. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Nishizawa S, Hirohashi Y, Torigoe T,
Takahashi A, Tamura Y, Mori T, Kanaseki T, Kamiguchi K, Asanuma H,
Morita R, et al: HSP DNAJB8 controls tumor-initiating ability in
renal cancer stem-like cells. Cancer Res. 72:2844–2854. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Watanabe Y, Tsukahara T, Murata K, Hamada
S, Kubo T, Kanaseki T, Hirohashi Y, Emori M, Teramoto A,
Nakatsugawa M, et al: Development of CAR-T cells specifically
targeting cancer stem cell antigen DNAJB8 against solid tumours. Br
J Cancer. 128:886–895. 2023. View Article : Google Scholar :
|
|
57
|
Morgan RA, Yang JC, Kitano M, Dudley ME,
Laurencot CM and Rosenberg SA: Case report of a serious adverse
event following the administration of T cells transduced with a
chimeric antigen receptor recognizing ERBB2. Mol Ther. 18:843–851.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Abdou AG, Kandil M, Asaad NY, Dawoud MM,
Shahin AA and Abd Eldayem AF: The prognostic role of Ezrin and
HER2/neu expression in osteosarcoma. Appl Immunohistochem Mol
Morphol. 24:355–363. 2016. View Article : Google Scholar
|
|
59
|
Xuan Y, Sheng Y, Zhang D, Zhang K, Zhang
Z, Ping Y, Wang S, Shi X, Lian J, Liu K, et al: Targeting CD276 by
CAR-T cells induces regression of esophagus squamous cell carcinoma
in xenograft mouse models. Transl Oncol. 14:1011382021. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Zhang Y, He L, Sadagopan A, Ma T, Dotti G,
Wang Y, Zheng H, Gao X, Wang D, DeLeo AB, et al: Targeting
radiation-resistant prostate cancer stem cells by B7-H3 CAR T
cells. Mol Cancer Ther. 20:577–588. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Du H, Hirabayashi K, Ahn S, Kren NP,
Montgomery SA, Wang X, Tiruthani K, Mirlekar B, Michaud D, Greene
K, et al: Antitumor responses in the absence of toxicity in solid
tumors by targeting B7-H3 via chimeric antigen receptor T cells.
Cancer Cell. 35:221–237.e8. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Theruvath J, Sotillo E, Mount CW, Graef
CM, Delaidelli A, Heitzeneder S, Labanieh L, Dhingra S, Leruste A,
Majzner RG, et al: Locoregionally administered B7-H3-targeted CAR T
cells for treatment of atypical teratoid/rhabdoid tumors. Nat Med.
26:712–719. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Talbot LJ, Chabot A, Funk A, Nguyen P,
Wagner J, Ross A, Tillman H, Davidoff A, Gottschalk S and DeRenzo
C: A novel orthotopic implantation technique for osteosarcoma
produces spontaneous metastases and illustrates dose-dependent
efficacy of B7-H3-CAR T cells. Front Immunol. 12:6917412021.
View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Majzner RG, Theruvath JL, Nellan A,
Heitzeneder S, Cui Y, Mount CW, Rietberg SP, Linde MH, Xu P, Rota
C, et al: CAR T cells targeting B7-H3, a pan-cancer antigen,
demonstrate potent preclinical activity against pediatric solid
tumors and brain tumors. Clin Cancer Res. 25:2560–2574. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Swart GWM: Activated leukocyte cell
adhesion molecule (CD166/ALCAM): Developmental and mechanistic
aspects of cell clustering and cell migration. Eur J Cell Biol.
81:313–321. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Federman N, Chan J, Nagy JO, Landaw EM,
McCabe K, Wu AM, Triche T, Kang H, Liu B, Marks JD and Denny CT:
Enhanced growth inhibition of osteosarcoma by cytotoxic polymerized
liposomal nanoparticles targeting the alcam cell surface receptor.
Sarcoma. 2012:1269062012. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
He S, Li S, Guo J, Zeng X, Liang D, Zhu Y,
Li Y, Yang D and Zhao X: CD166-specific CAR-T cells potently target
colorectal cancer cells. Transl Oncol. 27:1015752023. View Article : Google Scholar
|
|
68
|
Kang BH, Jensen KJ, Hatch JA and Janes KA:
Simultaneous profiling of 194 distinct receptor transcripts in
human cells. Sci Signal. 6:rs132013. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Pasquale EB: Eph receptors and ephrins in
cancer: Bidirectional signalling and beyond. Nat Rev Cancer.
10:165–180. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Wykosky J and Debinski W: The EphA2
receptor and ephrinA1 ligand in solid tumors: Function and
therapeutic targeting. Mol Cancer Res. 6:1795–1806. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Fritsche-Guenther R, Noske A, Ungethüm U,
Kuban RJ, Schlag PM, Tunn PU, Karle J, Krenn V, Dietel M and Sers
C: De novo expression of EphA2 in osteosarcoma modulates activation
of the mitogenic signalling pathway. Histopathology. 57:836–850.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Doronin II, Vishnyakova PA, Kholodenko IV,
Ponomarev ED, Ryazantsev DY, Molotkovskaya IM and Kholodenko RV:
Ganglioside GD2 in reception and transduction of cell death signal
in tumor cells. BMC Cancer. 14:2952014. View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Roth M, Linkowski M, Tarim J, Piperdi S,
Sowers R, Geller D, Gill J and Gorlick R: Ganglioside GD2 as a
therapeutic target for antibody-mediated therapy in patients with
osteosarcoma. Cancer. 120:548–554. 2014. View Article : Google Scholar
|
|
74
|
Long AH, Highfill SL, Cui Y, Smith JP,
Walker AJ, Ramakrishna S, El-Etriby R, Galli S, Tsokos MG, Orentas
RJ and Mackall CL: Reduction of MDSCs with all-trans retinoic acid
improves CAR therapy efficacy for sarcomas. Cancer Immunol Res.
4:869–880. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Suri M, Soni N, Okpaleke N, Yadav S, Shah
S, Iqbal Z, Alharbi MG, Kalra HS and Hamid P: A deep dive into the
newest avenues of immunotherapy for pediatric osteosarcoma: A
systematic review. Cureus. 13:e183492021.PubMed/NCBI
|
|
76
|
Park JA and Cheung NKV: GD2 or HER2
targeting T cell engaging bispecific antibodies to treat
osteosarcoma. J Hematol Oncol. 13:1722020. View Article : Google Scholar : PubMed/NCBI
|
|
77
|
Jiang J, Wang R, Yang L, Sha Y, Zhao S,
Guo J, Chen D, Zhong Z and Meng F: IL-11Rα-targeted nanostrategy
empowers chemotherapy of relapsed and patient-derived osteosarcoma.
J Control Release. 350:460–470. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Lokau J, Schoeder V and Garbers C: The
role of interleukin-11 in osteosarcoma. Der Pathologe. 41:163–167.
2020.In German. View Article : Google Scholar
|
|
79
|
Li YS, Liu Q, He HB and Luo W: The
possible role of insulin-like growth factor-1 in osteosarcoma. Curr
Probl Cancer. 43:228–235. 2019. View Article : Google Scholar
|
|
80
|
Duan Z, Choy E, Harmon D, Yang C, Ryu K,
Schwab J, Mankin H and Hornicek FJ: Insulin-like growth factor-I
receptor tyrosine kinase inhibitor cyclolignan picropodophyllin
inhibits proliferation and induces apoptosis in multidrug resistant
osteosarcoma cell lines. Mol Cancer Ther. 8:2122–2130. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
81
|
Tan X, Fan S, Wu W and Zhang Y:
MicroRNA-26a inhibits osteosarcoma cell proliferation by targeting
IGF-1. Bone Res. 3:150332015. View Article : Google Scholar : PubMed/NCBI
|
|
82
|
Liu Y, Zhu ST, Wang X, Deng J, Li WH,
Zhang P and Liu BS: MiR-100 inhibits osteosarcoma cell
proliferation, migration, and invasion and enhances
chemosensitivity by targeting IGFIR. Technol Cancer Res Treat.
15:NP40–NP48. 2016. View Article : Google Scholar
|
|
83
|
Chen G, Fang T, Huang Z, Qi Y, Du S, Di T,
Lei Z, Zhang X and Yan W: MicroRNA-133a inhibits osteosarcoma cells
proliferation and invasion via targeting IGF-1R. Cell Physiol
Biochem. 38:598–608. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
84
|
Hojjat-Farsangi M, Moshfegh A,
Daneshmanesh AH, Khan AS, Mikaelsson E, Osterborg A and Mellstedt
H: The receptor tyrosine kinase ROR1-an oncofetal antigen for
targeted cancer therapy. Semin Cancer Biol. 29:21–31. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
85
|
Dai B, Shen Y, Yan T and Zhang A:
Wnt5a/ROR1 activates DAAM1 and promotes the migration in
osteosarcoma cells. Oncol Rep. 43:601–608. 2020.PubMed/NCBI
|
|
86
|
Zhang T, Barber A and Sentman CL:
Generation of antitumor responses by genetic modification of
primary human T cells with a chimeric NKG2D receptor. Cancer Res.
66:5927–5933. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
87
|
Ding H, Yang X and Wei Y: Fusion proteins
of NKG2D/NKG2DL in cancer immunotherapy. Int J Mol Sci. 19:1772018.
View Article : Google Scholar : PubMed/NCBI
|
|
88
|
Baumeister SH, Murad J, Werner L, Daley H,
Trebeden-Negre H, Gicobi JK, Schmucker A, Reder J, Sentman CL,
Gilham DE, et al: Phase I trial of autologous CAR T cells targeting
NKG2D ligands in patients with AML/MDS and multiple myeloma. Cancer
Immunol Res. 7:100–112. 2019. View Article : Google Scholar
|
|
89
|
Tao K, He M, Tao F, Xu G, Ye M, Zheng Y
and Li Y: Development of NKG2D-based chimeric antigen receptor-T
cells for gastric cancer treatment. Cancer Chemother Pharmacol.
82:815–827. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
90
|
Zhang Y, Li X, Zhang J and Mao L: Novel
cellular immunotherapy using NKG2D CAR-T for the treatment of
cervical cancer. Biomed Pharmacother. 131:1105622020. View Article : Google Scholar : PubMed/NCBI
|
|
91
|
Sun B, Yang D, Dai H, Liu X, Jia R, Cui X,
Li W, Cai C, Xu J and Zhao X: Eradication of hepatocellular
carcinoma by NKG2D-based CAR-T cells. Cancer Immunol Res.
7:1813–1823. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
92
|
Wang X, Wang Y, Yu L, Sakakura K, Visus C,
Schwab JH, Ferrone CR, Favoino E, Koya Y, Campoli MR, et al: CSPG4
in cancer: multiple roles. Curr Mol Med. 10:419–429. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
93
|
Rolih V, Barutello G, Iussich S, De Maria
R, Quaglino E, Buracco P, Cavallo F and Riccardo F: CSPG4: A
prototype oncoantigen for translational immunotherapy studies. J
Transl Med. 15:1512017. View Article : Google Scholar : PubMed/NCBI
|
|
94
|
Casanova JM, Almeida JS, Reith JD, Sousa
LM, Fonseca R, Freitas-Tavares P, Santos-Rosa M and
Rodrigues-Santos P: Tumor-infiltrating lymphocytes and cancer
markers in osteosarcoma: Influence on patient survival. Cancers
(Basel). 13:60752021. View Article : Google Scholar : PubMed/NCBI
|
|
95
|
Deng Z, Niu G, Cai L, Wei R and Zhao X:
The prognostic significance of CD44V6, CDH11, and β-catenin
expression in patients with osteosarcoma. Biomed Res Int.
2013:4961932013. View Article : Google Scholar
|
|
96
|
Qiao GL, Song LN, Deng ZF, Chen Y and Ma
LJ: Prognostic value of CD44v6 expression in breast cancer: A
meta-analysis. Onco Targets Ther. 11:5451–5457. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
97
|
Saito S, Okabe H, Watanabe M, Ishimoto T,
Iwatsuki M, Baba Y, Tanaka Y, Kurashige J, Miyamoto Y and Baba H:
CD44v6 expression is related to mesenchymal phenotype and poor
prognosis in patients with colorectal cancer. Oncol Rep.
29:1570–1578. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
98
|
Nakajima K, Taniguchi K and Mutoh KI:
Expression of CD44v6 as matrix-associated ectodomain in the bone
development. J Vet Med Sci. 72:1017–1022. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
99
|
Ma S, Li X, Wang X, Cheng L, Li Z, Zhang
C, Ye Z and Qian Q: Current progress in CAR-T cell therapy for
solid tumors. Int J Biol Sci. 15:2548–2560. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
100
|
Liu B, Yan L and Zhou M: Target selection
of CAR T cell therapy in accordance with the TME for solid tumors.
Am J Cancer Res. 9:228–241. 2019.PubMed/NCBI
|
|
101
|
Saifullah MK, Fox DA, Sarkar S, Abidi SM,
Endres J, Piktel J, Haqqi TM and Singer NG: Expression and
characterization of a novel CD6 ligand in cells derived from joint
and epithelial tissues. J Immunol. 173:6125–6133. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
102
|
Ikeda K and Quertermous T: Molecular
isolation and characterization of a soluble isoform of activated
leukocyte cell adhesion molecule that modulates endothelial cell
function. J Biol Chem. 279:55315–55323. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
103
|
Zhang Q, Zhang Z, Liu G, Li D, Gu Z, Zhang
L, Pan Y, Cui X, Wang L, Liu G, et al: B7-H3 targeted CAR-T cells
show highly efficient anti-tumor function against osteosarcoma both
in vitro and in vivo. BMC Cancer. 22:11242022. View Article : Google Scholar : PubMed/NCBI
|
|
104
|
Majzner RG, Heitzeneder S and Mackall CL:
Harnessing the immunotherapy revolution for the treatment of
childhood cancers. Cancer Cell. 31:476–485. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
105
|
Mohammed S, Sukumaran S, Bajgain P,
Watanabe N, Heslop HE, Rooney CM, Brenner MK, Fisher WE, Leen AM
and Vera JF: Improving chimeric antigen receptor-modified T cell
function by reversing the immunosuppressive tumor microenvironment
of pancreatic cancer. Mol Ther. 25:249–258. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
106
|
Anderson KG, Stromnes IM and Greenberg PD:
Obstacles posed by the tumor microenvironment to T cell activity: A
case for synergistic therapies. Cancer Cell. 31:311–325. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
107
|
Pardoll DM: The blockade of immune
checkpoints in cancer immunotherapy. Nat Rev Cancer. 12:252–264.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
108
|
Wagner LM and Adams VR: Targeting the PD-1
pathway in pediatric solid tumors and brain tumors. Onco Targets
Ther. 10:2097–2106. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
109
|
Hashimoto K, Nishimura S and Akagi M:
Characterization of PD-1/PD-L1 immune checkpoint expression in
osteosarcoma. Diagnostics (Basel). 10:5282020. View Article : Google Scholar : PubMed/NCBI
|
|
110
|
Kawano M, Itonaga I, Iwasaki T and Tsumura
H: Enhancement of antitumor immunity by combining anti-cytotoxic T
lymphocyte antigen-4 antibodies and cryotreated tumor lysate-pulsed
dendritic cells in murine osteosarcoma. Oncol Rep. 29:1001–1006.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
111
|
Lussier DM, Johnson JL, Hingorani P and
Blattman JN: Combination immunotherapy with α-CTLA-4 and α-PD-L1
antibody blockade prevents immune escape and leads to complete
control of metastatic osteosarcoma. J Immunother Cancer. 3:212015.
View Article : Google Scholar
|
|
112
|
Sun CY, Zhang Z, Tao L, Xu FF, Li HY,
Zhang HY and Liu W: T cell exhaustion drives osteosarcoma
pathogenesis. Ann Transl Med. 9:14472021. View Article : Google Scholar : PubMed/NCBI
|
|
113
|
Liu Y, Luo J, Li Y, Cao J and Wang X: IFNγ
and TNFα synergistically promote galectin 9 secretion by human
osteosarcoma cells MG-63 to prevent T cell killing. Int J Clin Exp
Pathol. 13:2009–2017. 2020.
|
|
114
|
Sterner RC and Sterner RM: CAR-T cell
therapy: Current limitations and potential strategies. Blood Cancer
J. 11:692021. View Article : Google Scholar : PubMed/NCBI
|
|
115
|
Gabrilovich DI and Nagaraj S:
Myeloid-derived suppressor cells as regulators of the immune
system. Nat Rev Immunol. 9:162–174. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
116
|
Hattinger CM, Salaroglio IC, Fantoni L,
Godel M, Casotti C, Kopecka J, Scotlandi K, Ibrahim T, Riganti C
and Serra M: Strategies to overcome resistance to immune-based
therapies in osteosarcoma. Int J Mol Sci. 24:7992023. View Article : Google Scholar : PubMed/NCBI
|
|
117
|
Huang Q, Liang X, Ren T, Huang Y, Zhang H,
Yu Y, Chen C, Wang W, Niu J, Lou J and Guo W: The role of
tumor-associated macrophages in osteosarcoma
progression-therapeutic implications. Cell Oncol (Dordr).
44:525–539. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
118
|
Dumars C, Ngyuen JM, Gaultier A, Lanel R,
Corradini N, Gouin F, Heymann D and Heymann MF: Dysregulation of
macrophage polarization is associated with the metastatic process
in osteosarcoma. Oncotarget. 7:78343–78354. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
119
|
Sugiyama D, Hinohara K and Nishikawa H:
Significance of regulatory T cells in cancer immunology and
immunotherapy. Exp Dermatol. 32:256–263. 2023. View Article : Google Scholar
|
|
120
|
Taylor A, Verhagen J, Blaser K, Akdis M
and Akdis CA: Mechanisms of immune suppression by interleukin-10
and transforming growth factor-beta: The role of T regulatory
cells. Immunology. 117:433–442. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
121
|
Law AMK, Valdes-Mora F and Gallego-Ortega
D: Myeloid-derived suppressor cells as a therapeutic target for
cancer. Cells. 9:5612020. View Article : Google Scholar : PubMed/NCBI
|
|
122
|
Xiao H, Chen L, Luo G, Son H, Prectoni JH
and Zheng W: Effect of the cytokine levels in serum on
osteosarcoma. Tumor Biol. 35:1023–1028. 2014. View Article : Google Scholar
|
|
123
|
Tian B, Du X, Zheng S and Zhang Y: The
role of tumor microenvironment in regulating the plasticity of
osteosarcoma cells. Int J Mol Sci. 23:161552022. View Article : Google Scholar : PubMed/NCBI
|
|
124
|
Lamora A, Talbot J, Bougras G, Amiaud J,
Leduc M, Chesneau J, Taurelle J, Stresing V, Le Deley MC, Heymann
MF, et al: Overexpression of smad7 blocks primary tumor growth and
lung metastasis development in osteosarcoma. Clin Cancer Res.
20:5097–5112. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
125
|
Shen L, Li J, Liu Q, Song W, Zhang X,
Tiruthani K, Hu H, Das M, Goodwin TJ, Liu R and Huang L: Local
blockade of interleukin 10 and C-X-C motif chemokine ligand 12 with
nano-delivery promotes antitumor response in murine cancers. ACS
Nano. 12:9830–9841. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
126
|
Rossowska J, Anger N, Szczygieł A,
Mierzejewska J and Pajtasz-Piasecka E: Reprogramming the murine
colon cancer microenvironment using lentivectors encoding shRNA
against IL-10 as a component of a potent DC-based
chemoimmunotherapy. J Exp Clin Cancer Res. 37:1262018. View Article : Google Scholar : PubMed/NCBI
|
|
127
|
Zeng J, Chen S, Li C, Ye Z, Lin B, Liang
Y, Wang B, Ma Y, Chai X, Zhang X, et al: Mesenchymal stem/stromal
cells-derived IL-6 promotes nasopharyngeal carcinoma growth and
resistance to cisplatin via upregulating CD73 expression. J Cancer.
11:2068–2079. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
128
|
Yang C, Tian Y, Zhao F, Chen Z, Su P, Li Y
and Qian A: Bone microenvironment and osteosarcoma metastasis. Int
J Mol Sci. 21:69852020. View Article : Google Scholar : PubMed/NCBI
|
|
129
|
Chang AI, Schwertschkow AH, Nolta JA and
Wu J: Involvement of mesenchymal stem cells in cancer progression
and metastases. Curr Cancer Drug Targets. 15:88–98. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
130
|
Pietrovito L, Leo A, Gori V, Lulli M,
Parri M, Becherucci V, Piccini L, Bambi F, Taddei ML and Chiarugi
P: Bone marrow-derived mesenchymal stem cells promote invasiveness
and transendothelial migration of osteosarcoma cells via a
mesenchymal to amoeboid transition. Mol Oncol. 12:659–676. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
131
|
Lugano R, Ramachandran M and Dimberg A:
Tumor angiogenesis: Causes, consequences, challenges and
opportunities. Cell Mol Life Sci. 77:1745–1770. 2020. View Article : Google Scholar :
|
|
132
|
Tsukamoto S, Honoki K, Fujii H, Tohma Y,
Kido A, Mori T, Tsujiuchi T and Tanaka Y: Mesenchymal stem cells
promote tumor engraftment and metastatic colonization in rat
osteosarcoma model. Int J Oncol. 40:163–169. 2012.
|
|
133
|
Zhang R, Liu Q, Zhou S, He H, Zhao M and
Ma W: Mesenchymal stem cell suppresses the efficacy of CAR-T toward
killing lymphoma cells by modulating the microenvironment through
stanniocalcin-1. Elife. 12:e829342023. View Article : Google Scholar : PubMed/NCBI
|
|
134
|
Zhang L, Song J, Xin X, Sun D, Huang H,
Chen Y, Zhang T and Zhang Y: Hypoxia stimulates the migration and
invasion of osteosarcoma via up-regulating the NUSAP1 expression.
Open Med (Wars). 16:1083–1089. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
135
|
Lv X, Li J, Zhang C, Hu T, Li S, He S, Yan
H, Tan Y, Lei M, Wen M and Zuo J: The role of hypoxia-inducible
factors in tumor angiogenesis and cell metabolism. Genes Dis.
4:19–24. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
136
|
Guan G, Zhang Y, Lu Y, Liu L, Shi D, Wen
Y, Yang L, Ma Q, Liu T, Zhu X, et al: The HIF-1α/CXCR4 pathway
supports hypoxia-induced metastasis of human osteosarcoma cells.
Cancer Lett. 357:254–264. 2015. View Article : Google Scholar
|
|
137
|
Liu M, Wang D and Li N: MicroRNA-20b
downregulates HIF-1α and inhibits the proliferation and invasion of
osteosarcoma cells. Oncol Res. 23:257–266. 2016. View Article : Google Scholar
|
|
138
|
Galon J and Bruni D: Approaches to treat
immune hot, altered and cold tumours with combination
immunotherapies. Nat Rev Drug Discov. 18:197–218. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
139
|
de la Cruz-López KG, Castro-Muñoz LJ,
Reyes-Hernández DO, García-Carrancá A and Manzo-Merino J: Lactate
in the regulation of tumor microenvironment and therapeutic
approaches. Front Oncol. 9:11432019. View Article : Google Scholar : PubMed/NCBI
|
|
140
|
Kopecka J, Salaroglio IC, Perez-Ruiz E,
Sarmento-Ribeiro AB, Saponara S, De Las Rivas J and Riganti C:
Hypoxia as a driver of resistance to immunotherapy. Drug Resist
Updat. 59:1007872021. View Article : Google Scholar : PubMed/NCBI
|
|
141
|
Avnet S, Di Pompo G, Chano T, Errani C,
Ibrahim-Hashim A, Gillies RJ, Donati DM and Baldini N:
Cancer-associated mesenchymal stroma fosters the stemness of
osteosarcoma cells in response to intratumoral acidosis via NF-κB
activation. Int J Cancer. 140:1331–1345. 2017. View Article : Google Scholar :
|
|
142
|
Bobulescu IA, Di Sole F and Moe OW: Na+/H+
exchangers: Physiology and link to hypertension and organ ischemia.
Curr Opin Nephrol Hypertens. 14:485–494. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
143
|
Swietach P, Vaughan-Jones RD and Harris
AL: Regulation of tumor pH and the role of carbonic anhydrase 9.
Cancer Metastasis Rev. 26:299–310. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
144
|
Chiche J, Brahimi-Horn MC and Pouysségur
J: Tumor hypoxia induces a metabolic shift causing acidosis: A
common feature in cancer. J Cell Mol Med. 14:771–794. 2010.
View Article : Google Scholar
|
|
145
|
Yang Q, Liu J, Wu B, Wang X, Jiang Y and
Zhu D: Role of extracellular vesicles in osteosarcoma. Int J Med
Sci. 19:1216–1226. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
146
|
Chen C, Xie L, Ren T, Huang Y, Xu J and
Guo W: Immunotherapy for osteosarcoma: Fundamental mechanism,
rationale, and recent breakthroughs. Cancer Lett. 500:1–10. 2021.
View Article : Google Scholar
|
|
147
|
Prudowsky ZD and Yustein JT: Recent
insights into therapy resistance in osteosarcoma. Cancers (Basel).
13:832020. View Article : Google Scholar
|
|
148
|
Xie J, Wu H, Dai C, Pan Q, Ding Z, Hu D,
Ji B, Luo Y and Hu X: Beyond Warburg effect-dual metabolic nature
of cancer cells. Sci Rep. 4:49272014. View Article : Google Scholar
|
|
149
|
Tang HY, Guo JQ, Sang BT, Cheng JN and Wu
XM: PDGFRβ modulates aerobic glycolysis in osteosarcoma HOS cells
via the PI3K/AKT/mTOR/c-Myc pathway. Biochem Cell Biol. 100:75–84.
2022. View Article : Google Scholar
|
|
150
|
Zea AH, Rodriguez PC, Atkins MB, Hernandez
C, Signoretti S, Zabaleta J, McDermott D, Quiceno D, Youmans A,
O'Neill A, et al: Arginase-producing myeloid suppressor cells in
renal cell carcinoma patients: A mechanism of tumor evasion. Cancer
Res. 65:3044–3048. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
151
|
Rodríguez PC and Ochoa AC: Arginine
regulation by myeloid derived suppressor cells and tolerance in
cancer: Mechanisms and therapeutic perspectives. Immunol Rev.
222:180–191. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
152
|
Pietrobon V and Marincola FM: Hypoxia and
the phenomenon of immune exclusion. J Transl Med. 19:92021.
View Article : Google Scholar : PubMed/NCBI
|
|
153
|
Patel CH and Powell JD: Targeting T cell
metabolism to regulate T cell activation, differentiation and
function in disease. Curr Opin Immunol. 46:82–88. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
154
|
Schubert ML, Schmitt M, Wang L, Ramos CA,
Jordan K, Muller-Tidow C and Dreger P: Side-effect management of
chimeric antigen receptor (CAR) T-cell therapy. Ann Oncol.
32:34–48. 2021. View Article : Google Scholar
|
|
155
|
Almåsbak H, Aarvak T and Vemuri MC: CAR T
cell therapy: A game changer in cancer treatment. J Immunol Res.
2016:54746022016. View Article : Google Scholar : PubMed/NCBI
|
|
156
|
Maus MV, Grupp SA, Porter DL and June CH:
Antibody-modified T cells: CARs take the front seat for hematologic
malignancies. Blood. 123:2625–2635. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
157
|
Chen J, López-Moyado IF, Seo H, Lio CJ,
Hempleman LJ, Sekiya T, Yoshimura A, Scott-Browne JP and Rao A:
NR4A transcription factors limit CAR T cell function in solid
tumours. Nature. 567:530–534. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
158
|
Klebanoff CA, Gattinoni L and Restifo NP:
Sorting through subsets: Which T-cell populations mediate highly
effective adoptive immunotherapy? J Immunother. 35:651–660. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
159
|
Zhang BL, Qin DY, Mo ZM, Li Y, Wei W, Wang
YS, Wang W and Wei YQ: Hurdles of CAR-T cell-based cancer
immunotherapy directed against solid tumors. Sci China Life Sci.
59:340–348. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
160
|
Kershaw MH, Wang G, Westwood JA, Pachynski
RK, Tiffany HL, Marincola FM, Wang E, Young HA, Murphy PM and Hwu
P: Redirecting migration of T cells to chemokine secreted from
tumors by genetic modification with CXCR2. Hum Gene Ther.
13:1971–1980. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
161
|
Wang G, Lu X, Dey P, Deng P, Wu CC, Jiang
S, Fang Z, Zhao K, Konaparthi R, Hua S, et al: Targeting
YAP-dependent MDSC infiltration impairs tumor progression. Cancer
Discov. 6:80–95. 2016. View Article : Google Scholar :
|
|
162
|
Feig C, Jones JO, Kraman M, Wells RJ,
Deonarine A, Chan DS, Connell CM, Roberts EW, Zhao Q, Caballero OL,
et al: Targeting CXCL12 from FAP-expressing carcinoma-associated
fibroblasts synergizes with anti-PD-L1 immunotherapy in pancreatic
cancer. Proc Natl Acad Sci USA. 110:20212–20217. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
163
|
Xia AL, Wang XC, Lu YJ, Lu XJ and Sun B:
Chimeric-antigen receptor T (CAR-T) cell therapy for solid tumors:
Challenges and opportunities. Oncotarget. 8:90521–90531. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
164
|
Tian M, Cheuk AT, Wei JS, Abdelmaksoud A,
Chou HC, Milewski D, Kelly MC, Song YK, Dower CM, Li N, et al: An
optimized bicistronic chimeric antigen receptor against GPC2 or
CD276 overcomes heterogeneous expression in neuroblastoma. J Clin
Invest. 132:e1556212022. View Article : Google Scholar : PubMed/NCBI
|
|
165
|
Muhammad N, Wang R, Li W, Zhang Z, Chang
Y, Hu Y, Zhao J, Zheng X, Mao Q and Xia H: A novel TanCAR targeting
IL13Rα2 and EphA2 for enhanced glioblastoma therapy. Mol Ther
Oncolytics. 24:729–741. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
166
|
Han X, Wang Y, Wei J and Han W:
Multi-antigen-targeted chimeric antigen receptor T cells for cancer
therapy. J Hematol Oncol. 12:1282019. View Article : Google Scholar : PubMed/NCBI
|
|
167
|
Hudecek M, Sommermeyer D, Kosasih PL,
Silva-Benedict A, Liu L, Rader C, Jensen MC and Riddell SR: The
nonsignaling extracellular spacer domain of chimeric antigen
receptors is decisive for in vivo antitumor activity. Cancer
Immunol Res. 3:125–135. 2015. View Article : Google Scholar
|
|
168
|
Srivastava S and Riddell SR: Engineering
CAR-T cells: Design concepts. Trends Immunol. 36:494–502. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
169
|
Künkele A, Johnson AJ, Rolczynski LS,
Chang CA, Hoglund V, Kelly-Spratt KS and Jensen MC: Functional
tuning of CARs reveals signaling threshold above which CD8+ CTL
antitumor potency is attenuated due to cell fas-FasL-dependent
AICD. Cancer Immunol Res. 3:368–379. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
170
|
James SE, Greenberg PD, Jensen MC, Lin Y,
Wang J, Till BG, Raubitschek AA, Forman SJ and Press OW: Antigen
sensitivity of CD22-specific chimeric TCR is modulated by target
epitope distance from the cell membrane. J Immunol. 180:7028–7038.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
171
|
Wilkie S, Picco G, Foster J, Davies DM,
Julien S, Cooper L, Arif S, Mather SJ, Taylor-Papadimitriou J,
Burchell JM and Maher J: Retargeting of human T cells to
tumor-associated MUC1: The evolution of a chimeric antigen
receptor. J Immunol. 180:4901–4909. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
172
|
Hudecek M, Lupo-Stanghellini MT, Kosasih
PL, Sommermeyer D, Jensen MC, Rader C and Riddell SR: Receptor
affinity and extracellular domain modifications affect tumor
recognition by ROR1-specific chimeric antigen receptor T cells.
Clin Cancer Res. 19:3153–3164. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
173
|
Guest RD, Hawkins RE, Kirillova N, Cheadle
EJ, Arnold J, O'Neill A, Irlam J, Chester KA, Kemshead JT, Shaw DM,
et al: The role of extracellular spacer regions in the optimal
design of chimeric immune receptors: Evaluation of four different
scFvs and antigens. J Immunother. 28:203–211. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
174
|
Brudno JN, Lam N, Vanasse D, Shen YW, Rose
JJ, Rossi J, Xue A, Bot A, Scholler N, Mikkilineni L, et al: Safety
and feasibility of anti-CD19 CAR T cells with fully human binding
domains in patients with B-cell lymphoma. Nat Med. 26:270–280.
2020. View Article : Google Scholar : PubMed/NCBI
|
|
175
|
Alabanza L, Pegues M, Geldres C, Shi V,
Wiltzius JJW, Sievers SA, Yang S and Kochenderfer JN: Function of
novel Anti-CD19 chimeric antigen receptors with human variable
regions is affected by hinge and transmembrane domains. Mol Ther.
25:2452–2465. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
176
|
Hombach A, Hombach AA and Abken H:
Adoptive immunotherapy with genetically engineered T cells:
Modification of the IgG1 Fc 'spacer' domain in the extracellular
moiety of chimeric antigen receptors avoids 'off-target' activation
and unintended initiation of an innate immune response. Gene Ther.
17:1206–1213. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
177
|
Almåsbak H, Walseng E, Kristian A, Myhre
MR, Suso EM, Munthe LA, Andersen JT, Wang MY, Kvalheim G,
Gaudernack G and Kyte JA: Inclusion of an IgG1-Fc spacer abrogates
efficacy of CD19 CAR T cells in a xenograft mouse model. Gene Ther.
22:391–403. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
178
|
Watanabe N, Bajgain P, Sukumaran S, Ansari
S, Heslop HE, Rooney CM, Brenner MK, Leen AM and Vera JF:
Fine-tuning the CAR spacer improves T-cell potency. Oncoimmunology.
5:e12536562016. View Article : Google Scholar
|
|
179
|
Stoiber S, Cadilha BL, Benmebarek MR,
Lesch S, Endres S and Kobold S: Limitations in the design of
chimeric antigen receptors for cancer therapy. Cells. 8:4722019.
View Article : Google Scholar : PubMed/NCBI
|
|
180
|
Bridgeman JS, Hawkins RE, Bagley S,
Blaylock M, Holland M and Gilham DE: The optimal antigen response
of chimeric antigen receptors harboring the CD3zeta transmembrane
domain is dependent upon incorporation of the receptor into the
endogenous TCR/CD3 complex. J Immunol. 184:6938–6949. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
181
|
Guedan S, Posey AD Jr, Shaw C, Wing A, Da
T, Patel PR, McGettigan SE, Casado-Medrano V, Kawalekar OU,
Uribe-Herranz M, et al: Enhancing CAR T cell persistence through
ICOS and 4-1BB costimulation. JCI Insight. 3:e969762018. View Article : Google Scholar : PubMed/NCBI
|
|
182
|
Wan Z, Shao X, Ji X, Dong L, Wei J, Xiong
Z, Liu W and Qi H: Transmembrane domain-mediated Lck association
underlies bystander and costimulatory ICOS signaling. Cell Mol
Immunol. 17:143–152. 2020. View Article : Google Scholar :
|
|
183
|
Majzner RG, Rietberg SP, Sotillo E, Dong
R, Vachharajani VT, Labanieh L, Myklebust JH, Kadapakkam M, Weber
EW, Tousley AM, et al: Tuning the antigen density requirement for
CAR T-cell activity. Cancer Discov. 10:702–723. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
184
|
Fujiwara K, Tsunei A, Kusabuka H, Ogaki E,
Tachibana M and Okada N: Hinge and transmembrane domains of
chimeric antigen receptor regulate receptor expression and
signaling threshold. Cells. 9:11822020. View Article : Google Scholar : PubMed/NCBI
|
|
185
|
Yan Z, Cao J, Cheng H, Qiao J, Zhang H,
Wang Y, Shi M, Lan J, Fei X, Jin L, et al: A combination of
humanised anti-CD19 and anti-BCMA CAR T cells in patients with
relapsed or refractory multiple myeloma: A single-arm, phase 2
trial. Lancet Haematol. 6:e521–e529. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
186
|
Huang R, Li X, He Y, Zhu W, Gao L, Liu Y,
Gao L, Wen Q, Zhong JF, Zhang C and Zhang X: Recent advances in
CAR-T cell engineering. J Hematol Oncol. 13:862020. View Article : Google Scholar : PubMed/NCBI
|
|
187
|
Imai C, Mihara K, Andreansky M, Nicholson
IC, Pui CH, Geiger TL and Campana D: Chimeric receptors with 4-1BB
signaling capacity provoke potent cytotoxicity against acute
lymphoblastic leukemia. Leukemia. 18:676–684. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
188
|
Song DG, Ye Q, Poussin M, Harms GM, Figini
M and Powell DJ Jr: CD27 costimulation augments the survival and
antitumor activity of redirected human T cells in vivo. Blood.
119:696–706. 2012. View Article : Google Scholar
|
|
189
|
Maher J, Brentjens RJ, Gunset G, Riviere I
and Sadelain M: Human T-lymphocyte cytotoxicity and proliferation
directed by a single chimeric TCRzeta/CD28 receptor. Nat
Biotechnol. 20:70–75. 2002. View Article : Google Scholar
|
|
190
|
Guedan S, Chen X, Madar A, Carpenito C,
McGettigan SE, Frigault MJ, Lee J, Posey AD Jr, Scholler J,
Scholler N, et al: ICOS-based chimeric antigen receptors program
bipolar TH17/TH1 cells. Blood. 124:1070–1080. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
191
|
Mullard A: FDA approves first CAR T
therapy. Nat Rev Drug Discov. 16:6692017.
|
|
192
|
Carpenito C, Milone MC, Hassan R, Simonet
JC, Lakhal M, Suhoski MM, Varela-Rohena A, Haines KM, Heitjan DF,
Albelda SM, et al: Control of large, established tumor xenografts
with genetically retargeted human T cells containing CD28 and CD137
domains. Proc Natl Acad Sci USA. 106:3360–3365. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
193
|
Zhao Z, Condomines M, van der Stegen SJC,
Perna F, Kloss CC, Gunset G, Plotkin J and Sadelain M: Structural
design of engineered costimulation determines tumor rejection
kinetics and persistence of CAR T cells. Cancer Cell. 28:415–428.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
194
|
Guedan S, Madar A, Casado-Medrano V, Shaw
C, Wing A, Liu F, Young RM, June CH and Posey AD Jr: Single residue
in CD28-costimulated CAR-T cells limits long-term persistence and
antitumor durability. J Clin Invest. 130:3087–3097. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
195
|
van der Merwe PA and Dushek O: Mechanisms
for T cell receptor triggering. Nat Rev Immunol. 11:47–55. 2011.
View Article : Google Scholar
|
|
196
|
Gaud G, Lesourne R and Love PE: Regulatory
mechanisms in T cell receptor signalling. Nat Rev Immunol.
18:485–497. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
197
|
Feucht J, Sun J, Eyquem J, Ho YJ, Zhao Z,
Leibold J, Dobrin A, Cabriolu A, Hamieh M and Sadelain M:
Calibration of CAR activation potential directs alternative T cell
fates and therapeutic potency. Nat Med. 25:82–88. 2019. View Article : Google Scholar :
|
|
198
|
James JR: Tuning ITAM multiplicity on T
cell receptors can control potency and selectivity to ligand
density. Sci Signal. 11:eaan10882018. View Article : Google Scholar : PubMed/NCBI
|
|
199
|
Bachiller M, Dobaño-López C,
Rodríguez-García A, Castellsagué J, Gimenez-Alejandre M,
Antoñana-Vildosola A, Martin-Antonio B, Delgado J, Pérez-Galán P,
Juan M, et al: Co-Transduced CD19/BCMA dual-targeting CAR-T cells
for the treatment of non-hodgkin lymphoma. Blood. 140(Suppl 1):
S7386–S7387. 2022. View Article : Google Scholar
|
|
200
|
Ghorashian S, Lucchini G, Richardson R,
Nguyen K, Terris C, Oporto-Espuelas M, Yeung J, Pinner D, Chu J,
Williams L, et al: Dual antigen targeting with co-transduced
CD19/22 CAR T cells may prevent antigen-negative relapse after CAR
T cell therapy for relapsed/refractory ALL. Blood. 140(Suppl 1):
S10352–S10354. 2022. View Article : Google Scholar
|
|
201
|
Wang L, Tan Su Yin E, Zhao H, Ni F, Hu Y
and Huang H: CAR-T cells: The Chinese experience. Expert Opin Biol
Ther. 20:1293–1308. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
202
|
Sun C, Dotti G and Savoldo B: Utilizing
cell-based therapeutics to overcome immune evasion in hematologic
malignancies. Blood. 127:3350–3359. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
203
|
Dhupkar P, Gordon N, Stewart J and
Kleinerman ES: Anti-PD-1 therapy redirects macrophages from an M2
to an M1 phenotype inducing regression of OS lung metastases.
Cancer Med. 7:2654–2664. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
204
|
Rafiq S, Yeku OO, Jackson HJ, Purdon TJ,
van Leeuwen DG, Drakes DJ, Song M, Miele MM, Li Z, Wang P, et al:
Targeted delivery of a PD-1-blocking scFv by CAR-T cells enhances
anti-tumor efficacy in vivo. Nat Biotechnol. 36:847–856. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
205
|
Kenderian SS, Ruella M, Shestova O,
Klichinsky M, Kim M, Porter DL, June CH and Gill S: Identification
of PD1 and TIM3 As checkpoints that limit chimeric antigen receptor
T cell efficacy in leukemia. Biol Blood Marrow Transplant. 22(3
Suppl): S19–S21. 2016. View Article : Google Scholar
|
|
206
|
Suzuki E, Kapoor V, Jassar AS, Kaiser LR
and Albelda SM: Gemcitabine selectively eliminates splenic
Gr-1+/CD11b+ myeloid suppressor cells in tumor-bearing animals and
enhances antitumor immune activity. Clin Cancer Res. 11:6713–6721.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
207
|
Sevko A, Michels T, Vrohlings M, Umansky
L, Beckhove P, Kato M, Shurin GV, Shurin MR and Umansky V:
Antitumor effect of paclitaxel is mediated by inhibition of
myeloid-derived suppressor cells and chronic inflammation in the
spontaneous melanoma mode. J Immunol. 190:2464–2471. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
208
|
Eriksson E, Wenthe J, Irenaeus S, Loskog A
and Ullenhag G: Gemcitabine reduces MDSCs, tregs and TGFβ-1 while
restoring the teff/treg ratio in patients with pancreatic cancer. J
Transl Med. 14:2822016. View Article : Google Scholar
|
|
209
|
Vincent J, Mignot G, Chalmin F, Ladoire S,
Bruchard M, Chevriaux A, Martin F, Apetoh L, Rébé C and
Ghiringhelli F: 5-Fluorouracil selectively kills tumor-associated
myeloid-derived suppressor cells resulting in enhanced T
cell-dependent antitumor immunity. Cancer Res. 70:3052–3061. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
210
|
Kusmartsev S, Su Z, Heiser A, Dannull J,
Eruslanov E, Kübler H, Yancey D, Dahm P and Vieweg J: Reversal of
myeloid cell-mediated immunosuppression in patients with metastatic
renal cell carcinoma. Clin Cancer Res. 14:8270–8278. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
211
|
Yoshida K, Okamoto M, Sasaki J, Kuroda C,
Ishida H, Ueda K, Ideta H, Kamanaka T, Sobajima A, Takizawa T, et
al: Anti-PD-1 antibody decreases tumour-infiltrating regulatory T
cells. BMC Cancer. 20:252020. View Article : Google Scholar : PubMed/NCBI
|
|
212
|
Pyonteck SM, Akkari L, Schuhmacher AJ,
Bowman RL, Sevenich L, Quail DF, Olson OC, Quick ML, Huse JT,
Teijeiro V, et al: CSF-1R inhibition alters macrophage polarization
and blocks glioma progression. Nat Med. 19:1264–1272. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
213
|
Klug F, Prakash H, Huber PE, Seibel T,
Bender N, Halama N, Pfirschke C, Voss RH, Timke C, Umansky L, et
al: Low-dose irradiation programs macrophage differentiation to an
iNOS+/M1 phenotype that orchestrates effective T cell
immunotherapy. Cancer Cell. 24:589–602. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
214
|
Nadella V, Singh S, Jain A, Jain M,
Vasquez KM, Sharma A, Tanwar P, Rath GK and Prakash H: Low dose
radiation primed iNOS + M1macrophages modulate angiogenic
programming of tumor derived endothelium. Mol Carcinog.
57:1664–1671. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
215
|
Ruella M, Klichinsky M, Kenderian SS,
Shestova O, Ziober A, Kraft DO, Feldman M, Wasik MA, June CH and
Gill S: Overcoming the immunosuppressive tumor microenvironment of
Hodgkin lymphoma using chimeric antigen receptor T cells. Cancer
Discov. 7:1154–1167. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
216
|
Choi SH, Myers J, Tomchuck S, Bonner M,
Eid S, Kingsley D, VanHeyst K, Kim SJ, Kim BG and Huang AY: Oral
TGF-βR1 inhibitor vactosertib promotes osteosarcoma regression by
targeting tumor proliferation and enhancing anti-tumor immunity.
Res Sq. rs.3.rs-27092822023.
|
|
217
|
Tang N, Cheng C, Zhang X, Qiao M, Li N, Mu
W, Wei XF, Han W and Wang H: TGF-β inhibition via CRISPR promotes
the long-term efficacy of CAR T cells against solid tumors. JCI
Insight. 5:e1339772020. View Article : Google Scholar
|
|
218
|
Krenciute G, Prinzing BL, Yi Z, Wu MF, Liu
H, Dotti G, Balyasnikova IV and Gottschalk S: Transgenic expression
of IL15 improves antiglioma activity of IL13Rα2-CAR T cells but
results in antigen loss variants. Cancer Immunol Res. 5:571–581.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
219
|
Chmielewski M, Kopecky C, Hombach AA and
Abken H: IL-12 release by engineered T cells expressing chimeric
antigen receptors can effectively Muster an antigen-independent
macrophage response on tumor cells that have shut down tumor
antigen expression. Cancer Res. 71:5697–5706. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
220
|
Loschinski R, Böttcher M, Stoll A, Bruns
H, Mackensen A and Mougiakakos D: IL-21 modulates memory and
exhaustion phenotype of T-cells in a fatty acid oxidation-dependent
manner. Oncotarget. 9:13125–13138. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
221
|
Huang Y, Si X, Shao M, Teng X, Xiao G and
Huang H: Rewiring mitochondrial metabolism to counteract exhaustion
of CAR-T cells. J Hematol Oncol. 15:382022. View Article : Google Scholar : PubMed/NCBI
|
|
222
|
Sukumar M, Liu J, Ji Y, Subramanian M,
Crompton JG, Yu Z, Roychoudhuri R, Palmer DC, Muranski P, Karoly
ED, et al: Inhibiting glycolytic metabolism enhances CD8+ T cell
memory and antitumor function. J Clin Invest. 123:4479–4488. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
223
|
Geiger R, Rieckmann JC, Wolf T, Basso C,
Feng Y, Fuhrer T, Kogadeeva M, Picotti P, Meissner F, Mann M, et
al: L-arginine modulates T cell metabolism and enhances survival
and anti-tumor activity. Cell. 167:829–842.e13. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
224
|
Ghassemi S, Martinez-Becerra FJ, Master
AM, Richman SA, Heo D, Leferovich J, Tu Y, García-Cañaveras JC,
Ayari A, Lu Y, et al: Enhancing chimeric antigen receptor T cell
anti-tumor function through advanced media design. Mol Ther Methods
Clin Dev. 18:595–606. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
225
|
Lee DW, Kochenderfer JN, Stetler-Stevenson
M, Cui YK, Delbrook C, Feldman SA, Fry TJ, Orentas R, Sabatino M,
Shah NN, et al: T cells expressing CD19 chimeric antigen receptors
for acute lymphoblastic leukaemia in children and young adults: A
phase 1 dose-escalation trial. Lancet. 385:517–528. 2015.
View Article : Google Scholar
|
|
226
|
Turtle CJ, Hanafi LA, Berger C, Hudecek M,
Pender B, Robinson E, Hawkins R, Chaney C, Cherian S, Chen X, et
al: Immunotherapy of non-Hodgkin's lymphoma with a defined ratio of
CD8+ and CD4+ CD19-specific chimeric antigen receptor-modified T
cells. Sci Transl Med. 8:355ra1162016. View Article : Google Scholar : PubMed/NCBI
|
|
227
|
Davila ML, Riviere I, Wang X, Bartido S,
Park J, Curran K, Chung SS, Stefanski J, Borquez-Ojeda O, Olszewska
M, et al: Efficacy and toxicity management of 19-28z CAR T cell
therapy in B cell acute lymphoblastic leukemia. Sci Transl Med.
6:224ra252014. View Article : Google Scholar : PubMed/NCBI
|
|
228
|
Mestermann K, Giavridis T, Weber J, Rydzek
J, Frenz S, Nerreter T, Mades A, Sadelain M, Einsele H and Hudecek
M: The tyrosine kinase inhibitor dasatinib acts as a pharmacologic
on/off switch for CAR T cells. Sci Transl Med. 11:eaau59072019.
View Article : Google Scholar : PubMed/NCBI
|
|
229
|
Varadarajan I and Lee DW: Management of
T-cell engaging immunotherapy complications. Cancer J. 25:223–230.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
230
|
Fukumura D, Xavier R, Sugiura T, Chen Y,
Park EC, Lu N, Selig M, Nielsen G, Taksir T, Jain RK and Seed B:
Tumor induction of VEGF promoter activity in stromal cells. Cell.
94:715–725. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
231
|
Chinnasamy D, Yu Z, Theoret MR, Zhao Y,
Shrimali RK, Morgan RA, Feldman SA, Restifo NP and Rosenberg SA:
Gene therapy using genetically modified lymphocytes targeting
VEGFR-2 inhibits the growth of vascularized syngenic tumors in
mice. J Clin Invest. 120:3953–3968. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
232
|
Wang W, Ma Y, Li J, Shi HS, Wang LQ, Guo
FC, Zhang J, Li D, Mo BH, Wen F, et al: Specificity redirection by
CAR with human VEGFR-1 affinity endows T lymphocytes with
tumor-killing ability and anti-angiogenic potency. Gene Ther.
20:970–978. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
233
|
Slaney CY, Kershaw MH and Darcy PK:
Trafficking of T cells into tumors. Cancer Res. 74:7168–7174. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
234
|
van Schalkwyk MC, Papa SE, Jeannon JP,
Guerrero Urbano T, Spicer JF and Maher J: Design of a phase I
clinical trial to evaluate intratumoral delivery of ErbB-targeted
chimeric antigen receptor T-cells in locally advanced or recurre
head and neck cancer. Hum Gene Ther Clin Dev. 24:134–142. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
235
|
Sridhar P and Petrocca F: Regional
delivery of chimeric antigen receptor (CAR) T-cells for cancer
therapy. Cancers (Basel). 9:922017. View Article : Google Scholar : PubMed/NCBI
|
|
236
|
Milner JJ, Toma C, Yu B, Zhang K, Omilusik
K, Phan AT, Wang D, Getzler AJ, Nguyen T, Crotty S, et al: Runx3
programs CD8(+) T cell residency in non-lymphoid tissues and
tumors. Nature. 552:253–257. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
237
|
Caruana I, Savoldo B, Hoyos V, Weber G,
Liu H, Kim ES, Ittmann MM, Marchetti D and Dotti G: Heparanase
promotes tumor infiltration and antitumor activity of
CAR-redirected T lymphocytes. Nat Med. 21:524–529. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
238
|
Craddock JA, Lu A, Bear A, Pule M, Brenner
MK, Rooney CM and Foster AE: Enhanced tumor trafficking of GD2
chimeric antigen receptor T cells by expression of the chemokine
receptor CCR2b. J Immunother. 33:780–788. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
239
|
Maus MV, Haas AR, Beatty GL, Albelda SM,
Levine BL, Liu X, Zhao Y, Kalos M and June CH: T cells expressing
chimeric antigen receptors can cause anaphylaxis in humans. Cancer
Immunol Res. 1:26–31. 2013. View Article : Google Scholar
|
|
240
|
Till BG, Jensen MC, Wang J, Chen EY, Wood
BL, Greisman HA, Qian X, James SE, Raubitschek A, Forman SJ, et al:
Adoptive immunotherapy for indolent non-Hodgkin lymphoma and mantle
cell lymphoma using genetically modified autologous CD20-specific T
cells. Blood. 112:2261–2271. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
241
|
Ajina A and Maher J: Prospects for
combined use of oncolytic viruses and CAR T-cells. Review. J
Immunother Cancer. 5:902017. View Article : Google Scholar
|
|
242
|
Kim DS, Dastidar H, Zhang C, Zemp FJ, Lau
K, Ernst M, Rakic A, Sikdar S, Rajwani J, Naumenko V, et al: Smac
mimetics and oncolytic viruses synergize in driving anticancer
T-cell responses through complementary mechanisms. Nat Commun.
8:3442017. View Article : Google Scholar : PubMed/NCBI
|
|
243
|
Scott EM, Duffy MR, Freedman JD, Fisher KD
and Seymour LW: Solid tumor immunotherapy with T cell engager-armed
oncolytic viruses. Macromol Biosci. 18:17001872018. View Article : Google Scholar
|
|
244
|
Liu X, Ranganathan R, Jiang S, Fang C, Sun
J, Kim S, Newick K, Lo A, June CH, Zhao Y and Moon EK: A chimeric
switch-receptor targeting PD1 augments the efficacy of
second-generation CAR T cells in advanced solid tumors. Cancer Res.
76:1578–1590. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
245
|
Chmielewski M and Abken H: TRUCKS, the
fourth-generation CAR T cells: Current developments and clinical
translation. Adv Cell Gene Ther. 3:e842020. View Article : Google Scholar
|
|
246
|
Zhang H, Zhao P and Huang H: Engineering
better chimeric antigen receptor T cells. Exp Hematol Oncol.
9:342020. View Article : Google Scholar : PubMed/NCBI
|
|
247
|
Köksal H, Müller E, Inderberg EM, Bruland
Ø and Wälchli S: Treating osteosarcoma with CAR T cells. Scand J
Immunol. 89:e127412019. View Article : Google Scholar
|
|
248
|
Noordam L, Kaijen MEH, Bezemer K,
Cornelissen R, Maat LAPWM, Hoogsteden HC, Aerts JGJV, Hendriks RW,
Hegmans JPJJ and Vroman H: Low-dose cyclophosphamide depletes
circulating naïve and activated regulatory T cells in malignant
pleural mesothelioma patients synergistically treated with
dendritic cell-based immunotherapy. Oncoimmunology. 7:e14743182018.
View Article : Google Scholar
|
|
249
|
Ge Y, Domschke C, Stoiber N, Schott S,
Heil J, Rom J, Blumenstein M, Thum J, Sohn C, Schneeweiss A, et al:
Metronomic cyclophosphamide treatment in metastasized breast cancer
patients: Immunological effects and clinical outcome. Cancer
Immunol Immunother. 61:353–362. 2012. View Article : Google Scholar
|
|
250
|
Hu J, Sun C, Bernatchez C, Xia X, Hwu P,
Dotti G and Li S: T-cell homing therapy for reducing regulatory T
cells and preserving effector T-cell function in large solid
tumors. Clin Cancer Res. 24:2920–2934. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
251
|
Alizadeh D, Trad M, Hanke NT, Larmonier
CB, Janikashvili N, Bonnotte B, Katsanis E and Larmonier N:
Doxorubicin eliminates myeloid-derived suppressor cells and
enhances the efficacy of adoptive T-cell transfer in breast cancer.
Cancer Res. 74:104–118. 2014. View Article : Google Scholar
|
|
252
|
Seliger B and Quandt D: The expression,
function, and clinical relevance of B7 family members in cancer.
Cancer Immunol Immunother. 61:1327–1341. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
253
|
Murad JP, Tilakawardane D, Park AK, Lopez
LS, Young CA, Gibson J, Yamaguchi Y, Lee HJ, Kennewick KT, Gittins
BJ, et al: Pre-conditioning modifies the TME to enhance solid tumor
CAR T cell efficacy and endogenous protective immunity. Mol Ther.
29:2335–2349. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
254
|
Kohli K, Pillarisetty VG and Kim TS: Key
chemokines direct migration of immune cells in solid tumors. Cancer
Gene Ther. 29:10–21. 2022. View Article : Google Scholar :
|
|
255
|
Srivastava S, Furlan SN, Jaeger-Ruckstuhl
CA, Sarvothama M, Berger C, Smythe KS, Garrison SM, Specht JM, Lee
SM, Amezquita RA, et al: Immunogenic chemotherapy enhances
recruitment of CAR-T cells to lung tumors and improves antitumor
efficacy when combined with checkpoint blockade. Cancer Cell.
39:193–208.e10. 2021. View Article : Google Scholar
|
|
256
|
Motyka B, Korbutt G, Pinkoski MJ, Heibein
JA, Caputo A, Hobman M, Barry M, Shostak I, Sawchuk T, Holmes CF,
et al: Mannose 6-phosphate/insulin-like growth factor II receptor
is a death receptor for granzyme B during cytotoxic T cell-induced
apoptosis. Cell. 103:491–500. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
257
|
Ramakrishnan R, Huang C, Cho HI, Lloyd M,
Johnson J, Ren X, Altiok S, Sullivan D, Weber J, Celis E and
Gabrilovich DI: Autophagy induced by conventional chemotherapy
mediates tumor cell sensitivity to immunotherapy. Cancer Res.
72:5483–5493. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
258
|
Trapani JA, Sutton VR, Thia KYT, Li YQ,
Froelich CJ, Jans DA, Sandrin MS and Browne KA: A
clathrin/dynaminand mannose-6-phosphate receptor-independent
pathway for granzyme B-induced cell death. J Cell Biol.
160:223–233. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
259
|
Parente-Pereira AC, Whilding LM, Brewig N,
van der Stegen SJ, Davies DM, Wilkie S, van Schalkwyk MC,
Ghaem-Maghami S and Maher J: Synergistic chemoimmunotherapy of
epithelial ovarian cancer using ErbB-Retargeted T cells combined
with carboplatin. J Immunol. 191:2437–2445. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
260
|
Proietti E, Moschella F, Capone I and
Belardelli F: Exploitation of the propulsive force of chemotherapy
for improving the response to cancer immunotherapy. Mol Oncol.
6:1–14. 2012. View Article : Google Scholar
|
|
261
|
Senovilla L, Vitale I, Martins I, Tailler
M, Pailleret C, Michaud M, Galluzzi L, Adjemian S, Kepp O,
Niso-Santano M, et al: An immunosurveillance mechanism controls
cancer cell ploidy. Science. 337:1678–1684. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
262
|
Martins I, Tesniere A, Kepp O, Michaud M,
Schlemmer F, Senovilla L, Séror C, Métivier D, Perfettini JL,
Zitvogel L and Kroemer G: Chemotherapy induces ATP release from
tumor cells. Cell Cycle. 8:3723–3728. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
263
|
Garg AD, Galluzzi L, Apetoh L, Baert T,
Birge RB, Bravo-San Pedro JM, Breckpot K, Brough D, Chaurio R,
Cirone M, et al: Molecular and translational classifications of
DAMPs in immunogenic cell death. Front Immunol. 6:5882015.
View Article : Google Scholar : PubMed/NCBI
|
|
264
|
Sistigu A, Yamazaki T, Vacchelli E, Chaba
K, Enot DP, Adam J, Vitale I, Goubar A, Baracco EE, Remédios C, et
al: Cancer cell-autonomous contribution of type I interferon
signaling to the efficacy of chemotherapy. Nat Med. 20:1301–1309.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
265
|
Vierboom MP, Bos GM, Ooms M, Offringa R
and Melief CJ: Cyclophosphamide enhances anti-tumor effect of
wild-type p53-specific CTL. Int J Cancer. 87:253–260. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
266
|
Higgins JP, Bernstein MB and Hodge JW:
Enhancing immune responses to tumor-associated antigens. Cancer
Biol Ther. 8:1440–1449. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
267
|
Walle T, Martinez Monge R, Cerwenka A,
Ajona D, Melero I and Lecanda F: Radiation effects on antitumor
immune responses: Current perspectives and challenges. Ther Adv Med
Oncol. 10:17588340177425752018. View Article : Google Scholar : PubMed/NCBI
|
|
268
|
Apetoh L, Ghiringhelli F, Tesniere A,
Obeid M, Ortiz C, Criollo A, Mignot G, Maiuri MC, Ullrich E,
Saulnier P, et al: Toll-like receptor 4-dependent contribution of
the immune system to anticancer chemotherapy and radiotherapy. Nat
Med. 13:1050–1059. 2007. View
Article : Google Scholar : PubMed/NCBI
|
|
269
|
Crouse J, Kalinke U and Oxenius A:
Regulation of antiviral T cell responses by type I interferons. Nat
Rev Immunol. 15:231–242. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
270
|
Burnette BC, Liang H, Lee Y, Chlewicki L,
Khodarev NN, Weichselbaum RR, Fu YX and Auh SL: The efficacy of
radiotherapy relies upon induction of type I interferon-dependent
innate and adaptive immunity. Cancer Res. 71:2488–2496. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
271
|
Diamond MS, Kinder M, Matsushita H,
Mashayekhi M, Dunn GP, Archambault JM, Lee H, Arthur CD, White JM,
Kalinke U, et al: Type I interferon is selectively required by
dendritic cells for immune rejection of tumors. J Exp Med.
208:1989–2003. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
272
|
Fuertes MB, Kacha AK, Kline J, Woo SR,
Kranz DM, Murphy KM and Gajewski TF: Host type I IFN signals are
required for antitumor CD8+ T cell responses through CD8{alpha}+
dendritic cells. J Exp Med. 208:2005–2016. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
273
|
Weiss T, Weller M, Guckenberger M, Sentman
CL and Roth P: NKG2D-based CAR T cells and radiotherapy exert
synergistic efficacy in glioblastoma. Cancer Res. 78:1031–1043.
2018. View Article : Google Scholar
|
|
274
|
Matsumura S, Wang B, Kawashima N,
Braunstein S, Badura M, Cameron TO, Babb JS, Schneider RJ, Formenti
SC, Dustin ML and Demaria S: Radiation-induced CXCL16 release by
breast cancer cells attracts effector T cells. J Immunol.
181:3099–3107. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
275
|
Zhang B, Bowerman NA, Salama JK, Schmidt
H, Spiotto MT, Schietinger A, Yu P, Fu YX, Weichselbaum RR, Rowley
DA, et al: Induced sensitization of tumor stroma leads to
eradication of established cancer by T cells. J Exp Med. 204:49–55.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
276
|
Ganss R, Ryschich E, Klar E, Arnold B and
Hämmerling GJ: Combination of T-cell therapy and trigger of
inflammation induces remodeling of the vasculature and tumor
eradication. Cancer Res. 62:1462–1470. 2002.PubMed/NCBI
|
|
277
|
Yovino S and Grossman SA: Severity,
etiology and possible consequences of treatment-related lymphopenia
in patients with newly diagnosed high-grade gliomas. CNS Oncol.
1:149–154. 2012. View Article : Google Scholar
|