Introduction
The incidence of diabetes mellitus is increasing
annually. Diabetic nephropathy (DN) is one of the most common and
serious chronic complications of diabetes mellitus, and is one of
the main causes of chronic renal failure. The main pathological
changes in DN include hypertrophy and thickening of the glomerular
basilemma and renal tubules, extracellular matrix (ECM)
accumulation in the mesangium and glomerulosclerosis. The
pathogenesis is complicated and is related to many factors, such as
glomerular hemodynamic changes, disturbance of biochemical
metabolism, oxidative stress (OS), cell factors and genetic
predisposition. Compared to other kidney diseases, DN has a poor
prognosis and is more difficult to treat, the reasons for which
have not yet been fully understood. Increasing evidence has shown
that OS plays an important role in DN (1). Kidney cells produce a mass of
reactive oxygen species (ROS) and the decrease in anti-oxidants
leads to OS. In addition, ROS directly attack unsaturated fatty
acids in the body to increase vascular permeability (2), which is closely related to DN,
destroy cell membranes in kidney tissues, activate protein kinase
and mitogen-activated protein kinase, initiate cytokines and
transcription of growth factors, promote proliferation and
hypertrophy of cells in kidney tissue and increase ECM synthesis.
They also decrease ECM degradation and indirectly injure kidney
tissues, resulting in the pathogenic mechanisms of DN.
Adiponectin (ADPN), also known as apM1, Acrp30,
GBP28 or AdipoQ, is a recently discovered specific protein secreted
by adipose cells; it is abundant in the blood circulation and makes
up approximately 0.01% of the total plasma protein. In addition,
ADPN is the most abundant adipose tissue-secreted protein in human
plasma. Many studies have shown that ADPN is anti-inflammatory,
improves insulin resistance, is anti-atherosclerotic and decreases
blood sugar and blood fat levels (3). Previous studies have shown that ADPN
has anti-oxidant effects and that its concentration in the blood
plasma is closely related to DN. Early studies investigating DN
have shown that circulatory ADPN concentration is negatively
correlated with urine protein, indicating that impaired endothelial
function is related to low ADPN concentration in early DN (4). ADPN concentrations in the urine and
blood of patients with progressive-stage DN increase (5,6)
(possibly due to the anti-inflammatory and anti-atherosclerotic
effects) to abate renal impairment and prevent the adjustment of
positive and negative feedback in DN progression (7). Therefore, the present study aimed to
investigate the effects of ADPN on OS in human glomerular mesangial
cells (HMCs) through the induction of high glucose (HG) levels. The
main objective of the present study was to investigate the
protective effects of ADPN on DN and to determine which signaling
pathway is involved in order to clarify the mechanism of its
inhibition of diabetic microangiopathy.
Materials and methods
HMC culture
The HMCs preserved in our research institute were
provided by Professor Mei Changlin of the Second Military Medical
University of the Chinese People’s Liberation Army. The HMCs were
cultured at 37°C using RPMI-1640 culture medium (Gibco) containing
10% fetal cattle serum in a 5% CO2 incubator. The cells
were digested and subcultured with 0.25% pancreatin 2–3 days later.
When the cells grew to 85% fusion, the HMCs were inoculated into a
6-pore plate with a density of 1×106 cells/ml and
cultured in the logarithmic growth phase after 24 h. The culture
medium was then changed to serum-free medium and used 12 h later in
a following experiment.
Experimental grouping
Four groups of HMCs were divided randomly into 5
wells/group. The groups were as follows: control group, HG group,
HG + globular ADPN group (HG + gAd group) and HG + gAd + adenine
arabinoside group (HG + gAd + AraA group). A 30-mM concentration of
culture fluid containing D-glucose was applied to the HG group. The
HG + gAd group was exposed to 10 μg/ml gAd (R&D Co.) for 30
min, to which a 30-mM concentration of D-glucose was added. The HG
+ gAd + AraA group (AraA, an inhibitor of AMP-activated protein
kinase (AMPK), was purchased from Sigma Co.) received HG and gAd
after 30 min of pre-treatment with 2 mmol/l AraA.
Determination of ROS levels
Following 24-h culture of the cells in all groups,
the orthotropic loading probe was applied according to the
manufacturer’s instructions. After removing the culture fluid, 1 ml
diluted DCFH-DA solution was added into each well to produce a
10-μM terminal concentration. The RPMI-1640 serum-free culture
medium was used to wash the cells three times after incubation at
37°C for 20 min. This was performed to completely remove the probes
that were unable to enter the cells. Using a fluorescence
spectrophotometer, the cells were collected to determine the
fluorescence intensity (FI). The excitation and emission wavelength
were 488 and 525 nm, respectively.
Determination of the transcription and
protein expression of endothelial nitric oxide (NO) synthase
(eNOS)
Cell culture continued for 16 h, after which the
cells were collected. TRIzol reagent was used to extract the total
RNA, after which 1 μg total RNA was removed using an RT-PCR kit.
This was carried out to synthesize cDNA and conduct PCR, while
amplifying eNOS and the reference gene, β-actin, at the same time.
The eNOS properties were as follows: forward primer,
5′-GAGTCCTCACCGCCTTCTC-3′; reverse primer,
5′-AGGAAGCGGGTGGCAGTA-3′; PCR products, 626 bp. The β-actin
properties were as follows: forward primer,
5′-CACCCTGAAGTACCCCATCG-3′; reverse primer,
5′-TTGCCAATGGTGATGACCTG-3′; PCR products, 561 bp. The amplification
conditions were as follows: the annealing temperature of eNOS was
54°C and that of β-actin was 58°C at 35 cycles. The PCR products
were subjected to electrophoresis using a 1.2% agarose gel and then
photographed following ethidium bromide staining. The PCR products
were sequenced using a 3730 sequencer.
For western blot analysis, 40 μg of total cellular
protein were extracted and separated using 8% SDS-PAGE gel
electrophoresis; the protein was also hybridized with polyclonal
eNOS antibody after being transferred onto a nitrocellulose
membrane. To detect the protein expression of eNOS, coloration was
performed using the ECL method and exposure to Kodak film.
Quantitative analysis was conducted using UVP. The experiment was
repeated 4 times, and the results are expressed as arbitrary units
(AU) (AU = target protein/β-actin ×100%).
Analysis of AMPK
expression/phosphorylation
The HMCs in the RPMI-1640 culture medium, which
contained 10% fetal cattle serum, were inoculated into a 6-pore
plate. To culture the cells to 85% fusion, the culture medium was
changed to serum-free medium and then 10 μg/ml recombinant human
ADPN were added. Following cell activation for 0, 5, 10, 15 and 30
min, the cytoplasmic protein was collected to detect the AMPK
expression/phosphorylation (the phospho-AMPKα rabbit polyclonal
antibodies were purchased from Cell Signaling Co.).
Statistical analysis
The SPSS11.0 software was used, and the experimental
results are expressed as the means ± SD. Comparisons between the
groups were performed using one-way analysis of variance and the
t-test. P<0.05 indicated a statistically significant
difference.
Results
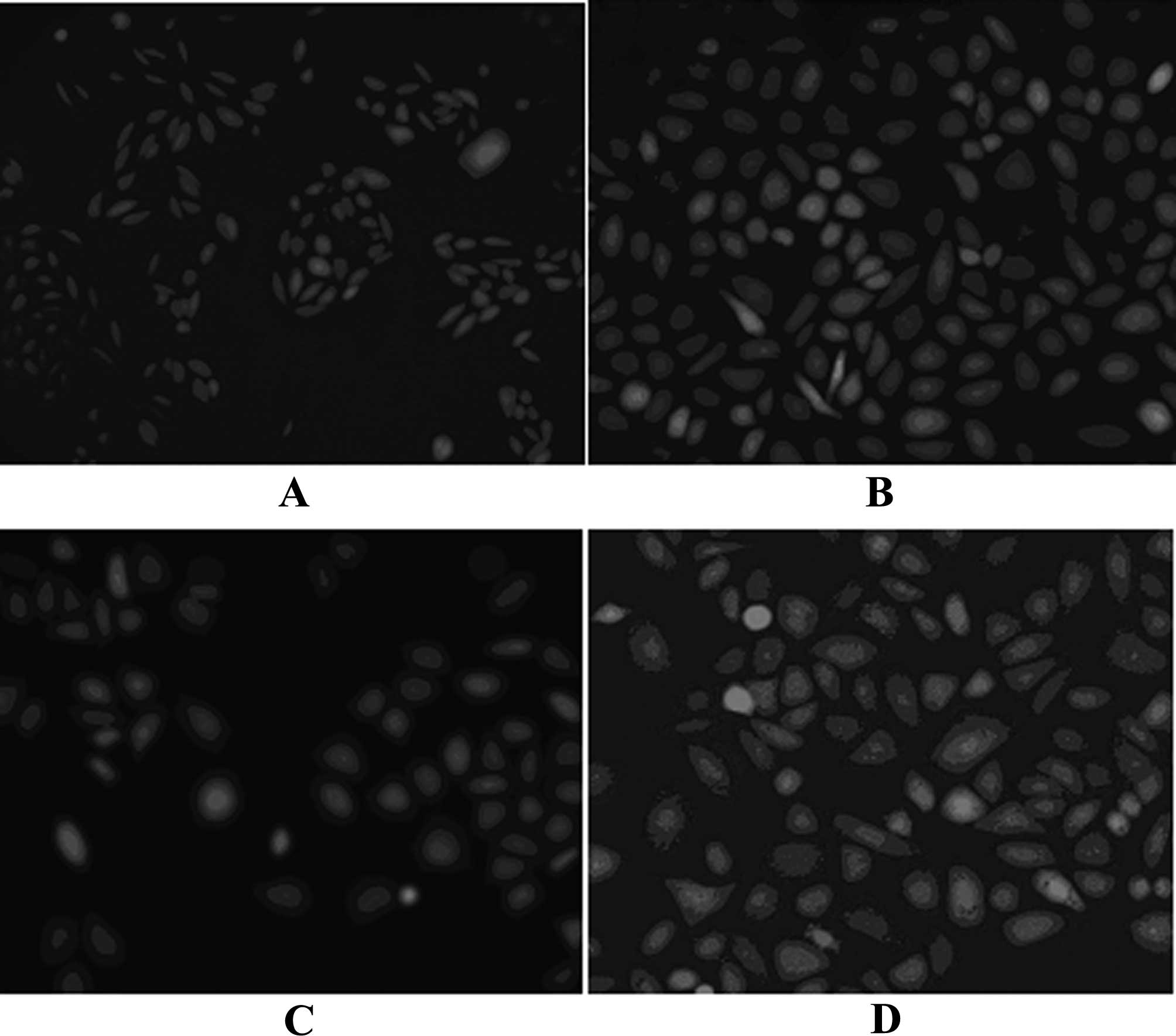
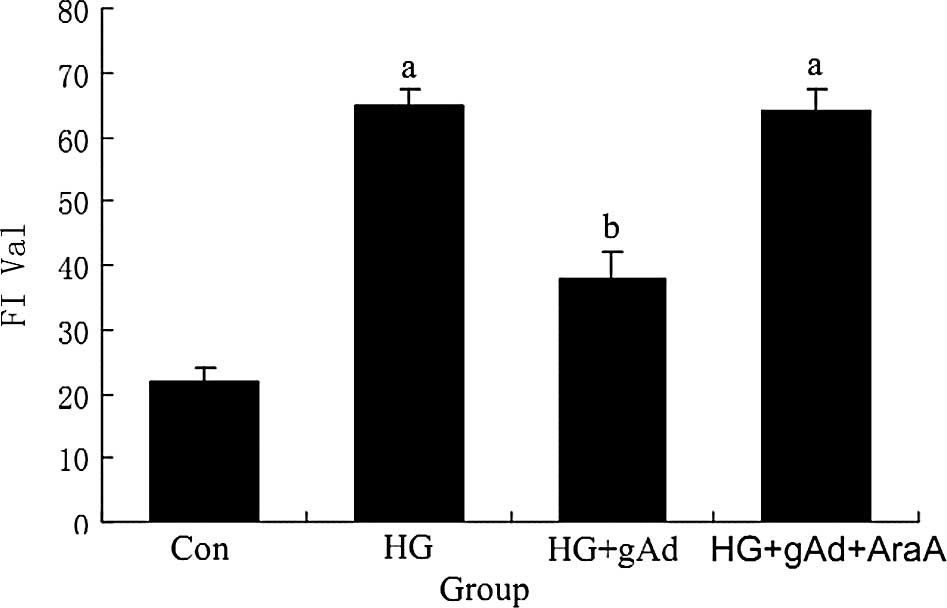
Effects of ADPN on the production of
ROS
DCFH-DA is lipotropic and freely diffuses into cells
and is hydrolyzed as hydrophilic DCFH by esterase in order to
remain in the neoplasm. DCFH is easily oxidized as epoptic
2′,7′-dichlorofluorescein (DCF) by peroxide (mainly hydrogen
peroxide) in the neoplasm. The amount of DCF is directly related to
the peroxide in the neoplasm. The experimental results revealed
that after 24 h under the influence of a high concentration of
glucose, the FI of the HMCs was significantly enhanced compared to
that of the control group (p<0.05), proving that HG levels
stimulated the HMCs to produce ROS. Compared to the HG group, the
fluorescence of the HG + gAd group was significantly weakened
(p<0.05), indicating that gAd inhibited the production of ROS.
When AraA was added, the effects of the inhibition of ADPN on ROS
were weakened; no significant difference was found compared to the
HG group (p>0.05). Thus, ADPN inhibited the production of ROS
under the stimulation of HG, and this effect was partly inhibited
by AraA (Figs. 1 and 2).
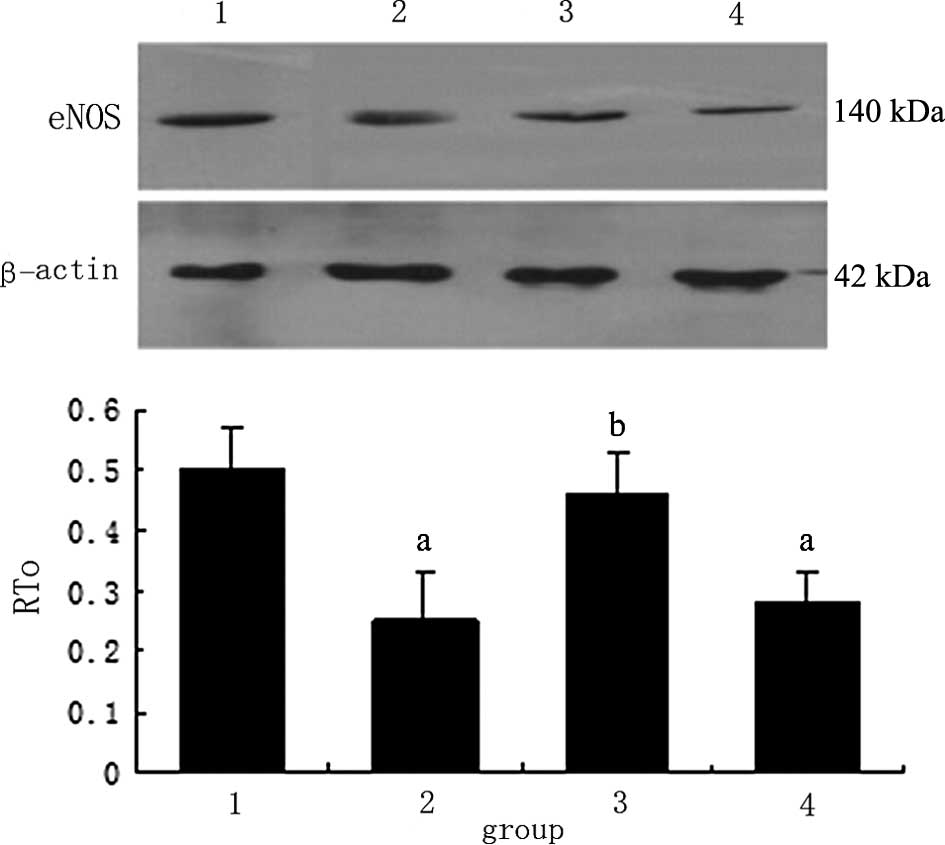
Effects of ADPN on the production of
eNOS
RT-PCR and western blot analysis results revealed
that, after 16 h of stimulation with 30 mM of D-glucose, the
expression of eNOS and mRNA was downregulated by 80.2% and the
protein expression was decreased by 35.3% (p<0.05) compared to
the control group. The expression of eNOS and mRNA in the HG + gAd
group was upregulated by 50.5% and that of protein increased by
30.2% (p<0.05) compared to the HG group. Moreover, after AraA
was added, the upregulation of eNOS and mRNA decreased; compared to
the HG group, no significant difference was found, indicating that
ADPN promoted the production of eNOS and subsequently protected the
endothelium (Figs. 3 and 4).
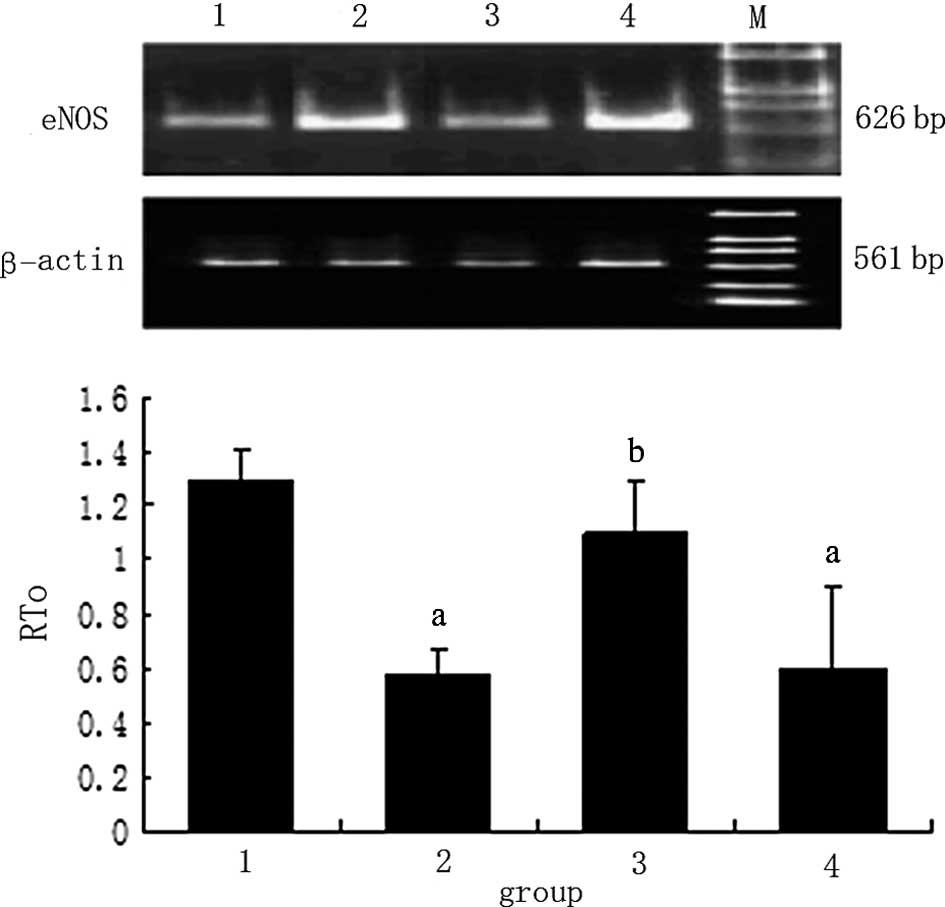 | Figure 3Human glomerular mesangial cell (HMC)
eNOS mRNA expression detected by RT-PCR. Lane 1, control group;
lane 2, HG group; lane 3, HG + gAd group; and lane 4, HG + gAd +
AraA group. M, DL2000 marker, compared to the control group
(ap<0.05), and compared to the HG group
(bp<0.05). eNOS, endothelial nitric oxide synthase;
HG, high glucose; gAd, globular adiponectin; AraA, adenine
arabinoside. |
Effects of ADPN on AMPK
expression/phosphorylation
ADPN stimulated the AMPK
expression/phosphorylationin HMCs in a time-dependent manner. The
UVP quantitative analysis revealed that without the stimulation of
ADPN, the AU value was 0.10 μg/ml. In addition, after the
stimulation of recombinant human ADPN for 5, 10, 15 and 30 min, the
AU values were 06.8±50.9, 487.9±92.2, 943.6±103.9 and 399.8±62.2,
respectively, peaking 15 min after the stimulation (Fig. 5). The stimulatory effect of ADPN on
the AMPK expression/phosphorylation was completely inhibited by
pre-treatment with AraA (2 mM) for 30 min.
Discussion
DN is a severe microangiopathy complication of
diabetes mellitus, and many studies have investigated its
pathogenesis. Multiple factors are attributed to DN and, among
these, hyperglycemia is considered an important initiating factor.
Nishikawa et al (8)
reported that hyperglycemia induces the increase of ROS from the
respiratory chain. In a diabetes mellitus state, (nicotinamide
adenine dinucleotide phosphate NADPH) oxidase in the kidney tissue
is activated by the stimulations of many extracellular signals to
mediate ROS generation (9,10). The study by Xu et al
(11) showed that in the kidney
tissues of diabetic rats, hyperglycemia decreased the production of
NADPH, lowered the intracellular glutathione (GSH) levels and
weakened the anti-oxidation of kidney tissue through the activation
of the protein kinase A, which in turn, reduced the activity of
glucose 6-phosphate dehydrogenase (G6PD) produced by NADPH. Ha
et al (12) proved that in
mesangial and renal tubular epithelial cells, hyperglycemia induces
DCF-sensitive ROS in a time-dependent manner. In accordance with
the above results, the present study found that the production of
ROS in HMCs increased and that the mesangial cells served as an
important glomerular source of ROS. Thus, under the conditions of
diabetes mellitus, the OS state resultes from the increase in ROS
production or the decrease in the clearing of ROS, which by itself
attacks the lipids, proteins and DNA to damage the kidney. Previous
animal experimental research has indicated that treatment with
anti-oxidants reduces proteinuria, abates pathological damage of
the kidney and effectively delays the formation and development of
DN (13,14).
ADPN is a type of specific protein secreted by
adipose cells; it is abundant in circulated blood and is excreted
though urine. Several studies have shown that ADPN has an
anti-inflammatory effect, improves insulin resistance and possesses
anti-atherosclerotic and anti-oxidative capabilities. It also
inhibits the stimulation of oxidized low-density lipoprotein for
the production of ROS in endothelial cells (15), increases the activity of eNOS and
protects the blood vessel endothelium from injury by TNF-α
(16). The results from the
present study prove that employing ADPN to stimulate mesangial
cells in vitro inhibits the stimulatory effect of HG on the
production of ROS. NO is the active substance in the blood vessels
released from endothelial cells. In the human body, NOS catalyzes
l-arginine to produce NO. eNOS is one of the main rate-limiting
enzymes, whose activity is positively correlated with the amount of
NO produced. eNOS continually produces a small amount of NO to
relax the smooth muscle; it also inhibits the proliferation of
endothelial cells, platelet aggregation and the production of ROS,
thereby decreasing OS of the endothelial cells.
In the later stages of DN, the production of eNOS
decreases and affects the normal function of NO, thus increasing
the production of superoxide (17). In the present study, we found that
the HMCs produced eNOS. During the stimulation of HG, the
production of eNOS clearly decreased; however, the expression of
eNOS and protein secretion evidently increased with the
pre-addition of ADPN, thereby protecting the cells. The mechanism
was possibly related to the inhibition of the production of ROS in
the cells brought about by ADPN. At present, the function of ROS in
signal transduction has received increasing attention from
investigators. ROS increase the production of ECM, injure DNA of
the endothelial cells and regulate the expression of apoptotic
genes (18) by activating the
signal transduction cascade waterfall and transcription factor and
by upregulating the gene expression of transforming growth factor
(TGF)- β1, angiotensin II (Ang II), monocyte chemoattractant
protein-1 (MCP-1) and plasminogen activator inhibitor type 1
(PAI-1). In our study, we found that ADPN upregulated the
expression of eNOS by inhibiting the production of ROS; however,
the function disappeared when a AMPK blocker was added.
AMPK is a type of protein kinase existing in the
tissues of most mammals; it perceives changes in the state of
cellular energy metabolism and maintains the balance of the supply
and demand of cellular energy by influencing many links to cellular
material metabolism. Many factors activate AMPK in mammalian cells,
and these factors, such as hypoxia, ischemia and lack of glucose,
mainly function by increasing the consumption of cellular adenosine
triphosphate (ATP) and inhibiting the production of ATP. Yamauchi
et al (19) reported that
the mechanism of the signal transduction of ADPN involves the
activation of AMPK. In skeletal muscle and hepatic cells, ADPN
activates AMPK to reduce blood sugar. Another study has proven that
AMPK possibly mediates the signal transmission of ADPN in
endothelial cells (20).
Activation of AMPK promotes fatty acid oxidation and net synthesis
of ATP. In relation to this, the activation of eNOS provides a
potential signal connection for the production of ADPN and NO. AMPK
plays a vital role in the downstream signal transduction pathways
of ADPN receptors and even in maintaining a steady blood glucose
level in an organism. In this study, ADPN inhibited the production
of ROS in mesangial cells and promoted the increased AMPK
expression/phosphorylation in a time-dependent manner. Applying
AraA as pre-treatment inhibited the phosphorylation of AMPK, partly
inhibited the production of eNOS and weakened the inhibition of
ADPN in ROS production during the induction of HG levels. AMPK was
also found to be included in the signaling pathways modulated by
ADPN. Based on studies on liver and skeletal muscle cells, the
anti-oxidation of ADPN on mesangial cells was at least partly
realized through the phosphorylation of AMPK.
In conclusion, the present study reveals that ADPN
protects HMCs against the production of ROS induced by HG levels
through anti-oxidation. Our results may provide another possible
mechanism for preventing DN by using ADPN. Further studies on the
signaling pathways modulated by ADPN and its functional mechanism
in the body would provide new ways to prevent and cure DN.
Acknowledgements
This study was supported by the project of the Hunan
Provincial Health Bureau (B2006-043).
References
|
1
|
Stojiljkovic L and Behnia R: Role of renin
angiotensin system inhibitors in cardiovascular and renal
protection: a lesson from clinical trials. Curr Pharm Des.
13:1335–1345. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Suzuki S, Hinokio Y, Komatu K, et al:
Oxidative damage to mitochondrial DNA and its relationship to
diabetic nephropathy. Diabetes Res Clin Pract. 45:161–168. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Okamoto Y, Kihara S, Ouchi N, et al:
Adiponectin reduces atherosclerosis in apolipoprotein E-deficient
mice. Circulation. 106:2767–2770. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Yilmaz MI, Saglam M, Qureshi AR, et al:
Endothelial dysfunction in type-2 diabetics with early diabetic
nephropathy is associated with low circulating adiponectin. Nephrol
Dail Transplant. 23:1621–1627. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Saraheimo M, Forsblom C and Fagerudd J:
Serum adiponectin is increased in type 1 diabetic patients with
nephropathy. Diabetes Care. 28:1410–1414. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Schalkwijk CG, Chaturvedi N, Schram MT, et
al: Adiponectin is inversely associated with renal function in type
1 diabetic patients. J Clin Endocrinol Metab. 91:129–135. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Fujita H, Morii T, Koshimura J, et al:
Possible relationship between adiponectin and renal tubular injury
in diabetic nephropathy. Endocr J. 53:745–752. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Nishikawa T, Edelstein D and Du XL:
Normalizing mitochondrial superoxide production blocks three
pathways of hyperglycaemic damage. Nature. 404:787–790. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Stokes KY, Clanton EC, Russell JM, et al:
NAD (P) H oxidase-derived superoxide mediates
hypercholesterolemia-induced leukocyte-endothelial cell adhesion.
Circ Res. 88:499–505. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Rueckschloss U, Galle J, Holtz J, et al:
Induction of NAD(P)H oxidase by oxidized low-density lipoprotein in
human endothelial cells: antioxidative potential of hydroxyme
thylglutaryl coenzyme A reductase inhibitor therapy. Circulation.
104:1767–1772. 2001. View Article : Google Scholar
|
|
11
|
Xu Y, Osborne BW and Stanton RC: Diabetes
causes inhibition of glucose-6-phosphate dehydrogenase via
activation of PKA, which contributes to oxidative stress in rat
kidney cortex. Am J Physiol Renal Physiol. 289:1040–1047. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Ha H, Yang Y, Lee HB, et al: Mechanisms of
reactive oxygen species generetion in LLC2PK1 cells cultured under
high glucose. J Am Soc Nephrol. 13:531A2002.
|
|
13
|
Davila-Esqueda ME, Vertiz-Hernandez AA and
Martinez-Morales F: Comparative analysis of the renoprotective
effects of pentoxifylline and vitamin E on streptozotocin-induced
diabetes mellitus. Ren Fail. 27:115–122. 2005.PubMed/NCBI
|
|
14
|
Chen L, Jia RH, Ding GH, et al: Effect of
irbesartan on oxidative stress and the activity of protein kinase C
in type 2 diabetic rats. Wu Han Da Xue Xue Bao (Yi Xue Ban).
25:150–153. 2004.
|
|
15
|
Motoshima H, Wu X, Mahadev K, et al:
Adiponectin suppresses proliferation an superoxide generation and
enhances eNOS activity in endothelial cells treated with oxidized
LDL. Biochem Biophys Res Commun. 315:264–271. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Ge Q, Deng HC and Liu JB: Effect of
adiponectin on endothelial function. Chin J Endocrinol Metab.
1:15–18. 2006.
|
|
17
|
Pritchard KA Jr, Groszek L, Smalley DM, et
al: Native low-density lipoprotein increases endothelial cell
nitric oxide synthase generation of superoxide anion. Circ Res.
77:510–518. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Ha H and Lee HB: Reactive oxygen species
amplify glucose signalling in renal cells cultured under high
glucose and in diabetic kidney. Nephrology. 1:7–10. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Yamauchi T, Kamon J, Ito Y, et al: Cloning
of adiponectin receptors that mediate antidiabetic metabolic
effects. Nature. 423:762–769. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Chen H, Montagnani M, Funahashi T, et al:
Adiponectin stimulates production of nitric oxide in vascular
endothelial cells. J Biol Chem. 278:45021–45026. 2003. View Article : Google Scholar : PubMed/NCBI
|