Introduction
Alzheimer’s disease (AD) is a neurodegenerative
disorder resulting in progressive cognitive impairment. It has been
reported that AD is the most common form of dementia among older
people and the worldwide prevalence of the disease is estimated at
>24 million cases (1). Medical
treatment for AD patients is placing an increasing burden on
physicians and families every year. Clinically, there are a variety
of drugs available for AD, such as cholinesterase inhibitors,
glutamate receptor antagonist and free radical scavengers. However,
these drugs cannot target the pathogenesis of the disease closely
and have significant side-effects (2). Therefore, it is important to find a
new type of drug and to clarify the mechanism of AD
pathophysiology. Caffeine is one of the most widely consumed
psychoactive substances in the world (3). Recently, studies have demonstrated
that caffeine intake may reduce the cognitive impairment in elderly
patients and the risk of AD in later life (4,5). It
also has been revealed that AD patients consume markedly less
caffeine than people without AD (6). Elevated levels of β-amyloid (Aβ) in
the brain and progressive cognitive impairment are the main
characteristics of AD. Several studies have indicated that caffeine
intake (1.5 mg/day) may reverse cognitive impairment and decrease
brain Aβ levels in aged AD mice (7,8).
Brain neurotrophic derived factor (BDNF), a member
of the neurotrophin family, is essential for growth, survival and
the differentiation of neurons. Furthermore, BDNF is involved in
learning and memory by binding to its main functional receptor
(TrkB), in the hippocampus, cortex and basal forebrain (9). The levels of BDNF and TrkB have been
reported to be lower in AD patients (10,11).
It has been demonstrated that BDNF signaling, through TrkB, is
involved in the pathophysiology and cognitive deficits of AD
(12). PS1/APP double transgenic
mice expressing the human APPswe and PS1-A246E mutations are a
widely used AD model which may imitate the main pathophysiology
process of AD. The present study was conducted in order to
investigate the effect of varying caffeine doses on memory
impairment and the expression of brain BNDF and TrkB in PS1/APP
double transgenic mice.
Materials and methods
Drugs
Caffeine (lot number, 1001176428) was purchased from
Sigma Corporation (St. Louis, MO, USA).
Animals
PS1/APP double transgenic mice (genetic background
C57BL/6J), containing the human APPswe and PS1-A246E mutations,
were obtained from the Institute of Laboratory Animals at the
Chinese Academy of Medical Sciences (Beijing, China). Wild-type
C57/BL6J mice were used as controls. All mice were housed in the
Laboratory Animal Center of Liaoning Medical University (Jinzhou,
Liaoning, China). All mice were maintained in an air-conditioned
room with a 12-h light and 12-h dark cycle, fed a standard diet and
water was available ad libitum. The ethical approval for
this study was obtained from the Ethics Committee of the Liaoning
Medical University.
Caffeine treatment
In this study, 24 PS1/APP double transgenic mice
were randomly divided into three groups (age, 24 months; n=8) and
0.3 ml/day of saline (Tg-control), 1.5 mg/day of caffeine (Caff-H)
and 0.75 mg/day of caffeine (Caff-L) were administered into the
stomachs of mice in the three groups, respectively. Furthermore,
eight wild-type C57/BL6J (NT) mice were administered 0.3 ml/day of
saline at the same time (WT). All mice were treated for eight
weeks.
Water maze
At the end of the seventh week of the experiment, a
water maze experiment was performed in order to evaluate the
spatial reference memory of the mice. We used a circular inflatable
pool (diameter 120 cm; height 90 cm) filled with opaque water,
containing a submerged escape platform (diameter, 9 cm) 2.0 cm
below the surface of the water. The water was maintained at a
constant temperature throughout the experiment (25±0.5°C) and the
pool was divided into four equal quadrants by black lines drawn on
the floor of the pool. The experiment was divided into place
navigation and spatial probe tests. The place navigation test
lasted for four days. During this test, mice were randomly placed
into one of the four equal quadrants of the pool. Mice were allowed
to search for the platform for 60 sec. If the mice did not locate
the platform within 60 sec, they were guided to it and left on the
platform for 10 sec. The latency time (the time from entering the
water to standing on the platform) was recorded. The spatial probe
test was performed on the fifth day of the experiment. The platform
was removed from the pool and the mice were placed into the water
at any location. The mice were allowed to swim in the maze for 60
sec. Swimming time, the time that the mice spent in target quadrant
(where the platform was), was recorded.
Western blotting
At the end of the experiment, the mice were
decapitated and the brains were rapidly removed on ice, the
hippocampus was quickly dissected and stored at −70°C until
required. The hippocampal tissue was homogenized in a lysis buffer
(50 mmol/l Tris-HCl, 5 mmol/l EDTA, 1% sodium deoxycholate, 150
mmol/l NaCl, 0.5% Triton X-100, 500 μmol/l
Na3VO4, 10 μmol/l AEBSF, 10 mmol/l NaF). The
homogenates were subsequently centrifuged at 12,000 × g for 10 min
at 4°C, and the supernatants were collected for protein
concentration determination using a protein assay (Bio-Rad,
Hercules, CA, USA). Equal amounts of protein extract were added to
8% SDS-polyacrylamide gel and transferred onto a PVDF membrane.
Western blotting was performed using rabbit anti-BDNF and anti-TrkB
[1:1,000, SC546, Santa Cruz Biotechnology, Inc. (Santa Cruz, CA,
USA); 1:1,000, ab38306, Abcam (Cambridge, MA, USA)] followed by a
chemiluminescence substrate (32109, ECL Plus, Amersham; kit
contents: luminol/enhancer, 25 ml, stable peroxide buffer, 25 ml)
and quantified. The BDNF and TrkB antisera detected the distinct
mature form of BDNF (mBDNF) and TrkB bands as described previously
(13). Recombinant human BDNF and
TrkB (BioVision, Mountain View, CA, USA) were added to act as a
positive control. The blots were stripped and reprobed with
anti-actin (1:4,000) to control loading variations. Quantity One
software (Bio-Rad) was used to quantify the protein bands. The
results were expressed as the means ± SEM of the ratio of
immunoreactivity normalized by β-actin.
Statistical analysis
Statistical analysis was performed using the
Student’s t-test and one-way ANOVA followed by Newman-Keuls
multiple comparisons test. P<0.05 was considered to indicate a
statistically significant difference.
Results
In order to investigate the effect of caffeine
treatment on spatial learning and memory of PS1/APP double
transgenic mice, a water maze test was performed. The results are
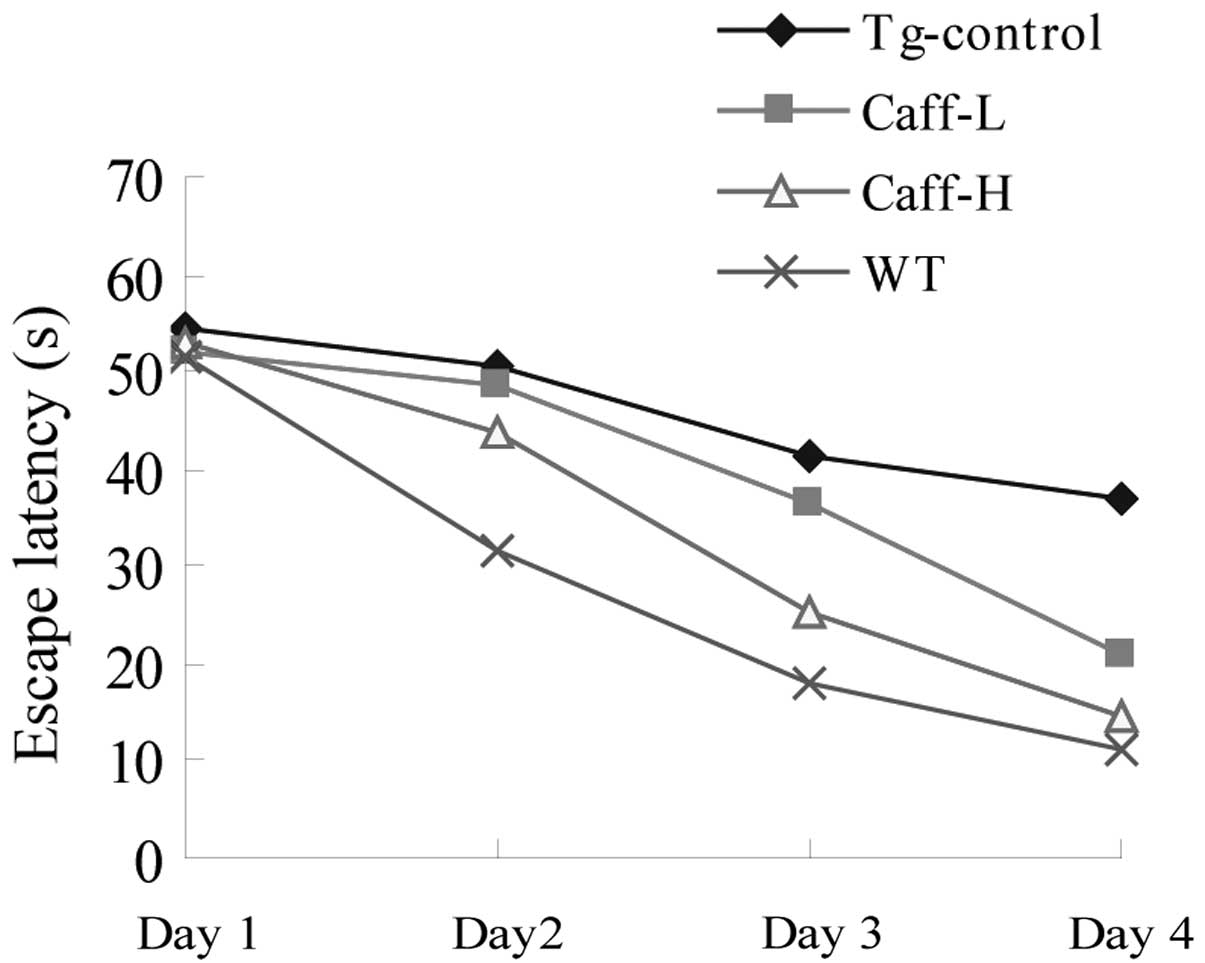
shown in Figs. 1 and 2. As shown in Fig. 1, the Tg-control mice took
significantly longer to locate the platform than the WT mice
(P<0.01), suggesting that the ability for spatial learning had
decreased in the AD model mice. Administering varying doses of
caffeine may significantly decrease the escape latency time of
PS1/APP mice (P<0.01). The escape latency time of Caff-H mice
was significantly higher compared with that of the Caff-L mice
indicating that there is a dose-response effect for caffeine
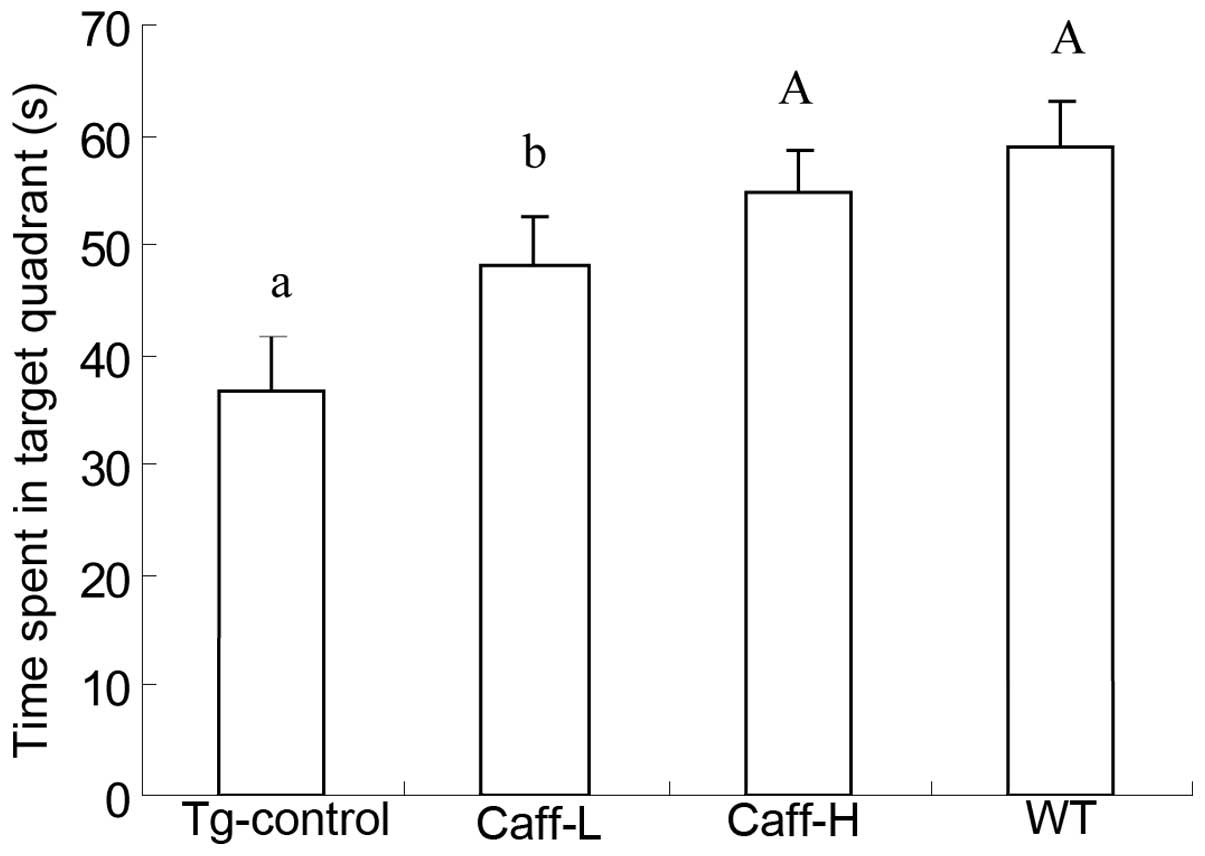
treatment. For the spatial probe test (Fig. 2), the results demonstrated that
caffeine treatment could significantly decrease the time that the
PS1/APP mice spent in the target quadrant (P<0.01), and Caff-H
mice spent longer in the target quadrant than Caff-L mice
(P<0.05). The water maze test revealed that caffeine treatment
was capable of significantly increasing the ability of spatial
learning and memory in PS1/APP mice with a dose-response
effect.
In order to determine the effect of caffeine
treatments on the expression of hippocampal BDNF and TrkB in mice,
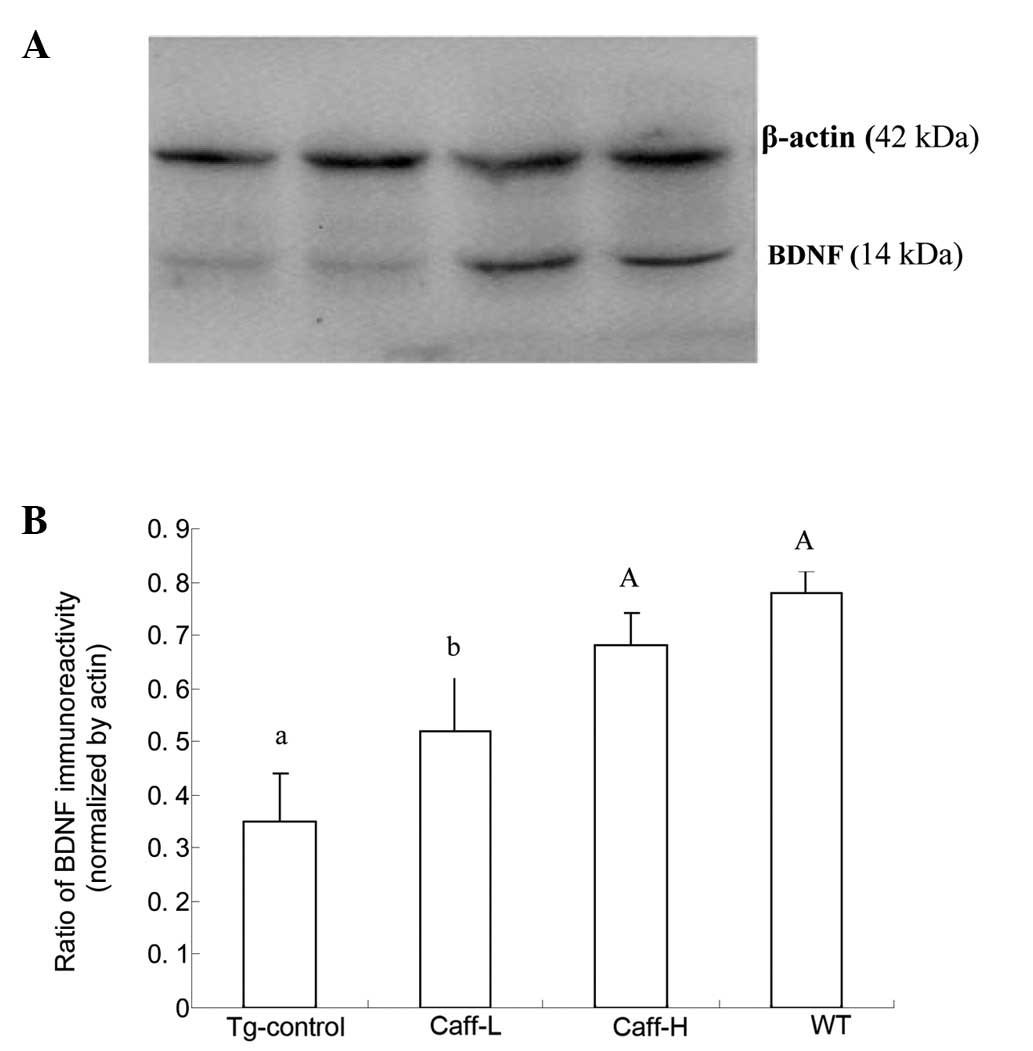
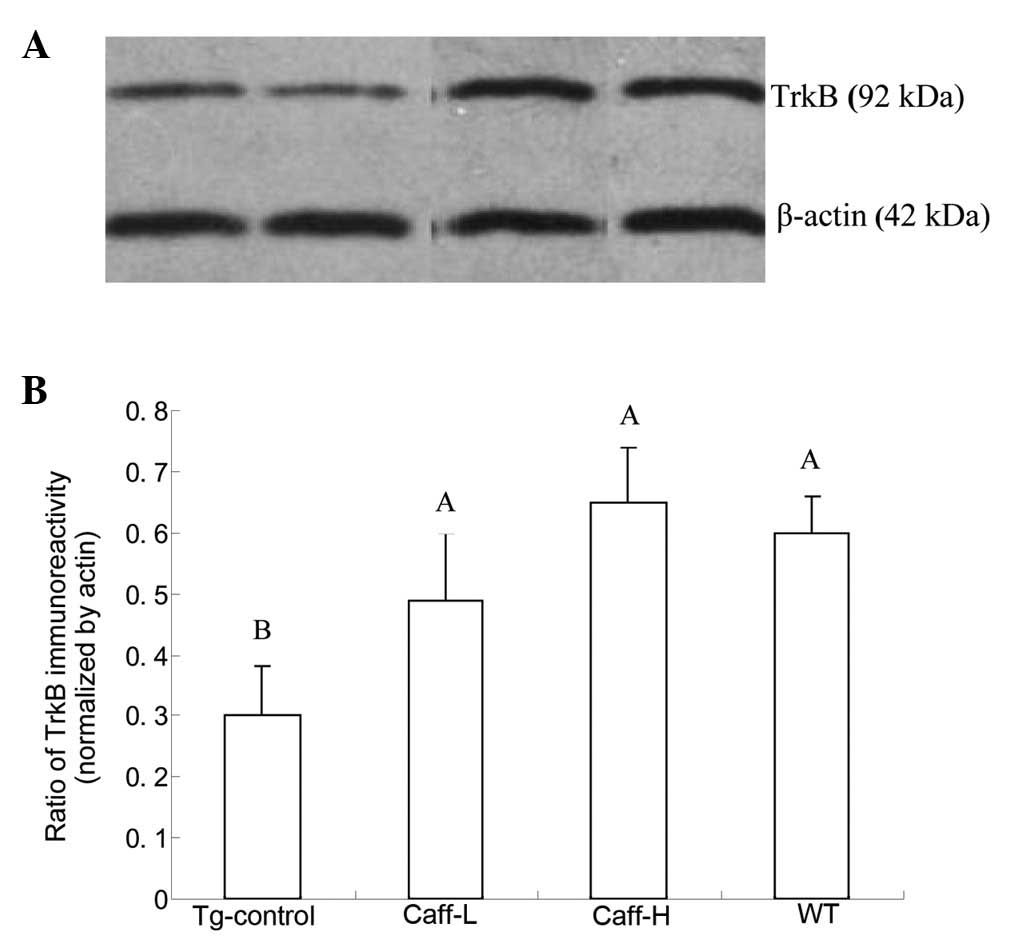
western blotting was performed. The results are shown in Figs. 3 and 4. The expression of hippocampal BDNF and
TrkB of PS1/APP mice treated with saline was significantly lower
compared with that of WT mice (P<0.01). It was also demonstrated
that caffeine treatment significantly increased the expression of
hippocampal BDNF and TrkB, and a high dose caffeine treatment
obtained a higher level of expression than a low dose of caffeine
(P<0.05).
Discussion
Several studies have demonstrated that caffeine
intake (1.5 mg/day) is capable of reversing cognitive impairment in
AD mice (7,8). As the effect of different doses of
caffeine on cognitive impairment and the expression of hippocampal
BDNF and TrkB in PS1/APP mice have been poorly investigated, the
present study was conducted. The results demonstrated that low
(0.75 mg/day) and high (1.5 mg/day) doses of caffeine increased
spatial learning ability and the memory and expression of
hippocampal BDNF and TrkB in PS1/APP mice with a dose-response
effect.
It had been shown that the cognitive ability of AD
mice decreased and 1.5 mg/day of caffeine was capable of reversing
the cognitive impairment (7,8). The
present study not only confirmed previous studies but also
investigated the effect of low doses of caffeine (0.75 mg/day) on
the cognitive impairment of AD mice. The results revealed that 0.75
mg/day of caffeine for eight weeks was capable of increasing
spatial learning ability and memory in 12 month old PS1/APP
transgenic mice. It has been reported that the oral administration
of 3 mg/day of caffeine for two weeks was capable of improving
cognitive impairment of 9.5 month old PS1/APP double transgenic
mice (14). A previous study has
demonstrated that 0.5 mg/day of caffeine in drinking water reduced
the cholesterol-induced increase in Aβ and phosphorylated τ, which
suggests that even particularly low doses of caffeine may protect
against sporadic AD-like pathology (15). The varying doses of caffeine
treatments in different studies may be caused by differences in the
ages of mice or treatment time.
Studies have revealed that caffeine intake may
reverse memory impairment and decrease the levels of Aβ in the
brains of AD mice (7,8,14,16);
however, the exact mechanism for the role of caffeine in memory
impairment is unclear. Previous studies have indicated that the
mechanism may be complex and involve a variety of aspects of memory
ability. Long-term caffeine administration may improve memory by
reducing the levels of Aβ through the suppression of the
Aβ-producing enzymes, β- and γ-secretase (7,17).
In another study, it has been demonstrated that caffeine is capable
of decreasing the expression of pro-apoptotic phospho-JNK and
phospho-ERK in the striatum and cortex, and stimulated PKA
signaling in the striatum of APPswe mice. BDNF is crucial in
neuronal plasticity, learning and memory. The levels of BDNF, and
its main receptor TrkB, have been reported to decrease in AD. We
hypothesized that BDNF and its receptor may be involved in the
protective role of caffeine against memory impairment. Results of
the present study have demonstrated that caffeine intake
significantly increased the expression of BDNF, and its main
receptor TrkB, in the brain, which is in agreement with our
hypothesis. There is evidence to support our results. It has been
demonstrated that BDNF, and TrkB, are capable of protecting against
memory impairment and regulate neurogenesis in the hippocampus of
AD (18). A recent study also
supports the role of BDNF signaling through TrkB in the
pathophysiology and cognitive deficits of AD (12). However, the exact mechanism of BDNF
and its receptor involving caffeine in AD remains unclear and
requires further investigation.
In conclusion, the present study reveals that 0.75
mg/day and 1.5 mg/day of caffeine for eight weeks is capable of
reversing memory impairment in 12 month old PS1/APP transgenic
mice, and BDNF and its receptor TrkB may be involved in the
protective role of caffeine against memory impairment in AD.
Acknowledgements
This study was supported by by the Principal Fund
for Clinical Medicine, the Science and Technology Initializing Fund
from the First Affiliated Hospital of Liaoning Medical College
(FYK201205), the Youth Science and Technology Initializing Fund
from the First Affiliated Hospital of Liaoning Medical College
(FY2012-01) and the Youth Science and Technology Initializing Fund
from Liaoning Medical College (Y2012Z001).
References
|
1
|
Massoud F and Gauthier S: Update on the
pharmacological treatment of Alzheimer’s disease. Curr
Neuropharmacol. 1:69–80. 2010.
|
|
2
|
Ritter JM: Drugs for Alzheimer’s disease.
Brit J Clin Pharmacol. 4:501–503. 2012.
|
|
3
|
Fisone G, Borgkvist A and Usiello A:
Caffeine as a psychomotor stimulant: mechanism of action. Cell Mol
Life Sci. 7:857–872. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Ritchie K, Carrière I, de Mendonca A, et
al: The neuroprotective effects of caffeine: a prospective
population study (the Three City Study). Neurology. 69:536–545.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Santos C, Lunet N, Azevedo A, et al:
Caffeine intake is associated with a lower risk of cognitive
decline: a cohort study from Portugal. J Alzheimers Dis.
20:S175–S185. 2010.PubMed/NCBI
|
|
6
|
Maia L and de Mendonça A: Does caffeine
intake protect from Alzheimer’s disease? Eur J Neurol. 9:377–382.
2002.
|
|
7
|
Arendash GW, Mori T, Cao C, et al:
Caffeine reverses cognitive impairment and decreases brain
amyloid-β levels in aged Alzheimer’s disease mice. J Alzheimers
Dis. 17:661–680. 2009.PubMed/NCBI
|
|
8
|
Arendash G, Schleif W, Rezai-Zadeh K, et
al: Caffeine protects Alzheimer’s mice against cognitive impairment
and reduces brain (beta)-amyloid production. Neuroscience.
142:941–952. 2006.
|
|
9
|
Bekinschtein P, Cammarota M, Igaz LM, et
al: Persistence of long-term memory storage requires a late protein
synthesis-and BDNF-dependent phase in the hippocampus. Neuron.
53:261–277. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Peng S, Wuu J, Mufson EJ and Fahnestock M:
Precursor form of brain-derived neurotrophic factor and mature
brain-derived neurotrophic factor are decreased in the pre-clinical
stages of Alzheimer’s disease. J Neurochem. 93:1412–1421.
2005.PubMed/NCBI
|
|
11
|
Ferrer I, Marín C, Rey MJ, et al: BDNF and
full-length and truncated TrkB expression in Alzheimer disease.
Implications in therapeutic strategies. J Neuropathol Exp Neurol.
58:729–739. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Kemppainen S, Rantamäki T, Jerónimo-Santos
A, et al: Impaired TrkB receptor signaling contributes to memory
impairment in APP/PS1 mice. Neurobiol Aging. 33:e23–e39.
2011.PubMed/NCBI
|
|
13
|
Lynch G, Kramar EA, Rex CS, et al:
Brain-derived neurotrophic factor restores synaptic plasticity in a
knock-in mouse model of Huntington’s disease. J Neurosci.
27:4424–4434. 2007.PubMed/NCBI
|
|
14
|
Zeitlin R, Patel S, Burgess S, et al:
Caffeine induces beneficial changes in PKA signaling and JNK and
ERK activities in the striatum and cortex of Alzheimer’s transgenic
mice. Brain Res. 1417:127–136. 2011.PubMed/NCBI
|
|
15
|
Prasanthi JR, Dasari B, Marwarha G, et al:
Caffeine protects against oxidative stress and Alzheimer’s
disease-like pathology in rabbit hippocampus induced by
cholesterol-enriched diet. Free Radic Biol Med. 49:1212–1220.
2010.PubMed/NCBI
|
|
16
|
Alhaider IA, Aleisa AM, Tran TT, et al:
Chronic caffeine treatment prevents sleep deprivation-induced
impairment of cognitive function and synaptic plasticity. Sleep.
33:437–444. 2010.PubMed/NCBI
|
|
17
|
Cao C, Wang L, Lin X, et al: Caffeine
synergizes with another coffee component to increase plasma GCSF:
linkage to cognitive benefits in Alzheimer’s mice. J Alzheimers
Dis. 25:323–335. 2011.PubMed/NCBI
|
|
18
|
Nagahara AH, Merrill DA, Coppola G, et al:
Neuroprotective effects of brain-derived neurotrophic factor in
rodent and primate models of Alzheimer’s disease. Nat Med.
15:331–337. 2009.
|