Introduction
Apolipoprotein B (ApoB), a major component of
low-density lipoproteins (LDLs), is important in lipid metabolism.
It is responsible for the transport of LDLs and mediates their
absorption by cells through LDL receptor-mediated endocytosis. High
plasma LDL levels contribute to the formation of plaques in
arterial walls, which may eventually block arteries and cause
cardiovascular disease (1,2).
In mammals, there are two isoforms of apoB: apoB-100
and apoB-48. The two isoforms are encoded by a single mRNA
transcript. The cytidine residue (C) at nucleotide 6666 of the apoB
mRNA is edited to a uracil (U) by the apolipoprotein B mRNA editing
enzyme APOBEC-1. This C-to-U editing generates a UAA translational
stop codon, resulting in the truncated gene product known as
apoB-48 (3). ApoB-48 lacks
apoB-100’s C-terminal LDL receptor binding region and is unable to
bind to LDL receptors. However, lipoprotein particles containing
apoB-48 possess multiple copies of apolipoprotein E, which assists
with the clearance of particles through LDL receptors and LDL
receptor-related proteins (4,5).
Lipoprotein particles containing apoB-48 clear from plasma more
rapidly than those containing apoB-100 (10 min compared with 3
days) (6). In theory, elevating
the expression of APOBEC-1 in the liver should increase the
efficiency of apoB mRNA editing, leading to a greater secretion of
lipoproteins containing apoB-48, a lower concentration of apoB-100
LDLs in the plasma and reduced formation of atherosclerotic
plaques.
Curcumin, an extract from the traditional Chinese
medicinal herb Curcuma longa L., possesses numerous
important biological activities. It has potential as a
chemotherapeutic agent and has hypocholesterolemic properties,
lowering the level of LDL-cholesterol in plasma (7–9). In
the present study, we used primary rat hepatocytes as a model
system to investigate whether the expression of APOBEC-1 and the
level of apoB mRNA editing may be improved by curcumin treatment.
Through this study, we aimed to identify a potential new therapy to
increase the amount of apoB-48 and reduce the amount of apoB-100 in
order to lower the incidence of atherosclerosis. Furthermore,
changes in APOBEC-1 expression levels and apoB mRNA editing
efficiency were analyzed to gain insight into the molecular
mechanisms of lipid metabolism underlying the effects of
curcumin.
Materials and methods
Animals
Animal experimental procedures were performed in
conformity with the Guide for the Care and Use of Laboratory
Animals (NIH publication no. 85–23, revised 1996) and were approved
by the Ethics Committee of the Animal Laboratory Research Centre of
Zhejiang Chinese Medical University [Zhejiang, China; approval no.
SYXK (Zhe) 2008-0115]. Sprague-Dawley (SD) rats within 24 h of
birth were purchased from Shanghai B&K Universal Group Limited
(Shanghai, China). Standard laboratory food and water were
available ad libitum.
Isolation of primary rat hepatocytes
Primary hepatocytes were obtained from male SD rats
within 24 h of birth. We used an improved collagenase/trypsin
digestion method, as follows: Rats were decapitated and the livers
were extracted and placed in cold D-Hanks solution. Liver capsules
were peeled off with forceps and the livers were cut into small
pieces with ophthalmic scissors. Tissues were thoroughly washed and
then digested with 0.25 mg/l trypsin for 3 min at 37°C. Dulbecco’s
modified Eagle’s medium (DMEM) plus 10% fetal bovine serum (FBS)
was added to terminate the digestion process. The digested tissues
were centrifuged at 800 g at 15–25°C for 5 min and the supernatant
was removed. Tissues were transferred to a sterile glass bottle and
digested for 15 min using 0.2 mg/l type IV collagenase. The
reaction was terminated by adding DMEM plus 10% FBS until the
digested material became flocculent. The dispersed cells were
filtered through a 200-mesh sieve and centrifuged for 5 min at 800
g. The cell pellet was washed twice with fresh medium and the
differential adhesion method was used to remove the fibroblasts.
Cell viability was >95%, which was measured using the trypan
blue exclusion test.
Culturing and identification of primary
rat hepatocytes
Primary rat hepatocytes (viability >95%) were
seeded in culture flasks at a density of 4×105 to
5×105 cells/ml and incubated in DMEM/F12 plus 5% FBS.
The day prior to identification, the cells (5×104
cells/ml) were plated onto glass cover slips in 24-well dishes and
grown for 24 h. Cells were washed twice with phosphate buffer
saline (PBS; pH 7.4) and then stained using a periodic acid-Schiff
kit according to the manufacturer’s instructions (Nanjing Jiancheng
Bioengineering Institute, Nanjing, Jiangsu, China). Stained cells
were mounted on glass slides and images were captured using a Nikon
Eclipse 80i microscope connected to a DS-5M-L1 camera.
Cell viability assay
Cell viability of primary rat hepatocytes was
measured using the MTT viability test. Briefly, cells were seeded
at a density of 6×103 cells/well in 96-well plates with
or without increasing concentrations of curcumin for 24 h. MTT (50
μl, 2 mg/ml) was added to each well and the plate was incubated for
an additional 4 h. Following removal of the medium, formazan
crystals were dissolved in 150 μl of dimethyl sulfoxide (DMSO). The
absorbance of MTT-formazan at 550 nm was measured using a
SpectraMaxM3 microplate reader.
Real-time PCR analysis
Following treatment of primary rat hepatocytes with
no curcumin or with 5, 15 or 25 μM curcumin for 24 h, total RNA was
isolated using TRIzol reagent (Invitrogen Life Technologies,
Carlsbad, CA, USA) according to the manufacturer’s instructions.
Complementary DNA (cDNA) was synthesized using a HiFi-MMLV cDNA kit
(Beijing Kang Century Biotechnology Co., Ltd., Beijing, China).
qRT-PCR was conducted using UltraSYBR mixture (Beijing Kang Century
Biotechnology Co., Ltd.). All primers were designed using primer5
(Premier Biosoft, Palo Alto, CA, USA). Primers for the β-actin gene
were 5′-GGCACCACACCTTCTACAAT-3′ (forward) and
5′-GTGGTGGTGAAGCTGTAGCC-3′ (reverse). Primers for the APOBEC-1 gene
were 5′-GAGCCCCACGAGTTTGAAGT-3′ (forward) and
5′-ACACCGCTGCTAATAAGGTC-3′ (reverse). All samples were run in
triplicate and changes in gene expression were calculated using the
ΔΔCt method. For quantitative analysis of apoB mRNA editing, a
regular polymerase chain reaction (PCR) was performed using Taq DNA
polymerase (Beijing Kang Century Biotechnology Co., Ltd.) and
primers 5′-GGCTTCCTCAGCAGATTCAT-3′ (forward) and
5′-ATCCAAGACGCACCACTACT-3′ (reverse).
Western blot analysis
Proteins from primary rat hepatocytes from the
control group (cultured for 24 h without curcumin) and the
treatment groups (cultured for 24 h with 5, 15 or 25 μM curcumin)
were extracted using Protein Extraction Reagent (Boster
Bioengineering, Wuhan, China) containing 1 mM phenylmethanesulfonyl
fluoride (PMSF; Roche Molecular Biochemicals, Indianapolis, IN,
USA). Protein concentrations were determined using the
bicinchoninic acid (BCA) protein assay (Nanjing KeyGen Biotech Co.
Ltd., Nanjing, Jiangsu, China). The proteins were separated by 10%
SDS-PAGE and then transferred onto a polyvinylidene difluoride
(PVDF) membrane (Pall Gelman Laboratory Corporation, Ann Arbor, MI,
USA). For western blot analyses, membranes were blocked for 2 h in
PBS containing 5% nonfat dried milk powder at room temperature,
incubated overnight with the primary antibody at 4°C, washed, and
then incubated with the secondary antibody for 2 h at room
temperature. Chemiluminescence was detected using an EZ-ECL
Chemiluminescence Detection kit for horseradish peroxidase (HRP;
Biological Industries, Beit HaEmek, Israel). Mouse antibody to
APOBEC-1 (Santa Cruz Biotechnology, Inc., Santa Cruz, CA, USA) and
β-actin (Boster Bioengineering) and goat anti-mouse immunoglobulin
G (IgG; Hangzhou HuaAn Biotechnology Co., Ltd., Hangzhou, Zhejiang,
China) were used in this study.
Quantification of RNA editing level
The editing level of apoB mRNA C6666
sites was determined from mixed C/T peaks at site 6666 in RT-PCR
production sequence chromatograms, where the genomically encoded
base was cytidine. The ratio between the C and T (U) peak heights
in individual chromatograms was measured using FinchTV. Percent
editing was calculated as the T peak height relative to the total
peak height (T/(C+T)x100). Since this method is not quantitative,
we also assessed editing levels by counting clones. We cloned the
RT-PCR products containing the edited cytidine into the pUC-T
vector (Beijing Kang Century Biotechnology Co., Ltd.). For each
sample, 90 clones were sequenced by Shanghai Sangon Biological
Engineering Technology & Services Co., Ltd. (Shanghai,
China).
Statistical analysis
Statistical analysis was performed using one-way
analysis of variance (ANOVA) for multiple comparisons and t-tests
among groups. P<0.05 was considered to indicate a statistically
significant difference and P<0.01 was considered to indicate an
extremely statistically significant difference.
Results
Curcumin has no significant effect on the
viability of primary rat hepatocytes
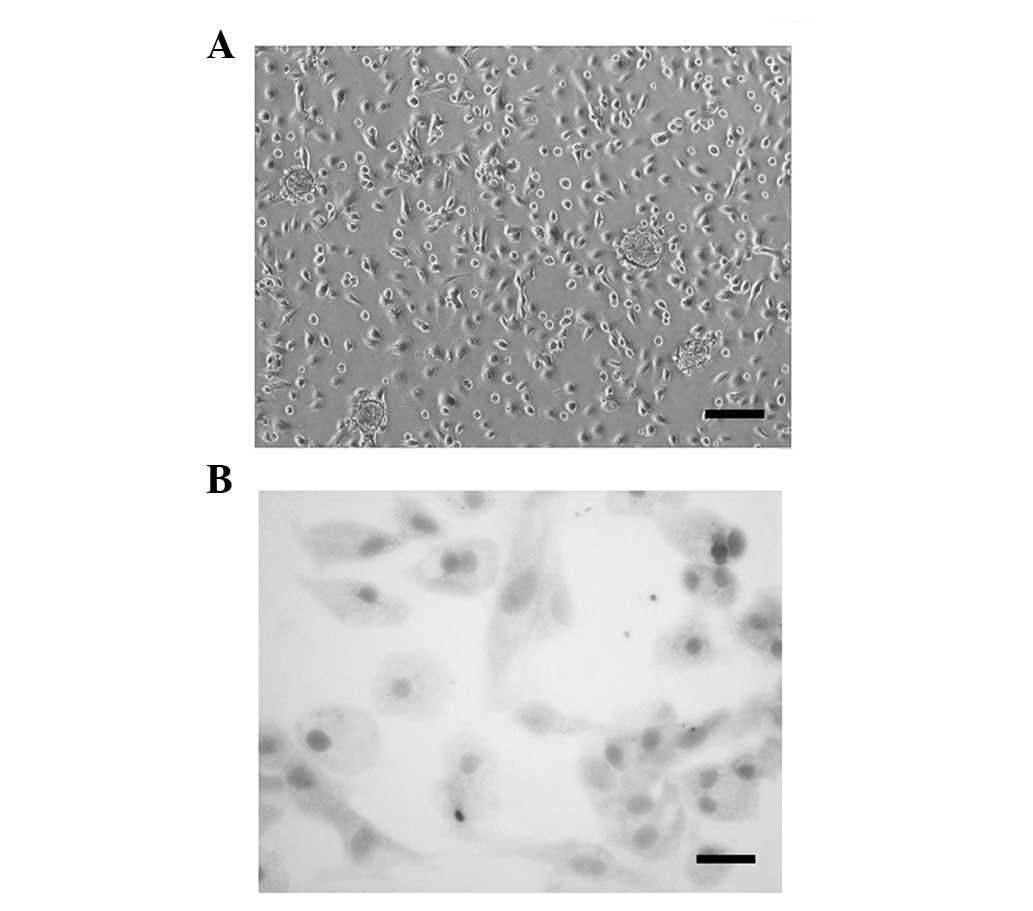
The cells isolated from newborn SD rats had a
rounded orpolygonal cell body with a translucent cytoplasm and
nearly covered the bottom of the flask following 5 days of culture
in DMEM/F12 (Fig. 1A). Following
periodic acid-Schiff staining, dense pink glycogen granules were
observed in the cytoplasm of the cells. Vacuoles were present
around nuclei, which were stained blue with hematoxylin (Fig. 1B). These observations indicated
that plenty of pure rat hepatocytes were harvested using the
improved collagenase/trypsin digestion method. These pure rat
hepatocytes were used in all the following experiments within three
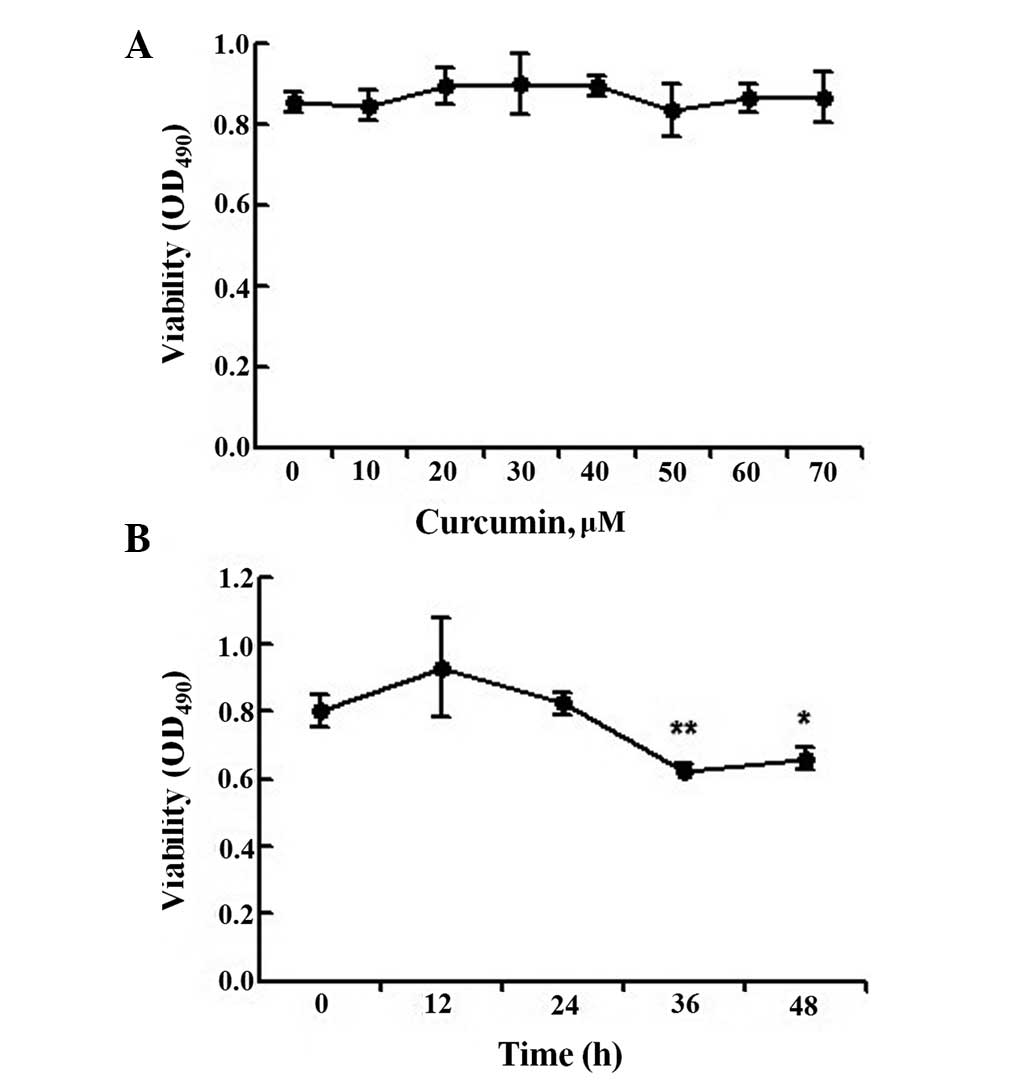
generations. The viability of hepatocytes treated with 10 to 70 μM
curcumin was not significantly different from that of the control
group (Fig. 2A), indicating that
curcumin concentrations up to 70 μM have no significant cytotoxic
effects on these cells. Therefore, test concentrations of 5, 15 and
25 μM were selected. We also studied the effect of 25 μM curcumin
at different time points. MTT assays revealed that while treatment
for 0–24 h had no effect on hepatocyte viability, cell viability
was reduced following 36 h of exposure (Fig. 2B). Therefore, primary rat
hepatocytes were treated with 5, 15 or 25 μM curcumin for 24 h.
Curcumin promotes the expression of
APOBEC-1 in a non dose-dependent manner
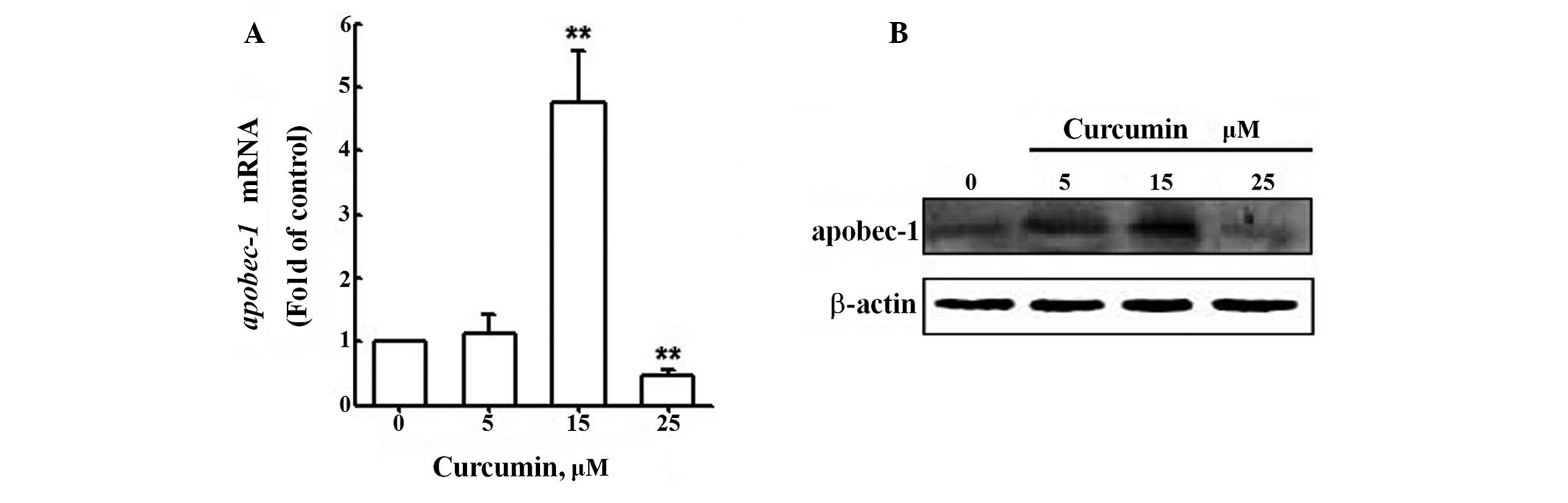
To obtain insights into the effect of curcumin on
apoB mRNA editing efficiency, we first evaluated the change in
APOBEC-1 mRNA expression following curcumin treatment by qRT-PCR
analysis of RNA samples from primary rat hepatocytes from the
control group and the treatment groups (Fig. 3A). APOBEC-1 mRNA levels were up to
more than three-fold higher in cells treated with 15 μM curcumin
than in untreated cells; however, mRNA levels were markedly reduced
in cells treated with 25 μM curcumin. Similar results were obtained
when APOBEC-1 protein levels were analyzed by western blotting
(Fig. 3B). These results indicate
that curcumin affects the expression of APOBEC-1 in primary rat
hepatocytes; however, not in a dose-dependent manner.
Curcumin (15 μM) increases the efficiency
of apoB mRNA editing
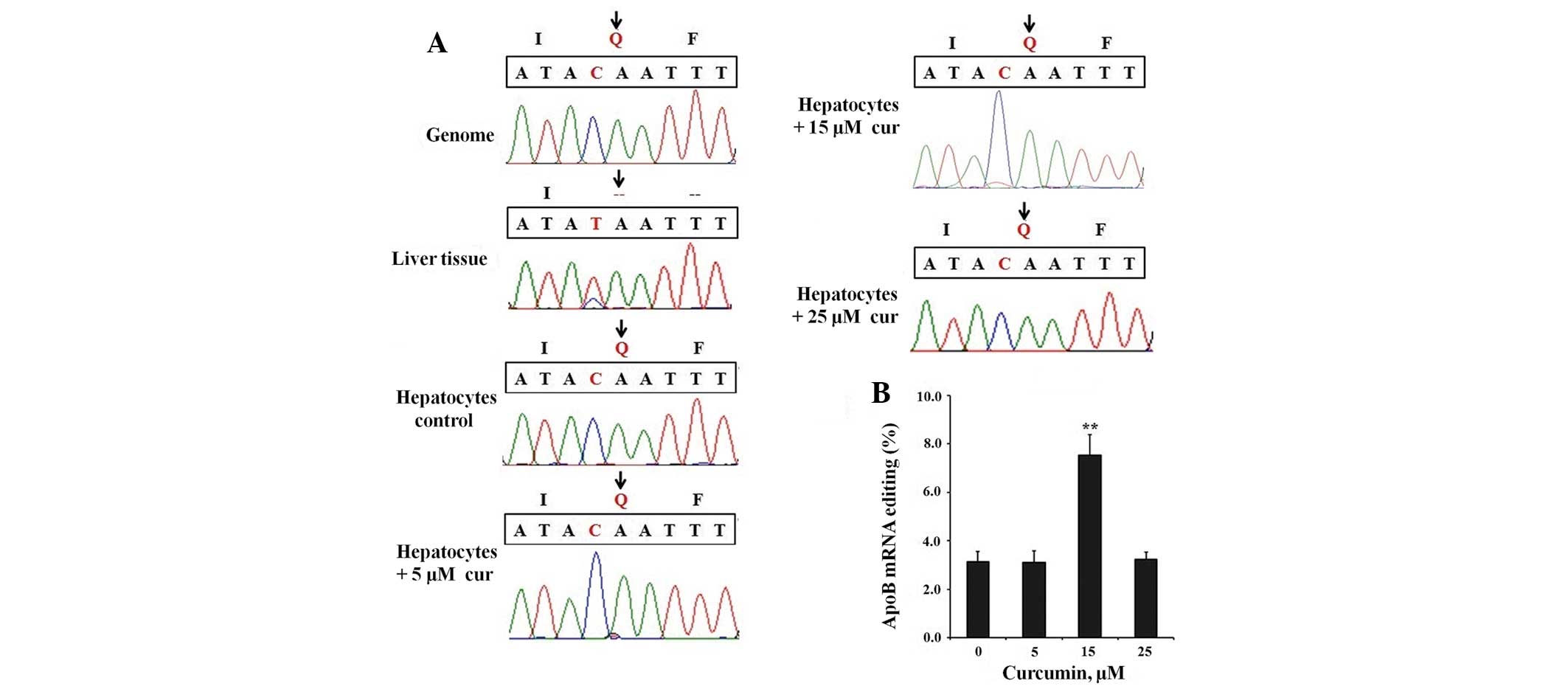
To evaluate the functional role of curcumin, apoB
mRNA editing levels in all groups were first investigated by
sequencing apoB mRNA RT-PCR products. As shown in Fig. 4A, the C-to-U editing signal was
only able to be detected in the 15 μM curcumin treatment group. We
also demonstrated that the apoB mRNA editing efficiency was
significantly higher in newborn rat liver tissues than in the
primary rat hepatocytes. To assess editing levels more
quantitatively, we sequenced at least 3×30 clones from each group
(Fig. 4B). The apoB mRNA editing
level increased from 3.13% in the control group (Fig. 4A, hepatocytes control) to 7.53% in
the 15 μM curcumin treatment group; however the other curcumin
concentrations had no significant effect. Furthermore, we
discovered no other editing site in the apoB mRNA besides the
normal C6666 site, in all groups. From these analyses,
we concluded that 15 μM curcumin increased the amount of apoB-48
and reduced the amount of apoB-100 in primary rat hepatocytes.
Discussion
Previous studies have demonstrated that curcumin is
able to reduce the LDL concentration in plasma in several different
ways, for example by upregulating the LDL receptor via the
SCAP-SREBP pathway to eliminate LDL-cholesterol in the blood
(10); by causing significant
activation of liver X receptor (LXR) and scavenger receptor class A
(SR-A) to decrease LDL-cholesterol synthesis (11) and by elevating the activity of
hepatic cholesterol-7α-hydroxylase, which is a key enzyme in
cholesterol degradation (12).
However, in the present study, we focused on the editing of the
mRNA transcript encoding apoB, the primary apolipoprotein of ‘bad
cholesterol’, and verified the LDL-cholesterol-lowering effect of
curcumin in a new way.
We used primary rat hepatocytes, isolated from
newborn SD rats by an improved collagenase/trypsin digestion
method, as a model system. Since primary cultured cells maintain
the basic properties of the original cells, they are extremely good
biological models for drug experiments (13). To investigate the effect of
curcumin, we performed qRT-PCR and western blot analyses to detect
the expression levels of APOBEC-1 in primary hepatocytes treated
with different concentrations of curcumin. In addition, an apoB
mRNA fragment covering C6666 was PCR amplified from cDNA
and cloned, and then the PCR products and the clones were sequenced
to determine differences in the mRNA editing efficiency between
curcumin-treated and untreated cells. The results clearly showed
that 15 μM curcumin increased the expression of APOBEC-1 at the
mRNA and protein levels; however, 25 μM curcumin reduced the
expression of this cytidine deaminase. Therefore, curcumin
increased the expression of APOBEC-1 in a non dose-dependent
manner. Furthermore, when the expression of APOBEC-1 increased, the
apoB mRNA editing level also increased (significant result,
P<0.01). In conclusion, our study demonstrated that an
intermediate concentration of curcumin increased the level of apoB
mRNA editing by increasing the expression of APOBEC-1, which
increased apoB-48 and reduced apoB-100.
Other studies have demonstrated that adenoviral
expression of exogenous APOBEC-1 is able to effectively increase
apoB mRNA editing efficiency and reduce plasma LDL levels; however,
the brief duration of transgene expression and the immune response
to adenoviral vectors make this method unsuitable for studies on
atherosclerosis (14). Curcumin is
a natural compound and has been demonstrated to be non toxic to
humans at dosages of up to 8000 mg/day when administered orally for
3 months (15). Therefore,
compared with the adenoviral delivery method, curcumin has the
potential to increase apoB mRNA editing efficiency for a longer
period without toxic side effects.
Transgenic animals provide another approach that is
capable of yielding long-term increases in APOBEC-1 expression in
the liver; however, hypermutation that is believed to induce liver
dysplasia and hepatocellular carcinomas through hyperediting of
apoB and other mRNAs has been discovered in such transgenic animals
(16). Therefore, there remains
numerous obstacles to the use of gene therapy for the clinical
treatment of atherosclerosis. In the present study, no editing site
other than the normal C6666 site was detected in primary
rat hepatocytes treated with 15 μM curcumin. Patterson et al
found that APOBEC-1 increased apoB mRNA hyperediting in a
dose-dependent manner, such that when the apoB mRNA editing level
was higher than 10%, additional editing sites were discovered. In
the present study, the C6666 site editing level in the
15 μM treatment group increased to only 7.53%. As the editing level
did not exceed 10%, hyperediting may have been avoided even though
curcumin increased the expression level of APOBEC-1. In conclusion,
compared with adenoviral delivery and transgenic animals, curcumin
treatment has clear advantages for therapeutic use to reduce
atherogenic lipoprotein levels and prevent atherosclerosis.
C-to-U editing of apoB mRNA is mediated not only by
APOBEC-1, but also by other proteins that form an enzyme complex
with APOBEC-1. Curcumin may also affect the expression of other
components in this system, possibly explaining why 25 μM curcumin
did not affect the apoB mRNA editing level despite reducing the
expression of APOBEC-1. Future studies of this mRNA editing system
are required in order to further understand how curcumin increases
the level of apoB mRNA editing.
To the best of our knowledge, this study is the
first to state that curcumin increases the level of apoB-48 and
reduces the level of apoB-100 through the C-to-U RNA editing enzyme
APOBEC-1 in primary rat hepatocytes without hyperediting and
toxicity. Compared with adenoviral delivery and transgenic animals,
curcumin treatment has clear advantages, therefore it may be used
therapeutically to reduce levels of atherogenic lipoproteins and
prevent atherosclerosis.
Acknowledgements
This study was supported by research grants from the
Natural Science Foundation of Zhejiang Province, Youth Fund Project
(no. LQ12C07001).
References
|
1
|
Young SG: Recent progress in understanding
apolipoprotein B. Circulation. 82:1574–1594. 1990. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Innerarity TL, Borén J, Yamanaka S and
Olofsson SO: Biosynthesis of apolipoprotein B48-containing
lipoproteins. Regulation by novel post-transcriptional mechanisms.
J Biol Chem. 271:2353–2356. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Anant S, Blanc V and Davidson NO:
Molecular regulation, evolutionary, and functional adaptations
associated with C to U editing of mammalian apolipoproteinB mRNA.
Prog Nucleic Acid Res Mol Biol. 75:1–41. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Anant S and Davidson NO: Molecular
mechanisms of apolipoprotein B mRNA editing. Curr Opin Lipidol.
12:159–165. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Pitas RE, Innerarity TL and Mahley RW:
Cell surface receptor binding of phospholipid protein complexes
containing different ratios of receptor active and inactive E
apoprotein. J Biol Chem. 255:5454–5460. 1980.
|
|
6
|
Innerarity TL and Mahley RW: Enhanced
binding by cultured human fibroblasts of apo-E-containing
lipoproteins as compared with low density lipoproteins.
Biochemistry. 17:1440–1447. 1978. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Gupta SC, Patchva S, Koh W and Aggarwal
BB: Discovery of curcumin, a component of the golden spice, and its
miraculous biological activities. Clin Exp Pharmacol Physiol.
39:283–299. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Joe B, Vijaykumar M and Lokesh BR:
Biological properties of curcumin-cellular and molecular mechanisms
of action. Crit Rev Food Sci Nutr. 44:97–111. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Egan ME, Pearson M, Weiner SA, et al:
Curcumin, a major constituent of turmeric, corrects cystic fibrosis
defects. Science. 304:600–602. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Dou X, Fan C, Wo L, Yan J, Qian Y and Wo
X: Curcumin up-regulates LDL receptor expression via the sterol
regulatory element pathway in HepG2 cells. Planta Med.
74:1374–1379. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Zhao JF, Ching LC, Huang YC, et al:
Molecular mechanism of curcumin on the suppression of cholesterol
accumulation in macrophage foam cells and atherosclerosis. Mol Nutr
Food Res. 56:691–701. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Srinivasan K and Sambaiah K: The effect of
spices on cholesterol 7 alpha-hydroxylase activity and on serum and
hepatic cholesterol levels in the rat. Int J Vitam Nutr Res.
61:364–369. 1991.PubMed/NCBI
|
|
13
|
Wang K, Shindoh H, Inoue T and Horii I:
Advantages of in vitro cytotoxicity testing by using primary rat
hepatocytes in comparison with established cell lines. J Toxicol
Sci. 27:229–237. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Teng B, Blumenthal S, Forte T, et al:
Adenovirus-mediated gene transfer of rat apolipoprotein B
mRNA-editing protein in mice virtually eliminates apolipoprotein
B-100 and normal low density lipoprotein production. J Biol Chem.
269:29395–29404. 1994.PubMed/NCBI
|
|
15
|
Cheng AL, Hsu CH, Lin JK, et al: Phase I
clinical trial of curcumin, a chemopreventive agent, in patients
with high-risk or pre-malignant lesions. Anticancer Res.
21:2895–2900. 2001.PubMed/NCBI
|
|
16
|
Hersberger M, Patarroyo-White S, Qian X,
et al: Regulatable liver expression of the rabbit apolipoprotein B
mRNA-editing enzyme catalytic polypeptide 1 (APOBEC-1) in mice
lacking endogenous APOBEC-1 leads to aberrant hyperediting. Biochem
J. 369:255–262. 2003. View Article : Google Scholar
|