Introduction
Muscle atrophy, dysfunction and resistance to
non-depolarizing neuromuscular blocking agents are commonly
associated with glucocorticoid (GC) treatment. Previous studies
have reported that partial recovery or termination of
pancuronium-induced paralysis occurs following large doses of
steroid administration (1–3). Soltész et al found a shorter
duration of atracurium and rocuronium-induced neuromuscular
blockage in patients receiving long-term prednisolone medication
(4,5). In addition, a retrospective clinical
review showed that patients receiving long-term pretreatment with
betamethasone required 75% more vecuronium (6). Preclinically, Arts et al
reported that the neuromuscular blocking effect of d-tubocurarine
at two doses and low-dose hemicholinium-3 may be partially or
wholly antagonized by prednisolone (7). Additionally, Robinson et al
and Parr et al reported betamethasone-induced resistance to
atracurium and vecuronium (8,9).
Findings of previous in vitro studies have
indicated that GCs have presynaptic facilitatory effects on
neuromuscular transmission at low concentrations (10–15).
The presynaptic effects are associated with enhanced active uptake
of choline into the nerve terminal, resulting in increased
synthesis and the release of acetylcholine (ACh). This effect may
contribute to resistance to neuromuscular blocking drugs following
GC treatment (15). However, it is
unclear whether these in vitro observations apply in
vivo, particularly when GCs are administered chronically.
Skeletal muscle atrophy induced by GCs is
characterized by a decrease in the size of fast-twitch or type II
muscle fibers with little or no impact observed on slow-twitch or
type I fibers. Skeletal muscle atrophy is also accompanied by a
reduction in the relative contribution of type II fibers to total
muscle fibers (16–21). Respiratory and peripheral muscles
are affected by corticosteroid treatment and GC-induced atrophy of
the diaphragm is often observed in clinical practice (21,22).
The diaphragm is composed of 40% type I and 60% type II fibers
(23). A shift towards slow-twitch
fibers induced by steroids may cause a marked impairment in
diaphragm function. Notably, GC-induced changes in isotonic
contractile properties due to an altered diaphragm-type composition
have been demonstrated in a number of animal studies (24–27).
Mechanisms of resistance to non-depolarizing muscle
relaxants (NDMRs) in GC treatment have not been investigated
extensively. Aside from featuring various contractile properties,
the type II fiber is more sensitive to NDMRs compared with the type
I fiber. As the planar area of the motor endplate in a type II
fiber is much larger compared with type I, the sarcoplasmic
reticulum density is twice that of the type I fiber. Drugs enter
the former junction more rapidly than the latter (28–30).
The composition of muscle fiber-types establishing various
sensitivities to particular muscle relaxants has been observed in a
number of clinical studies (31–36).
Donati et al examined vecuronium neuromuscular blockage of
two skeletal muscles in adult females and observed that the
adductor larynx, which contained mainly type II fibers, showed a
faster onset of muscle relaxant compared with the adductor
pollicis, which contained primarily type I fibers (36). Day et al compared the
ED50 of pancuronium in two muscle types, the
gastrocnemius (fast twitch) and soleus (slow twitch) and found that
the gastrocnemius was more sensitive (ED50, 105 μg/kg)
compared with the soleus (ED50, 150 μg/kg) (37). Thus, GC-induced fiber-type
transformation may be one of the mechanisms associated with
resistance to the action of NDMRs.
Therefore, the aim of the current study was to
investigate the effect of chronic administration of the popular
steroid, Dex, on diaphragm function and on the potency of a common
NDMR, rocuronium as well as to investigate whether any observed
effects are associated with muscle-fiber transformation.
Materials and methods
Animals and muscle preparation
This study was approved by the Animal Care and Use
Committee of the Shanghai Jiaotong University School of Medicine
and was performed in compliance with the WHO International Guiding
Principles for Animal Research. Sixty adult male Sprague-Dawley
rats (180–220 g) were randomized into three groups (n=20 per
group). The Dex group received a daily intraperitoneal injection of
Dex [600 μg/kg body mass; a dose that has been previously shown to
cause significant loss of muscle mass (17,38)]
suspended in an isotonic vehicle (0.9% NaCl) for 14 days. The
saline group received an equivalent volume of saline (0.9% NaCl)
daily for 14 days. All injections were performed at approximately
the same time of day in the two groups. A food-restricted group
(fr) was pair-fed with the Dex group for 14 days to evaluate
whether muscle dysfunction following Dex treatment was caused by
anorexia typically associated with GC therapy. This was achieved by
weighing the food intake of the Dex group rats daily and providing
the fr group rats with an equivalent amount of food. Food and water
was available ad libium to rats in the Dex and saline
groups. Each experimental group contained 20 rats. The animals were
included in the final data set.
Twenty-four hours following the final drug
administration, the rats were anesthetized with pentobarbital (60
mg/kg intraperitoneally). The left hemidiaphragm, with attached
phrenic nerve and the right hemidiaphragm, both with the central
tendon and ribcage intact, were rapidly removed from each rat to
investigate rocuronium potency and diaphragm contractibility.
Strips of diaphragm (8–10 mm wide, cut parallel to
the muscle fibers) with or without the phrenic nerve attached, were
dissected for subsequent experiments. Strips obtained from the
costal region of the diaphragm were embedded separately in plastic
holders and rapidly frozen in isopentane cooled in liquid nitrogen
(−160°C), followed by further freezing in liquid nitrogen for
ATPase staining. Each isolated strip was mounted vertically in a
tissue chamber, fixed at the rib cage, inferiorly positioned and
suspended at the central tendon from a force displacement
transducer using a 3-0 silk suture (ALC-M System for Isolated
Tissue-Organ Research; 40 ml in volume). The chamber was filled
with Krebs solution, maintained at 37°C and bubbled with 95%
oxygen/5% CO2. The composition of the Krebs solution was
as follows: 137 mM NaCl, 4 mM KCl, 2 mM CaCl2, 1 mM
MgCl2, 1 mM KH2PO3, 12 mM
NaHCO3 and 6.5 mM glucose, with a pH 7.40±0.05 during
bubbling. Isometric tension was elicited by indirect (phrenic
nerve) or direct (muscle) supramaximal constant-voltage stimulation
at 0.1 Hz for 0.05 and 0.2 msec in the cases of indirect and direct
stimulation, respectively, using a stimulator and a
constant-voltage unit. The twitch tension was recorded via the
force transducer on a recorder (ALC-MPA 2000m, Acquisition and
Analysis System for Life Science Research, Shanghai Alcott Biotech,
Shanghai, China). The stimulator was activated by a personal
computer. Twitch stimuli were used to determine the optimal length
(L0) at which skeletal muscle may generate the greatest
force, followed by a 15-min thermoequilibration period. A number of
measurements were performed as described in subsequent
sections.
Diaphragm contractile properties
General
For direct stimulation, the diaphragm was placed
between two large silver electrodes and pretreated with a
sufficient concentration (12 μM) of d-tubocurarine to completely
eliminate neuromuscular transmission.
Twitch characteristics
Five twitches were recorded at L0 to
obtain maximal twitch force (Pt), time-to-peak tension
(TPT) and half-relaxation time (1/2RT). The average values were
used for subsequent analysis.
Maximal tetanic contraction
Two maximal tetanic stimuli, with a frequency of 120
Hz and a train duration of 250 msec were generated to obtain the
maximal tetanic force (P0).
Force-frequency instructions
Muscle bundles were stimulated every 2 min at the
following frequencies: 10, 20, 40, 60, 80 and 120 Hz (train
duration, 250 msec).
Fatigue properties
Bundles were fatigued by means of 330 msec
stimulations repeated at 25 Hz and applied every sec for 5 min as
described by Burke et al with modifications. Fatigue index
(fatigue), adapted from Burke, is expressed as the ratio of the
tetanus force developed at 120 sec/the maximal force developed by
tetanus (39). Following the
experiment, the muscle was weighed and the cross-sectional area
(CSA) was estimated based on the formula: muscle weight
(g)/[L0 (cm) × 1.056 (g/cm3)], assuming
muscle density is 1.056 g/cm3 (26). Forces were normalized against the
CSA of the muscle segments.
Rocuronium potency
For indirect stimulation, the phrenic nerve was
positioned on wire bipolar silver electrodes. After the elicited
twitch tension had been stabilized for a minimum of 15 min, the
single-twitch tension, averaged in groups of five, was determined.
Rocuronium was then applied to the preparation. Drug concentrations
were determined by adding freshly prepared solutions with
calibrated micropipettes to modified Krebs solution (40 ml) in the
tissue chamber. Following stabilization of the drug effect for a
minimum of 10 min, single-twitch tension was again determined. Data
were accepted when twitch tension returned to 95–105% of the
initial value by rinsing the diaphragm preparation with Krebs
solution in each study. Rocuronium bromide was obtained from N.V.
Organon (Oss, The Netherlands) and all other drugs were purchased
from Sigma-Aldrich (St. Louis, MO, USA). Indirectly elicited twitch
tension in the same preparation with no neuromuscular blocker was
defined as the control value.
ATPase staining
Muscle blocks were serially sliced at 5 μm on a
cryostat (Leica CM1850, Nussloch, Germany). The modified ATPase
staining method was applied in addition to hematoxylin and eosin
(H&E) staining.
Muscle fiber types were classified as type I or II
based on their staining for myofibrillar ATPase following alkaline
pH 10.4 and acid pH 4.5 using a modification of the procedure
described by Brooke et al (40) and the criteria outlined by Peter
et al (41). The image
analysis software Image-Pro Plus, version 5.1 (Media Cybernetics,
Silver Spring, MD, USA) was used to evaluate the prepared samples.
Then, ~400 fibers/muscle were measured for muscle fiber-type
distribution and CSA of each fiber type. The distribution of muscle
fibers of a given fiber type (type I or II) was expressed as a
percentage of all the fibers in the section.
Statistical analysis
Statistical analyses were performed using the
SPSS/PC+ package (SPSS INC., Chicago, IL, USA). Data are expressed
as mean ± SD and were compared using one- or two-way
repeated-measures ANOVA with post hoc Bonferroni testing. Otherwise
repeated-measures ANOVA was used for force-frequency and the growth
curve analysis. P<0.05 was considered to indicate a
statistically significant difference.
Results
Body weight
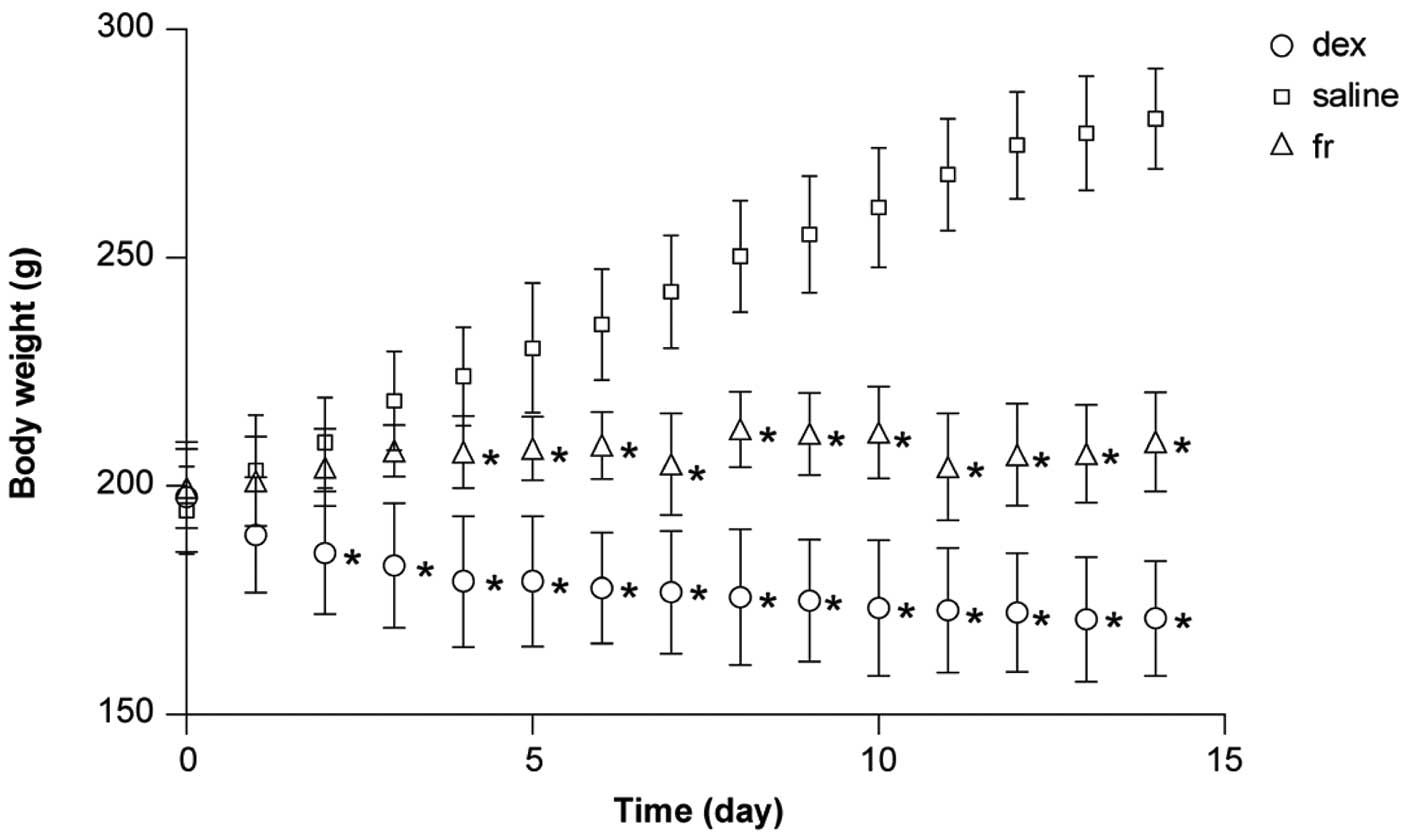
There was no difference in the initial body weight
between groups (P>0.05). Food intake decreased markedly in all
animals during the study, particularly in the initial three days
following Dex treatment (data not shown). At the end of the 14-day
treatment period, the body weights in the three experimental groups
were significantly different from one another (P<0.01; Table I). Repeated-measures ANOVA revealed
a significant effect of treatment on body weight during the study
(Fig. 1).
 | Table IEffect of experimental treatment on
body weight. |
Table I
Effect of experimental treatment on
body weight.
| Body weight |
|---|
|
|
|---|
| Group | Initial, g | Final, g |
|---|
| Dex | 197.55±2.688 |
171.10±2.812a,b |
| Saline | 194.70±2.163 | 280.95±2.423 |
| Fr | 199.55±1.941 | 209.70±2.43a |
Diaphragmatic contractile properties
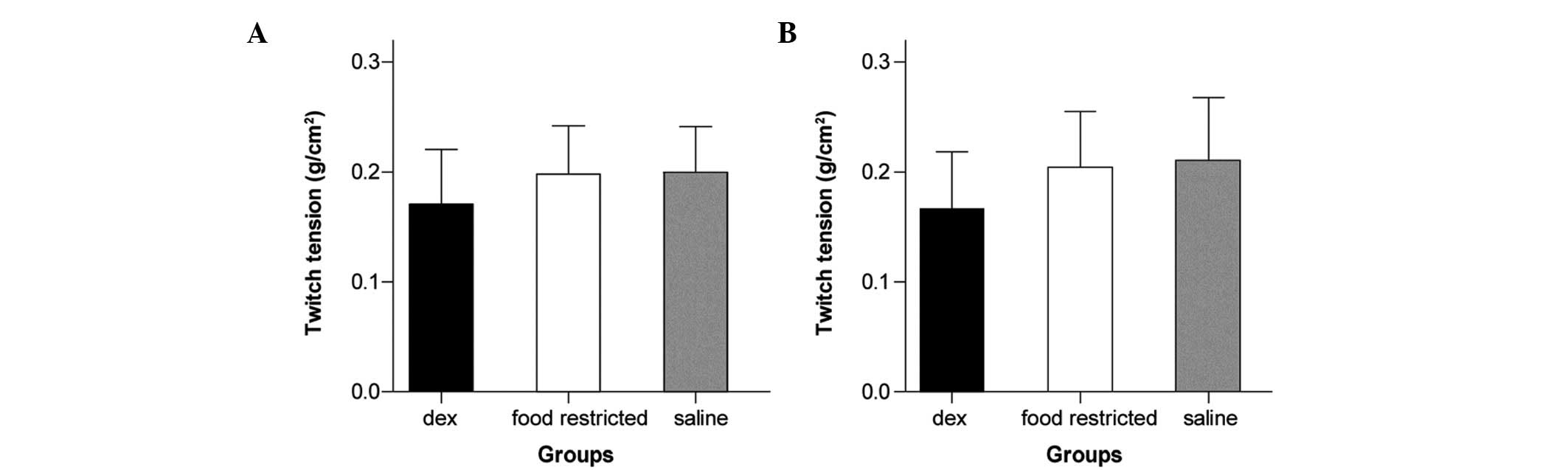
There was no significant difference between groups
with regard to diaphragm size (width and length) or in twitch
tensions elicited by indirect or direct stimulation (7.8–17.4 g;
P>0.05; Fig. 2A and B).
Twitch characteristics and maximal
tetanic tension
Data from the contractile properties of the
diaphragm are summarized in Table
II. P0 corrected by CSA (kg/cm2)
decreased in the Dex group (P<0.05). Pt and
Pt/P0 were decreased in the Dex group,
however, these differences did not reach statistical significance.
TPT and 1/2RT were significantly prolonged in the Dex group
compared with the other two groups (P<0.05).
| Table IIDiaphragmatic contractile
properties. |
Table II
Diaphragmatic contractile
properties.
| Group | Pt
(kg/cm2) | TPT (msec) | 1/2RT (msec) | P0
(kg/cm2) |
Pt/P0 | Fatigue (%) |
|---|
| Dex | 0.17 (0.05) | 23.3 (2.26)a,b | 24.3(2.12)a,b | 0.63 (0.12)a,b | 0.27 (0.04) | 49.6 (8.72)a,b |
| Saline | 0.21 (0.06) | 19.6 (1.98) | 20.2 (2.54) | 0.92 (0.31) | 0.23 (0.06) | 34.6 (9.64) |
| Fr | 0.21 (0.05) | 20.9 (2.34) | 21.4 (2.95) | 1.00 (0.37) | 0.21 (0.05) | 35.8 (7.93) |
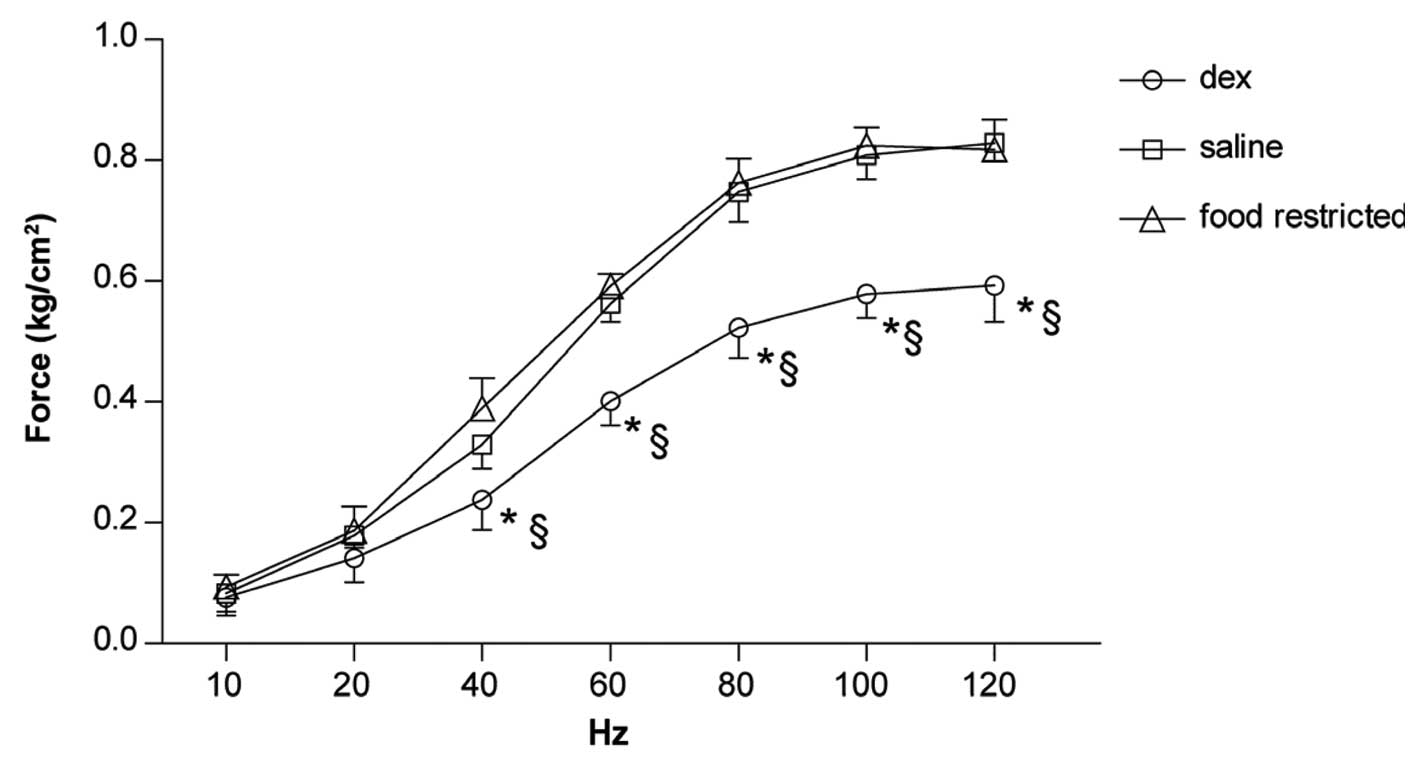
Force-frequency curve
The response of diaphragm strips to increasing
stimulus frequencies is shown in Fig.
3. A significant downward shift between 40 and 120 Hz was
observed in the Dex group (Fig.
3).
Fatigue properties
The fatigue index was significantly higher
(P<0.05) in the Dex group compared with the other two
groups.
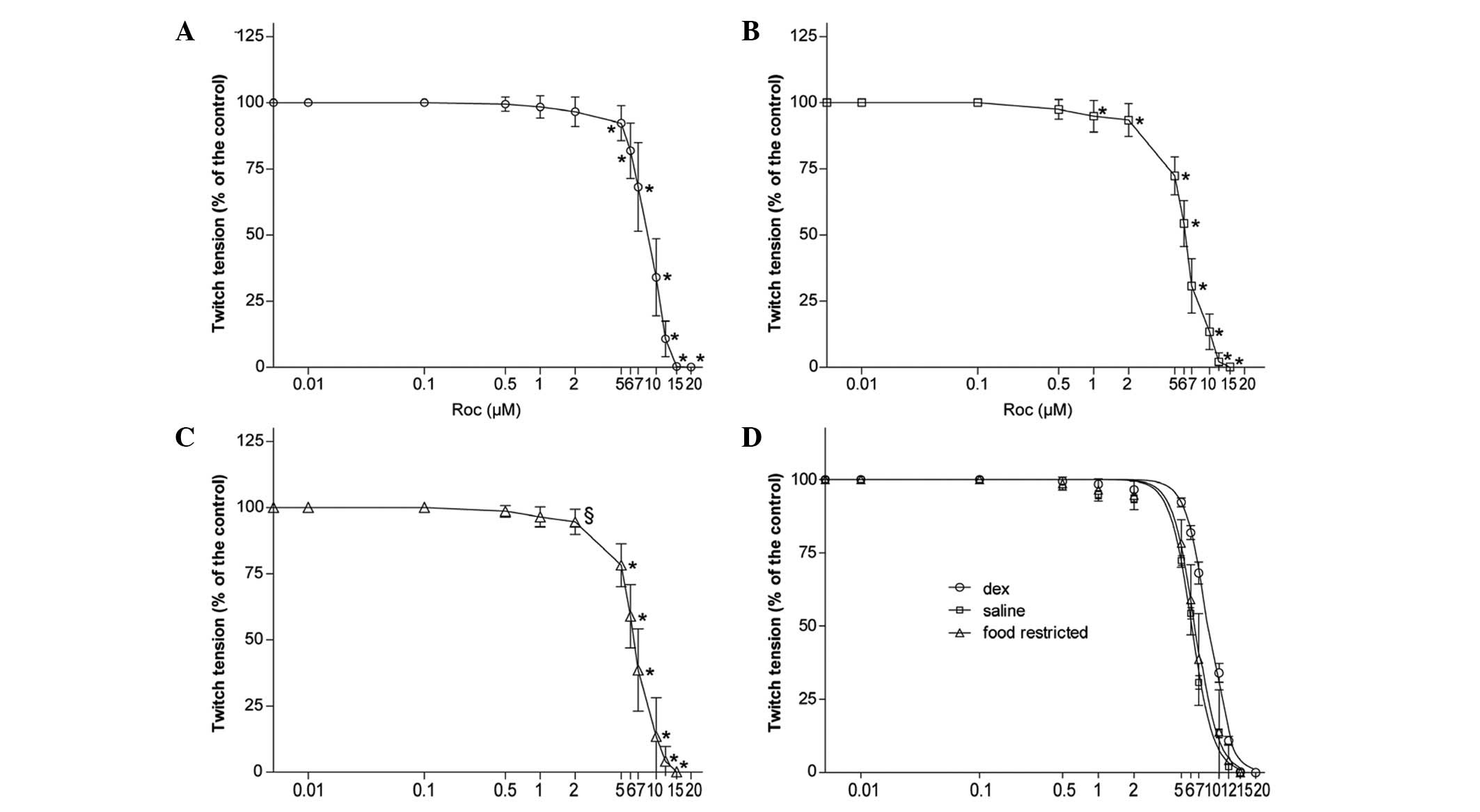
Rocuronium potency
Following rinsing of the diaphragms exposed to
rocuronium with Krebs solution, the indirectly elicited twitch
tension returned to 95–105% of the initial value in each study,
which indicated that the diaphragm remained responsive.
In all the groups, rocuronium reduced the magnitude
of indirectly elicited twitch tension in a concentration-dependent
manner (P<0.05 or P<0.01; Fig.
4A–C). IC50 values, which quantitatively indicate
the position of the dose-response curve for rocuronium, were
largest in the Dex group compared with the other two groups
(P<0.01; Table III). There
was no significant difference in rocuronium potency between the
saline and fr groups (P>0.05). There was no significant
difference in the slope among the three groups (P>0.05; Table III), which indicates no evident
drug interaction. Dex treatment significantly shifted the
concentration-twitch tension curve to the right compared with the
saline group (P<0.01), however, there was no Dex-dependent
effect compared with the fr group (P>0.05; Fig. 4D).
| Table IIIIC50 values and slopes of
the concentration-twitch tension curves of rocuronium. |
Table III
IC50 values and slopes of
the concentration-twitch tension curves of rocuronium.
| Rocuronium | Dex |
Food-restricted | Saline |
|---|
|
logIC50 |
0.920±0.0618a,b | 0.816±0.0532 | 0.786±0.0384 |
| Slope | −5.57±0.569 | −5.57±1.64 | −4.88±0.698 |
|
IC50 | 8.39±1.14 | 6.60±0.838 | 6.14±0.604 |
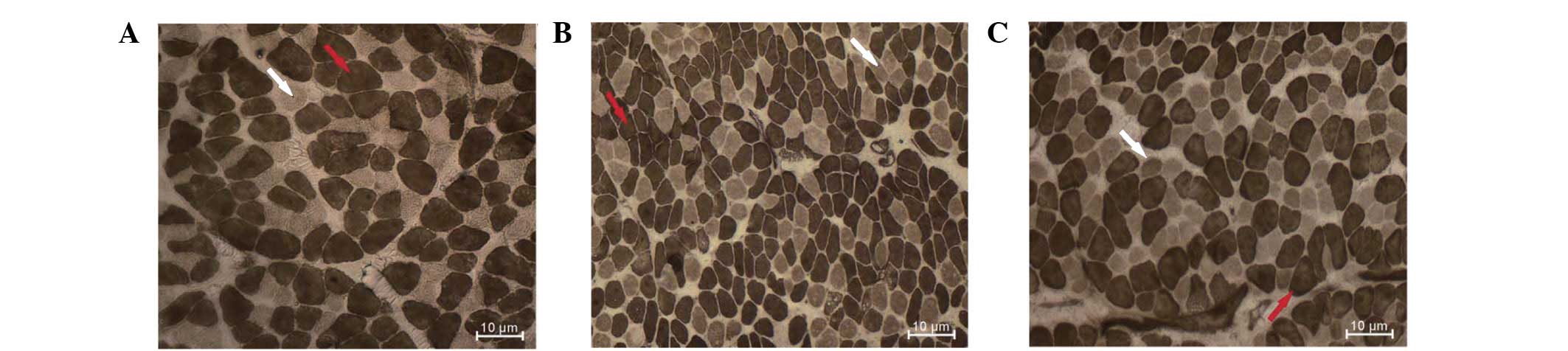
Histology and histochemistry
Histological examination of H&E-stained slides
showed a normal muscular pattern of the diaphragm in the three
groups. There was no inflammatory cell infiltration or
newly-generated muscle fibers observed in the Dex-treated
diaphragm. Morphometric analysis of ATPase-stained slides showed a
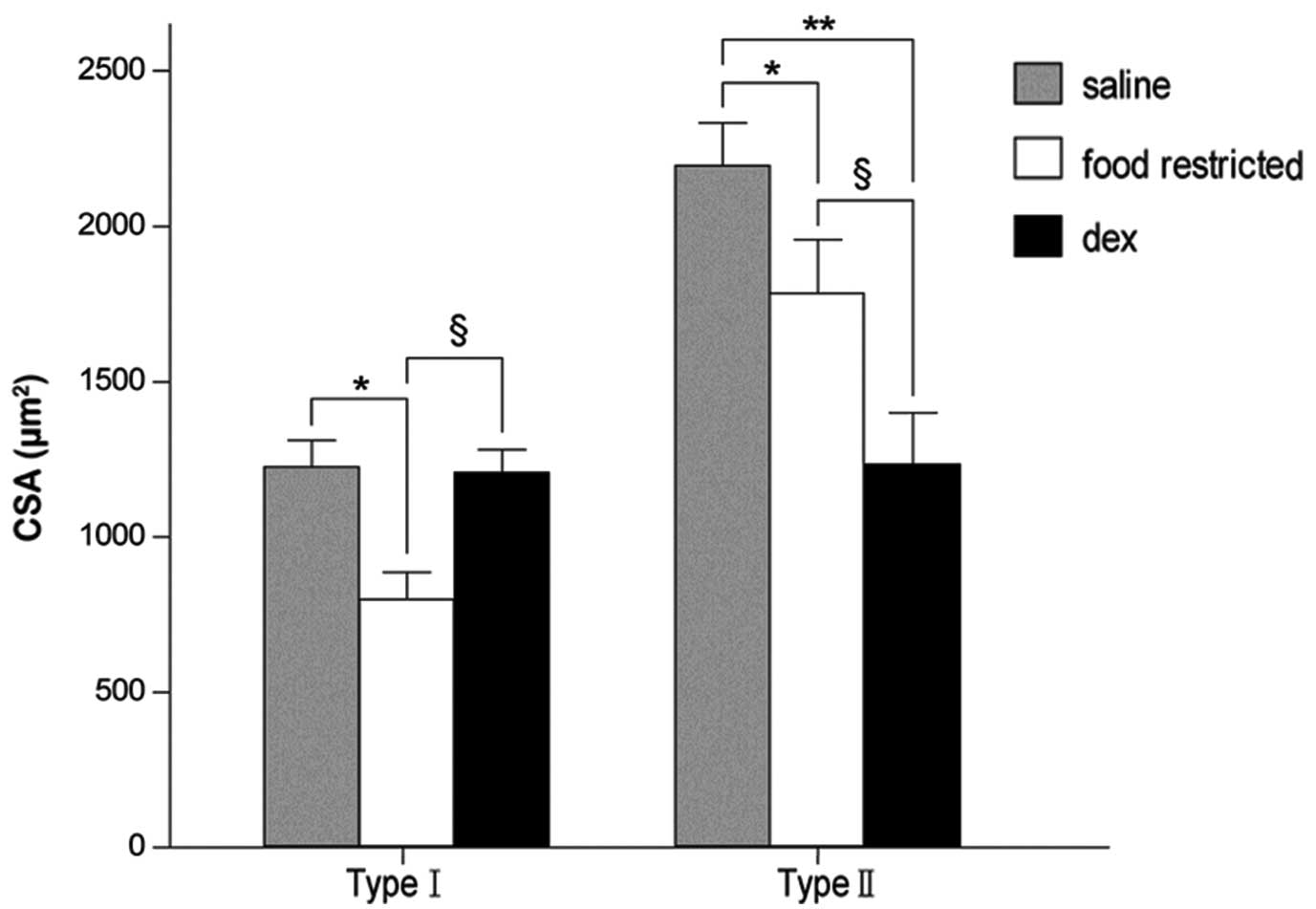
significant reduction in the percentage of type II fibers in the
Dex-treated group (P<0.05; Table
III). Dex treatment resulted in a significant reduction in the
CSA of type II fibers (P<0.05) with no change in the CSA of type
I fibers (P>0.05). In the fr group, there was generalized
atrophy of all the fiber types compared with the saline group
(P<0.05; Figs. 5 and 6; Table
IV).
| Table IVDiaphragm fiber distribution. |
Table IV
Diaphragm fiber distribution.
| Fiber-type
proportions, % total |
|---|
|
|
|---|
| Treatment | Type I | Type II |
|---|
| Dex | 45.5±2.2 | 54.2±1.9*§ |
| Saline | 34.2±1.4 | 63.8±1.7 |
| Fr | 33.8±1.6 | 66.5±2.4 |
Discussion
Over the 14-day period, the normal increase in body
weight observed in rats treated with saline was attenuated by Dex
treatment. Since alterations in nutritional status alone affect the
morphology and function of the rat diaphragm, interpretation of the
direct effects of Dex treatment is confounded (42,43).
The results of the current study demonstrate that
chronic Dex administration induces a selective atrophy and a
reduced distribution of type II fibers in the rat diaphragm,
indicating a shift towards slow fibers. This observation is
consistent with previous studies (24–26).
Matched food intake (fr group) caused a general atrophy of type I
and II fibers and the composition of type I and II fibers was
unaltered, which is consistent with observations from previous
studies (26,44,45).
In agreement with these morphological changes, alterations in
contractile properties were observed. Prolonged TPT and 1/2RT,
reduced force generation at a wide range of high stimulus
frequencies and resistance to fatigue, observed in the Dex group,
are all consistent with the changes of muscle morphology. No
significant change of muscle function was observed in the pair-fed
(fr) group.
The morphological and contractile adaptations of the
diaphragm in the pair-fed group were generally dissimilar to those
observed in the Dex-treated animals. These results are consistent
with previous studies indicating that the effects of GC treatment
on diaphragm structural and functional properties may not be
attributed to a non-selective catabolic effect alone (44–46).
In addition to the changes in diaphragm function and
structure, a marked decrease was observed in the potency of the
neuromuscular blocking action of rocuronium. In the current study,
the concentration-twitch tension curve of rocuronium shifted to the
right in the Dex-treated rats but not in the pair-fed rats,
indicating that weight loss was caused by GC-associated anorexia
itself and cannot account for the change in rocuronium potency.
Therefore, the resistance to rocuronium is more likely due to
Dex-induced changes in diaphragm-fiber composition.
Mechanisms of the GC-induced improved neuromuscular
transmission and hyposensitivities to the neuromuscular blocking
actions have not been investigated as extensively as those of
muscle contractility. A number of studies have demonstrated changes
in the electrophysiological properties of GC-treated neuromuscular
junctions (NMJs) (10–15). For example, Van Wilgenburg et
al reported that low-dose Dex and prednisolone resulted in
increased miniature endplate potential (MEPP) amplitude at rat
diaphragm NMJs, whereas high doses decreased MEPP amplitude
(14,47). Similar results at low CS doses were
also reported by Dalkara and Onur (11). In their study, Wilson et al
reported that prednisolone facilitated the spontaneous release of
ACh in the rat diaphragm, as manifested by a 2–3-fold increase in
MEPP frequency (48). A reduction
in MEPP amplitude was observed, indicating an additional
postsynaptic effect. Leeuwin et al reported an increase in
synaptic vesicle size that is indicative of an increase in quantal
size for neuromuscular transmission (12).
If GC treatment leads to an increase in the quantal
release of ACh at the NMJ, it may be hypothesized that such an
effect may improve neuromuscular transmission. However, with
repetitive stimulation, an increase in the quantal release of ACh
may lead to a rapid depletion of transmitter stores and
neuromuscular transmission failure. Greater resistance to fatigue
was revaled in the present study and a number of previous studies
(24,26,28).
Furthermore, the majority of experiments aiming to clarify the
mechanism were designed under acute conditions, whereas chronic
administration of steroids is more common in clinical practice.
To the best of our knowledge, no animal studies have
been performed to simultaneously examine morphological and
contractile effects, as well as sensitivity to NDMRs. The present
study examined the effects of Dex, a commonly used GC, at a low
dose, well within the range used clinically, on rat diaphragm
structure, contractile properties and susceptibility to rocuronium.
The altered diaphragm function was confirmed to be associated with
fiber transformation. Furthermore, Dex-induced resistance to
rocuronium is hypothesized to be associated with morphological
changes.
Dex was selected as a suitable corticosteroid in the
present study as it is a fluorinated drug with minimal
mineralocorticoid content and marked GC activity (49). In humans, Dex is administered
intravenously or intramuscularly at 6 mg/day for thrombocytopenia
(50). Prolonged Dex
administration at doses of 4–16 mg daily are frequently used during
intensive care unit hospitalization (51). Dex was also selected since
long-term Dex administration has a marked antagonizing effect on
the neuromuscular block caused by hemicholinium-3 and
d-tubocurarine (13,52–54).
It is well-established that among various species, the male rat is
similar to humans with respect to Dex metabolism (55). Intravenous low-dose Dex (0.3–0.5
mg/kg/day) was demonstrated to be efficacious during endotoxemia
and pulmonary fibrosis in rats (56,57).
In the present study, 0.6 mg/kg Dex may be equivalent to a dose of
6 mg/day in a 60-kg individual, thus, comparable to the dose that
is occasionally used in clinical treatment.
Rocuronium was used in the current study due to its
chemical stability and common use in anesthesia (58). The concentration of rocuronium
corresponding to 50% block (EC50) under propofol
anesthesia has been reported as 1.66 μM. The concentrations used in
the present study were relevant to this effective dose (59,60).
A limitation of the current study is that the
persistent effect of Dex following discontinuation of GC was not
examined. In a previous study, triamcinolone-induced weight loss
and type II fiber atrophy recovered 3 weeks following cessation of
the therapy (61). In patients
with GC-induced myopathy, an increase in muscle strength may be
observed within 3–4 weeks following discontinuation of the GC
(16). In addition, it is unclear
whether resistance to rocuronium changes over time. These areas
require further investigation.
In conclusion, the results of the present study
demonstrate that chronic Dex administration induces resistance to
rocuronium in the rat diaphragm. A reduced distribution of type II
fiber and associated changes in diaphragm function was noted. The
observations indicate that resistance to rocuronium in a
steroid-treated rat diaphragm is associated with Dex-induced fiber
transformation.
Acknowledgements
This study was supported by the National Natural
Science Foundation of China (no. 81171845). The authors would like
to thank Liang Zhu (Department of Pharmacology, School of Medicine,
Shanghai Jiaotong University) for technical assistance and
support.
References
|
1
|
Azar I, Kumar D and Betcher AM: Resistance
to pancuronium in an asthmatic patient treated with aminophylline
and steroids. Can Anaesth Soc J. 29:280–282. 1982. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Laflin MJ: Interaction of pancuronium and
corticosteroids. Anesthesiology. 47:471–472. 1977. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Meyers EF: Partial recovery from
pancuronium neuromuscular blockade following hydrocortisone
administration. Anesthesiology. 46:148–150. 1977. View Article : Google Scholar
|
|
4
|
Soltész S, Mencke T, Stunz M, Diefenbach
C, Ziegeler S and Molter GP: Attenuation of a rocuronium-induced
neuromuscular block in patients receiving prednisolone. Acta
Anaesthesiol Scand. 53:443–448. 2009.PubMed/NCBI
|
|
5
|
Soltész S, Mencke T, Mey C, Röhrig S,
Diefenbach C and Molter GP: Influence of a continuous prednisolone
medication on the time course of neuromuscular block of atracurium
in patients with chronic inflammatory bowel disease. Br J Anaesth.
100:798–802. 2008.
|
|
6
|
Parr SM, Galletly DC and Robinson BJ:
Betamethasone-induced resistance to vecuronium: a potential problem
in neurosurgery? Anaesth Intensive Care. 19:103–105.
1991.PubMed/NCBI
|
|
7
|
Arts WF and Oosterhuis HJ: Effect of
prednisolone on neuromuscular blocking in mice in vivo. Neurology.
25:1088–1090. 1975. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Robinson BJ, Lee E, Rees D, Purdie GL and
Galletly DC: Betamethasone-induced resistance to neuromuscular
blockade: a comparison of atracurium and vecuronium in vitro.
Anesth Analg. 74:762–765. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Parr SM, Robinson BJ, Rees D and Galletly
DC: Interaction between betamethasone and vecuronium. Br J Anaesth.
67:447–451. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Dal Belo CA, Leite GB, Fontana MD, et al:
New evidence for a presynaptic action of prednisolone at
neuromuscular junctions. Muscle Nerve. 26:37–43. 2002.PubMed/NCBI
|
|
11
|
Dalkara T and Onur R: Facilitatory effects
of dexamethasone on neuromuscular transmission. Exp Neurol.
95:116–125. 1987. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Leeuwin RS, Njio KD, Belling GA and van
den Hoven S: Glucocorticoid-induced changes in synaptic vesicles of
rat phrenic nerve terminals. Arch Int Pharmacodyn Ther.
266:200–207. 1983.PubMed/NCBI
|
|
13
|
Leeuwin RS, Veldsema-Currie RD, van
Wilgenburg H and Ottenhof M: Effects of corticosteroids on
neuromuscular blocking actions of d-tubocurarine. Eur J Pharmacol.
69:165–173. 1981. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Van Wilgenburg H, Njio KD, Belling GA and
Van den Hoven S: Effects of corticosteroids on the myoneural
junction. A morphometric and electrophysiological study. Eur J
Pharmacol. 84:129–137. 1982.PubMed/NCBI
|
|
15
|
Veldsema-Currie RD, Wolters E and Leeuwin
RS: The effect of corticosteroids and hemicholinium-3 on choline
uptake and incorporation into acetylcholine in rat diaphragm. Eur J
Pharmacol. 35:399–402. 1976. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Pereira RM and Freire de Carvalho J:
Glucocorticoid-induced myopathy. Joint Bone Spine. 78:41–44. 2011.
View Article : Google Scholar
|
|
17
|
Yamamoto D, Maki T, Herningtyas EH, et al:
Branched-chain amino acids protect against dexamethasone-induced
soleus muscle atrophy in rats. Muscle Nerve. 41:819–827. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Inder WJ, Jang C, Obeyesekere VR and
Alford FP: Dexamethasone administration inhibits skeletal muscle
expression of the androgen receptor and IGF-1 - implications for
steroid-induced myopathy. Clin Endocrinol (Oxf). 73:126–132.
2010.
|
|
19
|
Schakman O, Gilson H, Kalista S and
Thissen JP: Mechanisms of muscle atrophy induced by
glucocorticoids. Horm Res. 72(Suppl 1): 36–41. 2009. View Article : Google Scholar
|
|
20
|
Schakman O, Gilson H and Thissen JP:
Mechanisms of glucocorticoid-induced myopathy. J Endocrinol.
197:1–10. 2008. View Article : Google Scholar
|
|
21
|
Nava S, Gayan-Ramirez G, Rollier H, et al:
Effects of acute steroid administration on ventilatory and
peripheral muscles in rats. Am J Respir Crit Care Med.
153:1888–1896. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
van Balkom RH, van der Heijden HF, van
Herwaarden CL and Dekhuijzen PN: Corticosteroid-induced myopathy of
the respiratory muscles. Neth J Med. 45:114–122. 1994.PubMed/NCBI
|
|
23
|
Dekhuijzen PN, Gayan-Ramirez G, de Bock V,
Dom R and Decramer M: Triamcinolone and prednisolone affect
contractile properties and histopathology of rat diaphragm
differently. J Clin Invest. 92:1534–1542. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Fletcher LK, Powers SK, Coombes JS, et al:
Glucocorticoid- induced alterations in the rate of diaphragmatic
fatigue. Pharmacol Res. 42:61–68. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
van Balkom RH, Dekhuijzen PN, Folgering
HT, Veerkamp JH, Fransen JA and van Herwaarden CL: Effects of
long-term low-dose methylprednisolone on rat diaphragm function and
structure. Muscle Nerve. 20:983–990. 1997.PubMed/NCBI
|
|
26
|
Van Balkom RH, Zhan WZ, Prakash YS,
Dekhuijzen PN and Sieck GC: Corticosteroid effects on isotonic
contractile properties of rat diaphragm muscle. J Appl Physiol.
83:1062–1067. 1997.PubMed/NCBI
|
|
27
|
Dekhuijzen PN, Gayan-Ramirez G, Bisschop
A, et al: Rat diaphragm contractility and histopathology are
affected differently by low dose treatment with methylprednisolone
and deflazacort. Eur Respir J. 8:824–830. 1995.PubMed/NCBI
|
|
28
|
Sieck GC, van Balkom RH, Prakash YS, Zhan
WZ and Dekhuijzen PN: Corticosteroid effects on diaphragm
neuromuscular junctions. J Appl Physiol. 86:114–122.
1999.PubMed/NCBI
|
|
29
|
Sieck GC and Prakash YS: Morphological
adaptations of neuromuscular junctions depend on fiber type. Can J
Appl Physiol. 22:197–230. 1997. View
Article : Google Scholar : PubMed/NCBI
|
|
30
|
Prakash YS, Miller SM, Huang M and Sieck
GC: Morphology of diaphragm neuromuscular junctions on different
fibre types. J Neurocytol. 25:88–100. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Hemmerling TM and Donati F: Neuromuscular
blockade at the larynx, the diaphragm and the corrugator supercilii
muscle: a review. Can J Anaesth. 50:779–794. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Hemmerling TM, Schmidt J, Hanusa C, Wolf T
and Schmitt H: Simultaneous determination of neuromuscular block at
the larynx, diaphragm, adductor pollicis, orbicularis oculi and
corrugator supercilii muscles. Br J Anaesth. 85:856–860. 2000.
View Article : Google Scholar
|
|
33
|
Cantineau JP, Porte F, d’Honneur G and
Duvaldestin P: Neuromuscular effects of rocuronium on the diaphragm
and adductor pollicis muscles in anesthetized patients.
Anesthesiology. 81:585–590. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Donati F, Meistelman C and Plaud B:
Vecuronium neuromuscular blockade at the diaphragm, the orbicularis
oculi, and adductor pollicis muscles. Anesthesiology. 73:870–875.
1990. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Pansard JL, Chauvin M, Lebrault C, Gauneau
P and Duvaldestin P: Effect of an intubating dose of
succinylcholine and atracurium on the diaphragm and the adductor
pollicis muscle in humans. Anesthesiology. 67:326–330. 1987.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Donati F, Meistelman C and Plaud B:
Vecuronium neuromuscular blockade at the adductor muscles of the
larynx and adductor pollicis. Anesthesiology. 74:833–837. 1991.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Day NS, Blake GJ, Standaert FG and
Dretchen KL: Characterization of the train-of-four response in fast
and slow muscles: effect of d-tubocurarine, pancuronium, and
vecuronium. Anesthesiology. 58:414–417. 1983. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Ma K, Mallidis C, Bhasin S, et al:
Glucocorticoid-induced skeletal muscle atrophy is associated with
upregulation of myostatin gene expression. Am J Physiol Endocrinol
Metab. 285:E363–E371. 2003.PubMed/NCBI
|
|
39
|
Burke RE, Levine DN, Tsairis P and Zajac
FE III: Physiological types and histochemical profiles in motor
units of the cat gastrocnemius. J Physiol. 234:723–748. 1973.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Brooke MH and Kaiser KK: Three ‘myosin
adenosine triphosphatase’ systems: the nature of their pH lability
and sulfhydryl dependence. J Histochem Cytochem. 18:670–672.
1970.
|
|
41
|
Peter JB, Barnard RJ, Edgerton VR,
Gillespie CA and Stempel KE: Metabolic profiles of three fiber
types of skeletal muscle in guinea pigs and rabbits. Biochemistry.
11:2627–2633. 1972. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Sieck GC, Lewis MI and Blanco CE: Effects
of undernutrition on diaphragm fiber size, SDH activity, and
fatigue resistance. J Appl Physiol. 66:2196–2205. 1989.PubMed/NCBI
|
|
43
|
Lewis MI, Sieck GC, Fournier M and Belman
MJ: Effect of nutritional deprivation on diaphragm contractility
and muscle fiber size. J Appl Physiol. 60:596–603. 1986.PubMed/NCBI
|
|
44
|
Koerts-De Lang E, Schols AM, Wouters EF,
Gayan-Ramirez G and Decramer M: Contractile properties and
histochemical characteristics of the rat diaphragm after prolonged
triamcinolone treatment and nutritional deprivation. J Muscle Res
Cell Motil. 19:549–555. 1998.
|
|
45
|
Dekhuijzen PN, Gayan-Ramirez G, Bisschop
A, De Bock V, Dom R and Decramer M: Corticosteroid treatment and
nutritional deprivation cause a different pattern of atrophy in rat
diaphragm. J Appl Physiol. 78:629–637. 1995.PubMed/NCBI
|
|
46
|
Lewis MI and Sieck GC: Effect of acute
nutritional deprivation on diaphragm structure and function in
adolescent rats. J Appl Physiol. 73:974–978. 1992.PubMed/NCBI
|
|
47
|
Van Wilgenburg H: The effect of
prednisolone on neuromuscular transmission in the rat diaphragm.
Eur J Pharmacol. 55:355–361. 1979.
|
|
48
|
Wilson RW, Ward MD and Johns TR:
Corticosteroids: a direct effect at the neuromuscular junction.
Neurology. 24:1091–1095. 1974. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Kotsarini C, Griffiths PD, Wilkinson ID
and Hoggard N: A systematic review of the literature on the effects
of dexamethasone on the brain from in vivo human-based studies:
implications for physiological brain imaging of patients with
intracranial tumors. Neurosurgery. 67:1799–1815. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Kularatne SA, Walathara C, Mahindawansa
SI, et al: Efficacy of low dose dexamethasone in severe
thrombocytopenia caused by dengue fever: a placebo controlled
study. Postgrad Med J. 85:525–529. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Hardy JR, Rees E, Ling J, et al: A
prospective survey of the use of dexamethasone on a palliative care
unit. Palliat Med. 15:3–8. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Leeuwin RS and Wolters EC: Effect of
corticosteroids on sciatic nerve-tibialis anterior muscle of rats
treated with hemicholinium-3. An experimental approach to a
possible mechanism of action of corticosteroids in myasthenia
gravis. Neurology. 27:171–177. 1977. View Article : Google Scholar
|
|
53
|
Arts WF and Oosterhuis HJ: Long-term
effect of glucocorticosteroids on neuromuscular blocking in mice. J
Neurol Neurosurg Psychiatry. 40:675–677. 1977. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Wolters MJ and Leeuwin RS: Effect of
corticosteroids on the phrenic nerve-diaphragm of preparation
treated with hemicholinium. A possible model of myasthenia gravis.
Neurology. 26:574–578. 1976. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Tomlinson ES, Maggs JL, Park BK and Back
DJ: Dexamethasone metabolism in vitro: species differences. J
Steroid Biochem Mol Biol. 62:345–352. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Crossland H, Constantin-Teodosiu D,
Greenhaff PL and Gardiner SM: Low-dose dexamethasone prevents
endotoxaemia-induced muscle protein loss and impairment of
carbohydrate oxidation in rat skeletal muscle. J Physiol.
588:1333–1347. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Kitano M, Ishinaga H, Shimizu T, Takeuchi
K and Majima Y: Effects of clarithromycin and dexamethasone on
mucus production in isografted rat trachea. Pharmacology. 87:56–62.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Proost JH, Eriksson LI, Mirakhur RK, Roest
G and Wierda JM: Urinary, biliary and faecal excretion of
rocuronium in humans. Br J Anaesth. 85:717–723. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Ito S, Nagata O and Ozaki M: Estimated
blood concentration of rocuronium administrated by continuous
infusion to maintain an appropriate neuromuscular blockade under
propofol anesthesia. Masui. 59:82–86. 2010.(In Japanese).
|
|
60
|
Dragne A, Varin F, Plaud B and Donati F:
Rocuronium pharmacokinetic-pharmacodynamic relationship under
stable propofol or isoflurane anesthesia. Can J Anaesth.
49:353–360. 2002. View Article : Google Scholar
|
|
61
|
Braunstein PW Jr and DeGirolami U:
Experimental corticosteroid myopathy. Acta Neuropathol. 55:167–172.
1981. View Article : Google Scholar
|