Introduction
An abdominal aortic aneurysm (AAA) is a localized,
permanent dilatation of the aorta that affects ~8% of males >65
years-old (1). At present,
elective surgery is the major therapeutic option for AAA, however,
this is not applicable to small aneurysms despite the reported
growth rate of small aneurysms ranging between 1.5 and 3 mm per
year, which leads to a higher risk of rupture (2). With increased knowledge of aneurysm
pathophysiology, it is possible that aneurysm growth may be
retarded with medical therapy.
The role of inflammation in the pathogenesis of AAA
is well established. Infiltrating inflammatory cells enter the
aorta, release cytokines and proteases, inducing apoptosis of
vascular smooth muscle cells and ultimately, lead to destruction of
the vascular wall (1). Moreover,
the inflammatory microenvironment generates a large quantity of
oxidant species, largely through upregulation of nicotinamide
adenine dinucleotide phosphate (NADPH) oxidase in vascular cells
(3). Furthermore, emerging
evidence indicates that oxidative stress within the aortic wall is
closely involved in the pathogenesis of AAA. Oxidative stress
facilitates leukocyte recruitment into the vasculature by
modulating adhesion molecules and chemotactic cytokines (4). In addition, reactive oxygen species
(ROS) may alter the balance between destruction and regeneration of
the aortic wall by enhancing matrix proteolysis through
upregulation of matrix metalloproteinases (MMPs) (5). MMPs are the predominant extracellular
proteinases that participate in the degradation process of
structural proteins (1). Although
human data remains limited, several studies indicate that
antioxidant therapy may be effective in experimental AAA models
(6–8).
Quercetin (3,5,7,3′4′-Pentahydroxy flavon), a
typical member of the flavonoid family, is one of the most widely
recognized dietary polyphenolic compounds. It is ubiquitously
present in foods and is claimed to exert beneficial effects on
vascular disease (9), which has
been largely associated with its antioxidant and anti-inflammatory
properties. Within the flavonoid family, quercetin is proven to be
the most potent scavenger of free radicals (10). There is evidence that quercetin
reduces low-density lipoprotein oxidation (11) and prevents the development of
atherosclerotic lesions (12), in
which oxidative stress is assumed to have a pivotal role. Although
atherosclerosis and AAA are separate diseases, they have certain
similar pathological characteristics, including inflammation and
proteolysis (13). It is also
reported that quercetin in vitro inhibits the production of
O2•− in the rat aorta and decreases protein
expression of the NADPH oxidase subunit, p47phox (14,15).
A previous study from our research group indicated that quercetin
treatment inhibits inflammation and prevents
CaCl2-induced aneurysmal dilation in a mouse AAA model
(16). The present study was
designed to test the hypothesis that an antioxidative mechanism is
also involved in the protection afforded by quercetin.
Materials and methods
Pharmacological treatments
Quercetin was purchased from Sigma-Aldrich (Q4951;
Shanghai, China). Drug solutions were prepared by suspending the
compound in 0.5% carboxymethyl cellulose sodium. Animals were
gavaged daily with 0.1 ml solution of quercetin (60 mg/kg) or
vehicle alone, which began 2 weeks prior to AAA induction and
continued for 8 weeks. The dose regimen for quercetin was based on
previous studies demonstrating beneficial effects of the drug in
mouse models of aortic atherosclerosis (12).
Animal groups and the AAA model
A total of 60 male C57BL/6 wild-type mice (age, 6–7
weeks) were obtained from Vital River Laboratory Animal Technology
(Beijing, China). All animals were treated and cared for in
accordance with the Guide for the Care and Use of Laboratory
Animals (National Institutes of Health, Washington DC, 1996) and
the experimental protocols were approved by the Animal Care and Use
Committee (Nanjing University, Nanjing, China). The mice were
randomly assigned to one of four groups (n=15 in each group):
Vehicle treatment plus sham operation control (VC), vehicle
treatment plus AAA (VA), quercetin treatment plus AAA (QA) and
quercetin treatment plus sham operation control (QC). AAA was
induced in the infrarenal abdominal aorta (age, 8 weeks) by
periaortic application of CaCl2, as previously described
(16). NaCl (0.9%) was substituted
for CaCl2 in sham operation animals. Six weeks later,
the mice were laparotomied and the aortic diameters (ADs) were
measured; the abdominal incision was carried upwards as a
thoracoabdominal incision, the animals were then sacrificed by
left-heart injection of potassium chloride and the aortic tissues
were collected. An aneurysm was defined as an increase in the AD of
>50% of the original AD.
In vivo hemodynamic measurements
A computerized, non-invasive tail-cuff system with a
four-channel mouse platform (BP-2000; Visitech Systems, Inc., Apex,
NC, USA) was used to measure blood pressure and heart rate. To
train mice, daily measurements were performed for five consecutive
days prior to the actual recorded measurements. Hemodynamic
parameters were measured one day pre- and 6 weeks post-AAA
induction. The first 10 of 30 values recorded at each session were
disregarded and the remaining 20 values were averaged and used for
analysis, according to the manufacturer’s instructions.
ROS analysis, lipid peroxidation
determination and manganese-superoxide dismutase (Mn-SOD) activity
assay
Dihydroethidium (DHE) oxidative fluorescence dye was
used to evaluate in situ production of ROS (17). DHE stock solution was prepared by
dissolving DHE (D7008; Sigma-Aldrich) in dimethylsulfoxide at a
concentration of 5 mM. The stock solution was stored in the dark
and diluted in phosphate-buffered saline (PBS) to a final
concentration of 5 μM immediately prior to use. The abdominal aorta
was harvested and the aortic segment (10 mm) was embedded in
Tissue-Tek OCT compound (Sakura Finetech Japan, Tokyo, Japan) and
snap-frozen. DHE working solution (200 μl) was topically applied to
the aortic sections and the slides were subsequently incubated at
37°C in the dark for 30 min. Excess DHE was rinsed off twice with
PBS and the images were immediately captured with a fluorescent
microscope (BX51; Olympus, Tokyo, Japan) at excitation and emission
wavelengths of 520 and 610 nm, respectively.
A portion of the snap-frozen aortic tissue (n=5 per
group) was crushed in a prechilled mortar and resuspended in PBS at
a concentration of 50 mg/ml. The homogenate was centrifuged at
10,000 × g for 10 min at 4°C to collect the supernatant. The lipid
peroxidation product, malondialdehyde (MDA), was assessed using the
thiobarbituric acid reactive substances (TBARS) assay kit (A003;
Jiancheng Bioengineering, Shanghai, China). Briefly, 100 μl
supernatant was added to 100 μl sodium dodecyl sulfate (SDS) lysis
solution and mixed thoroughly. Following the addition of 250 μl
thiobarbituric acid (TBA) reagent, samples were incubated at 95°C
for 1 h and centrifuged at 1,500 × g at room temperature for 15
min. The absorbance of each supernatant was measured at 532 nm
using a spectrophotometer (BioPhotometer; Eppendorf, Hamburg,
Germany). Values of TBARS are expressed as nmol equivalents of MDA
per mg protein. Mn-SOD activity was measured using an assay kit
(A001-2; Jiancheng Bioengineering) according to the manufacturer’s
instructions. Assay conditions were 65 μmol phosphate buffer (pH
7.8), 1 μmol hydrochloric hydroxylamine, 0.75 μmol xanthine and
2.3×10−3 IU xanthine dismutase. The supernatant (50 μl)
was incubated in the system for 40 min at 37°C and terminated with
2 ml 3.3 g/l p-aminobenzene sulfonic acid and 10 g/l naphthylamine.
For inhibition of CuZn-SOD activity, the assay was conducted in the
presence of 10 mm KCN following preincubation for 30 min. The
supernatant was transferred to a microplate (Eppendorf) for
determination of the absorbance at 550 nm and 1 unit SOD was
defined as the quantity of enzyme required to produce 50%
dismutation of superoxide radical. Mn-SOD activity was calculated
by subtraction of CuZn-SOD activity from total SOD activity. The
standard curves were created as described in the manufacturer’s
instructions. Images were assessed by Image J 1.44 software
(National Institute of Health, Bethesda, MD, USA).
Histological analysis
The infrarenal abdominal aorta (n=5 per group) was
dissected and fixed in 10% neutral-buffered formalin. Specimens
were dehydrated through graded ethanols, embedded in paraffin and
sliced into 4–6-μm sections. Immunohistochemical staining with a
rabbit polyclonal anti-nitrotyrosine antibody (1:500; 06–284;
Millipore, Temecula, CA, USA) was used as an indicator of
peroxynitrite formation (18).
Briefly, the slides were incubated in 3% hydrogen peroxide for 5
min to quench endogenous peroxidase activity and were then
incubated with primary antibody overnight at 4°C. Subsequently,
slides were washed with PBS and incubated (15 min; 37°C) with
peroxidase-conjugated goat anti-rabbit IgG (AP132P; Millipore).
Finally, the slides were incubated with diaminobenzidine and
counterstained with hematoxylin.
Reverse transcription-polymerase chain
reaction (RT-PCR)
RT-PCR was used to define the expression of
glutathione peroxidase (GPx)-1, GPx-3, inducible nitric oxide
synthase (iNOS) and p47phox NADPH oxidase mRNA. Total RNA was
prepared with the TRIzol total RNA extraction kit (SK1321; Sangon
Biotech, Shanghai, China). Primer sequences were as follows:
Forward: 5′-ACC CCA AGT ACA TCA TTT GGT C-3′ and reverse: 5′-GCA
GGG TTT CTA TGT CAG GTT C-3′ for GPx-1; forward: 5′-ATC TAC GAG TAT
GGA GCC CTC A-3′ and reverse: 5′-GGC CCA AGT TCT TCT TGT AGT G-3′
for GPx-3; forward: 5′-CTT TGA CGC TCG GAA CTG TAG-3′ and reverse:
5′-AAC TCC AAG GTG GCA GCA T-3′ for iNOS; forward: 5′-CCC ATC ATC
CTT CAG ACC TAT C-3′ and reverse: 5′-AAC CTC GCT TTG TCT TCA TCT
G-3′ for p47 NADPH oxidase; and forward: 5′-AGG CCG GTG CTG AGT ATG
TC-3′ and reverse: 5′-TGC CTG CTT CAC CAC CTT CT-3′ for GAPDH.
Reverse transcription was performed with oligo-dT primers and the
AMV First Strand cDNA Synthesis kit (SK2029; Sangon Biotech),
according to the manufacturer’s instructions. The resultant cDNA
was amplified by Taq DNA polymerase (SK2442; Sangon Biotech)
in an Access RT-PCR System (Promega Corp., Madison, WI, USA). GAPDH
mRNA was also amplified to serve as an internal control. The
resultant PCR products were detected using an MSF-300G Scanner
(Microtek Lab, Carson, CA, USA) and expressed as the ratio to
GAPDH.
Western blotting
Total protein was extracted from the supernatants of
tissue homogenate with T-PER tissue protein extraction reagent
(Pierce Biotechnology Inc., Rockford, IL, USA) and stored at −80°C.
Equal quantities (30 μg) of total protein were separated on 10%
polyacrylamide gels and transferred to nitrocellulose membranes
using a semidry transfer cell (#164-5052; Bio-Rad, Hercules, CA,
USA) at 10 V for 40 min. The membranes were blocked for 60 min with
5% nonfat milk in Tris-buffered saline with Tween-20 (TBST) and
subsequently washed. Primary antibodies for p47phox NADPH oxidase
(sc-14015), c-Jun N-terminal kinase (JNK; sc-571) and
phosphorylated JNK (sc-6254) (all Santa Cruz Biotechnology, Santa
Cruz, CA, USA) were added at a 1:500 dilution and incubated
overnight at 4°C. Additionally, all blots were incubated with the
anti-β-actin antibody (1:5,000; 4970; Cell Signaling Technology,
Beverly, MA, USA) to confirm protein loading levels. Membranes were
washed with TBST, incubated with horseradish peroxidase-conjugated
species-appropriate secondary antibodies (Santa Cruz Biotechnology)
for 1 h at room temperature and developed using an enhanced
chemiluminescence kit (Pierce, Rockford, IL, USA). Quantification
of images was performed by scanning densitometry with Image J
1.44.
Electrophoretic mobility shift assay
(EMSA)
Nuclear protein lysates were harvested using NE-PER
nuclear and cytoplasmic extraction reagents (Pierce Biotechnology,
Inc.) according to the manufacturer’s instructions. Activator
protein (AP)-1 DNA-binding activities were analyzed using Gel Shift
Assay systems (Promega Corporation, Madison, WI, USA), according to
the instructions previously described (16). Briefly, the AP-1 consensus
oligonucleotide probe (5′-CGC TTG ATG AGT CAG CCG GAA-3′) was
end-labeled with [γ-32P]-ATP (Furui Biotech, Beijing,
China). The extracted nuclear proteins (10 μg) were incubated for
20 min at 37°C with the 32P-labeled oligonucleotide
(0.30 pmol) in a binding buffer. Reaction products were then
separated in a 4% polyacrylamide gel, followed by autoradiography.
The reactive bands were quantified as described in western blot
analysis.
Gelatin zymography
Protein extracts (10 μg) were mixed with SDS buffer
and separated by electrophoresis on 10% SDS-polyacrylamide gels
containing 1.0% gelatin. Following electrophoresis, the gels were
renatured in renaturing buffer (LC2670; Invitrogen Life
Technologies, Carlsbad, CA, USA) and incubated with developing
buffer (LC2671; Invitrogen Life Technologies) for 30 min at room
temperature. Subsequently, the gel was incubated in fresh
developing buffer overnight at 37°C. The gel was stained with 0.5%
Coomassie blue R-250 for 30 min and destained with destaining
solution containing 10% acetic acid and 40% methanol. The relative
molecular weight of each band was determined using protein
standards (Pierce Biotechnology, Inc.). Areas of protease activity
appeared as unstained bands against a blue background. Images were
assessed by Image J 1.44.
Statistical Analysis
All values are expressed as the mean ± standard
deviation. Statistical analyses were performed with SPSS for
Windows version 17.0 (SPSS, Inc., Chicago, IL, USA). Within-group
comparisons of hemodynamic parameters at various intervals were
performed using paired Student’s t tests. Between-group comparisons
were performed using the Fisher’s exact test or analysis of
variance. P<0.05 was considered to indicate a statistically
significant difference.
Results
AAA incidence
No significant difference was found in AD at the
time of surgery among the groups (data not shown). Six weeks later,
the VA mice showed a marked increase in AD following
CaCl2 treatment with 10/15 (66.7%) developing aneurysms.
Only 3/15 (20%) of the aortas became aneurysmal in QA mice; this
difference in aneurysm incidence was considered to be highly
significant (P<0.05). No aneurysm formation was observed in the
VC or QC mice. Quercetin treatment had no effect on mean arterial
pressure or heart rate when measured pre- and post-surgery
(Table I).
 | Table IHeart rate and mean arterial pressure
prior to and following NaCl/CaCl2 treatment. |
Table I
Heart rate and mean arterial pressure
prior to and following NaCl/CaCl2 treatment.
| 2 weeks prior to
surgery | Surgery | 6 weeks
post-surgery |
|---|
|
|
|
|
|---|
| Group (n=15) | HR, bpm | MAP, mmHg | HR, bpm | MAP, mmHg | HR, bpm | MAP, mmHg |
|---|
| VC | 630±30 | 83±8 | 626±34 | 85±6 | 633±27 | 85±6 |
| VA | 628±28 | 85±7 | 632±29 | 82±5 | 631±35 | 86±9 |
| QA | 631±33 | 84±6 | 629±33 | 84±8 | 627±34 | 83±5 |
| QC | 634±31 | 84±9 | 633±34 | 85±5 | 630±32 | 85±8 |
ROS generation, nitrotyrosine formation
and lipid peroxidation production
To evaluate the effect of quercetin on ROS
generation, aortic sections were exposed to DHE, which is
transformed to the highly fluorescent molecule, oxyethidium, in the
presence of superoxide (17). As
shown in Fig. 1A, ROS production
was extremely low in aortas from VC and QC mice. At 6 weeks
post-AAA induction in VA mice, oxyethidium fluorescence was higher,
being significantly enhanced throughout the vascular wall (Fig. 1A and B). However, it was attenuated
in QA mice, indicating decreased ROS production due to quercetin
treatment.
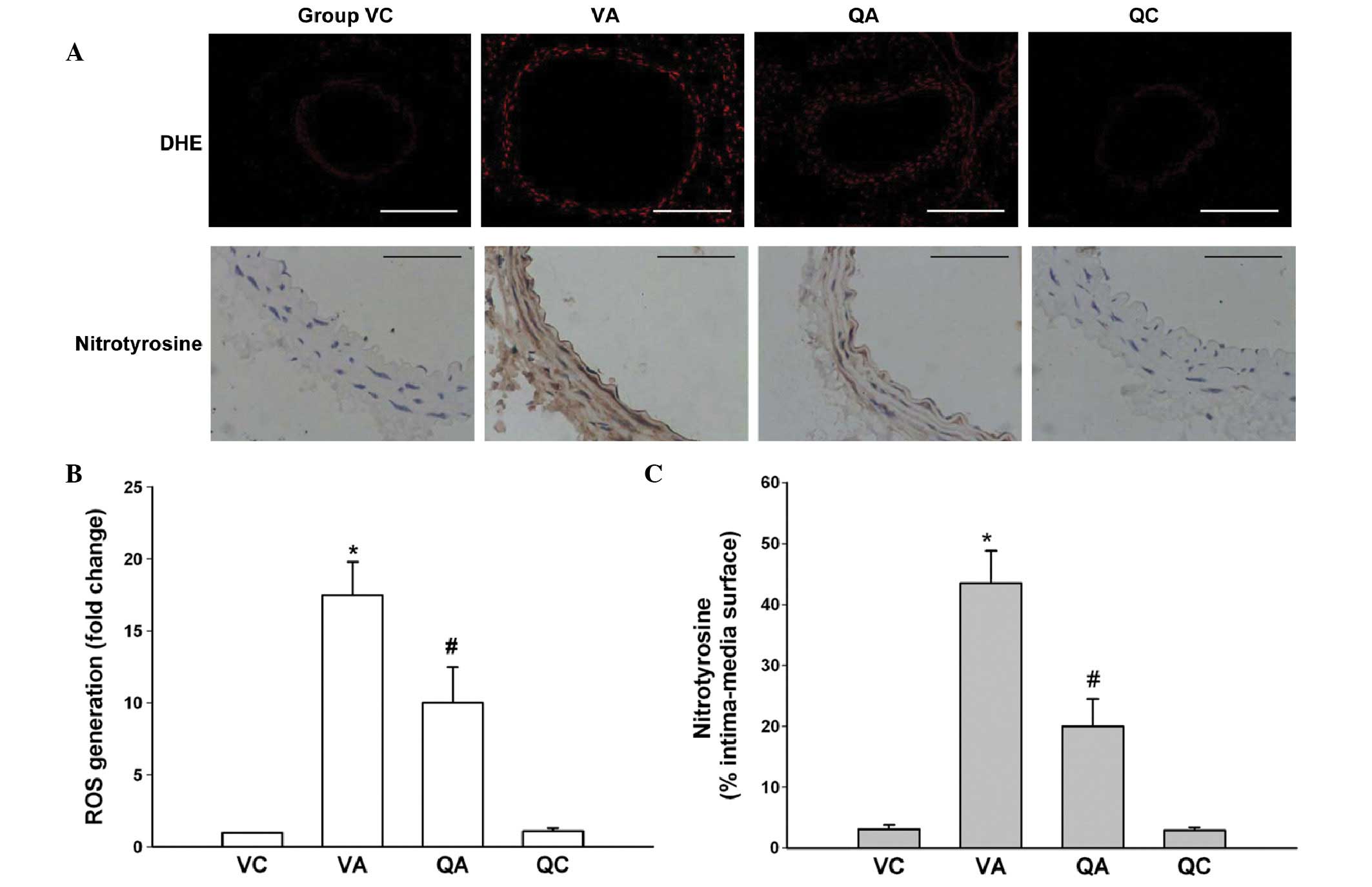 | Figure 1Effect of quercetin on ROS generation
and nitrotyrosine formation in the aortic wall. (A) DHE
fluorescence imaging and immunohistochemical staining for
nitrotyrosine in the mouse aortic wall (details in Materials and
methods). Scale bars are 500 and 50 μm, respectively. (B)
Densitometric analysis of DHE fluorescence relative to VC mice. (C)
Positively stained areas in the aortic section of each group.
Values are presented as the mean ± standard deviation with n=5 per
group. *P<0.05, vs. VC; #P<0.05, vs.
VA. ROS, reactive oxygen species; DHE, dihydroethidium; AAA,
abdominal aortic aneurysm; VC, vehicle treatment plus sham
operation control; VA, vehicle treatment plus AAA; QA, quercetin
treatment plus AAA; QC, quercetin treatment plus sham operation
control. |
As increased production of ROS may lead to further
peroxynitrite accumulation, which induces protein damage by
formation of nitrotyrosine (18),
immunohistochemistry was performed with a polyclonal antibody
against nitrotyrosine in aortic cross sections. Staining appeared
only weakly in the aorta of VC and QC animals, however, VA mice
revealed marked brown nitrotyrosine staining in the aortic wall. By
contrast, a decreased immunoreactivity was observed in QA mice
(Fig. 1A and C). Similar
observations were noted for lipid peroxidation production (MDA)
levels (TBARS) in aortic tissues. The TBARS concentration in QA
mice was found to be significantly lower than that in the VA mice
(Table II).
| Table IIEffect of quercetin on MDA levels and
Mn-SOD activities following NaCl/CaCl2 incubation. |
Table II
Effect of quercetin on MDA levels and
Mn-SOD activities following NaCl/CaCl2 incubation.
| Group | TBARS (nm MDA/mg
protein) | Mn-SOD activities
(U/mg protein) |
|---|
| VC | 0.075±0.027 | 2.8±1.5 |
| VA | 0.342±0.035a | 28.2±5.4a |
| QA | 0.231±0.029b | 15.5±4.9b |
| QC | 0.069±0.015 | 2.5±1.8 |
Endogenous vascular antioxidant defense
systems
Increased levels of Mn-SOD activity were observed at
AAA regions of the VA mice, while quercetin significantly decreased
its activity in QA mice (Table
II). In addition, quercetin caused a relative decrease in mRNA
expression of GPx-1 and GPx-3, which are involved in antioxidative
status (Fig. 2).
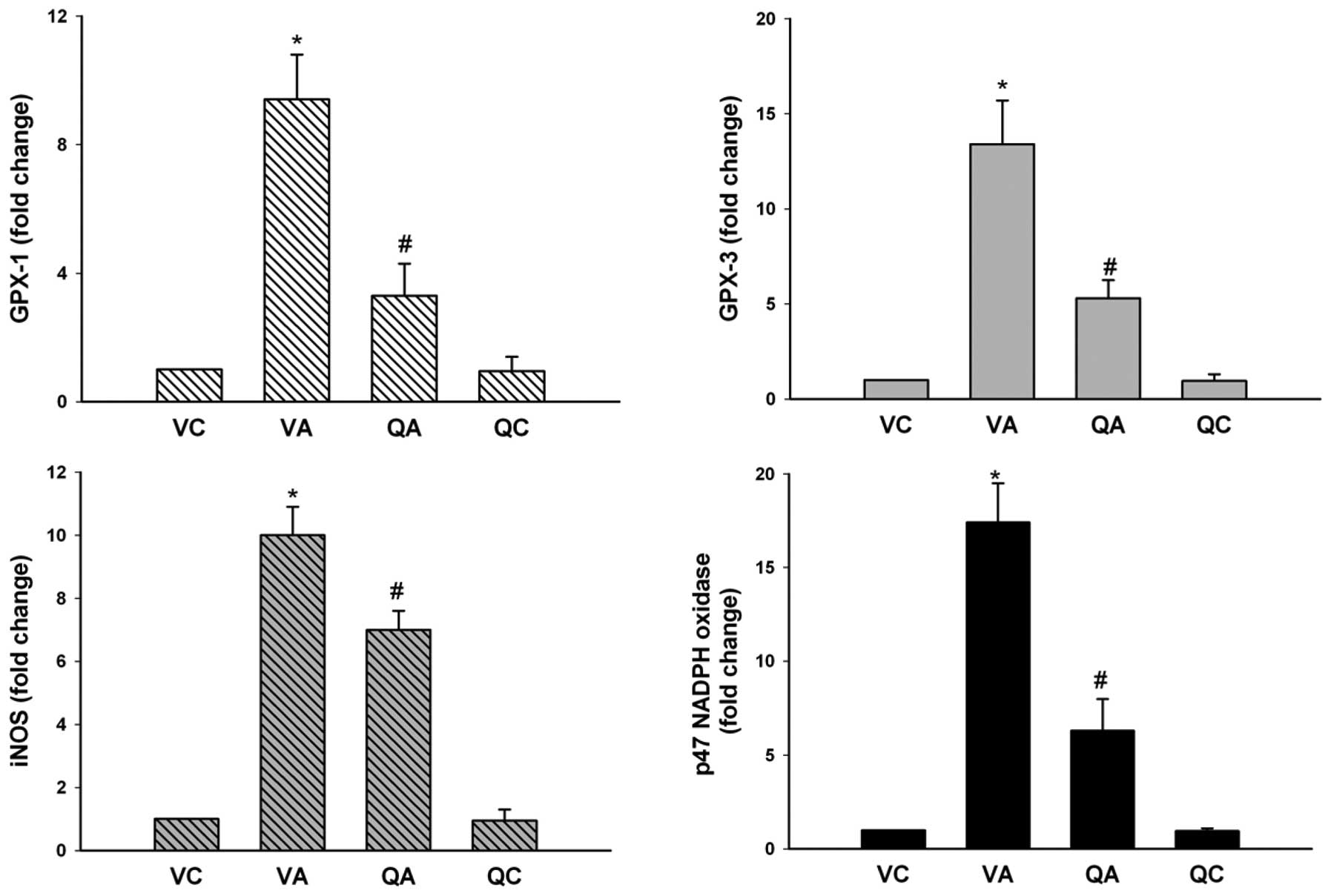 | Figure 2mRNA expression of GPx-1, GPx-3, iNOS
and p47 NADPH oxidase. Data are expressed as fold changes compared
with values of VC animals (n=5 in each group).
*P<0.05, vs. VC; #P<0.05, vs. VA. AAA,
abdominal aortic aneurysm; iNOS, inducible nitric oxide synthase;
Gx, glutathione peroxidase; NADPH, nicotinamide adenine
dinucleotide phosphate; VC, vehicle treatment plus sham operation
control; VA, vehicle treatment plus AAA; QA, quercetin treatment
plus AAA; QC, quercetin treatment plus sham operation control. |
Expression of iNOS and p47phox NADPH
oxidase
Expression of iNOS and the NADPH oxidase subunit,
p47phox, was also examined. Compared with the VA group, QA mice
showed relative decreases in iNOS and p47phox mRNA levels (Fig. 2) and this result was confirmed by
western blot analysis of p47phox (Fig.
3A and B).
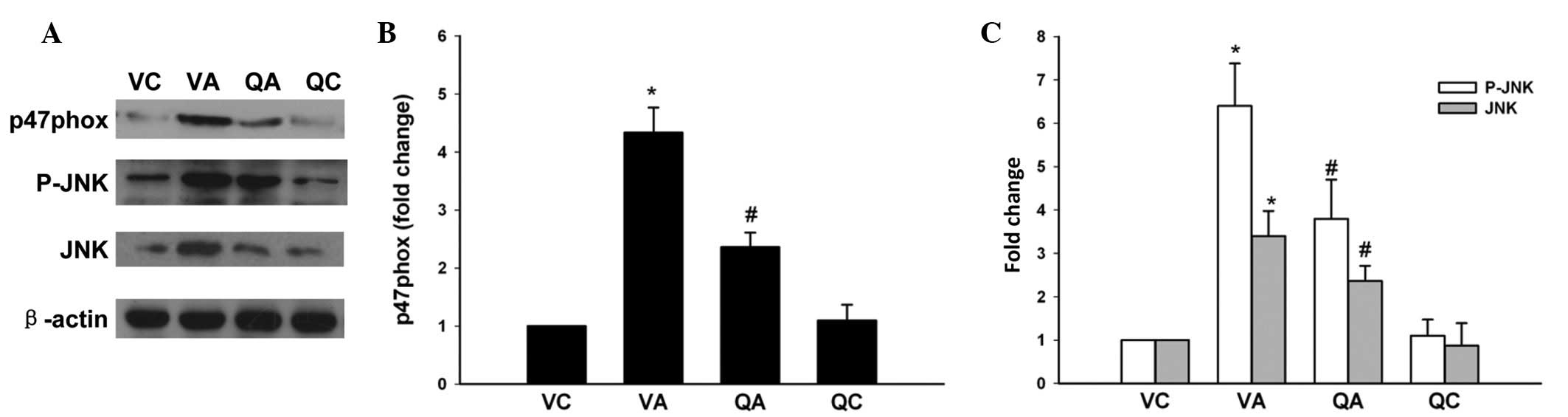 | Figure 3Expression of p47phox NADPH oxidase,
phosphorylated JNK and total JNK (A). Data are presented as the
mean ± standard deviation with n=5 per group and expressed as fold
changes compared with values in the VC animals (B).
*P<0.05, vs. VC; #P<0.05, vs. VA. AAA,
abdominal aortic aneurysm; JNK, c-Jun N-terminal kinase; NADPH,
nicotinamide adenine dinucleotide phosphate; VC, vehicle treatment
plus sham operation control; VA, vehicle treatment plus AAA; QA,
quercetin treatment plus AAA; QC, quercetin treatment plus sham
operation control. |
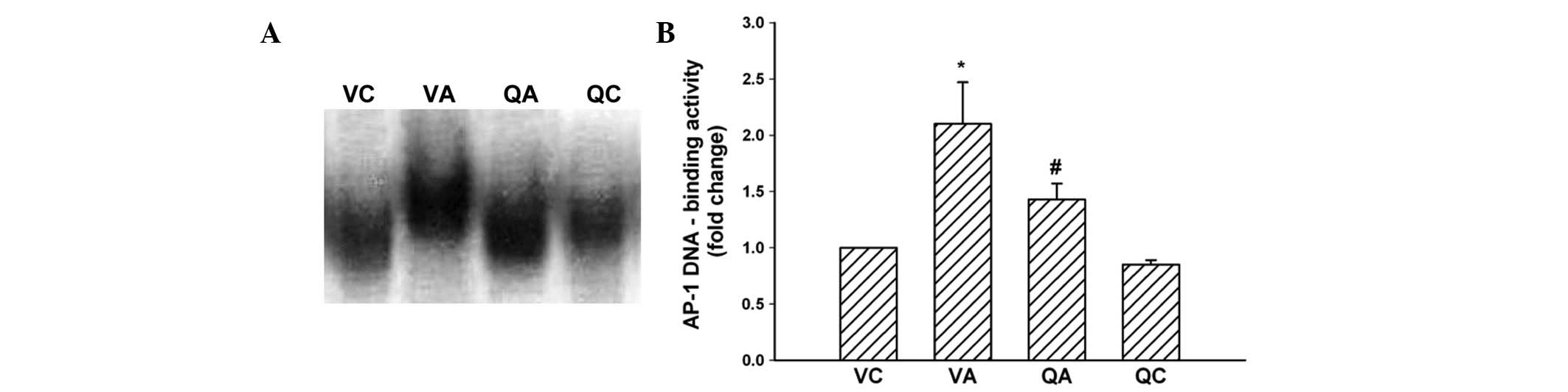
JNK/AP-1 signaling pathway and enzymatic
activities of MMPs
Since quercetin reduced oxidative stress, the
specific contribution of quercetin to the regulation of AP-1
activation in experimental AAA was examined. Levels of AP-1 DNA
binding activity in QA mice, determined by EMSA, were significantly
inhibited by quercetin when compared with controls in the VA group
(Fig. 4A and B). Western blotting
showed that phosphorylated-JNK was significantly upregulated in VA
mice and downregulated following treatment with quercetin. In
addition, QA animals had significantly less total JNK than VA
controls (Fig. 3A and B).
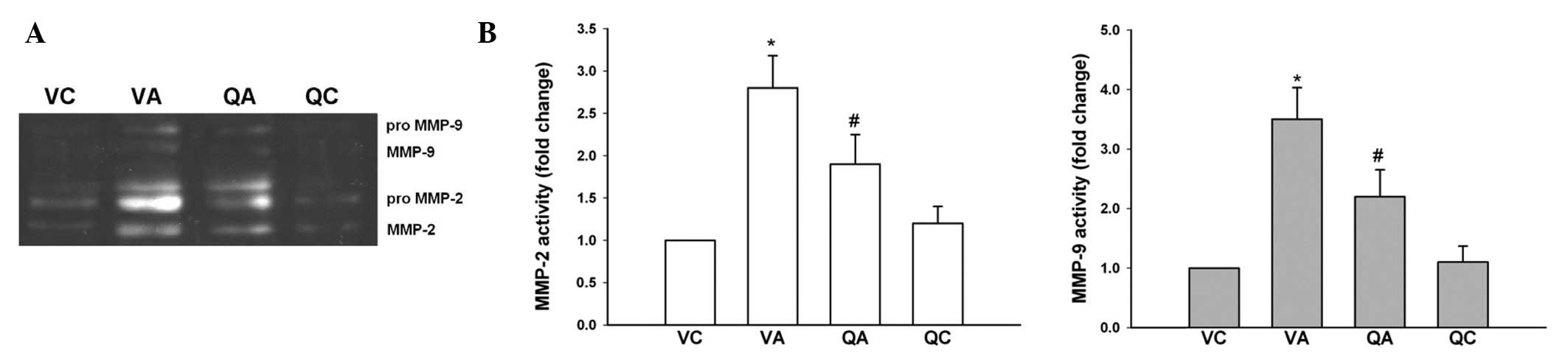
Gelatin zymography revealed that MMP-2 and -9
activities were elevated in VA mice, however, quercetin treatment
resulted in a marked decrease in MMP-2 and -9 activities (Fig. 5).
Discussion
The CaCl2-induced AAA model has been
widely employed to gain further understanding of the mechanisms
involved in aneurysm development, in order to identify potential
novel medical treatments (19). As
the results of this study showed, the development of
CaCl2-induced AAA in mice was accompanied by elevated
aortic ROS levels, increased nitrotyrosine formation and lipid
peroxidation products, indicating an enhancement in overall
oxidative stress. Previous studies have demonstrated that markers
of oxidative damage are present in human (20) and animal (21) aneurysmal lesions. However, these
oxidative stress markers were significantly inhibited by
supplementation of quercetin, a dietary antioxidant with a
polyphenolic structure. The antioxidant activity of polyphenols has
attracted much attention in relation to their possible role in the
prevention of chronic diseases (22). In particular, it was previously
reported that resveratrol, another polyphenolic compound,
counteracts systemic (23) and
local (24) oxidative stress and
limits experimental AAA progression. Moreover, a variety of
medications and interventions have been proven to successfully
suppress experimental aneurysm formation through a ROS-based
mechanism (6–8). Thus, it was hypothesized that the
aneurysm-inhibitory effect of quercetin in the present study may,
in part, associate with its lower oxidation-reduction
potential.
Oxidative stress is the result of a redox imbalance
between the generation of ROS and the secondary response from the
endogenous antioxidant network. Results from the present study
indicate a local upregulation of the endogenous antioxidant system,
including Mn-SOD and GPxs during CaCl2-induced AAA
formation. Mn-SOD and GPxs are key scavengers of ROS, for example,
H2O2 and lipid hydroperoxides (25,26).
Therefore, increases in Mn-SOD and GPxs may be a compensatory
response for an increase in ROS in the mouse aorta following
exposure to CaCl2. Quercetin, by restraining ROS levels,
prevents the elevation of those antioxidant enzymes, coinciding
with other study results (27–29),
which have reported the protective effect of quercetin on organ
injury.
The enhanced expression of NADPH oxidase, an enzyme
that catalyzes the production of O2•− from
oxygen and NADPH, is a major pathway of ROS formation in the
vascular wall (3). Inhibition of
ROS production by oral administration of apocynin, a specific
inhibitor of NADPH oxidases, attenuates AAA formation in a murine
model (8). It was also reported
that quercetin prevented the increase in aortic
O2•− production through downregulation of
p47phox expression in vivo and in vitro (14,15).
Furthermore, Thomas et al (30) have shown that p47phox deficiency
reduced oxidative stress and markedly attenuated AAA formation. The
present study found that quercetin treatment significantly
eliminated gene and protein expression of p47phox NADPH oxidase and
these data, together, demonstrate that quercetin is able to reduce
ROS formation via modulation of the p47phox subunit during AAA
development.
It is noteworthy that, in a previous study, iNOS
deficient mice were partly resistant to aneurysm induction by
CaCl2 (8). Nitric oxide
synthase (NOS) is also a source of ROS and increased cellular
expression of iNOS is specifically associated with large quantities
of nitric oxide produced during chronic inflammation (31). The present study found that
expression of iNOS in the aortic wall was inhibited following
quercetin treatment. It has been reported that under
inflammation-mimicking conditions, quercetin may inhibits iNOS
expression in cultured monocytes (32,33).
This result indicates another mechanism through which quercetin may
impact ROS generation.
More importantly, oxidative stress has been reported
to activate MMPs (34,35), a family of enzymes with the
capacity to cleave several components of the extracellular matrix,
including elastin and collagen. It is generally hypothesized that
MMPs are putative therapeutic targets in the prevention of AAA
(1). Our study group has
previously reported that treatment of mice with quercetin prevents
aortic wall destruction in the CaCl2-induced AAA model,
which is associated with a reduction in the expression of MMP-2 and
-9 (16). In the current study,
similar results were observed when determining the enzymatic
activities of MMPs in vitro by gelatin zymography. MMPs are
primarily regulated at the gene transcriptional level by various
factors, including cytokines, growth factors, ROS and reactive
nitrogen species (RNS). AP-1, a major downstream target of JNK, is
an essential transcription factor for MMP expression (36–38).
The present study shows that quercetin treatment significantly
inhibited AP-1 activation, accompanied by decreased phosphorylation
of JNK in AAA tissues. JNK, also known as stress-activated protein
kinase, is hypothesized to be involved in a number of cellular
stress responses. It is well established that ROS produced from
NADPH oxidase and RNS are potent inducers of JNK (39). Moreover, the existing evidence
indicates that JNK has an important role in AAA. Yoshimura et
al (40) have demonstrated
that pharmacological inhibition of JNK reduces MMP levels and
prevents the development of AAA. Furthermore, JNK inhibition caused
regression of established aneurysm in CaCl2- and
angiotensin II-infusion-induced AAA models. Activation of JNK leads
to modulation of other kinases, their nuclear translocation and
subsequent phosphorylation of a number of transcription factors,
including AP-1 (41). Thus, data
from the present study indicated that quercetin reduces oxidative
stress and blocks aneurysm formation, which may occur via the
mediation of the JNK/AP-1 pathway and MMP modulation.
In conclusion, the present study demonstrated that
an antioxidative mechanism is involved in the preventive action of
quercetin on CaCl2-induced AAA. This is notable as AAA
is a chronic and serious condition for which no medical treatment
currently exists. In addition, the compound has been observed to be
effective in reducing the risk factors of cardiovascular disease
that often occur simultaneously with AAA (1,9).
Although it is unclear whether these experimental observations
extend to aneurysmal degeneration as it occurs in humans, it is
likely to be a point of interest to explore in future
investigations.
Acknowledgements
The present study was supported by grants from the
Innovation Projects in Medical Science and Technology of Nanjing
Military Region (grant nos. 08Z028 and 10MA090) and the National
Natural Science Foundation of China (grant no. 81172032).
References
|
1
|
Sakalihasan N, Limet R and Defawe OD:
Abdominal aortic aneurysm. Lancet. 365:1577–1589. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Lederle FA: The natural history of
abdominal aortic aneurysm. Acta Chir Belg. 109:7–12.
2009.PubMed/NCBI
|
|
3
|
Keaney JF: Oxidative stress and the
vascular wall. Circulation. 112:2585–2588. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Marumo T, Schini-Kerth VB, Fisslthaler B
and Busse R: Platelet-derived growth factor-stimulated superoxide
anion production modulates activation of transcription factor
NF-kappaB and expression of monocyte chemoattractant protein 1 in
human aortic smooth muscle cells. Circulation. 96:2361–2367. 1997.
View Article : Google Scholar
|
|
5
|
McCormick ML, Gavrila D and Weintraub NL:
Role of oxidative stress in the pathogenesis of abdominal aortic
aneurysms. Arterioscler Thromb Vasc Biol. 27:461–469. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Wang M, Lee E, Song W, et al: Microsomal
prostaglandin E synthase-1 deletion suppresses oxidative stress and
angiotensin II-induced abdominal aortic aneurysm formation.
Circulation. 117:1302–1309. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Satoh K, Nigro P, Matoba T, et al:
Cyclophilin A enhances vascular oxidative stress and the
development of angiotensin II-induced aortic aneurysms. Nat Med.
15:649–656. 2009. View
Article : Google Scholar : PubMed/NCBI
|
|
8
|
Xiong WF, Mactaggart J, Knispel R, et al:
Inhibition of reactive oxygen species attenuates aneurysm formation
in a murine model. Atherosclerosis. 202:128–134. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Perez-Vizcaino F and Duarte J: Flavonols
and cardiovascular disease. Mol Aspects Med. 31:478–494. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Hanasaki Y, Ogawa S and Fukui S: The
correlation between active oxygens scavenging and antioxidative
effects of flavonoids. Free Radic Biol Med. 16:845–850. 1994.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Loke WM, Proudfoot JM, McKinley AJ, et al:
Quercetin and its in vivo metabolites inhibit neutrophil-mediated
low-density lipoprotein oxidation. J Agric Food Chem. 56:3609–3615.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Loke WM, Proudfoot JM, Hodgson JM, et al:
Specific dietary polyphenols attenuate atherosclerosis in
apolipoprotein E knockout mice by alleviating inflammation and
endothelial dysfunction. Arterioscler Thromb Vasc Biol. 30:749–757.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Golledge J and Norman PE: Atherosclerosis
and abdominal aortic aneurysm: cause, response, or common risk
factors? Arterioscler Thromb Vasc Biol. 30:1075–1077. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Sanchez M, Galisteo M, Vera R, et al:
Quercetin downregulates NADPH oxidase, increases eNOS activity and
prevents endothelial dysfunction in spontaneously hypertensive
rats. J Hypertens. 24:75–84. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Romero M, Jiménez R, Sánchez M, et al:
Quercetin inhibits vascular superoxide production induced by
endothelin-1: Role of NADPH oxidase, uncoupled eNOS and PKC.
Atherosclerosis. 202:58–67. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Wang L, Wang B, Li H, et al: Quercetin, a
flavonoid with anti-inflammatory activity, suppresses the
development of abdominal aortic aneurysms in mice. Eur J Pharmacol.
690:133–141. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Zhao H, Kalivendi S, Zhang H, et al:
Superoxide reacts with hydroethidine but forms a fluorescent
product that is distinctly different from ethidium: potential
implications in intracellular fluorescence detection of superoxide.
Free Radic Biol Med. 34:1359–1368. 2003. View Article : Google Scholar
|
|
18
|
Pryor WA and Squadrito GL: The chemistry
of peroxynitrite: a product from the reaction of nitric oxide with
superoxide. Am J Physiol. 268:L699–L722. 1995.PubMed/NCBI
|
|
19
|
Wang Y, Krishna S and Golledge J: The
calcium chloride-induced rodent model of abdominal aortic aneurysm.
Atherosclerosis. 226:29–39. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Miller FJ Jr, Sharp WJ, Fang X, Oberley
LW, Oberley TD and Weintraub NL: Oxidative stress in human
abdominal aortic aneurysms: a potential mediator of aneurysmal
remodeling. Arterioscler Thromb Vasc Biol. 22:560–565. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Yajima N, Masuda M, Miyazaki M, Nakajima
N, Chien S and Shyy JYJ: Oxidative stress is involved in the
development of experimental abdominal aortic aneurysm: A study of
the transcription profile with complementary DNA microarray. J Vasc
Surg. 36:379–385. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Schini-Kerth VB, Etienne-Selloum N,
Chataigneau T and Auger C: Vascular protection by natural
product-derived polyphenols: in vitro and in vivo evidence. Planta
Med. 77:1161–1167. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Palmieri D, Pane B, Barisione C, et al:
Resveratrol counteracts systemic and local inflammation involved in
early abdominal aortic aneurysm development. J Surg Res.
171:e237–e246. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Kaneko H, Anzai T, Morisawa M, et al:
Resveratrol prevents the development of abdominal aortic aneurysm
through attenuation of inflammation, oxidative stress, and
neovascularization. Atherosclerosis. 217:350–357. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Griendling KK and FitzGerald GA: Oxidative
stress and cardiovascular injury: Part I: basic mechanisms and
in vivo monitoring of ROS. Circulation. 108:1912–1916. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Miyamoto Y, Koh YH, Park YS, et al:
Oxidative stress caused by inactivation of glutathione peroxidase
and adaptive responses. Biol Chem. 384:567–574. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Dias AS, Porawski M, Alonso M, Marroni N,
Collado PS and González-Gallego J: Quercetin decreases oxidative
stress, NF-kappaB activation, and iNOS overexpression in liver of
streptozotocin-induced diabetic rats. J Nutr. 135:2299–2304.
2005.PubMed/NCBI
|
|
28
|
Mahesh T and Menon VP: Quercetin
allievates oxidative stress in streptozotocin-induced diabetic
rats. Phytother Res. 18:123–127. 2004. View
Article : Google Scholar : PubMed/NCBI
|
|
29
|
Moreira AJ, Fraga C, Alonso M, et al:
Quercetin prevents oxidative stress and NF-kappa B activation in
gastric mucosa of portal hypertensive rats. Biochem Pharmacol.
68:1939–1946. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Thomas M, Gavrila D, McCormick ML, et al:
Deletion of p47(phox) attenuates angiotensin II-induced abdominal
aortic aneurysm formation in apolipoprotein E-deficient mice.
Circulation. 114:404–413. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Stuehr DJ: Mammalian nitric oxide
synthases. Biochim Biophys Acta. 1411:217–230. 1999. View Article : Google Scholar
|
|
32
|
Kim BH, Cho SM, Reddy AM, Kim YS, Min KR
and Kim Y: Down-regulatory effect of quercitrin gallate on nuclear
factor-kappaB-dependent inducible nitric oxide synthase expression
in lipopolysaccharide-stimulated macrophages RAW 264.7. Biochem
Pharmacol. 69:1577–1583. 2005. View Article : Google Scholar
|
|
33
|
Bhaskar S, Shalini V and Helen A:
Quercetin regulates oxidized LDL induced inflammatory changes in
human PBMCs by modulating the TLR-NF-kappaB signaling pathway.
Immunobiology. 216:367–373. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Castier Y, Brandes RP, Leseche G, Tedgui A
and Lehoux S: p47phox-Dependent NADPH oxidase regulates
flow-induced vascular remodeling. Circ Res. 97:533–540. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Ejiri J, Inoue N, Tsukube T, et al:
Oxidative stress in the pathogenesis of thoracic aortic aneurysm.
Cardiovasc Res. 59:988–996. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Liu X, Manzano G, Lovett DH and Kim HT:
Role of AP-1 and RE-1 binding sites in matrix metalloproteinase-2
transcriptional regulation in skeletal muscle atrophy. Biochem
Biophys Res Commun. 396:219–223. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Kandasamy AD, Chow AK, Ali MAM and Schulz
R: Matrix metalloproteinase-2 and myocardial oxidative stress
injury: beyond the matrix. Cardiovasc Res. 85:413–423. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Kar S, Subbaram S, Carrico PM and Melendez
JA: Redox-control of matrix metalloproteinase-1: A critical link
between free radicals, matrix remodeling and degenerative disease.
Respir Physiol Neurobiol. 174:299–306. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Shen HM and Liu ZG: JNK signaling pathway
is a key modulator in cell death mediated by reactive oxygen and
nitrogen species. Free Radic Biol Med. 40:928–939. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Yoshimura K, Aoki H, Ikeda Y, et al:
Regression of abdominal aortic aneurysm by inhibition of c-Jun
N-terminal kinase. Nat Med. 11:1330–1338. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Manning AM and Davis RJ: Targeting JNK for
therapeutic benefit: from junk to gold? Nat Rev Drug Discov.
2:554–565. 2003. View Article : Google Scholar : PubMed/NCBI
|