Introduction
Retinoblastoma is the most common intraocular
malignancy in children. The incidence of retinoblastoma is constant
at 1/15,000–1/20,000 live births worldwide, which corresponds to
~9,000 new cases every year (1).
If diagnosed early and treated appropriately, children with
retinoblastoma are able to be cured. However, the prognosis remains
poor in developing countries (2),
where mortality due to retinoblastoma is common.
It is well accepted that retinoblastoma is initiated
by the mutation of the tumor suppressor gene retinoblastoma 1
(RB1) (3). Numerous studies
have suggested that Knudson’s M1 and M2 mutational events are
necessary for initiating the oncogenesis of retinoblastoma;
however, other tumor suppressor genes or oncogenes may also
participate in the aggressive progression of this tumor (4). The initiating events are only
sufficient for driving the formation of the benign retina tumor, or
retinoma; however, they are not enough to lead to the development
of malignancy (5). Thus,
additional mutational events or other biological behaviors are
required for the development from retinoma into retinoblastoma. In
addition to its cell biology, there are also numerous unclear
aspects associated with the presence of chemoresistance that must
be addressed (6). Therefore, the
development of novel strategies for the treatment of retinoblastoma
is highly desirable for improving the survival outcome of this
disease.
Notch signaling is a highly conserved pathway, which
mediates signaling between adjacent cells directly in metazoans
(7). In mammals, five canonical
Notch ligands (Jagged1 and 2 and Delta-like 1, 3 and 4) and four
Notch receptors (Notch1–4) have been described to date. Notch
receptors are activated by binding to their transmembrane ligands
on nearby cells. This interaction further triggers sequential
receptor cleavages within the transmembrane domain and the release
of the Notch intracellular domain (NICD). Following moving into the
nucleus, NICD binds to the transcriptional factor CBF/Su(H)/LAG-2
(CSL; also known as RBP-J in rodents), which is normally suppressed
by a corepressor complex. Corepressors are replaced by a
coactivating complex containing Mastermind-like protein when NICD
associates with CSL. Downstream genes, including Hes-1 and Hey-1,
are activated subsequently.
Aberrant expression and dysfunction of the Notch
pathway is frequently observed in various types of cancer (8,9). In
tumorigenesis, the Notch pathway is a double-edged sword (10). It can be an adjudicator of survival
versus death, proliferation versus growth arrest or differentiation
versus ‘stemness’. Despite the progress in delineating the
expression profiles and functional derangements of Notch signaling
pathways in carcinogenesis during the last decade, its biological
significance in retinoblastoma has received little attention, if
any.
The present study first examined the expression
profiles of Notch ligands and receptors in the human retinoblastoma
cell line SO-Rb50, and assessed the effect of
N-[N-(3,5-difluorophenacetyl)-l-alanyl]-S-phenylglycine
t-butyl ester (DAPT), a Notch signaling inhibitor, on the
proliferation of these cells, as well as its underlying
mechanism.
Materials and methods
Cell culture and treatment
Human retinoblastoma cells (SO-Rb50 cells) were
obtained from the Department of Pathology of Zhongshan Ophthalmic
Center, Sun Yat-sen University (Guangzhou, Guangdong, China). The
cells were cultured in RPMI-1640 medium (Gibco-BRL, Grand Island,
NY, USA) supplemented with 10% fetal bovine serum (Gibco-BRL) at
37°C in a humidified atmosphere of 95% air and 5% CO2.
DAPT (Sigma, St. Louis, MO, USA) was dissolved in dimethyl
sulfoxide (DMSO; Sigma) to yield a final concentration equal to 10
mmol/l. Cells were then treated with various concentrations of DAPT
(5, 10, 30 and 60 μmol/l) or DMSO of the same volume (control).
Cell proliferation analysis
The standard colorimetric Cell Counting kit-8
(CCK-8; Dojindo Laboratories, Kumamoto, Japan) was used for the
determination of the number of viable cells in cell proliferation
assays. SO-Rb50 cells were seeded with a volume of 200 μl cell
suspension (10,000 cells/well) into 96-well plates. Following
treatment with DAPT, the cells were incubated for 24 or 48 h.
Following that, 20 μl CCK-8 was added to each well and incubated
for 2 h at 37°C. Optical densities were measured using a microplate
reader (Elx800; Bio-Tek Inc., North Brunswick, NJ, USA) by scanning
at 450 nm. Cell survival rates were measured at two time-points of
the cell growth curve.
Western blot analysis
Cells were collected and lysed with cell lysis
buffer. The cell lysates were collected following centrifugation.
An equal amount of protein was loaded and separated on 10%
SDS-PAGE. Following being transferred onto polyvinylidene
difluoride membranes (Millipore, Bedford, MA, USA), the membranes
were inhibited in 5% skimmed milk for 1 h and then incubated with
primary antibodies at 4°C overnight. Following washing three times
with phosphate-buffered saline containing 0.1% Tween-20, the
membranes were incubated with horseradish peroxidase
(HRP)-conjugated secondary antibodies for 1 h at room temperature.
The membranes were incubated with SignalFire™ Elite ECL Reagent
(Cell Signaling Technology, Beverly, MA, USA) and exposed to X-ray
film. β-actin was used as the loading control. Primary antibodies,
including rabbit mAb anti-Jagged1, rabbit mAb anti-Jagged2, rabbit
mAb anti-Notch1, rabbit mAb anti-Notch2, rabbit mAb anti-Notch3,
rabbit mAb anti-PI3K, rabbit mAb anti-Akt, rabbit mAb anti-p-Akt,
rabbit mAb anti-p38MAPK, rabbit mAb anti-p-Src, rabbit mAb
anti-p-Erk1/2, rabbit mAb anti-β-catenin, mouse mAb anti-β-actin,
mouse mAb anti-Erk1/2, mouse mAb anti-p-p38MAPK and mouse mAb
anti-Src, together with horse anti-mouse and goat anti-rabbit
HRP-conjugated secondary antibodies were purchased from Cell
Signaling Technology.
Statistical analysis
Each experiment was repeated at least three times.
The results are expressed as the mean ± standard error. Student’s
t-test was performed for the estimation of statistical
significance. P<0.05 was considered to indicate a statistically
significant difference.
Results
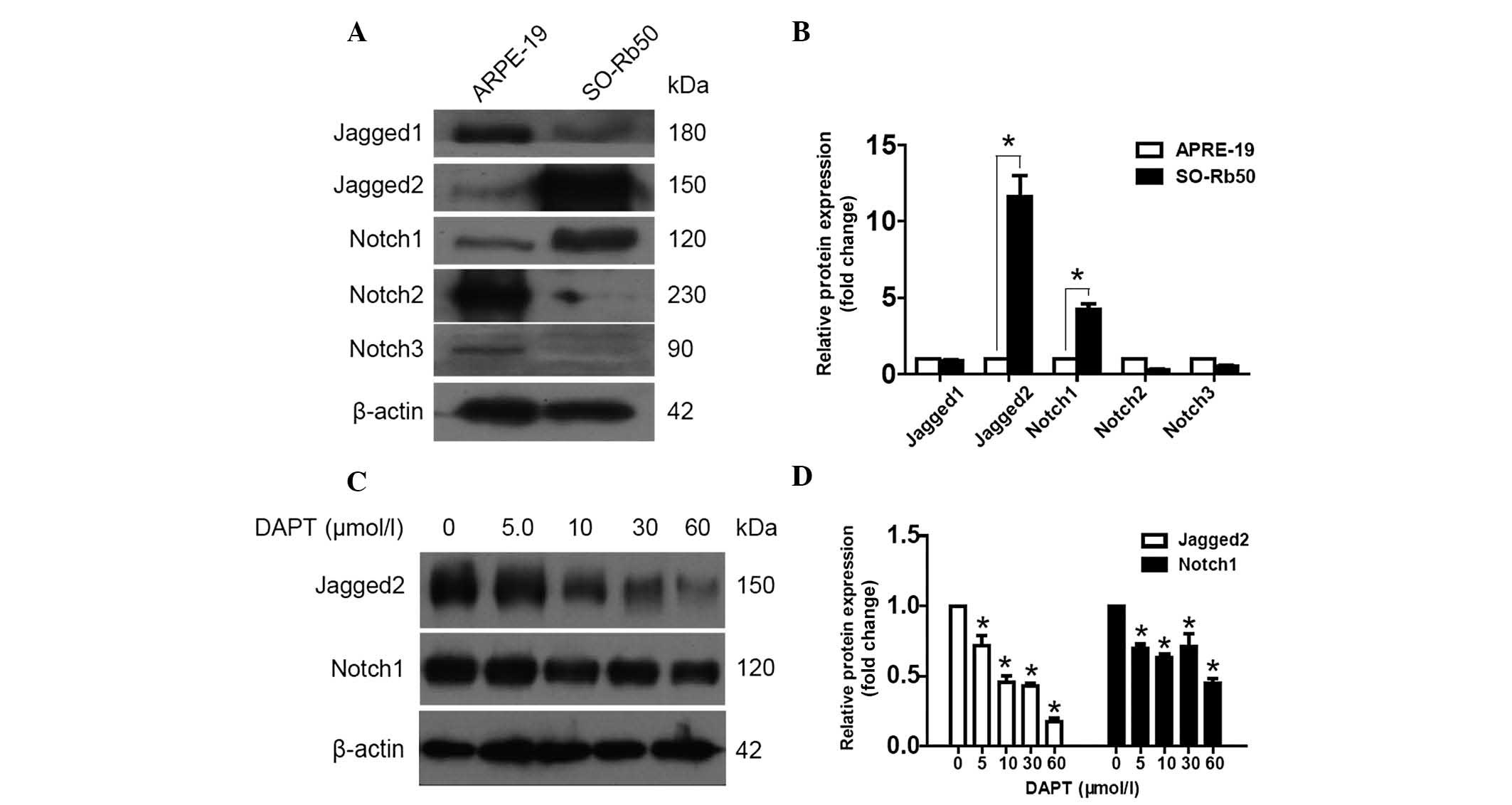
Notch1 and Jagged2 are highly expressed
in SO-Rb50 retinoblastoma cells
In order to understand the role of Notch signaling
in retinoblastoma, the present study initially sought to identify
which Notch receptors and ligands were primarily expressed in
SO-Rb50 cells. Western blot analysis was performed to measure the
protein expression levels of Notch1, Notch2, Notch3, Jagged1 and
Jagged2. Their expression in a human retinal pigment epithelial
cell line, ARPE-19, was set as the control. A significantly higher
expression of Jagged2 (4.23±0.35 fold change; P<0.05) and Notch1
(11.6±1.40 fold change; P<0.05) was detected in SO-Rb50 cells
(Fig. 1A and B). The expression
levels of Jagged1 in these two types of cell lines were comparable.
However, ARPE-19 exhibited much higher expression levels of Notch2
and Notch3, while they were minimal in SO-Rb50 cells. These results
suggest that Notch signaling is important in oncogenesis in
retinoblastoma.
Inhibition of the Notch pathway by the
γ-secretase inhibitor DAPT is able to downregulate the expression
of Jagged2 and Notch1
To determine the effects of a γ-secretase inhibitor
on the Notch signaling pathway, the protein expression levels of
Notch1 and Jagged2 in DAPT-treated SO-Rb50 cells were examined by
western blot analysis. The expression of Jagged2 and Notch1 were
downregulated following treatment with DAPT for 24 h. This
alteration was more profound for Jagged2 levels, which decreased
with increasing DAPT concentration in a dose-dependent manner
(P<0.05; Fig. 1C and D).
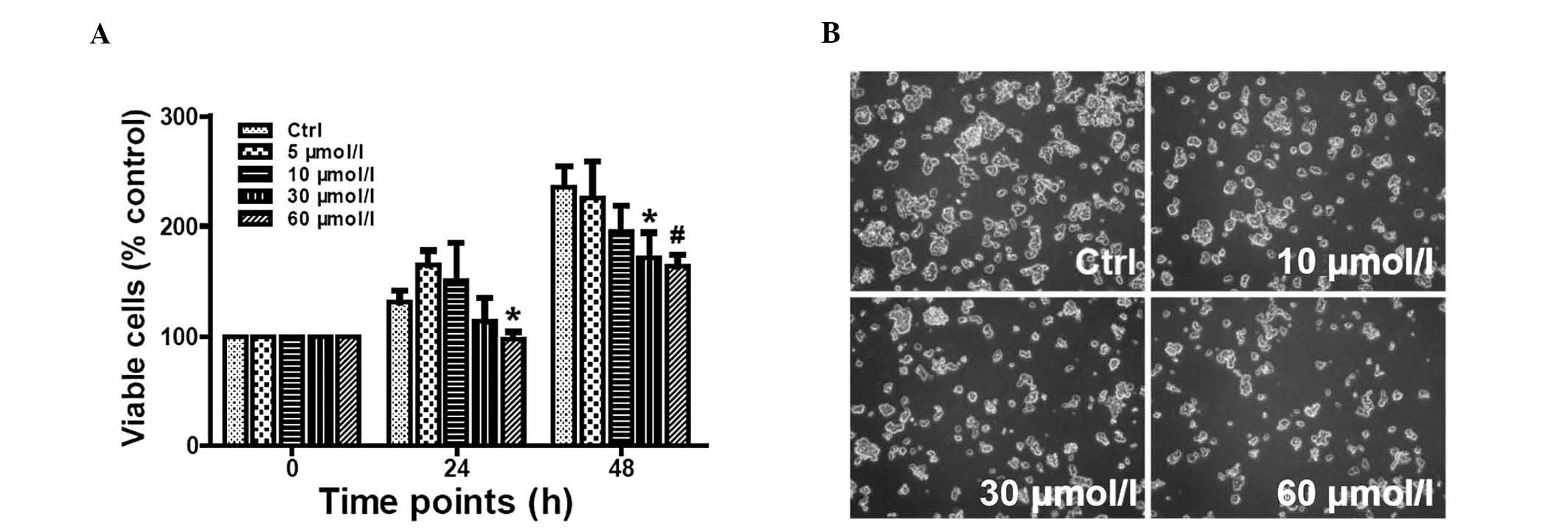
Inhibition of the Notch pathway inhibited
SO-Rb50 cell growth
To determine whether DAPT is able to inhibit SO-Rb50
cell proliferation, its effect on cell growth was examined by the
CCK-8 assay. Cells were treated with various concentrations of DAPT
(0, 5, 10, 30 or 60 μmol/l). The results demonstrated that DAPT
dose-dependently inhibited SO-Rb50 cell growth. The inhibitory
effect was significant when SO-Rb50 cells were treated with 60
μmol/l DAPT for 24 h or with 30 or 60 μmol/l DAPT for 48 h
(*P<0.05, #P<0.01; Fig. 2A). Nevertheless, the suppression
effect was not significantly different between these two doses at
either 24 or 48 h (P>0.05). Under the phase-contrast microscope,
the cell density was found to be markedly reduced when treated with
30 or 60 μmol/l DAPT for 24 h (Fig.
2B). These results demonstrated that the inhibition of the
Notch pathway with DAPT was able to inhibit SO-Rb50 cell
proliferation.
DAPT modulates cell growth through the
regulation of the PI3K/Akt, Src, p38MAPK and wnt/β-catenin
pathways; however, it has no effect on the Erk1/2 pathway
The Notch signaling pathway has been demonstrated to
crosstalk with the epidermal growth factor receptor pathway
(11). Therefore, several key
signaling pathways associated with cell growth, including PI3K/Akt,
Src, p38MAPK, wnt/β-catenin and Erk, were assessed by western blot
analysis following 24 h treatment with various concentrations of
DAPT. A significant inhibitory effect on the expression of PI3K and
β-catenin was observed, as well as on the phosphorylation of Akt,
Src and p38MAPK, following treatment with 30 or 60 μmol/l DAPT
(Fig. 3A–H). Although several
studies have demonstrated the crosstalk between the Notch pathway
and the Erk1/2 pathway (12), no
effect on p-Erk1/2 levels was observed in any of the treatments
(P>0.05; Fig. 3I and J).
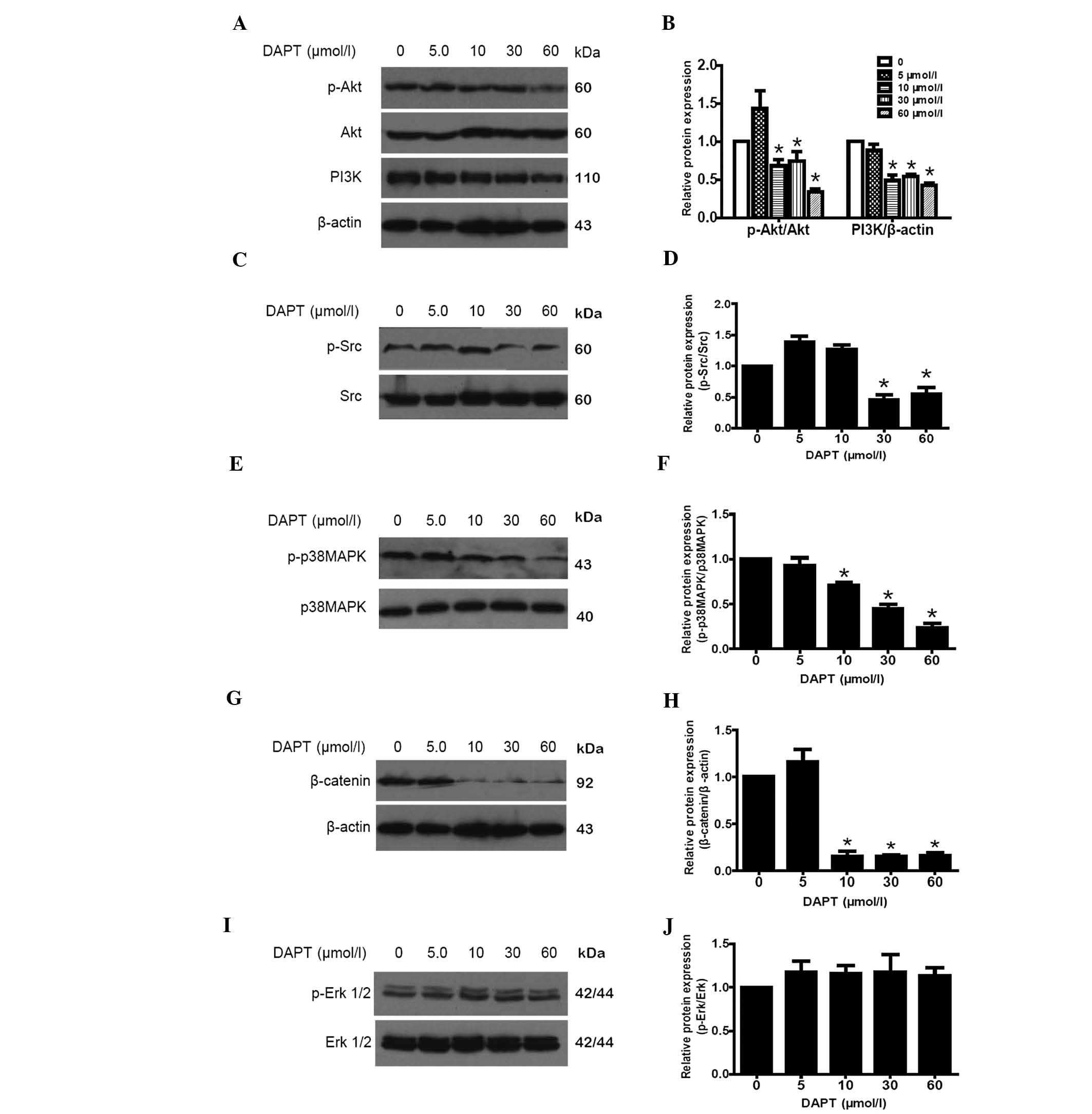 | Figure 3DAPT inhibits SO-Rb50 cell growth via
co-operating with multiple pathways. Western blot analysis was
performed to detect the activation of several classic pathways,
including the (A and B) PI3K/Akt, (C and D) Src, (E and F) p38MAPK,
(G and H) wnt/β-catenin and (I and J) Erk1/2 pathways. Total
protein expression levels of Akt, p38MAPK, Src and Erk1/2 were used
as the internal control for their phosphorylated isoforms,
respectively, and β-actin was set as the loading control for PI3K
and β-catenin. Expression of PI3K and β-catenin, as well as
phosphorylation of Akt, p38MAPK and Src, were significantly abated
when treated with 30 or 60 μmol/l DAPT for 24 h. (I and J) Of note,
the total or phosphorylated Erk1/2 were not changed. DAPT, DAPT,
N-[N-(3,5-difluorophenacetyl)-l-alanyl]-S-phenylglycine
t-butyl ester; MAPK, mitogen activated protein kinase; PI3K,
phosphoinositide 3-kinase; Erk, extracellular signal-regulated
kinase. *P<0.05. |
Discussion
Among various molecular targets, Notch signaling is
attractive as it is involved in numerous cellular processes,
including cell fate specification, differentiation, proliferation
and survival. Previous studies have demonstrated that Notch
signaling is involved in the carcinogenesis, progression, invasion
and neurovascular formation of numerous malignant tumors. Notch is
able to function as an oncogene or a tumor suppressor, depending on
the cell type and context. It acts as a tumor suppressor in skin
tumors and non-small cell lung cancer (13,14).
However, it acts as an oncogene in renal, pancreatic, breast and
prostate cancer (15). In the
present study, Notch1 and Jagged2 were found to be highly expressed
in the human retinoblastoma cell line. Inhibition of the
Jagged2/Notch1 signaling pathway with DAPT suppressed cell growth
via multiple crosstalk with other pathways. Therefore, the present
study provided evidence supporting the role of Jagged2/Notch1 as
oncogenes in human retinoblastoma cells.
Previous studies have demonstrated that Notch is
aberrantly expressed in various types of human cancers, including
breast, colon, cervical, lung, head and neck, renal cell carcinoma
and pancreatic cancer (16).
Typically, Notch1 is overexpressed in prostate cancer cell lines
and human prostate cancer tissues (17). Furthermore, Notch1 expression
levels in human prostate cancer tissues are elevated with
increasing tumor grade (17).
However, the role of the Notch pathway in retinoblastoma remains to
be elucidated. Therefore, in the present study, the expression
profiles of Notch ligands (Jagged1 and 2) and receptors (Notch1–3)
were investigated for the first time, to the best of our knowledge.
Notch1 and Jagged2 were identified to be highly expressed in the
human retinoblastoma cell line, which is similar to findings in
myeloma cells, tongue carcinoma tissues and lung adenocarcinoma
(18–20). These results further demonstrate
that the expression of Notch signaling pathway proteins is
frequently altered in malignancies.
Previous studies reported that components of the
Notch signaling pathway are also the downstream targets of Notch
signaling itself (21). In the
present study, the expression levels of Jagged2 and Notch1 were
consistently and significantly suppressed when the cells were
treated with various concentrations of DAPT. This suppressive
effect was more profound on the ligand Jagged2 and was
dose-dependent. The effect of DAPT on the gene expression levels of
Jagged1, Notch2 and Notch3 were not examined, as their expression
was relatively low in SO-Rb50 cells.
Several studies have demonstrated that the
downregulation of the Notch signaling pathway elicited a marked
effect on the cell growth inhibition of cancer cells (22,23).
In the present study, when the Notch signaling pathway was
inhibited with DAPT, SO-Rb50 cells underwent cell cycle arrest,
which was in agreement with previous studies. The present study
demonstrated that DAPT inhibited SO-Rb50 cell proliferation in a
dose-dependent manner.
It is well accepted that numerous signaling
pathways, including PI3K/Akt, Erk, p38MAPK, Src and wnt/β-catenin
signaling, are involved in tumor cell growth, invasion and
metastasis. Since DAPT inhibited the proliferation of SO-Rb50
cells, the present study examined the effects of DAPT on the
PI3K/Akt, Erk, p38MAPK, Src and wnt/β-catenin signaling
pathways.
The protein kinase Akt, which is able to induce
γ-secretase, has been implicated in transducing Notch signaling
(24). Notch1 activation enhanced
melanoma cell survival and this effect was mediated by activation
of the Akt pathway (25).
Activating mutation of Notch1 comprises the most prominent genetic
abnormality in T-cell acute lymphoblastic leukemia (T-ALL) in
children. However, the majority of T-ALL cell lines with Notch1
mutations demonstrate chemoresistance to γ-secretase inhibitors
(26). Previous studies revealed
that PI3K/Akt and PTEN/AKT/mTOR pathways are involved in
tumorigenesis (27). The present
study examined whether the growth inhibition of SO-Rb50 cells was
indeed due to the regulation of the Akt pathway. It was
consistently found that the downregulation of the Notch signaling
pathway by DAPT decreased Akt phosphorylation, suggesting that Akt
inactivation may contribute to DAPT-induced cell growth arrest.
Src family kinases are non-receptor kinases
overexpressed in the majority of pancreatic cancers and involved in
cancer progression and metastasis. The expression of Src kinase is
significantly higher in various types of invasive tumors (28,29).
In pancreatic cancer cells, Notch1 and c-Src proteins are
physically associated, and this association mediates Notch1
processing and activation (30).
Poorly differentiated tumors demonstrated a higher expression of
Src kinase when compared with moderately- and well-differentiated
tumors (31). Although Src kinase
is moderately expressed in normal retinal tissues, Src kinase and
phosphorylated signal transducer and activator of transcription 3
proteins in invasive retinoblastoma tumors are highly expressed,
suggesting that they may be important in invasiveness by
contributing to multiple aspects of tumor progression. In the
present study, a high expression of p-Src and Src in SO-Rb50 cells
was detected, whereas DAPT inhibited the phosphorylation of Src
without affecting the total Src expression. These results indicated
that Src kinases are inactivated by DAPT.
The Wnt signaling pathway, which is involved in stem
cell self-renewal and differentiation in various types of cancer,
is a promising candidate pathway for cancer therapy (32). In chick and monkey retina,
canonical Wnt signaling regulates the proliferation of retinal
stem/progenitor cells, maintains them in an uncommitted state and
suppresses neuronal differentiation. Although previous studies
demonstrated that Wnt signaling activation significantly decreased
the viability of retinoblastoma cells by inducing cell cycle arrest
associated with the upregulation of p53 (33), sorted adenosine
triphosphate-binding cassette sub-family G member 2
(ABCG2)-positive stem-like cells exhibited higher levels of
β-catenin than ABCG2-negative non-stem cells. This suggests that
elevated canonical Wnt signaling regulates cancer stem-like cell
populations in retinoblastoma. Therefore, inhibiting the canonical
Wnt pathway may potentially suppress stem-like cell formation in
retinoblastoma. β-catenin, which is involved in numerous
protein-protein interactions (34), is considered the central hub of the
Wnt signaling pathway. The present study found that DAPT also
inhibited the expression of β-catenin in SO-Rb50 cells, which
indicates an interaction between the Wnt and Notch signaling
pathways.
Crosstalk among the intracellular signaling pathways
is important in the determination of signaling specificity. In
myogenesis, Notch was reported to specifically indirectly
inactivate p38MAPK by inducing the expression of MKP-1, a member of
the dual-specificity MAPK phosphatase (35). However, crosstalk between the Notch
and the MAPK pathways in cancer has remained to be fully
elucidated. In human nasopharyngeal carcinoma cells, a γ-secretase
inhibitor suppressed human cell proliferation via inactivating
Erk1/2 signaling pathways, without affecting p38MAPK signaling
(22). In melanoma, Notch1
signaling promoted primary melanoma progression, partially mediated
through the activation of the MAPK pathway (25). Notably, in the present study,
examination of the Erk1/2 and p38MAPK levels in DAPT-treated
SO-Rb50 cells revealed that p38MAPK decreased in a dose-dependent
manner in response to DAPT, whereas the expression of p-Erk1/2 was
not affected. These data indicated that DAPT treatment resulted in
the inhibition of the MAPK signaling pathway with specificity on
p38MAPK signaling (however, not on Erk1/2 signaling). In addition,
the crosstalk between Notch and other oncogenic pathways was likely
to be context-dependent.
In conclusion, the present study suggested that
Jagged2/Notch1 signaling was involved in oncogenesis in
retinoblastoma. Inhibition of the Notch pathway with DAPT inhibited
the proliferation of human retinoblastoma cells via suppressing the
PI3K/Akt, Src, p38MAPK and Wnt/β-catenin signaling pathways.
Acknowledgements
The authors would like to thank the Department of
Pathology, Zhongshan Ophthalmic Center for providing the human
retinoblastoma cell line SO-Rb50.
References
|
1
|
Kivelä T: The epidemiological challenge of
the most frequent eye cancer: retinoblastoma, an issue of birth and
death. Br J Ophthalmol. 93:1129–1131. 2009.PubMed/NCBI
|
|
2
|
Rodriguez-Galindo C, Wilson MW, Chantada
G, et al: Retinoblastoma: one world, one vision. Pediatrics.
122:e763–e770. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Friend SH, Bernards R, Rogelj S, et al: A
human DNA segment with properties of the gene that predisposes to
retinoblastoma and osteosarcoma. Nature. 323:643–646. 1986.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Corson TW and Gallie BL: One hit, two
hits, three hits, more? Genomic changes in the development of
retinoblastoma. Genes Chromosomes Cancer. 46:617–634. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Dimaras H, Khetan V, Halliday W, et al:
Loss of RB1 induces non-proliferative retinoma: increasing genomic
instability correlates with progression to retinoblastoma. Hum Mol
Genet. 17:1363–1372. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Dyer MA and Bremner R: The search for the
retinoblastoma cell of origin. Nat Rev Cancer. 5:91–101. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Kidd S, Lockett TJ and Young MW: The Notch
locus of Drosophila melanogaster. Cell. 34:421–433. 1983.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Mullendore ME, Koorstra JB, Li YM, et al:
Ligand-dependent Notch signaling is involved in tumor initiation
and tumor maintenance in pancreatic cancer. Clin Cancer Res.
15:2291–2301. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Nam DH, Jeon HM, Kim S, et al: Activation
of notch signaling in a xenograft model of brain metastasis. Clin
Cancer Res. 14:4059–4066. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
South AP, Cho RJ and Aster JC: The
double-edged sword of Notch signaling in cancer. Semin Cell Dev
Biol. 23:458–464. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Aguirre A, Rubio ME and Gallo V: Notch and
EGFR pathway interaction regulates neural stem cell number and
self-renewal. Nature. 467:323–327. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Zayzafoon M, Abdulkadir SA and McDonald
JM: Notch signaling and ERK activation are important for the
osteomimetic properties of prostate cancer bone metastatic cell
lines. J Biol Chem. 279:3662–3670. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Ye YZ, Zhang ZH, Fan XY, et al: Notch3
overexpression associates with poor prognosis in human
non-small-cell lung cancer. Med Oncol. 30:5952013. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Reichrath J and Reichrath S:
Notch-signaling and nonmelanoma skin cancer: an ancient friend,
revisited. Adv Exp Med Biol. 727:265–271. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Purow B: Notch inhibition as a promising
new approach to cancer therapy. Adv Exp Med Biol. 727:305–319.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Miele L, Miao H and Nickoloff BJ: NOTCH
signaling as a novel cancer therapeutic target. Curr Cancer Drug
Targets. 6:313–323. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Bin Hafeez B, Adhami VM, Asim M, et al:
Targeted knockdown of Notch1 inhibits invasion of human prostate
cancer cells concomitant with inhibition of matrix
metalloproteinase-9 and urokinase plasminogen activator. Clin
Cancer Res. 15:452–459. 2009.
|
|
18
|
Chiron D, Maïga S, Descamps G, et al:
Critical role of the NOTCH ligand JAG2 in self-renewal of myeloma
cells. Blood Cells Mol Dis. 48:247–253. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Zhang TH, Liu HC, Zhu LJ, et al:
Activation of Notch signaling in human tongue carcinoma. J Oral
Pathol Med. 40:37–45. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Yang Y, Ahn YH, Gibbons DL, et al: The
Notch ligand Jagged2 promotes lung adenocarcinoma metastasis
through a miR-200-dependent pathway in mice. J Clin Invest.
121:1373–1385. 2011. View
Article : Google Scholar : PubMed/NCBI
|
|
21
|
Rodilla V, Villanueva A, Obrador-Hevia A,
et al: Jagged1 is the pathological link between Wnt and Notch
pathways in colorectal cancer. Proc Natl Acad Sci USA.
106:6315–6320. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Chen SM, Liu JP, Zhou JX, et al:
Suppression of the notch signaling pathway by gamma-secretase
inhibitor GSI inhibits human nasopharyngeal carcinoma cell
proliferation. Cancer Lett. 306:76–84. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Shao H, Huang Q and Liu ZJ: Targeting
Notch signaling for cancer therapeutic intervention. Adv Pharmacol.
65:191–234. 2012. View Article : Google Scholar
|
|
24
|
Guo D, Ye J, Dai J, et al: Notch-1
regulates Akt signaling pathway and the expression of cell cycle
regulatory proteins cyclin D1, CDK2 and p21 in T-ALL cell lines.
Leuk Res. 33:678–685. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Liu ZJ, Xiao M, Balint K, et al: Notch1
signaling promotes primary melanoma progression by activating
mitogen-activated protein kinase/phosphatidylinositol 3-kinase-Akt
pathways and up-regulating N-cadherin expression. Cancer Res.
66:4182–4190. 2006. View Article : Google Scholar
|
|
26
|
Mungamuri SK, Yang X, Thor AD and
Somasundaram K: Survival signaling by Notch1: mammalian target of
rapamycin (mTOR)-dependent inhibition of p53. Cancer Res.
66:4715–4724. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Guo D, Teng Q and Ji C: NOTCH and
phosphatidylinositide 3-kinase/phosphatase and tensin homolog
deleted on chromosome ten/AKT/mammalian target of rapamycin (mTOR)
signaling in T-cell development and T-cell acute lymphoblastic
leukemia. Leuk Lymphoma. 52:1200–1210. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Mohan A, Mallikarjuna K, Venkatesan N,
Abhyankar D, Parikh PM and Krishnakumar S: The study of c-Src
kinase and pStat3 protein expression in retinoblastoma. Exp Eye
Res. 83:736–740. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Ishizawar R and Parsons SJ: c-Src and
cooperating partners in human cancer. Cancer Cell. 6:209–214. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Ma YC, Shi C, Zhang YN, et al: The
tyrosine kinase c-Src directly mediates growth factor-induced
Notch-1 and Furin interaction and Notch-1 activation in pancreatic
cancer cells. PLoS One. 7:e334142012. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Chen YS, Wu MJ, Huang CY, Lin SC, Chuang
TH, Yu CC and Lo JF: CD133/Src axis mediates tumor initiating
property and epithelial-mesenchymal transition of head and neck
cancer. PLoS One. 6:e280532011. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Reya T and Clevers H: Wnt signalling in
stem cells and cancer. Nature. 434:843–850. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Tell S, Yi H, Jockovich ME, Murray TG and
Hackam AS: The Wnt signaling pathway has tumor suppressor
properties in retinoblastoma. Biochem Biophys Res Commun.
349:261–269. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Hahne G and Grossmann TN: Direct targeting
of beta-catenin: Inhibition of protein-protein interactions for the
inactivation of Wnt signaling. Bioorg Med Chem. 21:4020–4026. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Kondoh K, Sunadome K and Nishida E: Notch
signaling suppresses p38 MAPK activity via induction of MKP-1 in
myogenesis. J Biol Chem. 282:3058–3065. 2007. View Article : Google Scholar : PubMed/NCBI
|