Introduction
Alzheimer’s disease (AD) is an age-related
neurodegenerative disorder. This disease is pathologically
characterized by the deposition of extracellular amyloid-β (Aβ),
intracellular neurofibrillary tangles (NFTs) and degenerated
neurons. Evidence suggests that brain tissues are continually
exposed to oxidative stress during the development of AD (1). Oxidative stress is generally caused
by an imbalance between the production of reactive oxygen species
(ROS) and the antioxidative defense system, which is considered to
be responsible for the decreasing memory (2). Oxidative stress can provoke the
cellular adaptive response to injury induced by ROS, but this
response often requires the upregulation of endogenous antioxidant
enzymes (3). Heme oxygenases (HOs)
are considered to be important antioxidant enzymes, which catalyze
heme to biliverdin, carbon monoxide and ferrous iron (4). The two main isoforms of HO are HO-1
and HO-2. HO-1 expression is limited to a small number of neurons
and glial cells (5), and HO-2 is
constitutively expressed in the normal brain (6). It has previously been shown that the
expression of HO-1 is increased, while that of HO-2 is reduced, in
the brains of patients with AD (7). In addition, it has been shown that
the sustained upregulation of HO-1 may contribute to the
pathological iron deposition, oxidative damage and mitochondrial
dysfunction in AD (8).
Previous studies have shown that is an association
between HO expression and AD (9,10),
but whether HO-1 and HO-2 have different roles in the early stages
of AD has not yet been studied. In the present study, using the
APPswe/PS1ΔE9 (APP/PS1) transgenic mouse
model, HO-1 and HO-2 expression was monitored with
immunofluorescence and western blotting methods. Furthermore, the
differential expression patterns of HO-1 and HO-2 in neuronal and
glial cells were observed using immunofluorescence methods, and the
plasma concentrations of HO-1 and HO-2 were determined by
ELISA.
Materials and methods
Experimental animals
Six-month-old male APP/PS1 transgenic and wild-type
mice were used in this study, with 12 mice per group. The APP/PS1
mouse strain is a double-transgenic hemizygote that expresses a
chimeric mouse/human amyloid precursor protein and mutant human
presenilin-1. APP/PS1 transgenic mice were purchased from the Model
Animal Research Center of Nanjing University (Nanjing, China).
Wild-type mice were purchased from the Experimental Animal Center
of Shanghai Academy (Shanghai, China). All procedures were
performed in accordance with the Guide for the Care and Use of
Medical Laboratory Animals (Ministry of Health, China, 1998) and
the guidelines of the Shanghai University of Traditional Chinese
Medicine (Shanghai, China) Laboratory Animal Care and Use
Committee.
Morris water maze test
Since the Morris water maze is a test evaluating
spatial learning (11), the test
was selected in the present study to assess hippocampus-dependent
spatial learning and memory. The water maze was divided into four
equal quadrants and a hidden platform was submerged 1 cm beneath
the water surface. The water temperature was kept between 20 and
23°C. Each mouse was tested in four trials per day with an
inter-trial interval of 30–40 min, which continued for 5 days. In
each trial, the mouse was released facing the wall of the pool from
one of four starting points (north, east, south or west). The mouse
was allowed to search for the platform for ≤70 sec, allowing it to
rest 30 sec on the platform. The time the mice spent finding the
platform was recorded as the escape latency. The experiments were
recorded with a camera connected to a video recorder and a
computerized tracking system.
Measurement of ROS levels
Intracellular ROS levels of the hippocampus were
measured as previously described (12). Briefly, hippocampi were homogenized
in 0.01 M phosphate-buffered saline (PBS) (pH 7.2–7.4). The
homogenized cells (0.4 mg/ml) were loaded with the cell permeant
probe 2,7-dichlorofluorescein diacetate (DCFH-DA, 20 μM) for 60 min
at 37°C in the dark, then the fluorescence was measured through a
spectrofluorometer (Synergy HT, BioTek Instruments, Winooski, VT,
USA) using 485 nm as the excitation and 525 nm as the emission
wavelength. The normalized data were expressed as a value of
100%.
Immunofluorescence analysis
Mice were anesthetized with 5% chloral hydrate and
perfused through the heart with saline solution followed by 4%
paraformaldehyde. The brains were then removed, post-fixed in 4%
paraformaldehyde for 24 h and immersed in 30% sucrose until they
sank. Thereafter, coronal sections (30 μm) were cut using a
freezing microtome (Microm™ HM 525 Cryostat, Thermo Scientific,
Walldorf, Germany). Sections were permeabilized with 0.2% Triton
X-100, then blocked in 5% bovine serum albumin for 30 min. The
sections were subsequently incubated with primary antibodies,
including polyclonal anti-Aβ1-42 (1:200; Abcam,
Cambridge, MA, USA), monoclonal anti-HO-1 (1:200; Abcam),
monoclonal anti-HO-2 (1:100; Santa Cruz Biotechnology, Inc.,
Dallas, TX, USA), monoclonal anti-Neuronal Nuclei (1:500;
Millipore, Billerica, MA, USA) and monoclonal anti-glial fibrillary
acidic protein (GFAP; 1:500; Abcam), overnight at 4°C. Having been
washed with 0.01 M PBS three times, the sections were then
incubated with fluorescein isothiocyanate (1:200; Santa Cruz
Biotechnology, Inc.) or Cy3®-conjugated secondary
antibodies (1:200; Abcam) at 37°C for 1 h. Fluorescent signals were
detected by fluorescence microscopy.
Western blotting
Total protein concentration was determined using a
Micro BCA™ Protein Assay kit (Pierce Biotechnology, Inc., Rockford,
IL, USA). Protein samples were resolved in SDS sample buffer. The
protein samples of ~30 mg were run on a 10% SDS-PAGE gel and the
protein in the gel was transferred onto polyvinylidene fluoride
membrane. The membranes were incubated with anti-HO-1 (1:2,000;
Abcam), anti-HO-2 (1:1,000; Abcam) and anti-GAPDH (1:5,000; Abcam)
primary antibodies at 4°C overnight. The membranes were washed with
Tris-buffered saline Tween-20 buffer three times every 10 min. The
membranes were incubated with IRDye 800CW (Li-Cor, Inc., Lincoln,
NE, USA) secondary antibodies for 1 h at room temperature and the
blots were visualized using an Odyssey® scanner (Li-Cor,
Inc.).
ELISA
Briefly, 100 μl diluted plasma was loaded onto
96-well plates. Levels of HO-1 and HO-2 were determined using an
ELISA kit (Shanghai Westtang Bio-tech, Shanghai, China) according
to the manufacturer’s instructions.
Statistical analysis
Statistical analysis was performed using GraphPad
Prism version 5 software (GraphPad Software, Inc., La Jolla, CA,
USA). Measurement data are expressed as the mean ± standard error
of the mean. Differences were assessed using the Student’s t-test
for comparisons, A value of P<0.05 was considered to indicate a
statistically significant difference.
Results
Memory impairment in APP/PS1 transgenic
mice determined through Morris water maze analysis
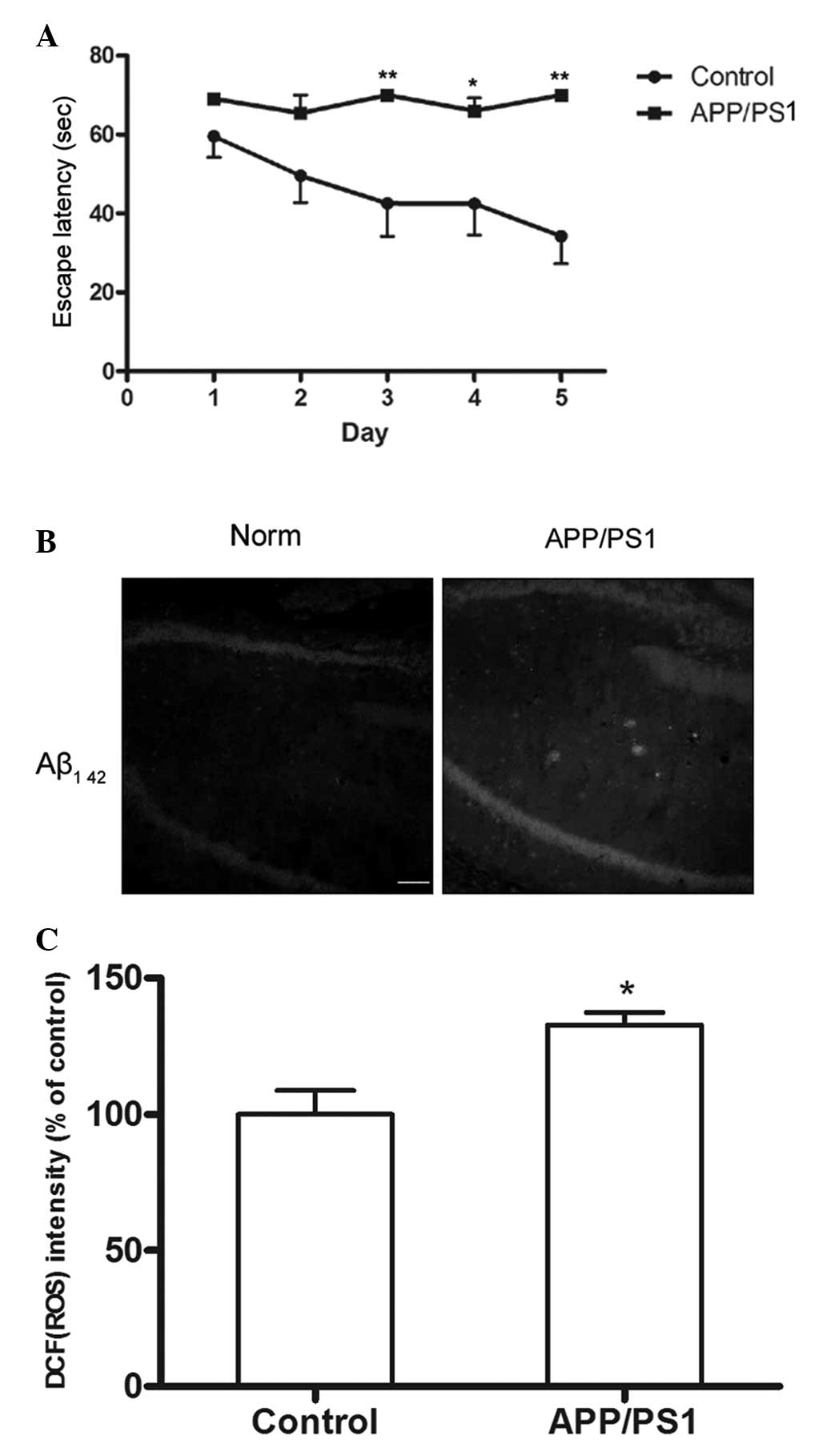
The Morris water maze was employed to detect the
memory ability of the 6-month-old APP/PS1 transgenic mice. It was
found that the mean latency of the APP/PS1 group became
significantly longer than that of the control group between the
third and the fifth day (P<0.05; Fig. 1A). The results showed that the
spatial learning and memory of the APP/PS1 transgenic mice was
impaired.
Increased Aβ plaques and ROS levels in
the hippocampi of APP/PS1 transgenic mice
Small, diffuse Aβ deposits were observed in the
hippocampus, but not in the cortex, of APP/PS1 transgenic mice;
this was due to the fact that the transgenic mice used were only 6
months old. Less Aβ staining was observed in the hippocampus or
cortex of wild-type mice (Fig.
1B). In the present study, the generated ROS were assessed with
the membrane-permeable fluorescent probe DCFH-DA. The ROS levels in
the hippocampi of the APP/PS1 transgenic mice were found to be
significantly higher than those of the wild-type mice (P<0.05;
Fig. 1C).
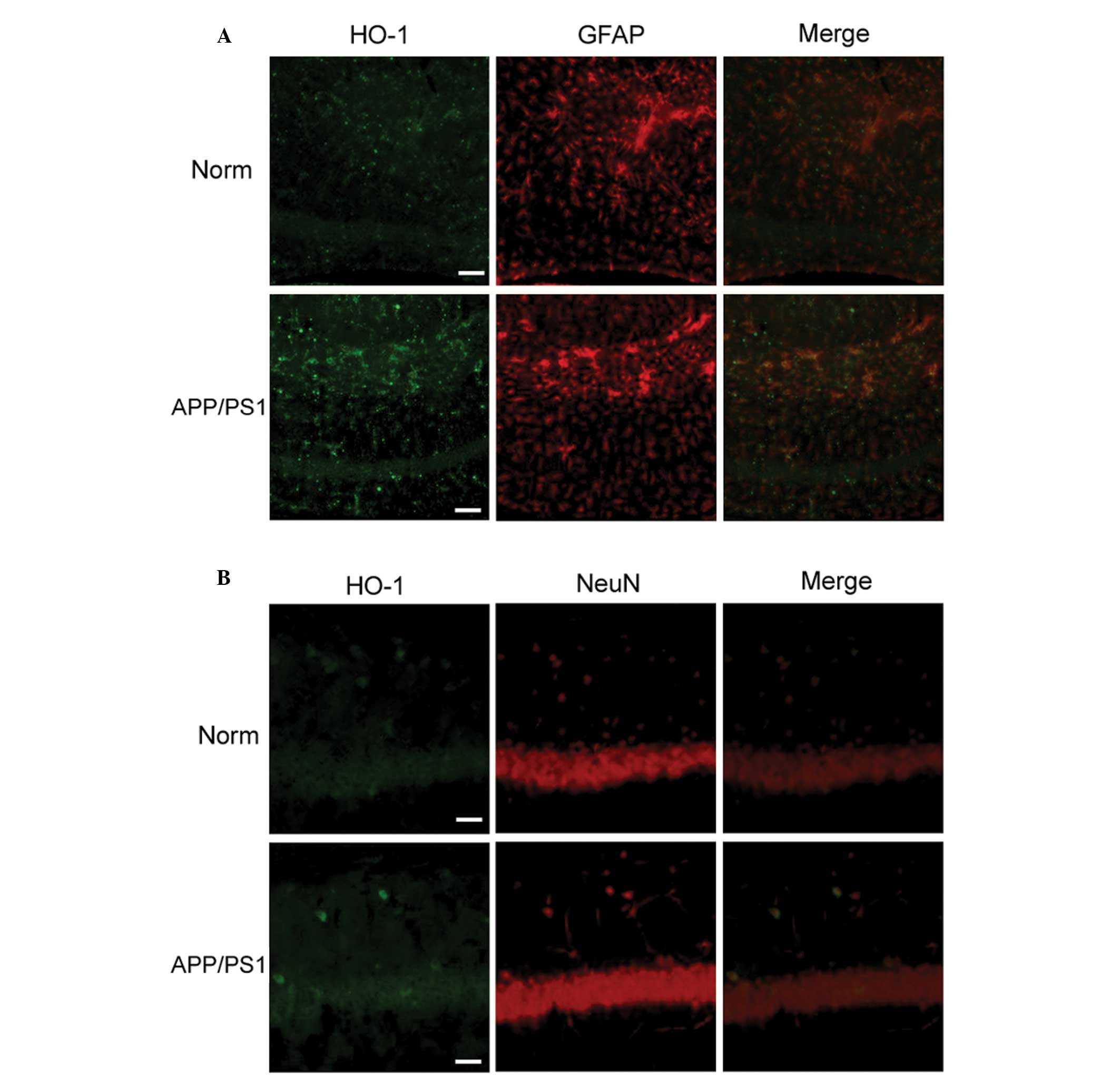
Increased HO-1 expression in APP/PS1
transgenic mice
HO-1 is restricted in expression to small groups of
dispersed neurons and glial cells (6). Using immunofluorescence labeling, it
was observed that astrocytes exhibited increased HO-1
immunoreactivity in the hippocampi of APP/PS1 transgenic mice
compared with that of normal mice. In contrast to the
overexpression of HO-1 in GFAP-positive astrocytes (Fig. 2A), it was observed that neurons
exhibited faint HO-1 immunoreactivity in the hippocampi of APP/PS1
transgenic mice, but that the immunoreactivity was still higher
than that in the control group (Fig.
2B). The result indicated that induced overexpression of HO-1
in astrocytes is an early event in the pathogenesis of AD.
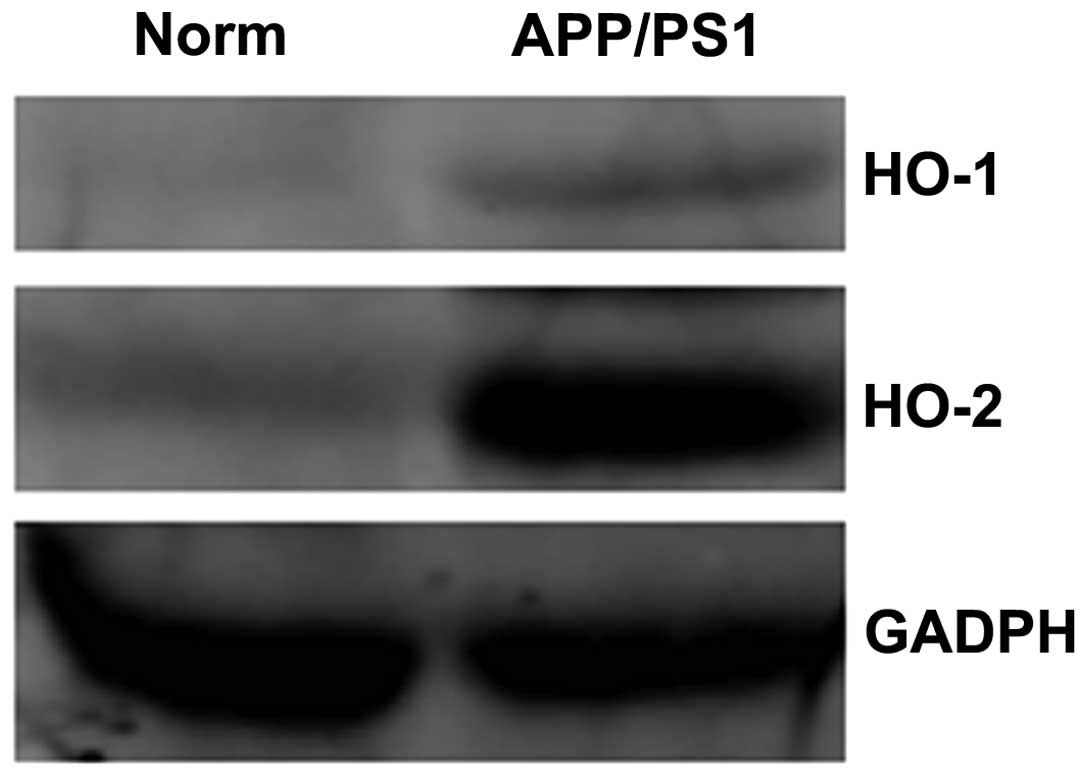
For further confirmation of HO-1 overexpression in
the hippocampi of APP/PS1 transgenic mice, protein extracts were
prepared and analyzed by western blotting. Higher expression levels
of HO-1 were observed in the transgenic mice than in the wild-type
mice (Fig. 3).
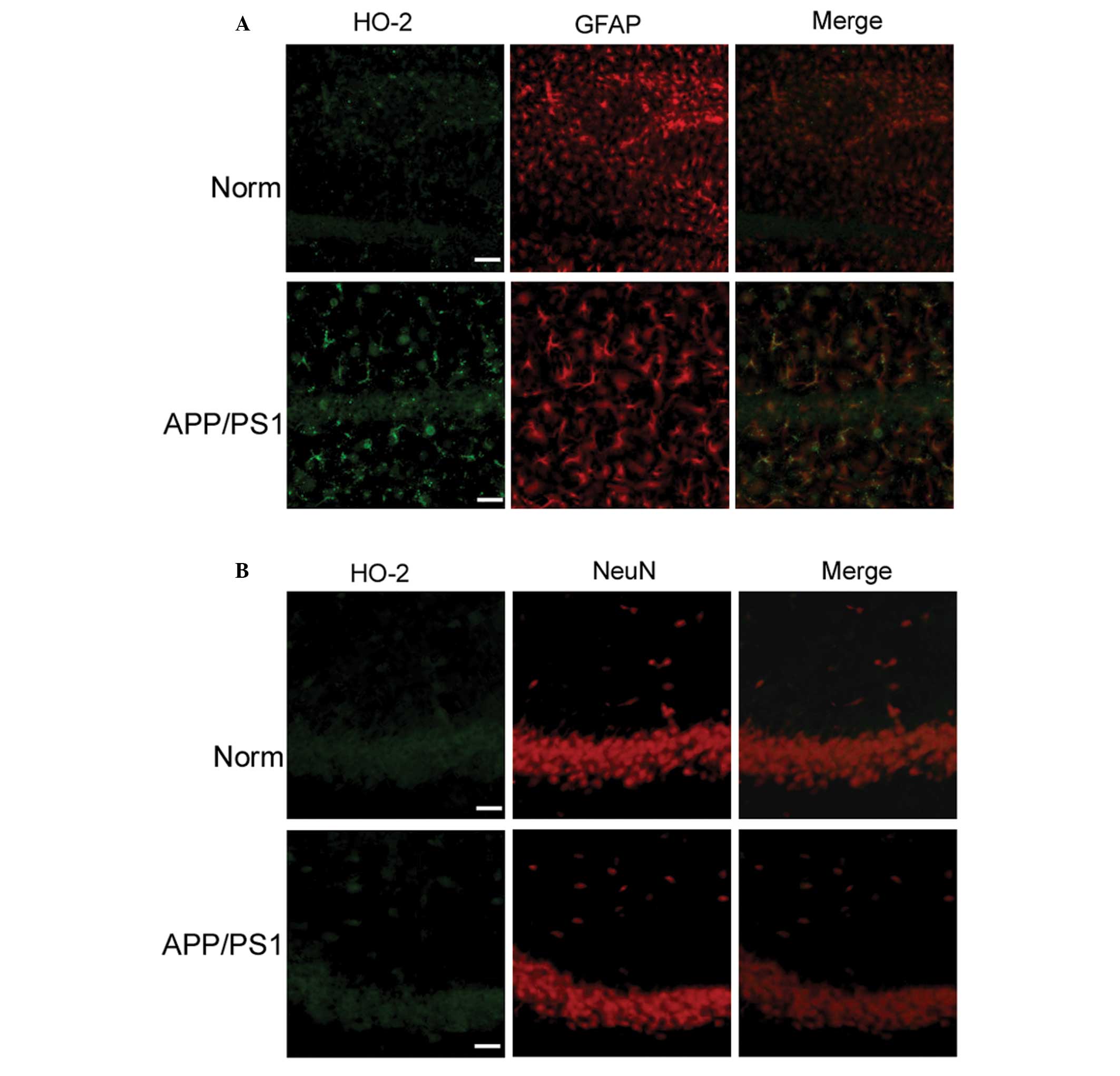
Increased HO-2 expression in APP/PS1
transgenic mice
As an antioxidant enzyme, HO is markedly induced
under conditions of oxidative stress, and several reports have
shown that HO-2 accounts for ~80% of the total rodent brain HO
activity (13,14). Therefore, the changes in HO-2 in
the early stages of AD were investigated in the present study.
Using double-label fluorescence microscopy, it was observed that
the hippocampi of APP/PS1 transgenic mice exhibited more numerous
immunoreactive GFAP cells coexpressing HO-2 protein than the
control mice (Fig. 4A); however,
HO-2 immunoreactivity was hardly detectable in neurons of
transgenic and wild-type mice (Fig.
4B). In order to confirm the overexpression of HO-2 in the
hippocampi of APP/PS1 transgenic mice, protein extracts were
prepared and analyzed by western blotting. HO-2 expression was
shown to be higher in the APP/PS1 transgenic mice than that in the
wild-type mice (Fig. 3). In
addition, more GFAP-positive astrocytes showing HO-2
immunoreactivity were observed than those exhibiting HO-1
immunoreactivity. These results indicated that HO-2 has a more
important role in the early stages of AD.
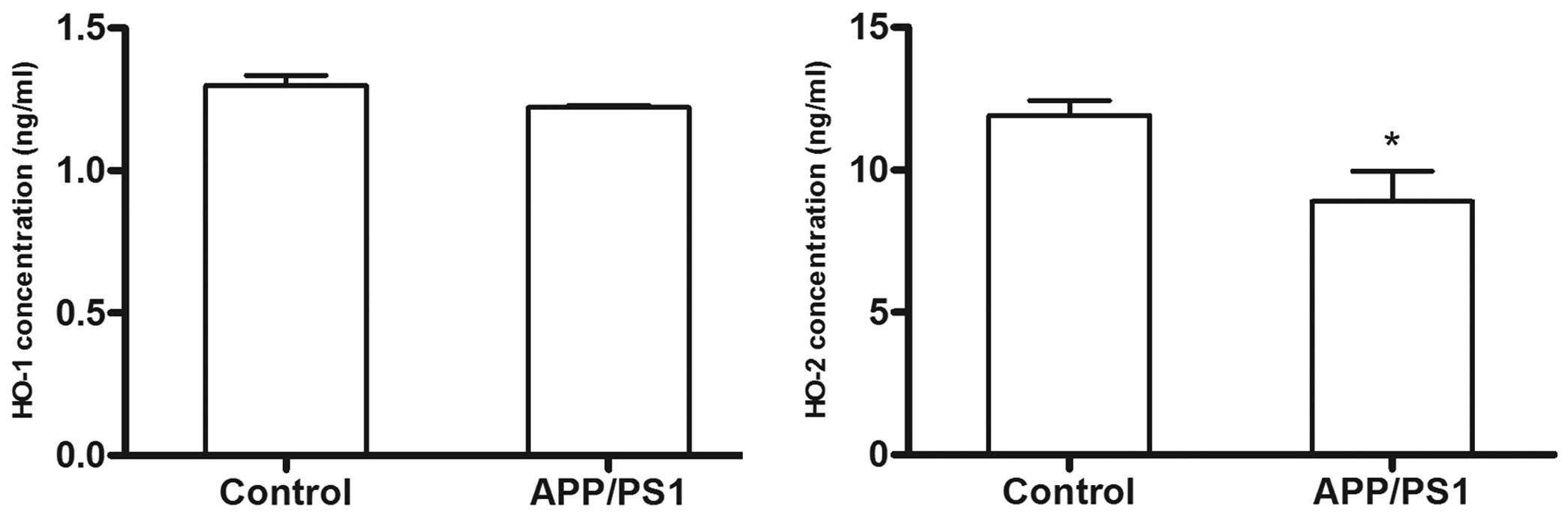
HO-1 and HO-2 concentration in
plasma
In contrast to the overexpression of HO-1 protein in
the brains of the 6-month-old APP/PS1 transgenic mice, the plasma
HO-1 concentration in the transgenic mice showed little difference
from that in the control mice, as determined by ELISA (P>0.05).
In addition, unlike the overexpression pattern of HO-2 in the
brains of the transgenic mice, the plasma HO-2 concentration in the
transgenic mice was lower than that of the control group
(P<0.05). Since the results of plasma HO-2 concentration were
not consistent with those for the AD brain (Fig. 5), it was hypothesized that the HO-2
suppressor activity was more robust than the HO-2 activity in the
plasma of AD mice models.
Discussion
AD is recognized as a clinical pathological disease
with multiple causes, including genetic, environmental and
lifestyle factors (15); oxidative
stress may be one of the underlying factors contributing to the
progression of AD. Oxidative stress is considered to be the
equilibrium state where the production of ROS exceeds the
capability of the antioxidant systems (16). The brain is an organ rich in
cholesterol and polyunsaturated fatty acids and is vulnerable to
oxidative stress (17–20). In a previous study, products of
oxidative stress were found in senile plaques or NFTs (21). As oxidative stress can be induced
by deposited amyloid peptide and accumulated tau (22,23),
it was speculated in the present study that endogenous antioxidant
systems play important roles in the pathogenesis of AD. The HO
system is one of the major endogenous antioxidant systems involved
in AD (24,25).
In a previous study, Huang et al (26) found that injection of Aβ into the
brains of adult Sprague Dawley rats could markedly reduce the
expression of HO-1; however, the present study indicated that HO-1
expression was increased in the hippocampi of 6-month-old APP/PS1
transgenic mice, which was consistent with a study by Schipper
et al (10). At a later
stage, it is likely that HO-1 overexpression began facilitating the
deposits of NFTs and amyloid protein due to toxic product of heme
metabolism (27,28).
The main roles of oxidative stress in the
pathogenesis of AD have been largely investigated, and numerous
studies have reported that plasma levels of oxidative products are
increased in patients with AD compared with controls (29–31).
In support of the hypothesis that plasma antioxidant capacity is
impaired in AD, the present study investigated the HO
concentrations in the plasma of AD disease models. A significant
reduction in HO-2 plasma concentration was observed in 6-month-old
transgenic mice compared with age-matched controls, but plasma HO-1
concentration remained unchanged. These results indicated that HO-2
may play a more important role than HO-1 in the pathological
process of AD.
The present results partially supported the study by
Barone et al (7) on
subjects with AD (particularly HO-1 data); however, the present
HO-2 data were not in agreement with those from patients with AD.
Plasma HO-1 and HO-2 concentrations may provide a novel insight
that the induction of antioxidant capacity, particularly HO-2, is
an early event in the progression of AD.
Acknowledgements
This study was supported by grants from the Shanghai
Health Bureau Youth Fund (no. 20114Y104) and the Shanghai Nature
Science Fund (no. 13ZR1439800).
Abbreviations:
|
AD
|
Alzheimer’s disease
|
|
Aβ
|
amyloid-β
|
|
NFT
|
neurofibrillary tangles
|
|
ROS
|
reactive oxygen species
|
|
HO
|
heme oxygenase
|
|
CO
|
carbon monoxide
|
References
|
1
|
Sayre LM, Perry G and Smith MA: Oxidative
stress and neurotoxicity. Chem Res Toxicol. 21:172–188. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Perrig WJ, Perrig P and Stähelin HB: The
relation between antioxidants and memory performance in the old and
very old. J Am Geriatr Soc. 45:718–724. 1997.PubMed/NCBI
|
|
3
|
Fujita K, Yamafuji M, Nakabeppu Y and Noda
M: Therapeutic approach to neurodegenerative diseases by medical
gases: focusing on redox signaling and related antioxidant enzymes.
Oxid Med Cell Longev. 2012:3242562012. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Pae HO, Lee YC and Chung HT: Heme
oxygenase-1 and carbon monoxide: emerging therapeutic targets in
inflammation and allergy. Recent Pat Inflamm Allergy Drug Discov.
2:159–165. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Barañano DE and Snyder SH: Neural roles
for heme oxygenase: contrasts to nitric oxide synthase. Proc Natl
Acad Sci USA. 98:10996–11002. 2001.PubMed/NCBI
|
|
6
|
Schipper HM: Heme oxygenase expression in
human central nervous system disorders. Free Radic Biol Med.
37:1995–2011. 2004. View Article : Google Scholar
|
|
7
|
Barone E, Di Domenico F, Sultana R, et al:
Heme oxygenase-1 posttranslational modifications in the brain of
subjects with Alzheimer disease and mild cognitive impairment. Free
Radic Biol Med. 52:2292–2301. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Schipper HM, Song W, Zukor H, Hascalovici
JR and Zeligman D: Heme oxygenase-1 and neurodegeneration:
expanding frontiers of engagement. J Neurochem. 110:469–485. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Schipper HM, Cissé S and Stopa EG:
Expression of heme oxygenase-1 in the senescent and
Alzheimer-diseased brain. Ann Neurol. 37:758–768. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Schipper HM, Bennett DA, Liberman A, et
al: Glial heme oxygenase-1 expression in Alzheimer disease and mild
cognitive impairment. Neurobiol Aging. 27:252–261. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Su D, Zhao Y, Xu H, et al: Isoflurane
exposure during mid-adulthood attenuates age-related spatial memory
impairment in APP/PS1 transgenic mice. PLoS One. 7:e501722012.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Ghosh A, Sarkar S, Mandal AK and Das N:
Neuroprotective role of nanoencapsulated quercetin in combating
ischemia-reperfusion induced neuronal damage in young and aged
rats. PLoS One. 8:e577352013. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Ishikawa M, Kajimura M, Adachi T, et al:
Carbon monoxide from heme oxygenase-2 Is a tonic regulator against
NO-dependent vasodilatation in the adult rat cerebral
microcirculation. Circ Res. 97:e104–e114. 2005. View Article : Google Scholar
|
|
14
|
Vreman HJ, Wong RJ, Kadotani T and
Stevenson DK: Determination of carbon monoxide (CO) in rodent
tissue: effect of heme administration and environmental CO
exposure. Anal Biochem. 341:280–289. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Feng Y and Wang X: Antioxidant therapies
for Alzheimer’s disease. Oxid Med Cell Longev. 2012:4729322012.
|
|
16
|
Poon HF, Calabrese V, Scapagnini G and
Butterfield DA: Free radicals and brain aging. Clin Geriatr Med.
20:329–359. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Mancuso M, Coppede F, Migliore L,
Siciliano G and Murri L: Mitochondrial dysfunction, oxidative
stress and neurodegeneration. J Alzheimers Dis. 10:59–73.
2006.PubMed/NCBI
|
|
18
|
Migliore L and Coppede F:
Environmental-induced oxidative stress in neurodegenerative
disorders and aging. Mutat Res. 674:73–84. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Moulton PV and Yang W: Air pollution,
oxidative stress, and Alzheimer’s disease. J Environ Public Health.
2012:4727512012.
|
|
20
|
Adibhatla RM and Hatcher JF: Role of
lipids in brain injury and diseases. Future Lipidol. 2:403–422.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Markesbery WR and Carney JM: Oxidative
alterations in Alzheimer’s disease. Brain Pathol. 9:133–146.
1999.
|
|
22
|
Butterfield DA, Castegna A, Lauderback CM
and Drake J: Evidence that amyloid beta-peptide-induced lipid
peroxidation and its sequelae in Alzheimer’s disease brain
contribute to neuronal death. Neurobiol Aging. 23:655–664.
2002.PubMed/NCBI
|
|
23
|
Cente M, Filipcik P, Pevalova M and Novak
M: Expression of a truncated tau protein induces oxidative stress
in a rodent model of tauopathy. Eur J Neurosci. 24:1085–1090. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Di Domenico F, Barone E, Mancuso C, et al:
HO-1/BVR-a system analysis in plasma from probable Alzheimer’s
disease and mild cognitive impairment subjects: a potential
biochemical marker for the prediction of the disease. J Alzheimers
Dis. 32:277–289. 2012.PubMed/NCBI
|
|
25
|
Butterfield DA, Barone E, Di Domenico F,
et al: Atorvastatin treatment in a dog preclinical model of
Alzheimer’s disease leads to up-regulation of haem oxygenase-1 and
is associated with reduced oxidative stress in brain. Int J
Neuropsychopharmacol. 15:981–987. 2012.PubMed/NCBI
|
|
26
|
Huang TC, Lu KT, Wo YY, Wu YJ and Yang YL:
Resveratrol protects rats from Aβ-induced neurotoxicity by the
reduction of iNOS expression and lipid peroxidation. PLoS One.
6:e291022011.
|
|
27
|
Hui Y, Wang D, Li W, et al: Long-term
overexpression of heme oxygenase 1 promotes tau aggregation in
mouse brain by inducing tau phosphorylation. J Alzheimers Dis.
26:299–313. 2011.PubMed/NCBI
|
|
28
|
Schipper HM: Brain iron deposition and the
free radical-mitochondrial theory of ageing. Ageing Res Rev.
3:265–301. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Schipper HM, Chertkow H, Mehindate K,
Frankel D, Melmed C and Bergman H: Evaluation of heme oxygenase-1
as a systemic biological marker of sporadic AD. Neurology.
54:1297–1304. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Yu HL, Chertkow HM, Bergman H and Schipper
HM: Aberrant profiles of native and oxidized glycoproteins in
Alzheimer plasma. Proteomics. 3:2240–2248. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Maes OC, Kravitz S, Mawal Y, et al:
Characterization of alpha1-antitrypsin as a heme oxygenase-1
suppressor in Alzheimer plasma. Neurobiol Dis. 24:89–100. 2006.
View Article : Google Scholar : PubMed/NCBI
|