Introduction
Hepatocellular carcinoma (HCC) is the sixth most
common solid tumor in the world and the third most common cause of
cancer mortality (1). Surgical
resection and liver transplantation surgery are the only potential
curative treatments for patients with early stage tumors. However,
a number of patients with advanced HCC are not suitable for surgery
due to the presence of metastases (2). Systemic chemotherapy is the main
treatment method for patients with advanced HCC. However,
traditional chemotherapy treatments provide little efficacy in
patients with advanced HCC and often provide no benefit to survival
rate (3). Recently, agents
targeting certain key pathways have provided new methods for the
treatment of HCC. For instance, 5-fluorouracil (5-Fu), an inhibitor
of thymidylate synthase, was reported as one of the first
chemotherapeutic agents assessed for the treatment of HCC with a
response rate of ~10% (4).
Therefore, novel pharmacological strategies for the treatment of
HCC are urgently required.
Multidrug resistance (MDR), including intrinsic or
acquired MDR, is a multi-factorial phenomenon and the major cause
of chemotherapeutic failure during cancer treatment. MDR markedly
restrains the management of clinical HCC chemotherapy. MDR
effectors include the transporter glycoproteins P-glycoprotein
(P-gp) and multidrug resistance-associated protein 1 (MRP1), whose
overexpression is generally considered to be the underlying
mechanism responsible for the MDR of tumor cells.
Metformin is a commonly used biguanide antidiabetic
drug that has anti-proliferative properties and its use in diabetic
patients has been linked to a reduction in the incidence of cancer
(5). Metformin activates
AMP-activated protein kinase (AMPK), most likely by interfering
with mitochondrial respiratory complex I (6). Activation of AMPK regulates the
growth of tumor cells through inhibition of the mammalian target of
rapamycin (mTOR) pathway. This pathway is generally involved in the
control of translation initiation and protein synthesis (7).
The mTOR pathway is aberrantly activated in HCC
(8). Inhibition of mTOR has been
demonstrated to suppress liver tumor growth and metastasis
(9,10). Several clinical and preclinical
studies have demonstrated that the mTOR inhibitor rapamycin and its
analogues can be used to treat various types of solid tumor,
including esophageal squamous cell carcinoma (11), lung cancer (12), renal cell carcinoma (13) and prostate cancer (14). Therefore, the mTOR pathway may be a
prospective target for the treatment of HCC. Furthermore, as
hypoxia has well-documented effects on cancer malignancy and
resistance to chemotherapy (15),
hypoxia inducible factor 1α (HIF-1α), a downstream protein of
mTORC1 that regulates the expression of P-gp and MRP1, has emerged
as an attractive target for cancer therapy (16).
Metformin is a potential anti-cancer agent targeting
the mTOR/HIF-1α pathway. In the present study, the role of
metformin as a chemosensitizer was investigated. The Bel-7402
derived multidrug-resistant cell line Bel/Fu, was used as an MDR
model.
Materials and methods
Cell culture
The present study was approved by the Second
Affiliated Hospital of Dalian Medical University Research Ethics
Committee (Dalian, China). The human hepatocellular carcinoma MDR
cell line Bel-7402/5-Fu (Bel/Fu) was obtained from KeyGen Biotech
Co., Ltd. (Nanjing, China). The cell line was cultured in RPMI-1640
medium (Gibco-BRL, Carlsbad, CA, USA) supplemented with 10% fetal
bovine serum (FBS; Gibco-BRL), 100 μg/ml penicillin and 100 μg/ml
streptomycin (Invitrogen Life Technologies, Carlsbad, CA, USA) at
37°C in a 5% CO2 atmosphere. The MDR Bel/Fu cells were
maintained in medium containing 20 μg/ml 5-Fu.
Reagents
Metformin (cat no. D150959-5G) and 5-Fu (cat no.
F6627-1G) were purchased from Sigma-Aldrich (St. Louis, MO, USA).
The Cell Counting kit-8 (CCK-8; cat no. KGA317), Annexin
V-fluorescein isothiocyanate (FITC) Apoptosis Detection kit (cat
no. KGA108) and the Cell Cycle Detection kit (cat no. KGA512) were
purchased from KeyGen Biotech Co., Ltd.
Antibodies
The following polyclonal rabbit anti-human
antibodies were used for western blot analysis: HIF-1α (N-term; cat
no. AP4776a; diluted 1:1,000), active caspase-3 (cat no. AJ1131b;
diluted 1:1,000), B-cell lymphoma 2 (Bcl-2; cat. no AJ1082a;
diluted 1:1,000), cyclin-dependent kinase 4 (CDK4; cat no. AP7520b;
diluted 1:1,000) and cyclin D1 (cat no. AP2612c, diluted 1:1,000)
obtained from Abgent (San Diego, CA, USA); AMPKα (Ab-172; cat no.
B0003, diluted 1:500), phosphorylated AMPKα (phospho-Thr172; cat
no. A0003; diluted 1:500), mTOR (cat no. B7156; diluted 1:500) and
phosphorylated mTOR (phospho-Ser2448; cat no. A7156; diluted 1:500)
purchased from Assay Biotech Co., Inc. (Sunnyvale, CA, USA); P-gp
(cat no. PA5-28801; diluted 1:500) and MRP1 (cat no. PA5-30594;
diluted 1:500) obtained from Thermo Fisher Scientific Inc.
(Rockford, IL, USA). The mouse anti-human monoclonal β-actin
antibody (cat no. sc-47778, diluted 1:1,000) was purchased from
Santa Cruz Biotechnology, Inc. (Santa Cruz, CA, USA). The
polyclonal goat anti-rabbit and polyclonal goat anti-mouse IgG,
peroxidase-conjugated secondary antibodies (cat nos. 31460 and
31430; diluted 1:10,000) were purchased from Thermo-Pierce
(Rockford, IL, USA).
Cell viability assay
Cell viability was determined using the CCK-8 assay
according to the manufacturer’s instructions. Cells
(5×103) were seeded into each well of a 96-well plate
and cultured in 100 μl RPMI-1640 medium supplemented with 10% FBS,
100 μg/ml penicillin and 100 μg/ml streptomycin. After 24 h, single
(100 μg/ml 5-Fu or 10 mM Met) or combined agents (100 μg/ml 5-Fu
plus 10 mM Met) were added into the culture medium. The cells were
incubated at 37°C for 24, 48 or 72 h, following which the medium
was exchanged for 100 μl RPMI-1640 medium and 10 μl CCK-8 reagent.
The cells were incubated for 2 h at 37°C. Finally, the optical
density was measured using an EnSpire™ 2300 Multilabel Reader
(PerkinElmer, Inc., Waltham, MA, USA) at 450 nm. Five replicates
were prepared for each condition. The mean values were calculated
and growth curves were produced.
Western blot analysis
Following the different treatments, the cells were
harvested and lysed in radioimmunoprecipitation assay buffer (cat
no. KGP702; KeyGen Biotech Co., Ltd.) supplemented with 1 mM
phenylmethylsulfonyl fluoride (cat no. KGP610; KeyGen Biotech Co.,
Ltd.) and 1 mM phosphatase inhibitor cocktail (cat no. KGP602;
KeyGen Biotech Co., Ltd.). The mixture was centrifuged at 12,000 ×
g for 20 min and the supernatant was collected. The protein
concentration was determined using a BCA assay kit (cat no. KGPBCA;
KeyGen Biotech Co., Ltd.) and each sample contained 30 μg protein
per 10 μl. The protein samples were mixed with loading buffer (cat
no. KGP101; KeyGen Biotech Co., Ltd.) and the proteins were
separated using 6 or 10% sodium dodecyl sulfate-polyacrylamide gel
electrophoresis and transferred onto polyvinylidene difluoride
membranes (Bio-Rad, Hercules, CA, USA). Following soaking in a
blocking buffer (TBS with 5% nonfat dry milk) for 2 h, the blots
were incubated at 4°C overnight with the primary antibody and
subsequently incubated at 37°C for 1 h with the horseradish
peroxidase-conjugated secondary antibody. The bands were visualized
using chemiluminescence and images were captured using a ChemiDoc
XRS imaging system (Bio-Rad). These were analyzed using Image Lab
software (Bio-Rad).
Reverse transcription-polymerase chain
reaction (RT-PCR)
Total RNA was isolated using an RNAiso™ Plus kit
(Takara Bio, Inc., Shiga, Japan). Total RNA (500 ng) was
reverse-transcribed into first-strand cDNA using a Takara RNA PCR
kit (AMV Ver. 3.0; Takara Bio, Inc.) according to the
manufacturer’s instructions. The following primers were used for
amplification: multidrug resistance 1 (MDR1), forward
5′-CCCATCATTGCAATAGCAGG-3′ and reverse 5′-GTT CAAACTTCTGCTCCTGA-3′;
MRP1, forward 5′-TGAAGG ACTTCGTGTCAGCC-3′ and reverse 5′-GTCCATGAT
GGTGTTGAGCC-3′; β-actin, forward 5′-GCATGGAGTCCT GTGGCAT-3′ and
reverse 5′-CTAGAAGCATTTGCG GTGG-3′. The PCR reactions were
subjected to the following amplification conditions: denaturation
at 94°C for 30 sec, annealing (53°C for MDR1, 57°C for MRP1 and
58°C for β-actin) for 30 sec and extension at 72°C for 30 sec. MDR1
was incubated for 32 cycles, MRP1 for 25 cycles and β-actin for 30
cycles. The PCR products were analyzed by 2% gel
electrophoresis.
Flow cytometric analysis
To evaluate the effects of single or combined agents
on cell cycle arrest and the induction of apoptosis, the cells were
examined using an Annexin V-FITC Apoptosis Detection kit and a Cell
Cycle Detection kit, according to the manufacturer’s instructions.
Bel/Fu cells were seeded into 6-well plates (1×105 and
2×105 cells per dish, respectively). For cell cycle
analysis, following the treatment with single or combined agents
for 48 h, a total of 1×106 cells were pelleted by
centrifugation at 1,000 × g for 5 min and washed twice with PBS.
The cell pellets were then resuspended in 500 μl ice-cold 70%
ethanol and incubated at 4°C overnight. The fixed cells were
centrifuged at 1,000 × g for 5 min and the pellets were washed with
PBS. Following incubation with 100 μl RNase A (10 μg/ml) at 37°C
for 30 min in the dark, the cells were resuspended in 400 μl
propidium iodide (PI; 50 μg/ml) and placed in the dark at 4°C for
30 min. The stained cells were analyzed using an Accuri C6 flow
cytometer (Accuri Cytometers, Inc., Ann Arbor, MI, USA). For
analysis of apoptosis, the cells were trypsinized, washed with cold
PBS and suspended in PBS. Subsequently, the cells were stained
using Annexin V-FITC reaction reagent (10 μl Annexin V-FITC, 5 μl
PI) at 37°C in the dark for 30 min. The stained cells were analyzed
using an Accuri C6 flow cytometer (Accuri Cytometers, Inc.).
Statistical analysis
SPSS 13.0 statistical software was used for
statistical analysis (SPSS, Inc., Chicago, IL, USA). Values are
presented as the mean ± standard deviation. Statistical analyses
were performed using Student’s t-test and the analysis of multiple
groups was performed using analysis of variance, followed by the
Student-Newman-Keuls post hoc test.
Results
Metformin has a synergistic
anti-proliferative effect with 5-Fu in Bel/Fu cells
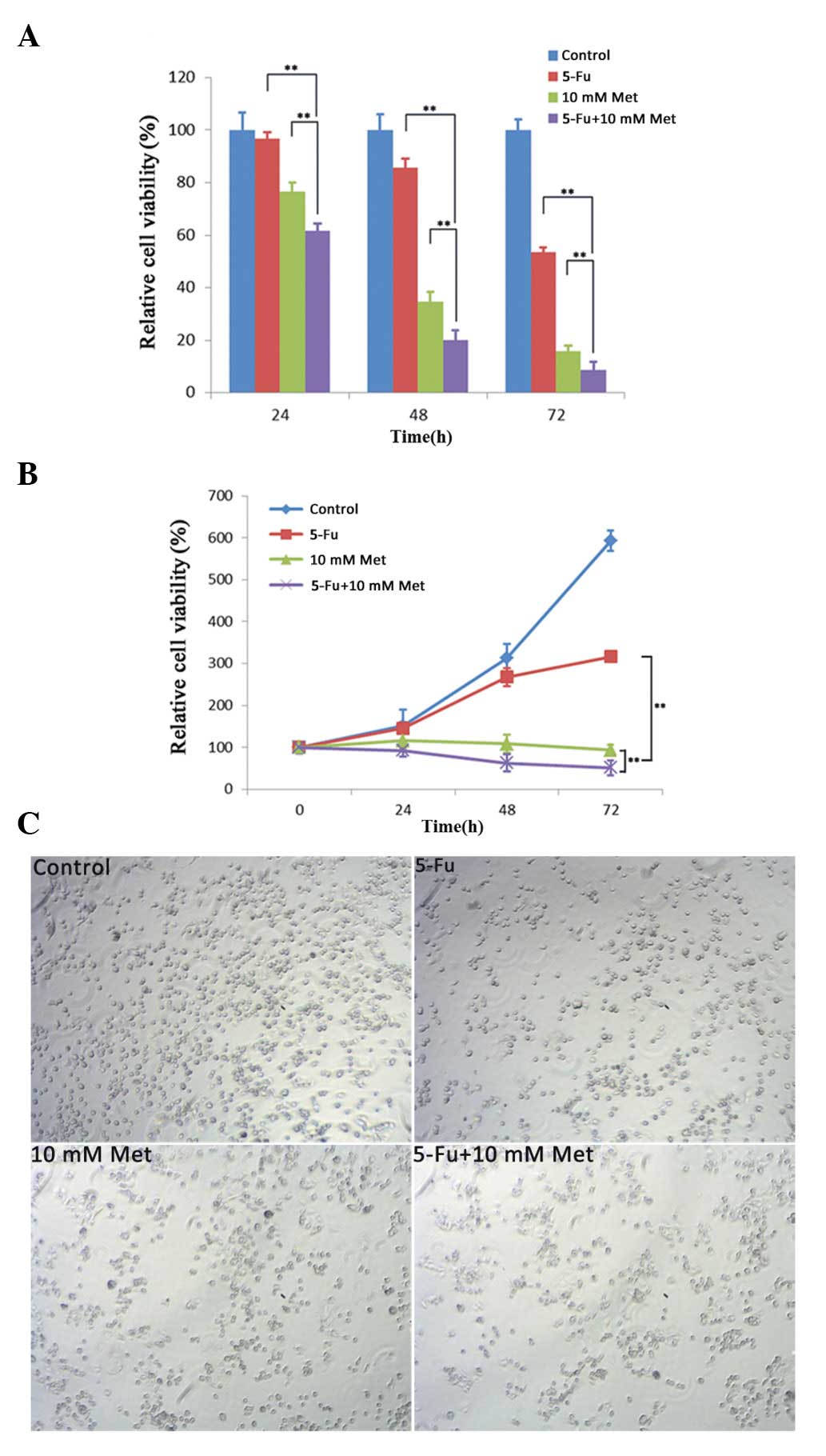
CCK-8 analysis was used to assess the effect of 5-Fu
and metformin on the proliferation of Bel/Fu cells. As shown in
Fig. 1A and B, the course of
proliferation was observed for 3 days after the addition of the
chemotherapeutic agents. All groups exhibited a time-dependent
inhibition of cell proliferation. Following treatment with 100
μg/ml 5-Fu, 10 mM metformin or a combination of the agents for 48
h, the cell viability rates were 85.58±3.76, 34.62±3.65 and
20.13±3.60%, respectively. The combination of the two agents
further decreased the proliferation rate of the Bel/Fu cells
compared with application of a single drug. Microscopic examination
demonstrated a significant decrease in cell density, which
correlated with the cell viability results. In conclusion,
metformin had a synergistic anti-proliferative effect with 5-Fu in
the Bel/Fu cells.
Combined treatment with 5-Fu and
metformin promotes apoptosis and induces cell cycle arrest in
Bel/Fu cells
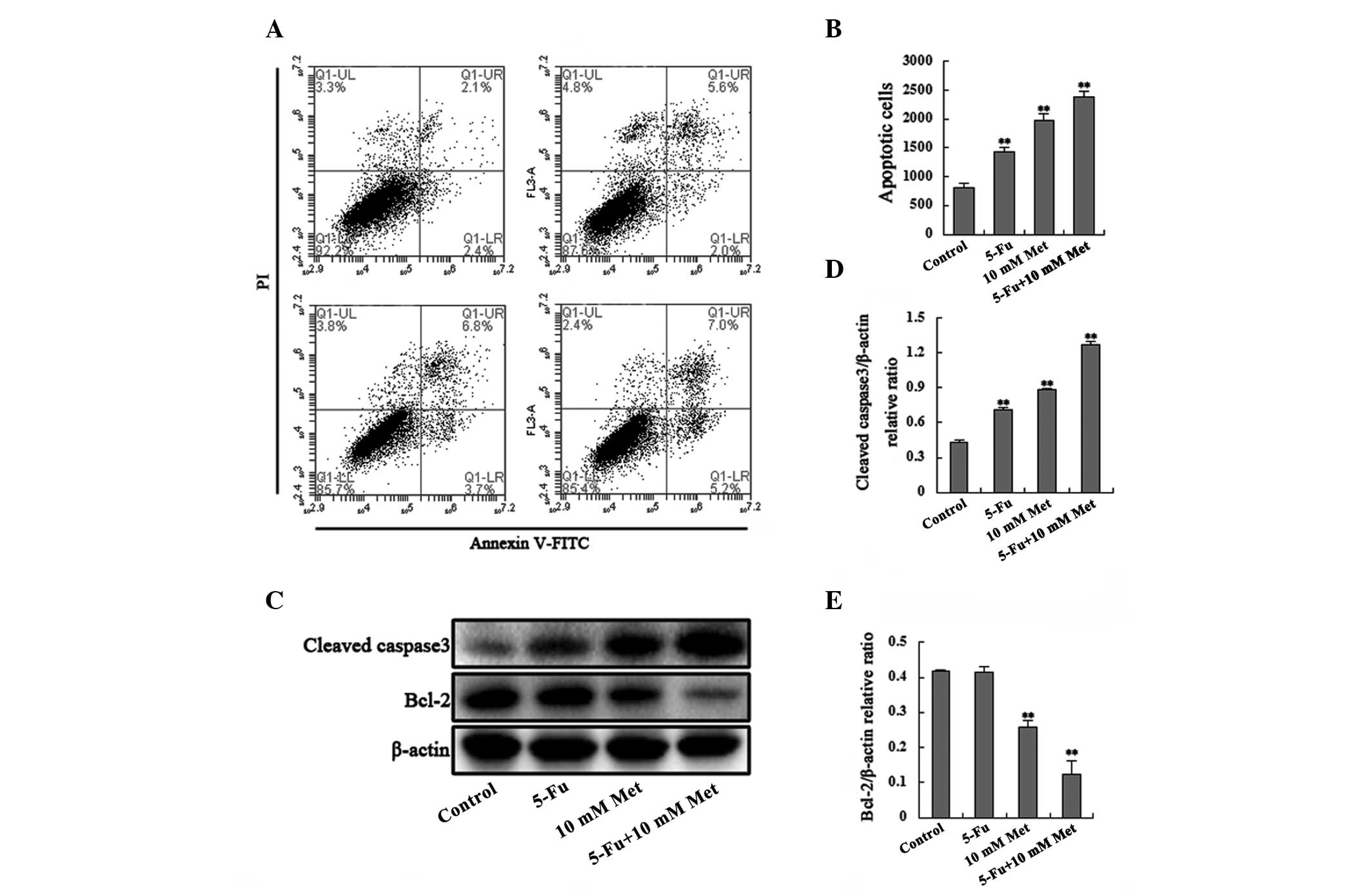
To determine whether the drug application-induced
decrease in cell viability was accompanied by cell cycle arrest or
apoptosis, cell cycle progression and induction of apoptosis were
analyzed. The number of apoptotic cells and the percentages of
cells in each phase of the cell cycle were examined using flow
cytometry after 48 h of incubation with the agents (100 μg/ml 5-Fu,
10 mM Met, or 100 μg/ml 5-Fu plus 10 mM Met). A significant
increase in the number of apoptotic cells and cells undergoing
G0/G1 cell cycle arrest was observed in the Bel/Fu cells treated
with the combined agents compared with the cells treated with a
single agent (Figs. 2A, 2B,
3A and 3B). The Bel/Fu cells were
cultured in the presence of the vehicle, 100 μg/ml 5-Fu, 10 mM
metformin or a combination of 5-Fu and metformin for 48 h. The
total percentage of apoptotic cells, including early apoptotic and
late apoptotic/necrotic cells, was 4.5±0.58, 7.6±0.74, 10.5±1.21
and 12.2±0.71%, respectively. In addition, the percentage of cells
in the G0/G1 phase was 24.52±2.82% for the vehicle control cells,
26.92±3.23% for the 5-Fu cells, 37.12±3.21% for the metformin cells
and 43.33±3.18% for the combined treatment.
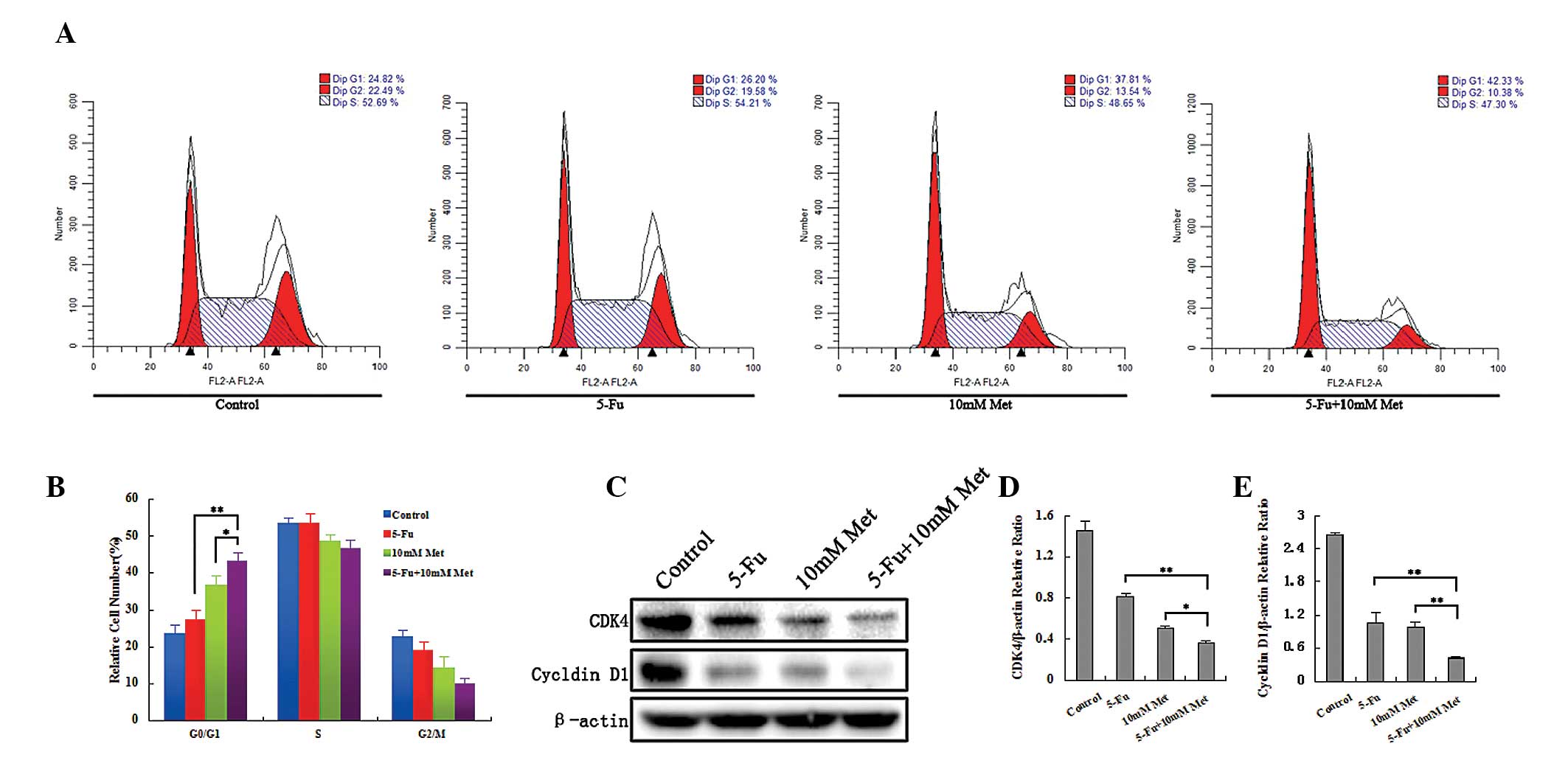 | Figure 3Combined treatment with 5-Fu and Met
induces cell cycle arrest in Bel/Fu cells. (A) Following single or
combined treatment with 100 μg/ml 5-Fu and 10 mM Met, Bel/Fu cells
were stained with propidium iodide. The cell cycle distribution was
measured using flow cytometric analysis. (B) Percentages of cells
in each phase of the cell cycle are shown in the bar graph.
(*P<0.05, **P<0.01, the combined
treatment group, compared with the single agent treatment group)
(C) Levels of cyclin D1 and CDK4 were monitored using western blot
analysis. (D and E) Band intensities were quantified using Image
Lab 5.0 software and were normalized to β-actin.
(*P<0.05, **P<0.01, the combined
treatment group, compared with the single agent treatment group).
5-Fu, 5-fluorouracil; Bcl-2, B-cell lymphoma 2; Met, metformin;
CDK4, cyclin-dependent kinase 4; Bel/Fu, Bel-7402/5-Fu. |
Furthermore, to confirm that the agents induced
apoptosis and G0/G1 cell cycle arrest in the Bel/Fu cells, the
levels of cleaved caspase-3, Bcl-2, cyclin D1 and CDK4 were
monitored using western blot analysis in the Bel/Fu cells (Fig. 2C–E and Fig. 3C–E). Activation of the
pro-apoptotic caspase-3 and downregulation of the apoptosis
suppressor Bcl-2 were associated with agent-induced apoptosis.
Cyclin D1 and CDK4, which are responsible for the transition from
G0/G1 phase to the S phase, were downregulated following combined
or single agent treatment. Thus, these results suggested that
metformin promoted apoptosis and induced G0/G1 cell cycle arrest in
the Bel/Fu cells.
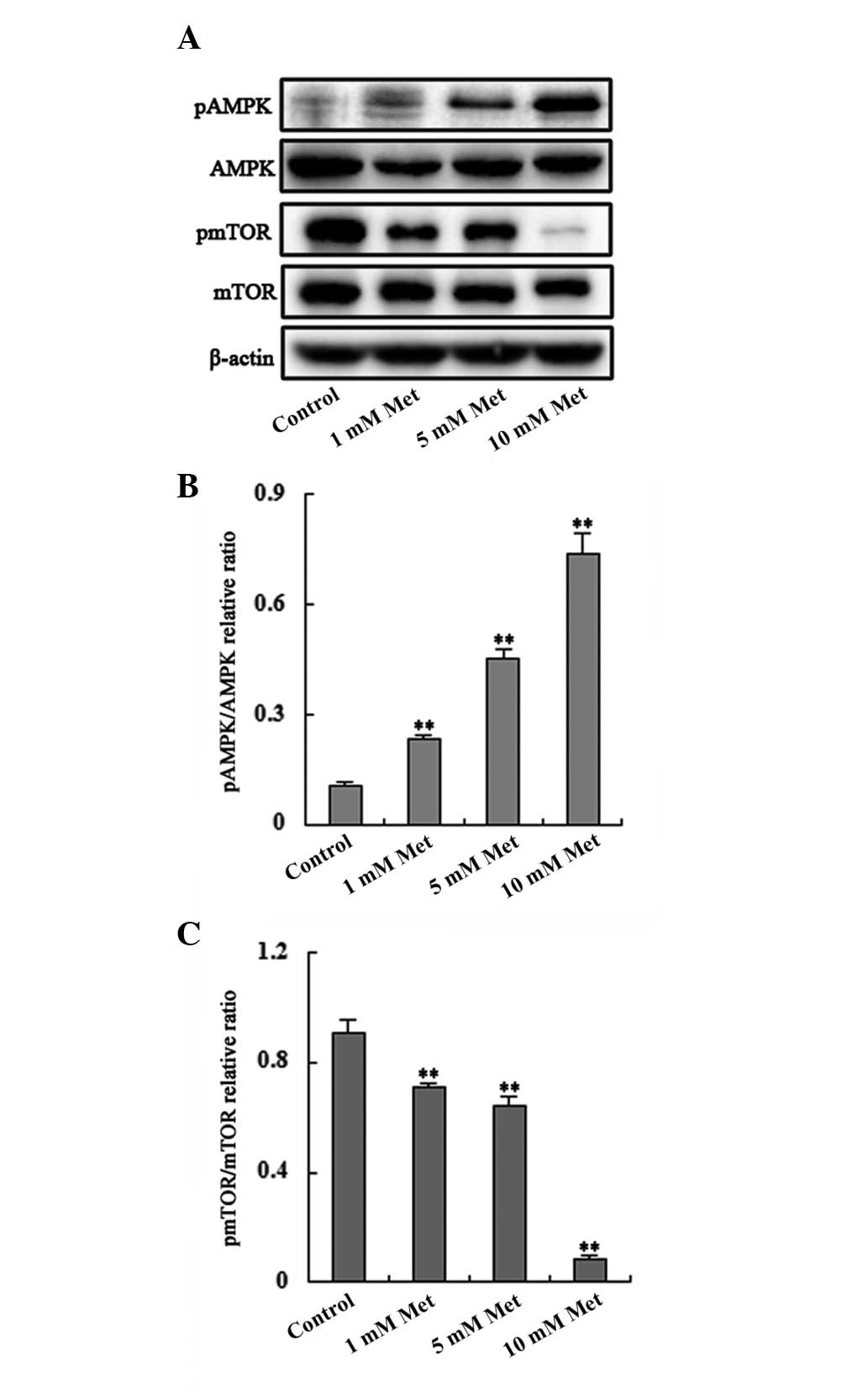
Metformin treatment results in increased
AMPK phosphorylation and decreased mTOR phosphorylation in a
dose-dependent manner
To evaluate the specific effect of metformin on the
AMPK/mTOR pathway, which is widely considered to be the most common
target of metformin, western blot analysis was used to evaluate the
AMPK/mTOR pathway in Bel/Fu cells. Metformin treatment resulted in
enhanced AMPK phosphorylation and reduced mTOR phosphorylation in a
dose-dependent manner (Fig.
4).
Metformin treatment results in decreased
HIF-1α, MDR1 and MRP1 expression in Bel-Fu cells
HIF-1α is a downstream protein of the mTOR pathway
that transcriptionally regulates the expression of P-gp/MDR1 and
MRP1, which are associated with MDR. Thus, the present study
examined the expression of these genes in Bel/Fu cells using RT-PCR
and western blot analysis in order to determine the effects of
metformin on these genes. The results demonstrated that the
expression of HIF-1α, MDR1 and MRP1 in the Bel/Fu cells decreased
significantly with increasing concentrations of metformin (Fig. 5). HIF-1α may transcriptionally
regulate the expression of P-gp/MDR1 and MRP1.
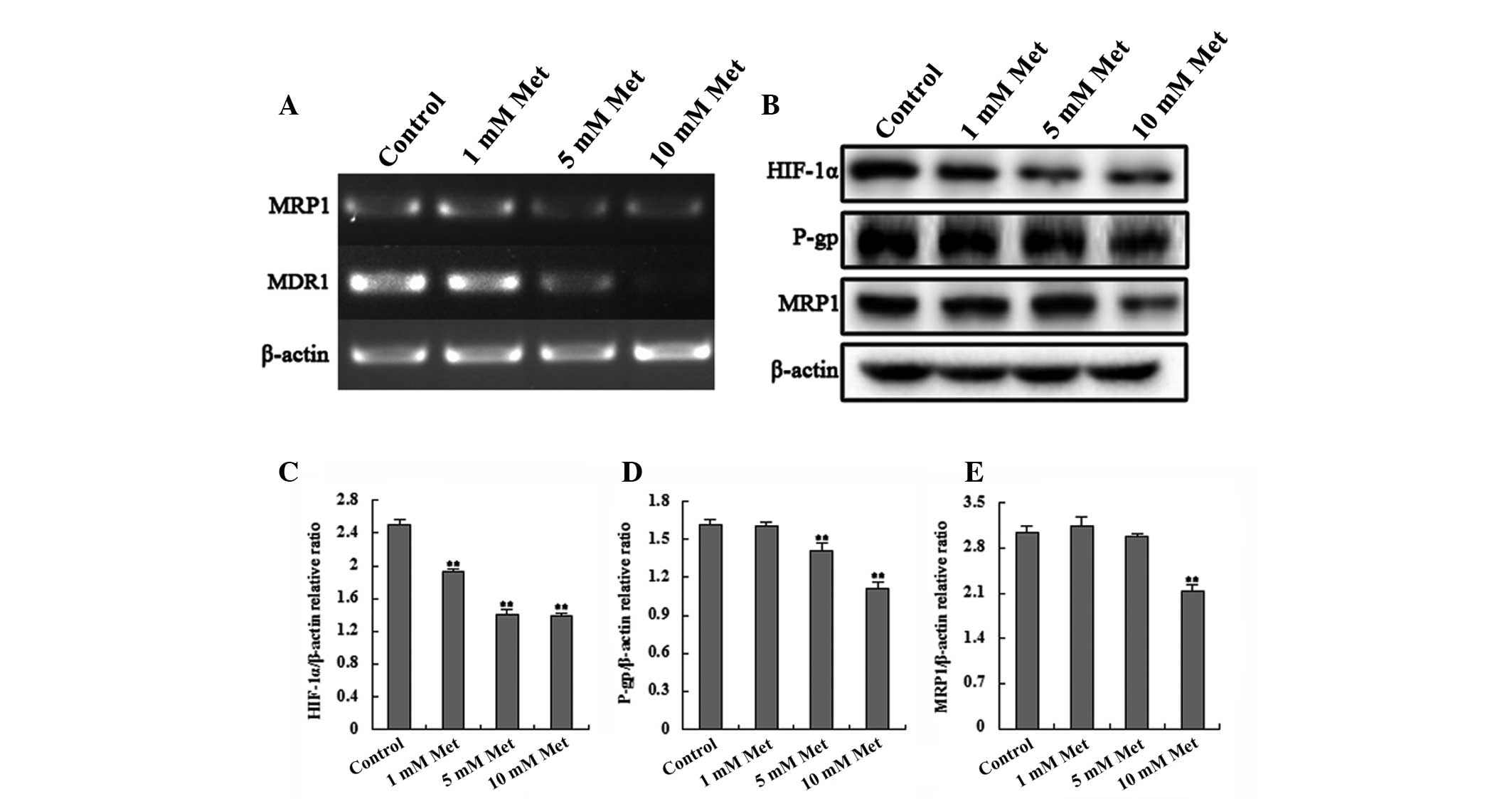 | Figure 5Met affects the expression of HIF-1α,
MDR1/P-gp and MRP1. (A) Reverse transcription-polymerase chain
reaction analysis of MDR1 and MRP1 expression in the different
treatment groups. (B) Western blot analysis of HIF-1α, MDR1 and
MRP1 in the different treatment groups. (C–E) Relative density of
the western blotting bands demonstrated that levels of HIF-1α, P-gp
and MRP1 decreased with increasing concentrations of Met in a
dose-dependent manner, with a statistically significant difference
between the treatments and the controls (**P <0.01,
compared with the control). P-gp and MRP1 protein band densities
were normalized to their corresponding β-actin bands.
(**P<0.01, compared with the control). HIF-1α,
hypoxia-inducible factor-1α; MDR1, multidrug resistance-associated
protein 1; P-gp, P-glycoprotein; MRP1, multidrug resistance protein
1; Met, metformin. |
Discussion
HCC is the sixth most common solid tumor in the
world and the third leading cause of cancer mortality worldwide
(1). MDR is a multifactorial
phenomenon and the major obstacle in the successful and effective
chemotherapeutic treatment of cancer. HCC can easily acquire MDR,
which leads to unsatisfactory effects during chemotherapy treatment
(17). Therefore, traditional
chemotherapy treatments achieve little efficacy in patients with
advanced HCC and are often no benefit to survival rates (3). Thus, approaches to reduce the MDR
properties of cancer cells may enhance the efficacy of
chemotherapeutic agents in the treatment of HCC.
Metformin is a representative therapeutic agent for
type 2 diabetes mellitus. Previous studies have reported that
metformin activates AMPK (18) and
inhibits hepatic cancer cell growth (19). However, the effect of metformin on
the MDR of HCC cells remains to be elucidated. The present study
demonstrated that metformin reversed the MDR of Bel/Fu cells, which
were used as an MDR model, by targeting the AMPK/mTOR/HIF-1α
pathway.
To evaluate the effect of metformin on the Bel/Fu
cells, the cell viability of cells treated with vehicle controls
(100 μg/ml 5-Fu, 10 mM metformin and combined treatment of 100
μg/ml 5-Fu and 10 mM metformin) was examined. The cell viability
was more significantly inhibited in the combination group than in
the single agent groups and the cell viability inhibition rate was
also time dependent. The combination of the two agents decreased
the proliferation rate of the Bel/Fu cells compared with the single
drug treatments. Additionally, the apoptotic rate increased
consistently with the decrease in cell viability. These results
indicated that metformin may reverse the MDR of Bel/Fu cells.
The sensitivity of cells to chemotherapy may be
evaluated by examining the cell cycle distribution and the present
study analyzed this in the different treatment groups. As mentioned
above, no significant different was identified between the 5-Fu
group and the control group. However, the proportion of cells in
the G1 phase was markedly increased in the 10 mM metformin group
and in the combination group, while the proportion of cells in the
S, G2 and M phases decreased concordantly. Therefore, the present
study demonstrated that the expression of cyclin D1 and CDK4, which
facilitate transition from the G1 to the S phase during cell
proliferation, were decreased in the 5-Fu, 10 mM metformin and
combination treatment groups compared with the control group. As
cell apoptosis is another important indicator used to measure the
chemotherapeutic sensitivity of cells, the apoptotic rate of each
group was analyzed using flow cytometry. The apoptotic rate of the
5-Fu + 10 mM metformin group was significantly higher than the
other groups. Coincidently, the expression of cleaved caspase-3
increased and the expression of Bcl-2 decreased in the 5-Fu, 10 mM
metformin and combination treatment groups compared with the
control group. These findings confirmed that metformin reversed the
MDR of the Bel/Fu cells.
Previous studies have demonstrated that P-gp and
MRP1, which act as energy-dependent drug efflux pumps, are
important in MDR (20). According
to previous studies, mTORI decreases the translation of HIF-1α
(21). Overexpression of HIF-1α
significantly upregulates the expression of P-gp and MRP1,
indicating that HIF-1α may confer hypoxia-induced drug resistance
by reducing intracellular drug accumulation (22,23).
However, metformin inhibits the phosphorylation of mTOR by
activating AMPK (23). Therefore,
the present study also evaluated the expression of these proteins
using western blot analysis. The activation of AMPK increased with
increasing concentrations of metformin. By contrast, the
phosphorylation of mTOR was inhibited, leading to a subsequent
decrease in HIF-1α. Coincidently, the expression of P-gp and MRP1
decreased in the metformin-treated cells in a dose-dependent manner
at the mRNA and protein levels; P-gp is a translational product of
the MDR1 gene.
Previous studies have suggested that metformin may
improve oxygenation and suppress the accumulation of HIF-1α in
diseases associated with tumors or diabetes through the activation
of the AMPK/mTOR pathway and the repression of oxygen consumption
(24–27). HIF-1, a basic helix-loop-helix
transcription factor, is important in regulating the transcription
of various target genes in response to hypoxia (28). HIF-1α is an oxygen-regulated
subunit that mediates the essential function of HIF-1. Previous
studies have demonstrated that the overexpression of HIF-1α may
contribute to the pathogenesis of tumor resistance to chemotherapy
(29,30). P-gp and MRP1 are regarded as
energy-dependent membrane efflux pumps and are widely considered to
be transcriptionally regulated by HIF-1α in multiple types of human
tumor (22,31–33).
Consequently, the results of the present study implied that
metformin reversed the MDR of Bel/Fu cells by targeting the
AMPK/mTOR/HIF-1α/P-gp and MRP1 signaling pathways. Therefore,
metformin may be important in combination chemotherapy treatments
for HCC and other malignances.
In conclusion, the present study revealed the role
of metformin, an inexpensive and widely used antidiabetic drug
without severe adverse effects, in reversing the MDR of HCC cells.
Metformin may target the AMPK/mTOR/HIF-1α/P-gp and MRP1 pathways,
resulting in sensitization of the cells to combination
chemotherapies with other agents for the treatment of HCC. Future
studies may focus on the chemotherapy sensitization effect of
metformin with other chemotherapeutics in vivo and on the
elucidation of the precise mechanism underlying the cytotoxic and
MDR reversal effects of metformin in HCC.
Acknowledgements
This study was supported by grants from the National
Natural Science Foundation of China (nos. 81272368 and
81102069).
References
|
1
|
Jemal A, Bray F, Center MM, Ferlay J, Ward
E and Forman D: Global cancer statistics. CA Cancer J Clin.
61:69–90. 2011. View Article : Google Scholar
|
|
2
|
Carr BI: Hepatocellular carcinoma: current
management and future trends. Gastroenterology. 127:S218–S224.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Patt YZ, Hassan MM, Aguayo A, et al: Oral
capecitabine for the treatment of hepatocellular carcinoma,
cholangiocarcinoma and gallbladder carcinoma. Cancer. 101:578–586.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Lin DY, Lin SM and Liaw YF: Non-surgical
treatment of hepatocellular carcinoma. J Gastroenterol Hepatol.
12:S319–S328. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Ben Sahra I, Le Marchand-Brustel Y, Tanti
JF and Bost F: Metformin in cancer therapy: a new perspective for
an old antidiabetic drug? Mol Cancer Ther. 9:1092–1099. 2010.
|
|
6
|
Larsson O, Morita M, Topisirovic I, et al:
Distinct perturbation of the translatome by the antidiabetic drug
metformin. Proc Natl Acad Sci USA. 109:8977–8982. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Zoncu R, Efeyan A and Sabatini DM: mTOR:
from growth signal integration to cancer, diabetes and ageing. Nat
Rev Mol Cell Biol. 12:21–35. 2011. View
Article : Google Scholar : PubMed/NCBI
|
|
8
|
Wang N, Pan W, Zhu M, et al: Fangchinoline
induces autophagic cell death via p53/sestrin2/AMPK signalling in
human hepatocellular carcinoma cells. Br J Pharmacol. 164:731–742.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Zhang DM, Liu JS, Deng LJ, et al:
Arenobufagin, a natural bufadienolide from toad venom, induces
apoptosis and autophagy in human hepatocellular carcinoma cells
through inhibition of PI3K/Akt/mTOR pathway. Carcinogenesis.
34:1331–1342. 2013. View Article : Google Scholar
|
|
10
|
Cui XD, Lee MJ, Kim JH, et al: Activation
of mammalian target of rapamycin complex 1 (mTORC1) and Raf/Pyk2 by
growth factor-mediated Eph receptor 2 (EphA2) is required for
cholangiocarcinoma growth and metastasis. Hepatology. 57:2248–2260.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Nishikawa T, Takaoka M, Ohara T, et al:
Antiproliferative effect of a novel mTOR inhibitor temsirolimus
contributes to the prolonged survival of orthotopic esophageal
cancer-bearing mice. Cancer Biol Ther. 14:230–236. 2013. View Article : Google Scholar
|
|
12
|
Reungwetwattana T, Molina JR, Mandrekar
SJ, et al: Brief report: a phase II “window-of-opportunity”
frontline study of the MTOR inhibitor, temsirolimus given as a
single agent in patients with advanced NSCLC, an NCCTG study. J
Thorac Oncol. 7:919–922. 2012.
|
|
13
|
Grgic T, Mis L and Hammond JM: Everolimus:
a new mammalian target of rapamycin inhibitor for the treatment of
advanced renal cell carcinoma. Ann Pharmacother. 45:78–83. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Nakabayashi M, Werner L, Courtney KD, et
al: Phase II trial of RAD001 and bicalutamide for
castration-resistant prostate cancer. BJU Int. 110:1729–1735. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Sullivan R, Pare GC, Frederiksen LJ,
Semenza GL and Graham CH: Hypoxia-induced resistance to anticancer
drugs is associated with decreased senescence and requires
hypoxia-inducible factor-1 activity. Mol Cancer Ther. 7:1961–1973.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Kunz M and Ibrahim SM: Molecular responses
to hypoxia in tumor cells. Mol Cancer. 2:232003. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Zhang Y, Liu G, Lin C, Liao G and Tang B:
Silencing the EZH2 gene by RNA interference reverses the drug
resistance of human hepatic multidrug-resistant cancer cells to
5-Fu. Life Sci. 92:896–902. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Miller RA and Birnbaum MJ: An energetic
tale of AMPK-independent effects of metformin. J Clin Invest.
120:2267–2270. 2010. View
Article : Google Scholar : PubMed/NCBI
|
|
19
|
Xiong Y, Lu QJ, Zhao J and Wu GY:
Metformin inhibits growth of hepatocellular carcinoma cells by
inducing apoptosis via mitochondrion-mediated pathway. Asian Pac J
Cancer Prev. 13:3275–3279. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Zhu MM, Tong JL, Xu Q, et al: Increased
JNK1 signaling pathway is responsible for ABCG2-mediated multidrug
resistance in human colon cancer. PLoS One. 7:e417632012.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Thomas GV, Tran C, Mellinghoff IK, et al:
Hypoxia-inducible factor determines sensitivity to inhibitors of
mTOR in kidney cancer. Nat Med. 12:122–127. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Zhu H, Luo SF, Wang J, et al: Effect of
environmental factors on chemoresistance of HepG2 cells by
regulating hypoxia-inducible factor-1alpha. Chin Med J (Engl).
125:1095–1103. 2012.PubMed/NCBI
|
|
23
|
Jin WS, Kong ZL, Shen ZF, Jin YZ, Zhang WK
and Chen GF: Regulation of hypoxia inducible factor-1alpha
expression by the alteration of redox status in HepG2 cells. J Exp
Clin Cancer Res. 30:612011. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Takiyama Y, Harumi T, Watanabe J, et al:
Tubular injury in a rat model of type 2 diabetes is prevented by
metformin: a possible role of HIF-1alpha expression and oxygen
metabolism. Diabetes. 60:981–992. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Ece H, Cigdem E, Yuksel K, Ahmet D, Hakan
E and Oktay TM: Use of oral antidiabetic drugs (metformin and
pioglitazone) in diabetic patients with breast cancer: how does it
effect serum Hif-1 alpha and 8Ohdg levels? Asian Pac J Cancer Prev.
13:5143–5148. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Zannella V, Dal Pra A, Muaddi H, et al:
Reprogramming metabolism with metformin improves tumor oxygenation
and radiotherapy response. Clin Cancer Res. 19:6741–6750. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Sheng B, Liu J and Li GH: Metformin
preconditioning protects Daphnia pulex from lethal hypoxic
insult involving AMPK, HIF and mTOR signaling. Comp Biochem Physiol
B Biochem Mol Biol. 163:51–58. 2012.PubMed/NCBI
|
|
28
|
Li Y and Ye D: Cancer therapy by targeting
hypoxia-inducible factor-1. Curr Cancer Drug Targets. 10:782–796.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Rho JK, Choi YJ, Lee JK, et al: Gefitinib
circumvents hypoxia-induced drug resistance by the modulation of
HIF-1alpha. Oncol Rep. 21:801–807. 2009.PubMed/NCBI
|
|
30
|
Huang C, Xu D, Xia Q, Wang P, Rong C and
Su Y: Reversal of P-glycoprotein-mediated multidrug resistance of
human hepatic cancer cells by Astragaloside II. J Pharm Pharmacol.
64:1741–1750. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Ding Z, Yang L, Xie X, et al: Expression
and significance of hypoxia-inducible factor-1 alpha and
MDR1/P-glycoprotein in human colon carcinoma tissue and cells. J
Cancer Res Clin Oncol. 136:1697–1707. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Li DW, Dong P, Wang F, Chen XW, Xu CZ and
Zhou L: Hypoxia induced multidrug resistance of laryngeal cancer
cells via hypoxia-inducible factor-1alpha. Asian Pac J Cancer Prev.
14:4853–4858. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Chen M, Huang SL, Zhang XQ, et al:
Reversal effects of pantoprazole on multidrug resistance in human
gastric adenocarcinoma cells by down-regulating the
V-ATPases/mTOR/HIF-1alpha/P-gp and MRP1 signaling pathway in vitro
and in vivo. J Cell Biochem. 113:2474–2487. 2012. View Article : Google Scholar
|