Introduction
Hepatic fibrosis (HF) is recognized as one of the
most prevalent types of liver disease. Biologically, HF is defined
as the wound-healing process that occurs as a result of a wide
range of inflammatory reactions in the liver (1–3).
There are numerous enivronmental toxins for chronic liver disease,
including cholestasis, circulatory disturbances, autoimmune and
nutrition disorders, environmental toxins and the use of particular
medicine; however, the two primary causes of liver fibrosis have
been identified as infections caused by the hepatitis virus and
alcoholism (4). Regardless of its
origin, liver fibrosis is progressive and eventually leads to
cirrhosis or hepatocellular carcinoma, ultimately resulting in
organ failure and risk of mortality (5).
HF is characterized by the overabundant deposition
of extracellular matrix (ECM) proteins, composed mainly of type I
and type III collagen proteins. These excessive depositions disturb
the normal structure of the hepatic lobule, resulting in
misdirected blood flow through the liver, thereby impairing normal
organ functioning. This is the most salient feature of liver
cirrhosis (6).
Early in the progression of hepatic fibrosis, a
potent, fibrinogenic cytokine, transforming growth factor-β
(TGF-β), was demonstrated to be locally and systemically increased
in response to acute as well as chronic liver injury (7). TGF-β has been reported to trigger the
activation of hepatic stellate cells (HSCs), inducing their
differentiation into fibroblasts. This transformation has been
shown to be the principle determinant for the accumulation of
extracellular matrix proteins (8,9).
Activation of HSCs is therefore thought to be a key step in the
progression of hepatic fibrosis, justifying their use as a major
therapeutic target for the prevention and treatment of liver
cirrhosis (10). Due to the
connection between TGF-β and HSCs, therapeutic modalities that may
inhibit or reverse the action of either in order to prevent the
progression of hepatic fibrosis are the focus of present
studies.
Recent studies indicated that hepatic fibrosis is a
complex pathological process that involves various cytokines and
numerous cell signaling pathways (11–14).
TGF-β1 has been established as the crucial fibrogenic cytokine
promoting liver fibrosis, due to its activation of HSCs via the
TGF-β/Smad signaling pathway (15). In addition, it has been reported
that fibrosis has a dynamic bidirectional nature (16,17);
however, no effective therapies or medicine aimed at its reversal
are currently available, making their development necessary and
urgent.
Astragalus and Paeoniae radix rubra
extract (APE) is composed primarily of paeoniflorin, astragalosides
and curzenone extracted from a variety of traditional Chinese herbs
(e.g. Astragali radix, Paeoniae radix rubra,
Curcumae rhizoma, Bupleuri radix and
Eupolyphaga), the majority of which have a long history as
remedies for the treatment of chronic liver disease. For example,
the use of Astragali radix in Chinese Medicine as a tonic
herb is used alone or in conjunction with other herbs for the
treatment of liver diseases. Pharmacological and clinical studies
have demonstrated its hepatoprotective and other beneficial effects
(18,19). The primary components of
Astragali radix, astragalosides, were found to significantly
inhibit the progression of CCl4-induced hepatic fibrosis
in vivo as well as inhibit the proliferation of
TGF-β1-stimulated HSCs in vitro (20). Another herb, Paeoniae radix
rubra, has also been considered to be potent for the treatment for
liver diseases. Paeoniae radix rubra extracts have been
reported to reduce CCl4-induced liver fibrosis in rats
as well as platelet-derived growth factor-stimulated hepatic
stellate cell migration (21).
APE was prepared from extracts of Astragali
radix, Paeoniae radix rubra, Curcumae rhizoma,
Bupleuri radix and Eupolyphaga in the standard ratio
30:30:15:12:10 as measured by crude herbal weight. Previous studies
have demonstrated that APE had protective effects on
chemically-induced, acute liver injury in mice through the
inhibition oxidative stress. In order to further investigate the
anti-fibrotic activity of APE, the present study was designed to
characterize the effects and elucidate the mechanism of APE
treatment on CCl4-induced liver fibrosis in rats.
Materials and methods
Preparation of APE
The following herbs were purchased from The Xi’an
Chinese Medicine Corporation, (Xi’an, China): Astragali
radix, Paeoniae radix rubra, Curcumae rhizoma,
Bupleuri radix and Eupolyphaga. All herbs used were
identified by Dr Genquan Qiu, a specialist in Traditional Chinese
Herbal Medicine and contributing author of the present study. Herb
samples were preserved in the specimen room of the Institute of
Clinical Pharmacology at Xi’an Medical College (Xi’an, China). APE
components of the five herbs were extracted and prepared as
follows:
A total of 8.45 kg of dried, sliced crude herbs
(Astragali radix, Paeoniae radix rubra,
Curcumae rhizoma, Bupleuri radix and
Eupolyphaga) were prepared at a standard ratio of
30:30:15:12:10. Samples were decocted three times using 80 l water
at 95°C for 35 min each time. The decocted solution was filtered
though a150-μm gauze (Abcam, Cambridge, UK) and the filtrate was
then concentrated to a mass of 4.22 kg (density, 1.225) using a
vacuum desiccator (5530000; Labconoco, Kansas City, MO, USA) at
70°C. The sediment was dried into power using a spray drier (WD645;
Titanium Industries, Inc., Taipei, Taiwan) at 80–160°C. This
process yielded 2.11 kg dry powder. It should be noted that in all
cellular in vitro experiments, APE powder was dissolved in
Hank’s solution at a ratio of 1:50 (H9494, Sigma-Aldrich).
Animals and treatment groups
Male Sprague-Dawley rats were obtained from the
Experimental Animal Center (Anhui Medical University, Hefei, China)
and had a weight range of 160–200g. All animals were housed in
plastic cages a room temperature of 22±1°C, relative humidity of
50±20% and under a 12-h light/dark cycle. The studies were
performed in accordance with the guidelines for the humane
treatment of animals as set forth by the Association of Laboratory
Animal Sciences and the Center for Laboratory Animal Sciences at
Anhui Medical University. This study was approved by the ethics
committee of Anhui Medical University (2011-002 SCXK; Hefei,
China)
Chronic liver injury was induced using
CCl4 injections as previously described (22–24).
Rats were divided at random into six groups (n=10 per group). The
control subjects were allowed ad libitum access to food and
water. All other groups were administered an intraperitoneal
injection of 1.0 ml/kg CCl4 in an olive oil vehicle (2:3
v/v) mixture, twice a week for eight weeks. The positive control
group was intragastrically (i.g.) administrated colchicine (0.1
mg/kg; C3915; Sigma-Aldrich, St. Louis, MO, USA), a clinically used
treatment for acute gout and other immunological diseases (25). Treatment groups received daily i.g.
doses of APE, with each group receiving either 1.3, 2.6 or 5.2
g/kg. Oral i.g. administration of either colchicine or APE occurred
on the day prior to CCl4 treatment. All subjects were
weighed once per week throughout the duration of the experiment and
the drug dose was adjusted to the body weight for each
administration.
The rats were anesthetized with diethyl ether and
sacrificed via cervical dislocation following the final injection
of CCl4. Blood samples were then collected from the
abdominal aorta. Samples were centrifuged for ten minutes (3,000 ×
g, 4°C) and serum was collected and then frozen at −80°C. Following
sacrificing of the animals, the liver, spleen and kidney were
promptly removed and weighed. A portion of the liver was fixed with
paraffin for histopathology and the remaining tissue was stored at
−80°C until further use.
Analysis of liver function
The serum activities of aspartate aminotransferase
(AST) and alanine aminotransferase (ALT) were determined using
spectrophotometry with an Olympus AU 600 Autoanalyzer (Alternative
Biomedical Solutions, Dallas, Texas, USA) and commercially
available alanine aminotransferase assay and aspartate
aminotransferase assay kits (Jiancheng Institute of Biotechnology,
Nanjing, China). All absorbances were read at 505 nm and the enzyme
activity was calculated as U/l.
Activity measurements of antioxidants,
antioxidant enzyme activity and malondialdehyde (MDA)
Liver homogenate (10%, w/v) was prepared by
homogenizing the liver tissue on ice in 150 mM Tris-HCl buffered
saline (pH 7.2; Sigma-Aldrich) using a polytron homogenizer
(PT3100D; Kinematical, Lucerne, Switzerland). A commercially
available kit (Jiancheng Biological Engineering Research Institute,
Nanjing, China) was used to determine the activities of SOD and
GSH-Px according to the manufacturer’s instructions. Data are
expressed as SOD U/mg protein and GSH-Px mg/g protein (26). MDA levels in liver tissues were
determined using the thiobarbituric acid method, provided by a
commercially available kit (Jiancheng Biological Engineering
Research Institute) and measured according to the manufacturer’s
instructions.
Measurement of fibrotic markers and serum
levels of TGF-β1
Levels of hexadecenoic acid (HA), laminin (LN) and
procollagen type III (PCIII) were assayed using a radioimmunoassay
method with a kit obtained from Beijing North Institute of
Biotechnology (Beijing, China). Hydroxyproline (hyp) and TGF-β1
serum levels were determined using an ELISA method using a kit
obtained from Sigma-Aldrich. All absorbances were read at 450 nm
using a microplate reader (FlexStation 3; Molecular Devices,
Sunnyvale, CA, USA).
Histopathological evaluation
Liver tissue was removed immediately following the
sacrification of the animals and fixed in 10% formalin, then
embedded in paraffin. Hematoxylin and eosin (HE) staining (E4283;
Sigma-Aldrich) was performed to measure the degree of liver injury,
while Masson staining (HT15; Sigma-Aldrich) was used to detect
collagen deposition. Each staining method was performed according
to the manufacturer’s instructions. Histopathological evaluation
was performed by an expert pathologist, blinded to the treatment
identity of each tissue sample. The degree of liver fibrosis was
categorized into five groups according to the following scoring
system: 0, no fibrosis, normal liver and absence of fibrosis; I,
fibrosis present (collagen fibers present that extend from the
portal triad or central vein to the peripheral region); II, mild
fibrosis (mild collagen fibers present with extension without
compartment formation); III, moderate fibrosis (moderate collagen
fibers present with a certain level of pseudo-lobe formation); or
IV, severe fibrosis (severe collagen fibers present with thickening
of the partial compartments and frequent pseudo-lobe formation)
(27). The degree of fibrosis was
expressed as the mean of ten fields sampled from each slide. The
final numerical score was calculated as follows: The sum of the
number per grade of affected rats, divided by the total number of
samples.
Immunoblot analysis
Liver tissue samples were homogenized in extraction
buffer (25 mmol 4-(2-hydroxyethyl)-1-piperazineethanesulfonic acid,
400 mmol KCl, 1 mmol EDTA and 1.5 mmol MgCl2)
supplemented with protease and phosphatase inhibitors
(Sigma-Aldrich) (1 mmol/l phenylmethyl sulfonyl fluoride, 0.1
mmol/l N-tosyl-L-phenylalanine chloromethyl ketone, 1 mg/ml
aprotinin, 1 mg/ml pepstatin, 0.5 g/ml leupeptin, 1 mmol/l NaF, 1
mmol/l Na4P2O4 and 2 mmol/l
Na3VO4). The extract was centrifuged at
14,000 × g for 10 min at 4°C and resulting supernatants were boiled
for five min in SDS sample buffer [100 mmol/l Tris-HCl (pH 6.8), 4%
SDS, 12% β-mercaptoethanol, 20% glycerol and 0.01% bromophenol
blue] (Sigma-Aldrich). Samples were subjected to SDS-PAGE and
transferred onto polyvinylidene difluoride (PVDF) membranes
(Millipore, Bedford, MA, USA). The membrane was blocked overnight
at 4°C by immersing in TBST-20 buffer, containing 10 mmol/l
Tris-HCl, 150 mmol/l NaCl, 0.08% Tween 20 and 10% non-fat dry milk.
PVDF membranes were then incubated with primary antibodies
overnight at 4°C, followed by the appropriate secondary antibody
for 2 h at room temperature. The primary antibodies used were as
follows: Mouse anti-human monoclonal antibodies against collagen
types I and III were purchased from Abcam (CAT: ab6308 and ab6310,
Cambridge, UK), mouse anti-human monoclonal antibodies against
TGF-β1 (CAT: sc-130348), mouse anti-Smad7 antibody (CAT: sc-365846)
and rabbit/mouse anti-β-actin antibody (CAT: sc-7210 and sc-8432)
were purchased from Santa Cruz Biotechnology (Santa Cruz, CA, USA).
Rabbit anti-human phospho-Smad2 antibody (CAT: BS4172) and rabbit
antihuman Smad2 antibody (CAT: BS2993) were purchased from Bioworld
Technology (St. Louis, MO, USA). Rabbit anti-human phospho-Smad3
antibody (CAT: 9520), rabbit anti-human Smad3 antibody (CAT: 9513),
rabbit anti-Smad4 (CAT: 9515) and rabbit anti-α-SMA antibody (CAT:
A2522) were purchased from Cell Signaling Technology (Boston, MA,
USA). The secondary antibodies used were goat anti-rabbit IgG and
goat anti-mouse IgG (43413; 42472; Sigma-Aldrich).
The membranes were washed three times with TBST-20
buffer, immunoreactive proteins were detected using enhanced
chemiluminescence medium (RPN 2106; Amersham Pharmacia Biotech,
Piscataway, NJ, USA) and visualized by autoradiography (Bio-Rad,
Hercules, CA, USA). Blots were then quantified via densitometric
analysis using Quantity One version 4.52 software and a GelDoc XR
(Bio-Rad).
Statistical analyses
Quantitative data are expressed as mean ± standard
deviation. The Student’s t-test was used to compare between groups
and the Mann-Whitney rank sum test was used for the degree of
histopathological liver injury. All tests were performed using SPSS
version 13.0 software (SPSS, Inc., Chicago, IL, USA). P<0.05 was
considered to indicate a statistically significant difference
between values.
Results
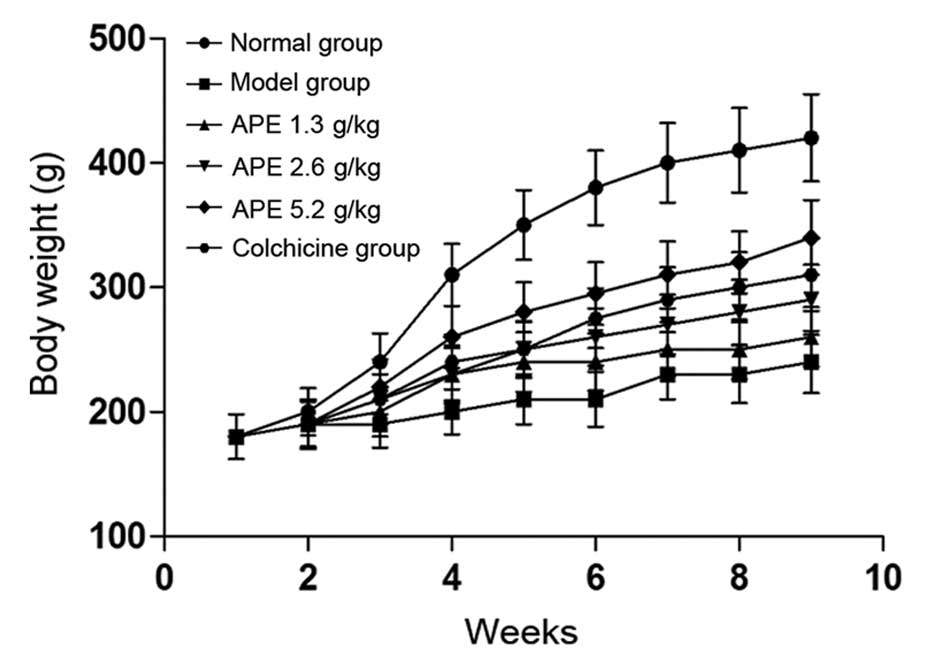
APE mitigates the decrease in body weight
induced by CCl4 treatment
In comparison to the normal group, the weight of
rats in the model group and the CCl4-treated groups was
markedly decreased (P<0.05) (Fig.
1). The weight of animals receiving APE treatments (2.6 and 5.2
g/kg) significantly increased in a dose-dependent manner
(P<0.05), compared to that of the model group. However, the
lowest APE dose (1.3 g/kg) and colchicine (0.1 mg/kg), used as a
positive control, had no significant effect on ameliorating
CCl4-induced weight loss (P>0.05).
APE treatment protects against
CCl4-induced increases in liver and spleen coefficients,
but not kidney coefficients
Liver, spleen and kidney coefficients were compared
to organ coefficients obtained from rats with
CCl4-induced fibrosis. As shown in Table I, the liver and spleen coefficients
were significantly increased when compared to those of the control
groups. More specifically, treatment with higher doses of APE (2.6
and 5.2 g/kg) markedly reduced the liver and spleen coefficients
following CCl4 treatment (P<0.05). In the positive
control group, colchicine (0.1 mg/kg) also reduced the liver
coefficient (P<0.05); however, it showed a protective effect
against the increased spleen coefficient following CCl4
treatment (P>0.05). No significant differences were observed in
kidney coefficients among all treatment groups when compared to
those of the controls (Table
I).
 | Table IEffect of APE on organ coefficients
in rats with CCl4-induced hepatic fibrosis (n=10). |
Table I
Effect of APE on organ coefficients
in rats with CCl4-induced hepatic fibrosis (n=10).
| Groups | Dose (g/kg) | Liver coefficient
(%) | Spleen coefficient
(%) | Kidney coefficient
(%) |
|---|
| Normal | — | 2.64±0.22 | 0.24±0.06 | 0.68±0.07 |
| Model | — | 4.65±0.41a | 0.38±0.07a | 0.69±0.11 |
| APE | 1.3 | 4.33±0.45 | 0.34±0.05 | 0.69±0.08 |
| 2.6 | 4.21±0.40b | 0.31±0.05b | 0.65±0.13 |
| 5.2 | 4.13±0.38c | 0.30±0.04c | 0.67±0.11 |
| Colchicine | 0.1 | 4.26±0.37b | 0.32±0.06 | 0.58±0.09 |
APE treatment mitigates
CCl4-induced increases in serum AST and ALT levels
Serum ALT and AST activities were evaluated in order
to measure the extent of liver injury following chronic
CCl4 treatment. Serum levels of ALT and AST
post-CCl4 treatment were increased three and four-fold,
respectively, when compared to those of the control group
(P<0.01) (Table II). Of note,
high doses of APE (2.6 and 5.2 g/kg) and colchicine (0.1 mg/kg)
demonstrated a significantly protective effect against the increase
of serum ALT and AST following long-term treatment with
CCl4 (P<0.05) (Table
II).
| Table IIEffect of APE on liver function,
hydroxyproline content and production of TGF-β1 in rats with
CCl4-induced hepatic fibrosis (n=10). |
Table II
Effect of APE on liver function,
hydroxyproline content and production of TGF-β1 in rats with
CCl4-induced hepatic fibrosis (n=10).
| Group | Dose (mg/kg) | ALT (U/l) | AST (U/l) | Hydroxyproline
(mg/g) |
|---|
| Normal | — | 42.62±10.36 | 36.49±9.88 | 1.08±0.29 |
| Model | — |
126.53±31.59a |
116.93±28.47a | 2.96±0.85a |
| APE | 1.3 | 109.48±20.86 | 98.86.46±18.53 | 2.54±0.58 |
| 2.6 | 91.42±18.28b | 89.36±19.48b | 2.18±0.48b |
| 5.2 | 86.94±20.84b | 81.75±18.47b | 2.08±0.36c |
| Colchicine | 0.1 | 96.58±18.93b | 90.86±21.61b | 2.24±0.34b |
Effect of APE on levels of MDA, SOD and
GSH-Px in hepatic tissue
GSH-Px and SOD have been reported to be capable of
scavenging the toxic CCl4 metabolite lipid peroxide
(28); therefore, levels of GSH-Px
and SOD were measured in hepatic tissue samples from rats in order
to determine the effect of APE on these compounds. As shown in
Table III, the levels of GSH-Px
and SOD were significantly decreased in the model group compared
with those of the control group (P<0.01). Treatment with high
doses of APE (2.6 and 5.2 g/kg) and colchicine (0.1 mg/kg)
mitigated CCl4-induced GSH-Px and SOD depletion
(P<0.05) (Table III).
Following treatment with low doses of APE (1.3 g/kg), levels of
GSH-Px and SOD were slightly, but not significantly, higher than
those of animals administered CCl4 only.
| Table IIIEffect of APE on MDA levels, and SOD
and GSH-Px activities in liver homogenates of
CCl4-induced liver fibrosis in rats (n=10). |
Table III
Effect of APE on MDA levels, and SOD
and GSH-Px activities in liver homogenates of
CCl4-induced liver fibrosis in rats (n=10).
| Group | Dose (g/kg) | GSH-Px (U/mg) | SOD (U/mg) | MDA (μmol/g) |
|---|
| Normal | — | 397.85±54.58 | 247.62±41.82 | 42.16±9.86 |
| Model | — |
215.51±42.43a |
146.83±28.68a | 71.83±11.35a |
| APE | 1.3 | 246.25±31.46 | 161.52±30.49 | 62.83±12.63 |
| 2.6 |
265.31±32.46b |
183.64±29.34b | 56.83±12.64b |
| 5.2 |
297.61±34.56c |
195.39±32.94b | 52.61±13.48b |
| Colchicine | 0.1 |
271.68±31.83b |
174.22±28.57b | 58.23±15.61 |
In addition, MDA, a marker for lipid peroxidation
levels, was significantly increased in rats with hepatic fibrosis
compared with those of the model group (P<0.01). Of note, APE
treatment (2.6 and 5.2 g/kg) resulted in an observable decrease in
MDA levels (P<0.05). However, the colchicine group (0.1 mg/kg)
showed no significant difference in MDA levels when compared with
those of the model group (Table
III).
APE administration decreases serum HA, LN
and PCIII and hydroxyproline levels in CCl4-induced
fibrotic livers
As shown in Table
IV, serum levels of three markers of liver fibrosis, HA, LN and
PC III, were markedly increased in hepatic fibrotic rats when
compared to those of the control group (P<0.01) (Table IV). Administration of APE (2.6 and
5.2 g/kg) or colchicine (0.1 mg/kg) effectively decreased
CCl4-induced serum levels of HA, LN and PC III
(P<0.05; P<0.01) (Table
IV).
| Table IVEffect of APE on serum HA, LN and
PCIII levels in rats with CCl4-induced liver fibrosis
(n=10). |
Table IV
Effect of APE on serum HA, LN and
PCIII levels in rats with CCl4-induced liver fibrosis
(n=10).
| Group | Dose (mg/kg) | HA (μg/l) | LN (μg/l) | PCIII (μg/l) |
|---|
| Normal | — | 106.38±27.69 | 116.95±36.56 | 108.62±28.64 |
| Model | — |
256.83±52.68a |
297.62±48.31a |
286.34±45.29a |
| APE | 1.3 | 221.96±38.94 | 264.83±41.66 | 257.34±46.92 |
| 2.6 |
208.53±38.42b |
238.18±38.71b |
228.67±42.31b |
| 5.2 |
195.21±39.48c |
224.73±34.16c |
208.71±43.53c |
| Colchicine | 0.1 |
211.62±37.49c |
240.68±46.48a |
248.57±42.16b |
APE reduces hapatic hydroxyproline, an
index of liver fibrosis
Following long-term treatment with CCl4,
rats were found to have increased levels of hydroxyproline when
compared with those of the control group (P<0.01) (Table II). Treatment with APE (2.6 and
5.2 g/kg) resulted in significantly reduced levels of
hydroxyproline in liver tissue when compared with those of the
model group (P<0.05; P<0.01) (Table II). Colchicine (0.1 mg/kg) had an
identical effect to that of high doses of APE on hydroxyproline
levels (P<0.05) (Table
II).
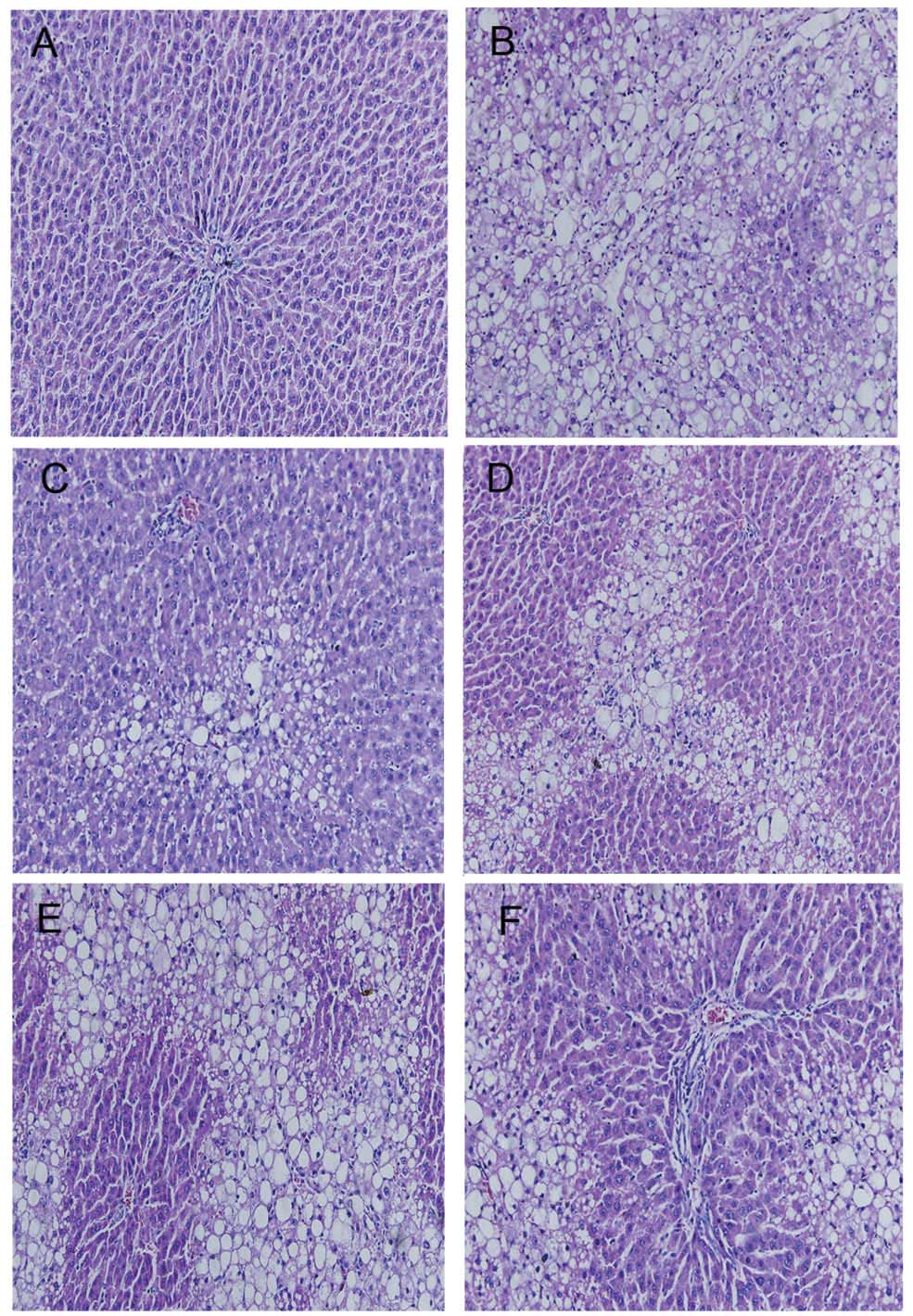
APE results in improved liver
histopathology
HE staining indicated that CCl4
administration resulted in extensive morphological changes to the
animals’ livers, including marked fatty acid degeneration,
necrosis, hepatocyte ballooning and infiltration of inflammatory
cells into the interstitial space of the livers. Liver tissue from
the control group tissue showed normal lobular architecture with
central veins and radiating hepatic cords (Fig. 2A). The histological pattern of
tissue of subjects treated with APE (1.3, 2.6 and 5.2 g/kg) showed
a lesser degree of liver injury and inflammation (Fig. 2C–E). A similar trend of reduced
hepatic injury and inflammation was also observed in the tissue of
colchicine-treated rats (Fig.
2F).
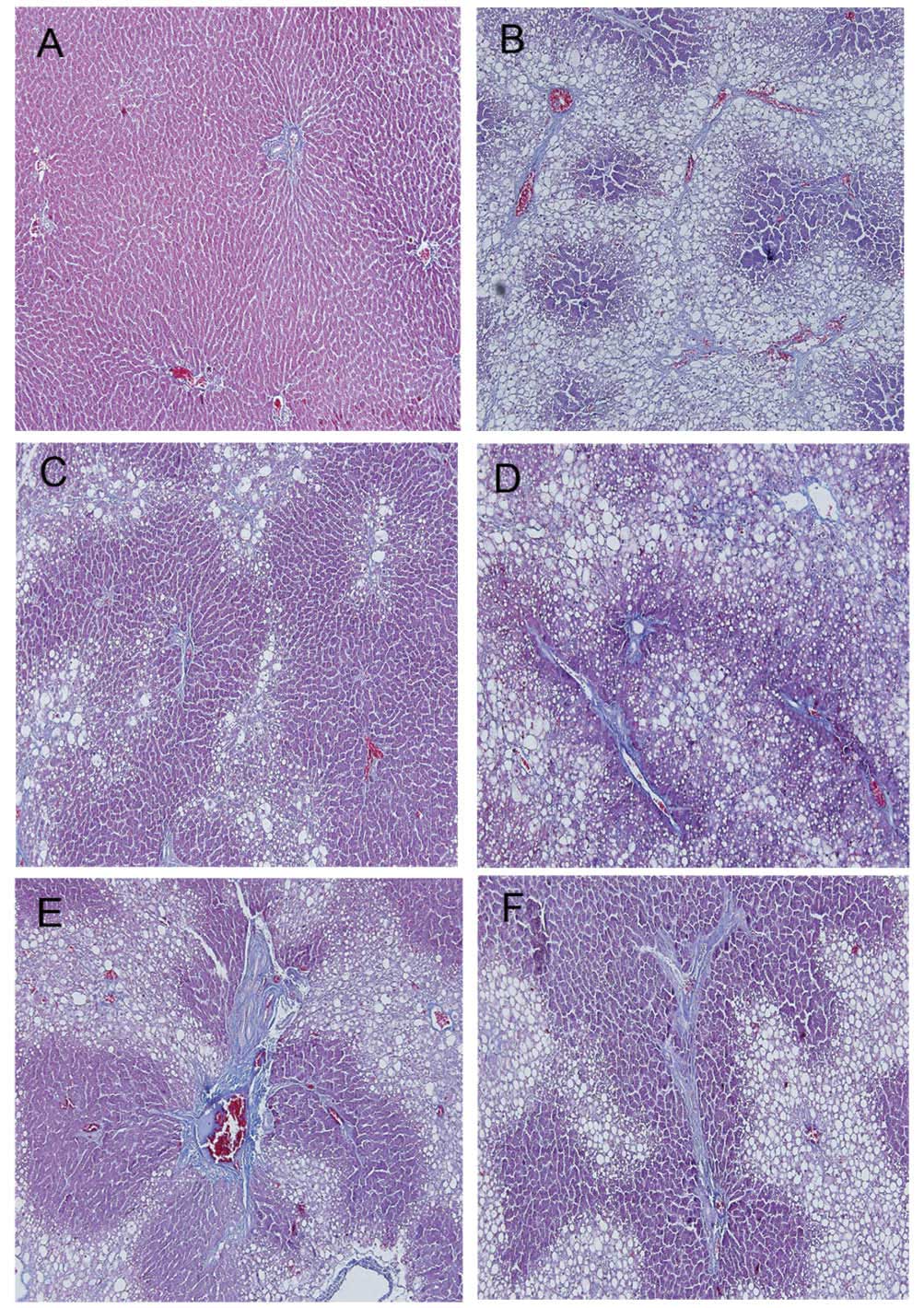
Masson staining was used to detect histopathological
changes that resulted from CCl4-induced fibrosis
(Fig. 3). The livers of rats
treated with CCl4 showed extensive accumulation of
connective tissues resulting in the formation of continuous
fibrotic septa, nodules of regeneration and noticeable alterations
to the central vein, compared to those of the healthy controls
(Fig. 3A and B). APE- (1.3, 2.6
and 5.2 g/kg), and colchicine- (0.1 mg/kg) treated tissues showed
reduced collagen deposition and fewer pseudolobule formation
(Fig. 3C–F). Microscopic
examination revealed that APE markedly reduced the grade of liver
fibrosis and improved CCl4-induced hepatic fibrosis
(P<0.05) (Table V).
| Table VEffect of APE on the pathological
grading of CCl4-induced liver fibrosis in rats. |
Table V
Effect of APE on the pathological
grading of CCl4-induced liver fibrosis in rats.
| | Pathological
grading of hepatic fibrosis | |
|---|
| |
| |
|---|
| Group | Dose (mg/kg) | 0 | I | II | III | IV | P-value |
|---|
| Normal | — | 10 | 0 | 0 | 0 | 0 | — |
| Model | — | 0 | 0 | 1 | 4 | 6 | 0.000a |
| APE | 1.3 | 0 | 1 | 2 | 3 | 4 | 0.467 |
| 2.6 | 0 | 2 | 4 | 2 | 2 | 0.031b |
| 5.2 | 1 | 4 | 2 | 3 | 0 | 0.014b |
| Colchicine | 0.1 | 0 | 1 | 3 | 3 | 2 | 0.029b |
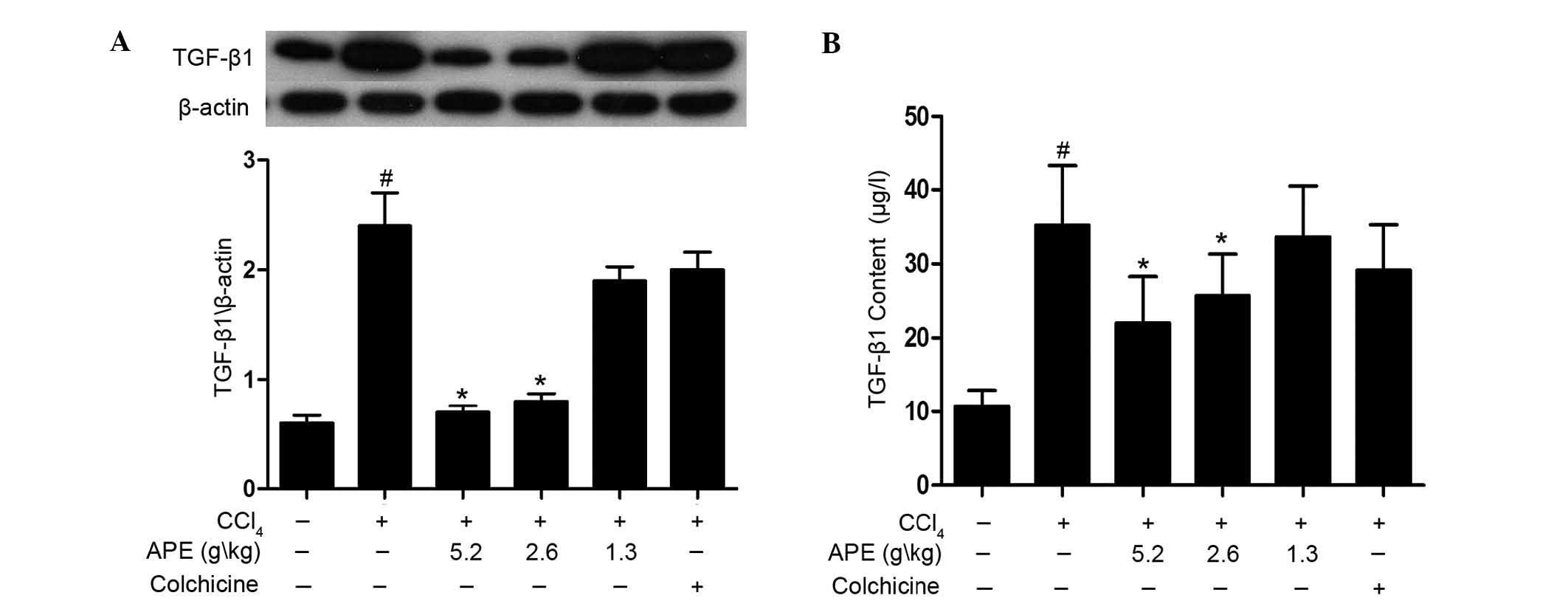
High doses of APE reduce TGF-β1 levels in
rats with hepatic fibrosis
The production of TGF-β1 was investigated using
western blot analysis and ELISA assay. As shown in Fig. 4A, western blot analysis revealed
that levels of TGF-β1 in hepatic fibrosis tissue were significantly
elevated when compared to those of the normal control group
(P<0.05). In addition, APE treatment (2.6 and 5.2 g/kg)
significantly decreased TGF-β1 levels when compared to those of the
CCl4-only control group (P<0.05). However, the
low-dose APE group (1.3 g/kg) and colchicine (0.1 mg/kg) group
showed no statistically significant differences to the
CCl4-only control group. The ELISA assay also revealed
that APE treatment (2.6 and 5.2 g/kg) reduced levels of TGF-β1 in
serum (P<0.05) (Fig. 4B). These
results indicated that high doses of APE (2.6 and 5.2 g/kg) have
the capacity to mitigate increased levels of TGF-β1 in rats with
CCl4-induced hepatic fibrosis.
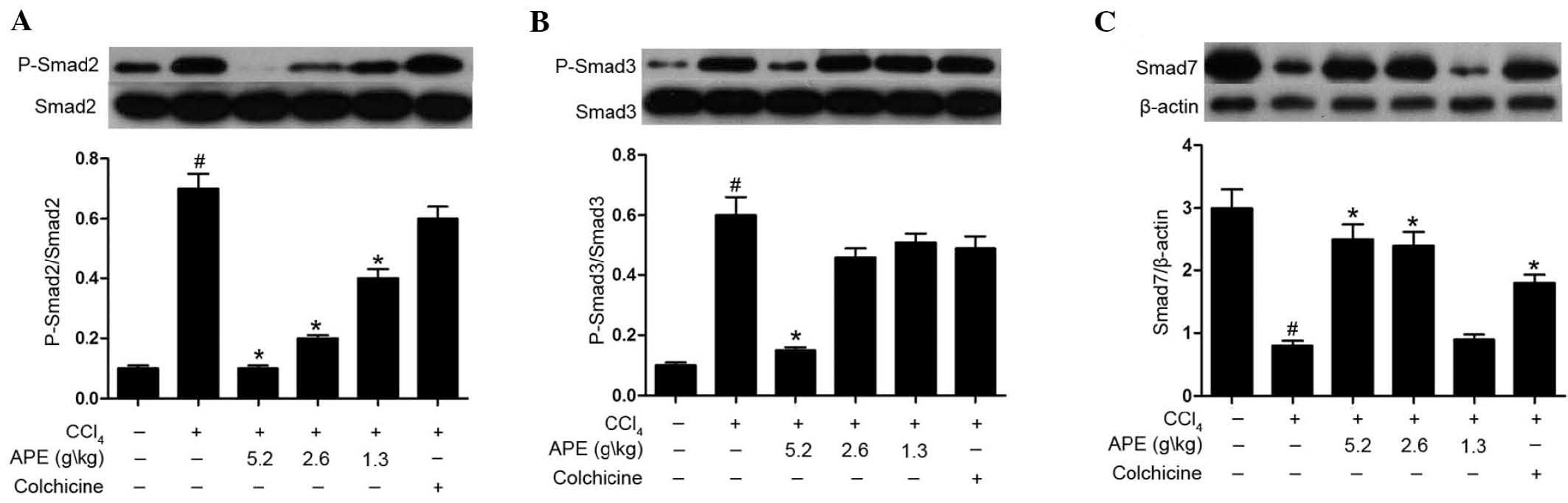
APE decreases Smad2/3 phosphorylation and
Smad7 expression in the liver tissue of rats with
CCl4-induced hepatic fibrosis
As shown in Fig. 5,
CCl4-induced hepatic fibrosis caused increased
phosphorylation of Smad2 and Smad3 when compared to that of the
controls. APE treatment (5.2 g/kg) significantly decreased Smad2
and Smad3 phosphorylation in liver tissue of rats with hepatic
fibrosis (P<0.05). Lower doses of APE (1.3 and 2.6 g/kg) also
downregulated the phosphorylation of Smad2, but had no significant
effect on Smad3 phosphorylation (P<0.05). Similarly, colchicine
treatment (0.1 mg/kg) had no significant effect on the
phosphorylation of Smad2 or Smad3 (Fig. 5A and B).
Levels of Smad7 expression in the model group were
markedly suppressed compared to those of the normal group
(P<0.05). By contrast, APE (2.6 and 5.2 g/kg) and colchicine
treatments (0.1 mg/kg) significantly increased levels of Smad7
expression compared to those of the model group (P<0.05).
In conclusion, these results revealed that high
doses of APE (2.6 and 5.2 g/kg) prevented CCl4-induced
fibrosis. The results also indicated that the mechanism of action
of APE proceeded through preventing the phosphorylation of Smad2/3
and enhancing the inhibition of Smad7 expression in the TGF-β/Smad
signaling pathway.
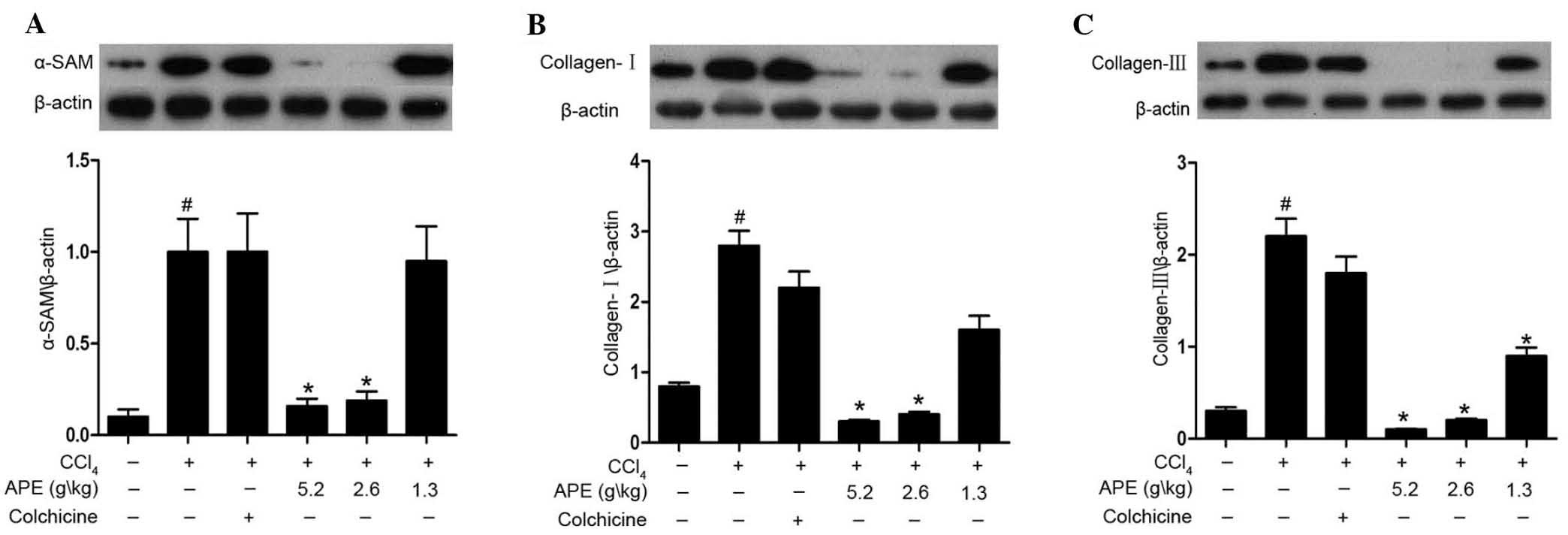
APE decreases α-SAM, collagen I and
collagen III expression in liver tissue of rates with
CCl4-induced hepatic fibrosis
Western blot analysis revealed increased protein
expression of α-SAM, collagen I and collagen III in
CCl4-induced hepatic fibrotic tissue when compared to
expression levels in normal control rats (Fig. 6). APE treatment (2.6 and 5.2 g/kg)
significantly decreased α-SAM, collagen I and collagen III
expression compared to expression levels in the model group.
Moreover, the low dose of APE (1.3 g/kg) was also able to decrease
collagen III expression (P<0.05). However, no significant
reductions were observed following treatment with colchicine (0.1
mg/kg).
Discussion
It is well documented that HF is a common feature in
numerous chronic hepatic diseases and that constant fibrosis may
lead to the development of hepatocellular carcinoma (29). However, HF is only a small part of
a dynamic cascade that begins with hepatocyte necrosis. Following
necrosis, HSCs are activated and proliferate, with the concurrent
release of fibrogenic transmitters or factors (30). However, studies have demonstrated
that interrupting or reversing the molecular pathways involved in
hepatic fibrosis may be promising for future therapies (31,32).
Previous studies have shown that APE had protective effects against
acute liver injury in murine models. To further investigate the
effects of APE on liver fibrosis, the present study used the
well-characterized CCl4-induced hepatic fibrosis model
(33). Liver injury has been
reported to induce the release of two aminotransferases, AST and
ALT, into the circulatory system (34). These proteins are therefore used as
conventional indicators of hepatic trauma. Throughout the present
study, body weight, organ coefficients and the levels of ALT and
AST were assessed to evaluate the extent and degree of liver injury
in rats undergoing long-term CCl4 treatment. The results
indicated that high doses of APE (2.6 and 5.2 g/kg) reduced
inflammation and alleviated CCl4-induced liver injury.
In addition, histopathological examinations using HE staining
provided further evidence for the hepatoprotective effect of APE in
rats.
HF is characterized by excessive deposition of ECM
components, including collagen protein, proteoglycan and osamine
protein (35). Therefore,
administration of APE may protect the liver against hepatic injury
by reducing collagen deposition. HA, LN and PC III are essential
components of the ECM and, due to their abnormally increased
expression in liver fibrosis, are conducive biomarkers for hepatic
fibrogenesis (36,37). Increased expression of hepatic hyp
is another liver index that represents the degree of HF (38). In the present study, the levels of
serum HA, LN, PC III and the content of hyp in hepatic tissue were
significantly increased in CCl4-treated rats, whereas
APE treatment (2.6 and 5.2 g/kg) markedly decreased the levels of
serum HA, LN, PC III and the content of hyp in hepatic tissue.
These significant results strongly suggested the presence of
inherent hepatoprotective effects of APE. Furthermore,
histopathological observations provided additional confirmation
that the severity of CCl4-induced liver fibrosis was
largely ameliorated by treatment with APE.
Oxidative stress has an important role in the
generation of CCl4-induced liver fibrosis (39). CCl4 damages the
hepatocellular membrane via lipid peroxidation, which is followed
by the release of inflammatory mediators from activated
inflammatory cells. The release of these mediators is thought to
potentiate CCl4-induced hepatic injury (28). Numerous antioxidants have been
shown to have therapeutic or protective effects against liver
injury (40,41). The concentration of the main
product of lipid peroxidation, MDA, is generally assayed as the
total levels of lipid peroxidation products (42). SOD and GSH-Px are key enzymes in
the antioxidant defense system and their mechanism of action
involes catalysis of the transformation of hydrogen peroxide into
water (43). In the present study,
CCl4-treated rats demonstrated elevated levels of MDA
and decreased activity of SOD and GSH-Px. However, treatment with
APE (2.6 and 5.2 g/kg) reduced the amount of MDA and increased the
activities of SOD and GSH-Px. This therefore indicates that APE may
have an anti-oxidative role in hepatic fibrosis through recovery of
the organism’s natural anti-oxidative defense system.
HSCs are a major type of fibrogenic liver cell found
during liver injury and have are responsible for the progression of
hepatic fibrosis (44). TGF-β1 has
been suggested to be an important factor in activating and
promoting the transformation of HSCs (45). The present study demonstrated that
TGF-β1 was highly expressed in CCl4-treated rats and
administration of APE resulted in a significant reduction of TGF-β1
levels.
TGF-β1 controls a diverse set of cellular processes
and its canonical signaling is mediated via TGF-β-induced
phosphorylation of receptor-activated Smad2 and Smad3 (46). Smad7 is the negative feedback
regulator for TGF-β signaling, acting to antagonize the activity of
the receptor-regulated Smads, which leads to the termination of
TGF-β signaling (47). The present
study showed that APE significantly inhibited the phosphorylation
of Smad2 and Smad3 and reversed the inhibitory effect of
CCl4 treatment on Smad7 expression. Therefore, it was
hypothesized that the anti-fibrotic effects of APE occur via the
TGF-β1/Smad pathway.
Following long-term CCl4-stimulation,
HSCs become activated and transdifferentiate into myofibroblasts
(MFBs). MFBs are characterized by numerous fibrotic functions,
including the induction of ECM deposition, α-SMA expression as well
as the synthesis and secretion of type I and type III collagen
(48). Western blot analysis
performed in the present study revealed that CCl4
enhanced the expression of α-SMA, type I and type III collagen,
while the administration of APE prevented fibrosis development
through inhibition of HSCs.
In conclusion, the present study demonstrated that
APE was effective in preventing necro-inflammation and fibrogenesis
in CCl4-induced hepatic fibrosis. Of note, the
hepatoprotective effects of APE appear to be associated with the
inhibition of lipid peroxidation and decreases in TGF-β1 levels.
However, further studies are required to determine whether APE
exerts these protective effects in human subjects in vivo
and whether it has an effect on other types of fibrogenesis. In
addition, the exact mechanism and pharmacological actions of APE
for clinical usage remain to be elucidated.
Acknowledgements
The authors would like to thank Miss Yin Zhou and
Mr. Xiaopeng Tian for their technical assistance and the Department
of Pathology at Peking University Shen Zhen Hospital for providing
and processing the samples. This study was supported by research
grants from the Nature Science Foundation of Xi’an Medical College
(no. L12C01).
References
|
1
|
Handa P and Kowdley KV: Chemokines: potent
mediators of hepatic inflammation and fibrosis in chronic liver
diseases. Ann Hepatol. 13:152–154. 2013.
|
|
2
|
Puche JE, Saiman Y and Friedman SL:
Hepatic stellate cells and liver fibrosis. Compr Physiol.
3:1473–1492. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Mera K, Uto H, Mawatari S, Ido A,
Yoshimine Y, Nosaki T, Oda K, Tabu K, Kumagai K, Tamai T, et al:
Serum levels of apoptosis inhibitor of macrophage are associated
with hepatic fibrosis in patients with chronic hepatitis C. BMC
Gastroenterol. 14:272014. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Friedman SL: Liver fibrosis in 2012:
convergent pathways that cause hepatic fibrosis in NASH. Nat Rev
Gastroenterol Hepatol. 10:71–72. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Bataller R and Brenner DA: Liver fibrosis.
J Clin Invest. 115:209–218. 2005. View
Article : Google Scholar : PubMed/NCBI
|
|
6
|
Tsukada S, Parsons CJ and Rippe RA:
Mechanisms of liver fibrosis. Clin Chim Acta. 364:33–60. 2006.
View Article : Google Scholar
|
|
7
|
Tahashi Y, Matsuzaki K, Date M, Yoshida K,
Furukawa F, Sugano Y, Matsushita M, Himeno Y, Inagaki Y and Inoue
K: Differential regulation of TGF-beta signal in hepatic stellate
cells between acute and chronic rat liver injury. Hepatology.
35:49–61. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Gabriel A, Ziólkowski A, Radlowski P,
Tomaszek K and Dziambor A: Hepatocyte steatosis in HCV patients
promotes fibrosis by enhancing TGF-beta liver expression. Hepatol
Res. 38:141–146. 2008.
|
|
9
|
Ray K: Liver: Hepatic stellate cells hold
the key to liver fibrosis. Nat Rev Gastroenterol Hepatol.
11:742014. View Article : Google Scholar
|
|
10
|
Cheng K, Yang N and Mahato RI: TGF-beta1
gene silencing for treating liver fibrosis. Mol Pharm. 6:772–779.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Qian H, Shi J, Fan TT, Lv J, Chen SW, Song
CY, Zheng ZW, Xie WF and Chen YX: Sophocarpine attenuates liver
fibrosis by inhibiting the TLR4 signaling pathway in rats. World J
Gastroenterol. 20:1822–1832. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Wu BR, Zheng YL, Sang XL, Jin M, Wang WZ,
Zhang QS, Zhao SX and Kong L: Role of the IGF-1/PI3K pathway and
the molecular mechanism of Fuzhenghuayu therapy in a spontaneous
recovery rat model of liver fibrosis. Chinese Journal of
Hepatology. 21:674–678. 2013.(In Chinese).
|
|
13
|
Ren WG, Kong LB, Mi HM, Zhao SX, Zhang YG,
Wang RQ and Nan YM: Activation of Fas/FasL and its downstream
signaling pathway promotes development of alcoholic steatohepatitis
and liver fibrosis in mice. Chinese Journal of Hepatology.
21:129–133. 2013.(In Chinese).
|
|
14
|
Zhai X, Yan K, Fan J, Niu M, Zhou Q and
Zhou Y, Chen H and Zhou Y: The beta-catenin pathway contributes to
the effects of leptin on SREBP-1c expression in rat hepatic
stellate cells and liver fibrosis. Br J Pharmacol. 169:197–212.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Lee JH, Lee H, Joung YK, Jung KH, Choi JH,
Lee DH, Park KD and Hong SS: The use of low molecular weight
heparin-pluronic nanogels to impede liver fibrosis by inhibition
the TGF-beta/Smad signaling pathway. Biomaterials. 32:1438–1445.
2011. View Article : Google Scholar
|
|
16
|
Czaja AJ: Review article: The prevention
and reversal of hepatic fibrosis in autoimmune hepatitis. Aliment
Pharmacol Ther. 39:385–406. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Pellicoro A, Ramachandran P and Iredale
JP: Reversibility of liver fibrosis. Fibrogenesis Tissue Repair.
5(Suppl 1): S262012.PubMed/NCBI
|
|
18
|
Liang XL and Yuan JY: Effect of Chinese
herbal compound on liver fibrosis in rabbits with schistosomiasis
by B-ultrasound. Asian Pac J Trop Med. 6:658–662. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Lv J, Zhao Z, Chen Y, Wang Q, Tao Y, Yang
L, Fan TP and Liu C: The chinese herbal decoction danggui buxue
tang inhibits angiogenesis in a rat model of liver fibrosis. Evid
Based Complement Alternat Med. 2012:2849632012. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Gui SY, Wei W, Wang H, Wu L, Sun WY, Chen
WB and Wu CY: Effects and mechanisms of crude astragalosides
fraction on liver fibrosis in rats. J Ethnopharmacol. 103:154–159.
2006. View Article : Google Scholar
|
|
21
|
Li CX, Li L, Lou J, Yang WX, Lei TW, Li
YH, Liu J, Cheng ML and Huang LH: The protective effects of
traditional Chinese medicine prescription, han-dan-gan-le, on
CCl4-induced liver fibrosis in rats. Am J Chin Med. 26:325–332.
1998. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Brattin WJ, Glende EA Jr and Recknagel RO:
Pathological mechanisms in carbon tetrachloride hepatotoxicity. J
Free Radic Biol Med. 1:27–38. 1985. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Nadkarni GD and D’Souza NB: Hepatic
antioxidant enzymes and lipid peroxidation in carbon
tetrachloride-induced liver cirrhosis in rats. Biochem Med Metab
Biol. 40:42–45. 1988. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Gassó M, Rubio M, Varela G, Cabré M,
Caballería J, Alonso E, Deulofem R, Camps J, Giménez A, Pajares M,
et al: Effects of S-adenosylmethionine on lipid peroxidation and
liver fibrogenesis in carbon tetrachloride-induced cirrhosis. J
Hepatol. 25:200–205. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Tian XP, Yin YY and Li X: Effects and
mechanisms of Acremoniumterricola milleretal mycelium on liver
fibrosis induced by carbon tetrachloride in rats. Am J Chin Med.
39:537–550. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Sid B, Verrax J and Calderon PB: Role of
oxidative stress in the pathogenesis of alcohol-induced liver
disease. Free Radic Res. 47:894–904. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Ruwart MJ, Wilkinson KF, Rush BD, Vidmar
TJ, Peters KM, Henley KS, Appelman HD, Kim KY, Schuppan D and Hahn
EG: The integrated value of serum procollagen III peptide over time
predicts hepatic hydroxyproline content and stainable collagen in a
model of dietary cirrhosis in the rat. Hepatology. 10:801–806.
1989. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Gebhardt R: Oxidative stress,
plant-derived antioxidants and liver fibrosis. Planta Med.
68:289–296. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Wallace MC and Friedman SL: Hepatic
fibrosis and the microenvironment: fertile soil for hepatocellular
carcinoma development. Gene Expr. 16:77–84. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Zhao G, Zhang ZQ, Zhang B, Luo M, Sun YW
and Wu ZY: Down-regulation of tTG expression by RNAi inhibits HSC
proliferation and attenuates liver fibrosis. Int J Clin Exp Pathol.
4:513–520. 2011.PubMed/NCBI
|
|
31
|
Song YH, Chen XL, Kong XJ, Liu NZ, Li W,
Wu XL, Lin JS and Jin YX: Ribozymes against TGFbeta1 reverse
character of activated hepatic stellate cells in vitro and inhibit
liver fibrosis in rats. J Gene Med. 7:965–976. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Nanji AA, Zakim D, Rahemtulla A, Daly T,
Miao L, Zhao S, Khwaja S, Tahan SR and Dannenberg SR: Dietary
saturated fatty acids down-regulate cyclooxygenase-2 and tumor
necrosis factor alfa and reverse fibrosis in alcohol-induced liver
disease in the rat. Hepatology. 26:1538–1545. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Hou G, Dick R and Brewer GJ: Improvement
in dissolution of liver fibrosis in an animal model by
tetrathiomolybdate. Exp Biol Med (Maywood). 234:662–665. 2009.
View Article : Google Scholar
|
|
34
|
Guéchot J, Boisson RC, Zarski JP, Sturm N,
Calès P and Lasnier E: AST/ALT ratio is not an index of liver
fibrosis in chronic hepatitis C when aminotransferase activities
are determinate according to the international recommendations.
Clin Res Hepatol Gastroenterol. 37:467–472. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Su X, Wang Y, Zhou G, Yang X, Yu R, Lin Y
and Zheng C: Probucol attenuates ethanol-induced liver fibrosis in
rats by inhibiting oxidative stress, extracellular matrix protein
accumulation and cytokine production. Clin Exp Pharmacol Physiol.
41:73–80. 2014. View Article : Google Scholar
|
|
36
|
Xu HG, Fang JP, Huang SL, Li HG, Zhong FY,
Guo HX and Su H: Diagnostic values of serum levels of HA, PC III, C
IV and LN to the liver fibrosis in children with beta-thalassemia
major. Chinese Journal of Pediatrics. 41:603–606. 2003.(In
Chinese).
|
|
37
|
Kaneda H, Hashimoto E, Yatsuji S,
Tokushige K and Shiratori K: Hyaluronic acid levels can predict
severe fibrosis and platelet counts can predict cirrhosis in
patients with nonalcoholic fatty liver disease. J Gastroenterol
Hepatol. 21:1459–1465. 2006.PubMed/NCBI
|
|
38
|
Dang SS, Wang BF, Cheng YA, Song P, Liu ZG
and Li ZF: Inhibitory effects of saikosaponin-d on CCl4-induced
hepatic fibrogenesis in rats. World J Gastroenterol. 13:557–563.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Hsu CL, Hsu CC and Yen GC:
Hepatoprotection by freshwater clam extract against CCl4-induced
hepatic damage in rats. Am J Chin Med. 38:881–894. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Koca SS, Bahcecioglu IH, Poyrazoglu OK,
Ozercan IH, Sahin K and Ustundag B: The treatment with antibody of
TNF-alpha reduces the inflammation, necrosis and fibrosis in the
non-alcoholic steatohepatitis induced by methionine- and
choline-deficient diet. Inflammation. 31:91–98. 2008. View Article : Google Scholar
|
|
41
|
Wu SJ, Tam KW, Tsai YH, Chang CC and Chao
JC: Curcumin and saikosaponin a inhibit chemical-induced liver
inflammation and fibrosis in rats. Am J Chin Med. 38:99–111. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Koh YH, Park YS, Takahashi M, Suzuki K and
Taniguchi N: Aldehyde reductase gene expression by lipid
peroxidation end products, MDA and HNE. Free Radic Res. 33:739–746.
2000. View Article : Google Scholar
|
|
43
|
Djordjevic A, Spasic S, Jovanovic-Galovic
A, Djordjevic R and Grubor-Lajsic G: Oxidative stress in diabetic
pregnancy: SOD, CAT and GSH-Px activity and lipid peroxidation
products. J Matern Fetal Neonatal Med. 16:367–372. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Park KC, Park JH, Jeon JY, Kim SY, Kim JM,
Lim CY, Lee TH, Kim HK, Lee HG, Kim SM, et al: HNHA, new histone
deacetylase inhibitor, improves liver fibrosis in BDL rats through
suppression of hepatic stellate cells. Br J Pharmacol. 2014.(Epub
ahead of print). View Article : Google Scholar
|
|
45
|
Kang KH, Qian ZJ, Ryu B, Karadeniz F, Kim
D and Kim SK: hepatic fibrosis inhibitory effect of peptides
isolated from navicula incerta on tgf-beta1 induced activation of
lx-2 human hepatic stellate cells. Prev Nutr Food Sci. 18:124–132.
2013. View Article : Google Scholar
|
|
46
|
Meng XM, Huang XR, Chung AC, Qin W, Shao
X, Igarashi P, Ju W, Bottinger EP and Lan HY: Smad2 protects
against TGF-beta/Smad3-mediated renal fibrosis. J Am Soc Nephrol.
21:1477–1487. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
del Pilar Alatorre-Carranza M,
Miranda-Díaz A, Yañez-Sánchez I, Pizano-Martínez O,
Hermosillo-Sandoval JM, Vázquez-Del Mercado M, Hernández-Hoyos S,
Martínez-Abundis R, Fafutis-Morris M, Segura-Ortega J and
Delgado-Rizo V: Liver fibrosis secondary to bile duct injury:
correlation of Smad7 with TGF-beta and extracellular matrix
proteins. BMC Gastroenterol. 9:812009. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Brenner DA, Kisseleva T, Scholten D, Paik
YH, Iwaisako K, Inokuchi S, Schnabl B, Seki E, De Minicis S,
Oesterreicher C and Taura K: Origin of myofibroblasts in liver
fibrosis. Fibrogenesis Tissue Repair. 5(Suppl 1): S172012.
View Article : Google Scholar : PubMed/NCBI
|