Introduction
Osteosarcoma is the most common primary bone
malignancy, which occurs most frequently in late childhood and
early adulthood (1). In the last
25 years, the introduction of dose intensive combination
chemotherapy has resulted in a five-year tumor-free survival rate
of 60–75% (2). However, in
patients with metastasis and local recurrence, prognosis remains
poor (3). Cancer stem cells (CSCs)
are a small subset of cells within malignant tumors that possess
stem cell-like properties, and are responsible for cancer
origination and progression (4–6).
Further research on the genetic network that controls the CSC-like
characteristics may provide novel strategies in cancer
treatment.
Recently, a growing number of long non-coding RNAs
(lncRNAs) have been identified as crucial regulators in various
types of cancer (7–9). Furthermore, other lncRNAs have been
shown to be associated with stem cell properties, such as
pluripotency and differentiation (10). However, lncRNAs associated with
CSCs have yet to be reported. Through bioinformatics analysis using
public data (http://genome.ucsc.edu/), the
possible cis- regulatory association between the lncRNA-mRNA pairs
were predicted and the genes transcribed within a 10 kbp window
upstream or downstream of lncRNAs were considered as cis target
genes. If one cis target gene of an lncRNA was also a known
cancer-associated mRNA, the lncRNA-mRNA pair was selected for the
experiments in the present study. An lncRNA, TCONS_00004241, which
is located at 2p21, on the antisense side of the promoter upstream
region of a cancer-associated mRNA, hypoxia-inducible factor-2α
(HIF-2α) was identified. This class of lncRNAs are known as
promoter upstream transcripts (PROMPTs), which appear to regulate
the transcriptional activity of host PCGs (11–14).
Therefore, the lncRNA TCONS_00004241 was termed HIF-2α promoter
upstream transcript (HIF2PUT). Transcript factors chip-seq data
from encode/analysis (http://genome.ucsc.edu/) also demonstrated that many
cancer-associated transcription factors interact with the HIF2PUT
locus, particularly at the promoter region, which HIF2PUT shares
with its mRNA partner, HIF-2α.
HIF-2α has previously been demonstrated to be
associated with stem cell-like properties in stem cells, and in the
CSCs of numerous types of cancer (15–18);
therefore the present study aimed to explore the possible function
of HIF2PUT in osteosarcoma CSCs, and the association with its host
PCG/HIF-2α.
Materials and methods
Patient samples
Patients with osteosarcoma, who underwent initial
surgery at the Chinese PLA General Hospital between 2012 and 2013,
were retrospectively selected for the present study (Table I). Prior to resection none of the
patients had received therapy. A total of 17 pairs of osteosarcoma
samples and adjacent tissues were used. The use of tumor material
for research was approved by the Ethical Committee of PLA General
Hospital (Beijing, China). Written informed consent was obtained
from the patient/the patient’s family.
 | Table ICharacteristics of the patients
participating in the present study. |
Table I
Characteristics of the patients
participating in the present study.
| Number | Age | Sex | Histological
diagnosis | Stage | Tumour location |
|---|
| 1 | 25 | F | osteosarcoma | II A | Distal end of
femur |
| 2 | 20 | M | osteosarcoma | II B | Proximal end of
tibia |
| 3 | 14 | M | osteosarcoma | II B | Proximal end of
humerus |
| 4 | 28 | M | osteosarcoma | II B | Distal end of
tibia |
| 5 | 8 | M | osteosarcoma | I B | Distal end of
femur |
| 6 | 15 | M | osteosarcoma | II A | Proximal end of
tibia |
| 7 | 14 | F | osteosarcoma | I B | Distal end of
femur |
| 8 | 10 | M | osteosarcoma | I A | Distal end of
femur |
| 9 | 15 | M | osteosarcoma | II A | Proximal end of
tibia |
| 10 | 15 | F | osteosarcoma | I B | Distal end of
femur |
| 11 | 11 | M | osteosarcoma | II A | Distal end of
femur |
| 12 | 11 | M | osteosarcoma | II A | Proximal end of
tibia |
| 13 | 12 | F | osteosarcoma | II B | Proximal end of
tibia |
| 14 | 18 | M | osteosarcoma | II A | Distal end of
femur |
| 15 | 12 | M | osteosarcoma | II B | Proximal end of
tibia |
| 16 | 21 | F | osteosarcoma | I B | Distal end of
femur |
| 17 | 19 | M | osteosarcoma | II A | Proximal end of
tibia |
Cancer cell lines
The SAOS2, MG63, and U2OS human osteosarcoma cell
lines were purchased from American Type Culture Collection
(Manassas, VA, USA). The OS-732 osteosarcoma cell line was obtained
from the Cell Culture Center of Peking Union Medical College
(Beijing, China). The cells were cultured in Dulbecco’s modified
Eagle’s medium (DMEM; Gibco Life Technologies, Carlsbad, CA, USA),
supplemented with 10% fetal bovine serum (Invitrogen Life
Technologies, Carlsbad, CA, USA), at 37°C in an atmosphere
containing 5% CO2.
Quantitative polymerase chain reaction
(qPCR)
Total RNA was isolated from the osteosarcoma tumor
tissues, matched adjacent normal tissues and osteosarcoma cells
using TRIzol® reagent (Invitrogen Life Technologies).
cDNA synthesis was performed with 2 μg total RNA using the
RevertAid™ H Minus First Strand cDNA Synthesis kit (Takara Bio,
Inc., Otsu, Japan). The primers were obtained from Sheng Gong
(Shanghai, China), and the sequences are included in Table II. A qPCR was performed using the
SYBR® PrimeScript RT-PCR kit (Takara Bio, Inc.) and the
Applied Biosystems 7500 Fluorescent Quantitative PCR system
(Applied Biosystems Life Technologies, Foster City, CA, USA). The
reaction mixtures were incubated at 95°C for 30 sec, followed by 40
amplification cycles of 95°C for 5 sec and 60°C for 34 sec. The
comparative cycle threshold method was used to quantify the
relative expression levels of mRNA and lncRNA (19). Expression levels of the
housekeeping gene β-actin were used to normalize the expression
levels of the genes-of-interest. The expression levels of the genes
were calculated in each patient using the following ratio: target
in cancerous tissue/target in non-cancerous tissue [R(C/N)].
| Table IIPrimers for quantitative polymerase
chain reaction analysis. |
Table II
Primers for quantitative polymerase
chain reaction analysis.
| Gene | Forward | Reverse |
|---|
| β-actin |
5′-CCACTGGCATCGTGATGGA-3′ |
5′-CGCTCGGTGAGGATCTTCAT-3′ |
| HIF2PUT |
5′-CGGAGGTGTTCTATGAGCTGG-3′ |
5′-AGCTTGTGTGTTCGCAGGAA-3′ |
| HIF-2α |
5′-TGGGATCTAACAGGAACAGC-3′ |
5′-CTAAATAGCCAGACAAGGGT-3′ |
Overexpression of HIF2PUT
The cDNA-HIF2PUT plasmid was constructed by
introducing a BamHI-EcoRI fragment containing HIF2PUT
cDNA, into the same site in a pcDNA3.1 vector, which was obtained
from GenePharma Ltd. (Shanghai, China). MG63 cells, which have low
expression levels of HIF2PUT, were transfected with pcDNA-HIF2PUT
using Lipofectamine® 2000 (Invitrogen Life
Technologies), according to the manufacturer’s instructions. The
cells were collected following transfection for RNA isolation, and
cell proliferation, scratch wound healing invasion and spheroid
formation assays.
Transfection of siRNA
For gene knockdown analysis, small interfering
(si)RNAs targeting the lncRNA-HIF2PUT sequence, and non-targeting
siRNA, were obtained from GenePharma Co., Ltd. (Shanghai, China).
The siRNAs used in the present study had the following sequences:
HIF2PUT siRNA1, sense 5′-CCUGCCACAUGCCUUAUCUTT-3′, and antisense
5′-AGAUAAGGCAUGUGGCAGGTT-3′; and HIF2PUT siRNA2, sense
5′-GUCUAUAUCUCUCCCUUUATT-3′, and antisense
5′-UAAAGGGAGAGAUAUAGACTT-3′. Approximately 5% MG63 cells were
plated in each well of 12-well plates, at least 24 h prior to
transfection, in order to achieve 30–50% confluence. siRNA
transfection was performed using the X-tremeGENE Transfection
reagent (Roche Diagnostics, Basel, Switzerland), according to the
manufacturer’s instructions. The cells were collected following
transfection for RNA isolation, and cell proliferation, scratch
wound healing, invasion and spheroid formation assays.
Flow cytometry (FCM)
The expression rate of osteosarcoma stem cell
surface marker CD133 was analyzed by fluorescence-activated cell
sorting (FACS) using a FACSVantage™ flow cytometer (BD Biosciences,
San Jose, CA, USA). Following treatment with FcR Blocking reagent
(Miltenyi Biotec, Bergisch Gladbach, Germany), the MG63 cells were
incubated with phycoerythrin-conjugated anti-CD133 (Miltenyi
Biotec) at 4°C for 15 min. Dead cells were eliminated using 50
μg/ml propidium iodide (EMD Millipore, Billerica, MA, USA). The
labeled cells were then analyzed and separated by FCM, and the data
were analyzed using CellQuest™ software (BD Biosciences). Gating
was implemented based on negative-control staining profiles.
Cell proliferation assay
Following transfection, cell proliferation was
assessed using an MTS assay (Promega Corporation, Madison, WI,
USA), according to the manufacturer’s instructions. MG63 cells
(2×103 cells/well) from each group were plated into a
96-well plate. A total of 20 μl MTS reagent was added to each well
containing 100 μl culture media. The plate was incubated for 2 h at
37°C in a humidified atmosphere containing 5% CO2. The
optical density of the samples was then measured at a wavelength of
490 nm using a plate reader (Molecular Devices, Sunnyvale, CA,
USA).
Spheroid formation assay
The self-renewing capabilities of the cells were
assessed using ultra-low attachment surface 96-well culture dishes
(Corning, Inc., Corning, NY, USA). The cells were re-suspended in
200 μl serum-free medium DMEM/F12 (Gibco Life Technologies)
supplemented with 20 ng/ml human epidermal growth factor
(Peprotech, Inc., Rocky Hill, NJ, USA), 20 ng/ml human basic
fibroblast growth factor (Peprotech, Inc.) and 1% N-2 supplement
(Gibco Life Technologies), at a density of 200 cells in each well.
Phase-contrast images were captured seven days later using an
Olympus fluorescence microscope (Olympus, Tokyo, Japan).
Scratch wound healing assay
Uniform wounds were scraped across plated MG63
cells, which had been grown on plastic six-well plates, using a
pipette tip, prior to transfection. The initial gap length (0 h)
and the residual gap length (48 h) after wounding were calculated
from photomicrographs using an Olympus fluorescence microscope
(Olympus).
Matrigel™ invasion assay
A cell invasion assay was carried out using a
modified Boyden Chamber consisting of
Transwell®-precoated Matrigel™ membrane filter inserts,
with 8 mm pores in 24-well tissue culture plates (BD Biosciences).
Minimum Essential Medium supplemented with 10% fetal bovine serum
was added to the lower chamber, and served as the
chemoattractant.
Statistical analysis
Differences between the groups were analyzed using
Student’s t-test. Correlations between gene expression levels were
studied using Pearson’s correlation. Statistical analyses were
performed using SPSS version 18.0 (SPSS, Inc., Chicago, IL, USA).
For all statistical analyses, P<0.05 was considered to indicate
a statistically significant difference.
Results
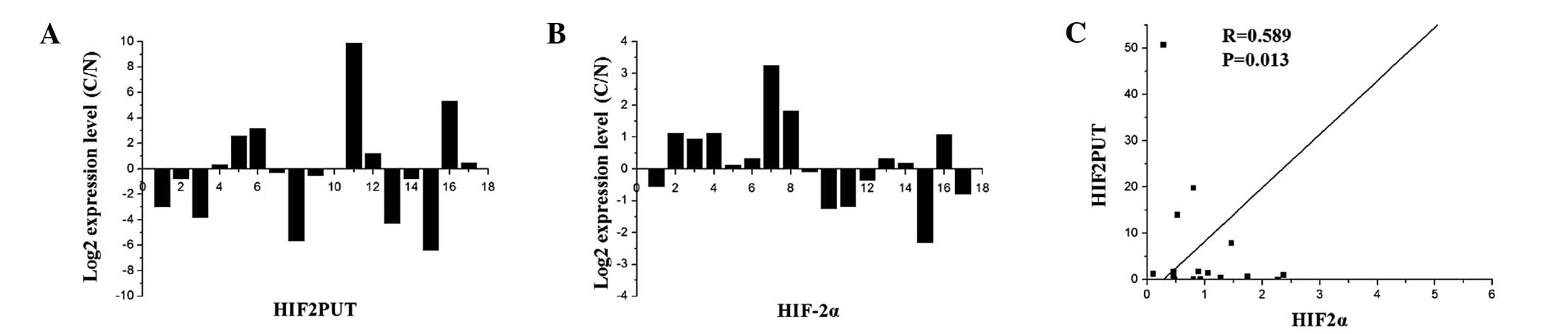
Expression of HIF2PUT is correlated with
HIF-2α in osteosarcoma
HIF2PUT and HIF-2α expression levels were assessed
in a group of 17 patients with osteosarcoma. For each patient, RNA
was isolated from cancerous and adjacent non-tumorous osteosarcoma
tissues. The expression levels of HIF2PUT were significantly
correlated with those of HIF-2α in the osteosarcoma tissue samples
(R=0.589, P<0.05, Fig.
1A–C).
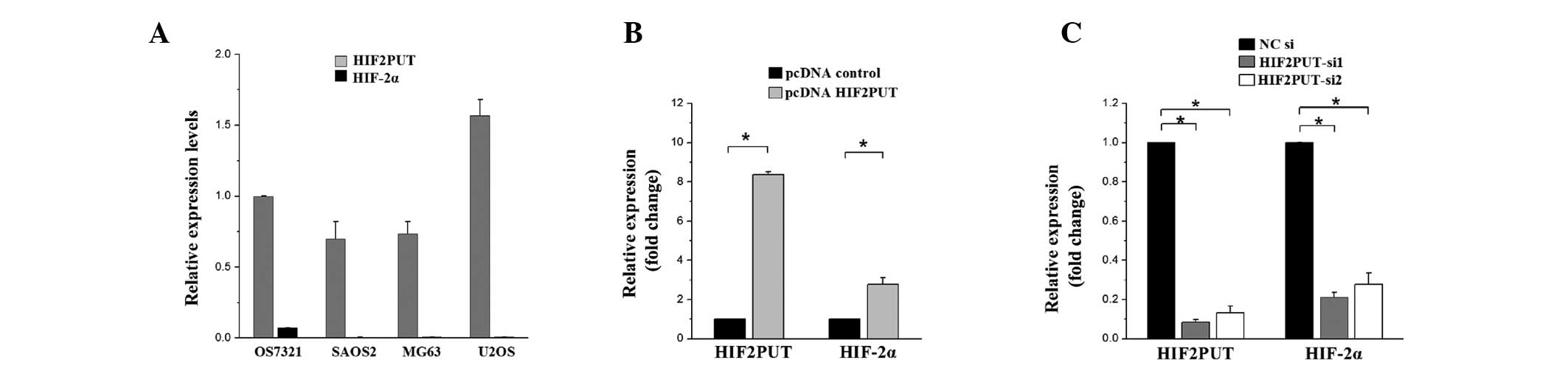
HIF-2α mRNA expression was co-regulated with the
lncRNA HIF2PUT overexpression or knockdown in the MG63 cells.
HIF2PUT and HIF-2α expression levels were analyzed by qPCR in four
osteosarcoma cell lines. The expression levels of HIF-2α were low
in all four cell lines (Fig. 2A),
whereas the expression levels of HIF2PUT were notably higher. The
MG63 cells were chosen as the candidate cell line for the
subsequent HIF2PUT overexpression and knockdown experiments, since
the expression levels of HIF2PUT in this cell line were relatively
moderate compared with the other cell lines. In the overexpression
experiment, a HIF2PUT cDNA plasmid was constructed and stably
transfected into the MG63 cells. lncRNA HIF2PUT expression levels
were markedly upregulated, and HIF-2α mRNA expression levels were
also moderately elevated (Fig.
2B). In the knockdown experiment, transfection of the cells
with HIF2PUT siRNA resulted in a significant downregulation of
HIF2PUT expression levels, and HIF-2α mRNA expression levels were
also markedly decreased (Fig.
2C).
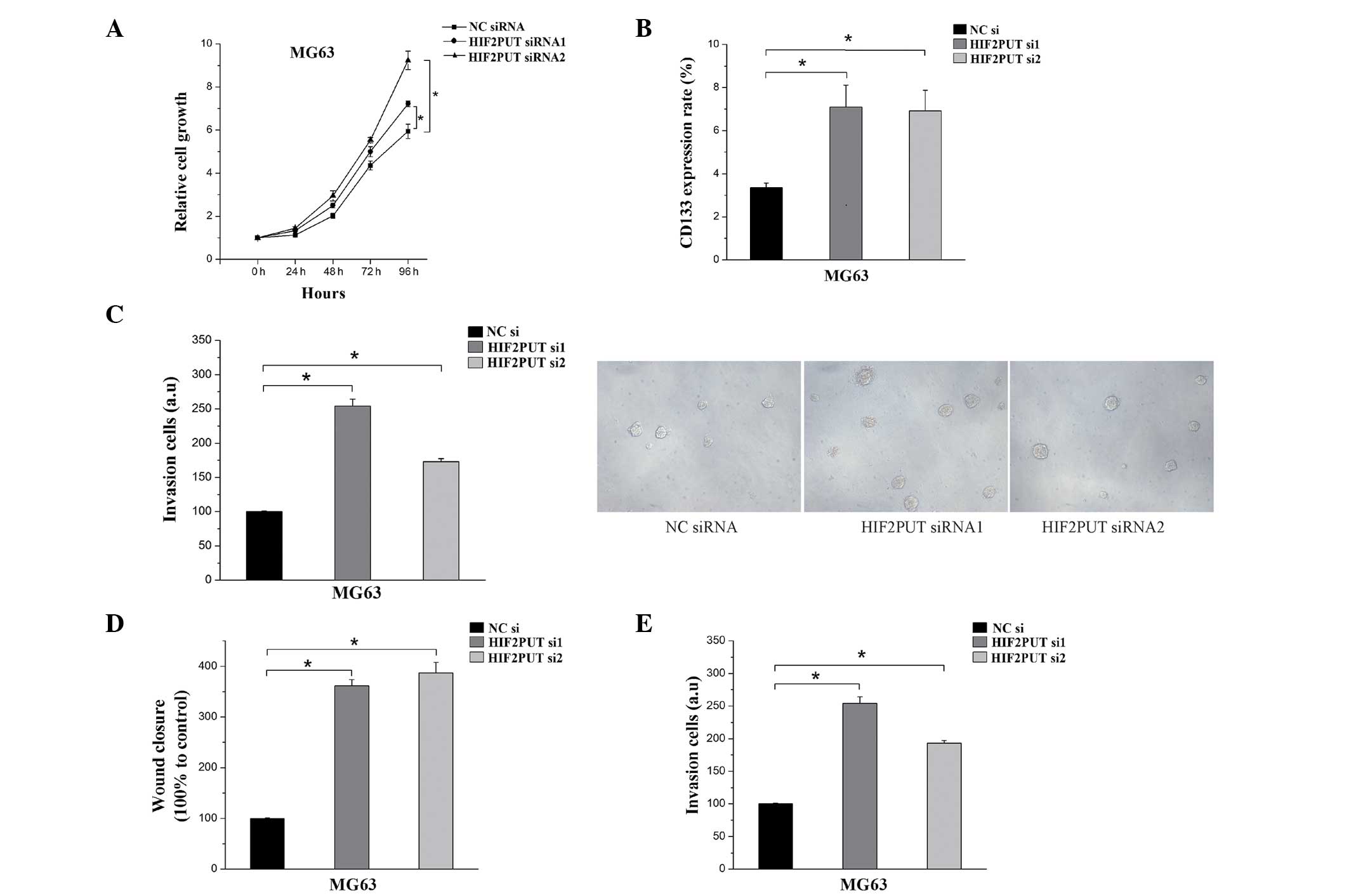
Transfection with HIF2PUT siRNA enhances
the proliferation, migration and self-renewal of MG63 cells
The proliferation of MG63 cells was evidently
accelerated following knockdown of HIF2PUT expression (Fig. 3A), as determined by MTS assay. The
FCM analysis demonstrated that the expression rate of the stem cell
marker CD133 was significantly elevated in the MG63 cells following
transfection with HIF2PUT siRNA (Fig.
3B). These results coincided with an increased rate of
spheroid-formation in the stem cell culturing media (Fig. 3C). To further investigate the
effects of HIF2PUT on cell migration and invasion, scratch wound
healing and Matrigel™ invasion assays were performed on the MG63
cells in vitro. The wound healing and Matrigel™ invasion
assays demonstrated a marked elevation of cell migration in the
cells transfected with the HIF2PUT siRNAs, as compared with the
negative control group (Fig. 3D and
E).
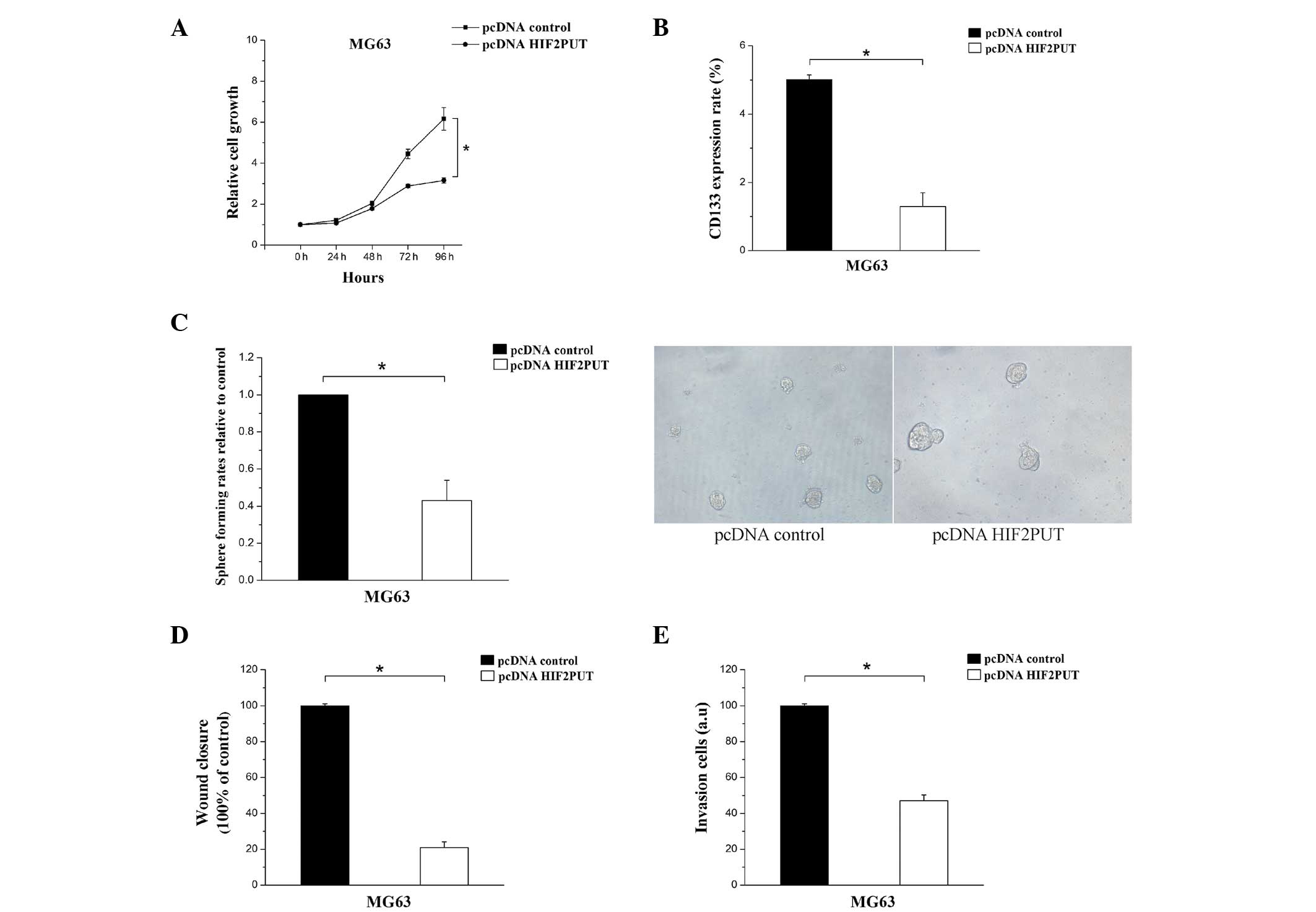
Overexpression of HIF2PUT decreases the
proliferation, migration and self-renewal of MG63 cells
To further identify the role of HIF2PUT in MG63
cells, functional assays were performed with MG63 cells with an
overexpression of HIF2PUT (pcDNA-HIF2PUT), as compared with the
MG63 cells transfected with pcDNA-control. The MG63 cells
transfected with pcDNA-HIF2PUT had a significantly slower rate of
growth, as compared with the cells transfected with the
pcDNA-control (Fig. 4A). To
determine the effects of HIF2PUT overexpression on CSCs in MG63
cells in vitro, the percentage of CD133+
expressing cells was determined by FCM, and the stem cell
sphere-forming ability was assessed by spheroid-formation assay.
Following transfection with pcDNA-HIF2PUT, the percentage of
CD133+ CSCs and the number of stem cell spheres were
significantly decreased in MG63 cells in vitro (Fig. 4B and C). Furthermore, the migratory
and invasive ability of the cells was markedly inhibited by
pcDNA-HIF2PUT transfection (Fig. 4D
and E).
Discussion
Osteosarcoma, and other types of cancer, contains a
small subset of cells with stem cell-like properties, which are
responsible for tumor aggression and recurrence (20). An improved understanding regarding
the molecular network modulating these stem cell characteristics is
essential for the development of effective therapeutic strategies
for osteosarcoma.
Recently, growing evidence has indicated that the
human genome is largely transcribed into noncoding RNAs, such as
microRNAs and lncRNAs. lncRNAs may have critical regulating roles
in the extensive biological processes of certain cells, including
stem cells (10). Previous studies
have suggested that lncRNAs may act as crucial modulators in cancer
development (7–9).
However, the functional role of lncRNAs in CSCs
remains unknown, particularly in osteosarcoma. In the present
study, by loss- and gain-of function experiments, a novel lncRNA,
HIF2PUT, was shown to inhibit the proliferation, self-renewal and
migration of osteosarcoma stem cells. CD133 is considered a CSC
marker of osteosarcoma (20). The
present study showed that overexpression of HIF2PUT markedly
decreased the CD133+ cell population in MG63 cells.
These results suggest that HIF2PUT may negatively modulate the CSC
population in osteosarcoma. The present study may provide the first
CSC-associated lncRNA: HIF2PUT, in osteosarcoma, which may be a
potential therapeutic target.
HIF2PUT is a PROMPT of HIF-2α. The cellular
biological effects of PROMPTs in the human body have yet to be
elucidated (8). Previous studies
have suggested that PROMPTs can bind other proteins. Growth
arrest-specific 5 is an important cancer associated lncRNA that can
regulate downstream genes by binding to the transcription factor,
ensuring the transcription factor can no longer bind to promoters
(22). However, other studies have
suggested that the majority of PROMPTs exert their function by
regulating the transcriptional activity of their host PCGs
(11–14).
The present study investigated the association
between HIF2PUT and its host PCG, HIF-2α. The expression levels of
HIF2PUT were shown to be positively correlated with those of HIF-2α
in osteosarcoma tissues. Furthermore, in the MG63 cells,
downregulation of HIF2PUT expression resulted in a concomitant
decrease in the mRNA expression levels of HIF-2α. These results
suggest that the HIF2PUT may be a regulator of HIF-2α in
osteosarcoma.
It has previously been reported that transcripts of
HIF-2α are restricted to particular cell types (15). The present study showed that
expression of HIF-2α in osteosarcoma tissues can be detected;
however, the expression levels in all four osteosarcoma cell lines
were low. This may be due to the osteosarcoma tissues containing
not only tumor cells, but also numerous types of supporting cells.
Knowles et al (23),
detected low expression levels of HIF-2α in MG63 cells in
non-hypoxic conditions; however, the expression levels were higher
in hypoxic conditions, which coincides with the results of the
present study.
Previous studies have indicated that the role of
HIF-2α may vary from cancer gene to tumor suppressor in numerous
types of cancer (17,24–26).
The present study could not identify the exact function of HIF-2α
in osteosarcoma, due to the extremely low expression. In
conclusion, the results of the present study may provide primary
evidence that lncRNA-HIF2PUT can suppress the properties of CSCs in
osteosarcoma, partly by regulating the expression of HIF-2α.
Further work targeting this novel lncRNA in osteosarcoma may
provide potential therapeutic targets in the future.
Acknowledgements
The authors of the present study are grateful for
the support provided by Dr Shuyun Liu, for the kind suggestions and
technical assistance.
References
|
1
|
Stiller CA: International patterns of
cancer incidence in adolescents. Cancer Treat Rev. 33:631–645.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Mankin HJ, Hornicek FJ, Rosenberg AE,
Harmon DC and Gebhardt MC: Survival data for 648 patients with
osteosarcoma treated at one institution. Clin Orthop Relat Res.
429:286–291. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Harting MT, Blakely ML, Jaffe N, et al:
Long-term survival after aggressive resection of pulmonary
metastases among children and adolescents with osteosarcoma. J
Pediatr Surg. 41:194–199. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Merlos-Suárez A, Barriga FM, Jung P, et
al: The intestinal stem cell signature identifies colorectal cancer
stem cells and predicts disease relapse. Cell Stem Cell. 8:511–524.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Takahashi S, Kamiyama T, Tomaru U, et al:
Frequency and pattern of expression of the stem cell marker CD133
have strong prognostic effect on the surgical outcome of colorectal
cancer patients. Oncol Rep. 24:1201–1212. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Ailles LE and Weissman IL: Cancer stem
cells in solid tumors. Curr Opin Biotechnol. 18:460–466. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Huarte M and Rinn JL: Large non-coding
RNAs: missing links in cancer? Hum Mol Genet. 19:R152–R161. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Spizzo R, Almeida MI, Colombatti A and
Calin GA: Long non-coding RNAs and cancer: a new frontier of
translational research? Oncogene. 31:4577–4587. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Tsai MC, Spitale RC and Chang HY: Long
intergenic noncoding RNAs: new links in cancer progression. Cancer
Res. 71:3–7. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Guttman M, Donaghey J, Carey BW, et al:
lincRNAs act in the circuitry controlling pluripotency and
differentiation. Nature. 477:295–300. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Preker P, Nielsen J, Kammler S, et al: RNA
exosome depletion reveals transcription upstream of active human
promoters. Science. 322:1851–1854. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Jacquier A: The complex eukaryotic
transcriptome: unexpected pervasive transcription and novel small
RNAs. Nat Rev Genet. 10:833–844. 2009. View
Article : Google Scholar : PubMed/NCBI
|
|
13
|
Preker P, Almvig K, Christensen MS, et al:
PROMoter uPstream Transcripts share characteristics with mRNAs and
are produced upstream of all three major types of mammalian
promoters. Nucleic Acids Res. 39:7179–7193. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Taft RJ, Kaplan CD, Simons C and Mattick
JS: Evolution, biogenesis and function of promoter-associated RNAs.
Cell Cycle. 8:2332–2338. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Ijichi N, Ikeda K, Fujita M, et al: EPAS1,
a dexamethasone-inducible gene in osteoblasts, inhibits
osteoblastic differentiation. Open Bone J. 1:28–37. 2009.
View Article : Google Scholar
|
|
16
|
Li Z, Bao S, Wu Q, et al:
Hypoxia-inducible factors regulate tumorigenic capacity of glioma
stem cells. Cancer Cell. 15:501–513. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Gordan JD, Bertout JA, Hu CJ, Diehl JA and
Simon MC: HIF-2alpha promotes hypoxic cell proliferation by
enhancing c-myc transcriptional activity. Cancer Cell. 11:335–347.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Covello KL, Kehler J, Yu H, et al:
HIF-2alpha regulates Oct-4: effects of hypoxia on stem cell
function, embryonic development, and tumor growth. Genes Dev.
20:557–570. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Yang X, Song JH, Cheng YL, et al: Long
non-coding RNA HNF1A-AS1 regulates proliferation and migration in
oesophageal adenocarcinoma cells. Gut. 63:881–890. 2014. View Article : Google Scholar
|
|
20
|
Siclari VA and Qin L: Targeting the
osteosarcoma cancer stem cell. J Orthop Surg Res. 5:782010.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Tirino V, Desiderio V, d’Aquino R, et al:
Detection and characterization of CD133+ cancer stem
cells in human solid tumours. PLoS One. 3:e34692008. View Article : Google Scholar
|
|
22
|
Williams GT, Mourtada-Maarabouni M and
Farzaneh F: A critical role for non-coding RNA GAS5 in growth
arrest and rapamycin inhibition in human T-lymphocytes. Biochem Soc
Trans. 39:482–486. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Knowles HJ, Schaefer KL, Dirksen U and
Athanasou NA: Hypoxia and hypoglycaemia in Ewing’s sarcoma and
osteosarcoma: regulation and phenotypic effects of
Hypoxia-Inducible Factor. BMC Cancer. 10:3722010. View Article : Google Scholar
|
|
24
|
Acker T, Diez-Juan A, Aragones J, et al:
Genetic evidence for a tumor suppressor role of HIF-2alpha. Cancer
Cell. 8:131–141. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Yu L and Hales CA: Long-term exposure to
hypoxia inhibits tumor progression of lung cancer in rats and mice.
BMC Cancer. 11:3312011. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Biswas S, Charlesworth PJ, Turner GD, et
al: CD31 angiogenesis and combined expression of HIF-1α and HIF-2α
are prognostic in primary clear-cell renal cell carcinoma (CC-RCC),
but HIFα transcriptional products are not: implications for
antiangiogenic trials and HIFα biomarker studies in primary CC-RCC.
Carcinogenesis. 33:1717–1725. 2012. View Article : Google Scholar : PubMed/NCBI
|