Introduction
Compared with percutaneous coronary intervention and
medical therapy, coronary artery bypass graft (CABG) has been
demonstrated to be a more effective method for the treatment of
coronary artery disease (1). The
saphenous vein is the most commonly used vessel for the graft in
CABG surgery; however, vein graft failure (VGF) is a serious
complication that affects the curative efficacy of CABG.
Endothelial dysfunction is a critical factor in VGF (2). Following CABG, the vein graft wall
sustains high blood pressure and is frequently injured by
mechanical stretch. This injury leads to endothelial dysfunction,
which in turn induces alterations in endothelial cell (EC)
proliferation, cytokine secretion, leukocyte adherence and platelet
aggregation (3–5). However, the mechanism underlying
endothelial dysfunction remains unclear.
MicroRNAs (miRNAs) are small non-coding RNA
molecules, 22 nucleotides in length, that are crucial in cell
growth, differentiation, metabolism, apoptosis and migration
(6). In a previous study, it was
demonstrated that early growth response 1 (EGR1) is a vital factor
in the development of VGF. Additionally, certain miRNAs that were
able to regulate EGR1 expression were predicted via miRBase
(http://mirbase.org), Target Scan Human (http://www.targetscan.org/) and miRDB (http://mirdb.org/miRDB/) (Zheng et al,
unpublished data). Several previous studies have suggested that
miRNAs are involved in various pathological cardiovascular
conditions, including atherosclerosis, hypertension and arrhythmia
(7–9). In 2013, McDonald et al
(10) demonstrated that in VGF,
miR-21 induces intimal hyperplasia by regulating the expression of
the phosphatase and tensin homolog deleted on chromosome 10, signal
transducer and activator of transcription 3 and proliferating cell
nuclear antigen. These results suggested that miRNAs are vital
regulated factors and may be used as novel genetic therapeutic
targets in the treatment of VGF (11). However, in endothelial dysfunction
induced by mechanical stretch, the expression and mechanistic
regulation of EC miRNAs remains unclear. An improved understanding
of miRNA expression in ECs with mechanical stretch stimulation may
lead to the development of novel biomarkers and therapeutic targets
for the initial stages of VGF. Therefore, in order to identify
miRNA expression in endothelial dysfunction, the current study
conducted miRNA profiling using samples obtained from ECs
undergoing mechanical stretch.
Materials and methods
Mechanical stretch stimulation
Human umbilical vein endothelial cells (HUVECs;
GeneChem Co., Ltd., Shanghai, China) were maintained in Dulbecco's
modified Eagle's medium 140 (Gibco Life Technologies, Grand Island,
NY, USA) with 10% fetal bovine serum (Gibco Life Technologies) at
37°C with 5% CO2. The ECs were then plated on silicone
elastomer-bottomed and collagen-coated plates (Flexcell
International Corporation, McKeesport, PA, USA). A
computer-controlled Cyclic Stress Unit (Flexcell 5100; Flexcell
International Corporation) was used to subject HUVECs to mechanical
stretch for 24 h (cyclic deformation at 60 cycles/min, elongation
of 15%).
RNA isolation
Total RNA was isolated with the mirVana miRNA
Isolation kit (Applied Biosystems, Foster City, CA, USA) in
accordance with the manufacturer's instructions. RNA quantity and
purity was assessed using a NanoDrop ND-1000 spectrophotometer
(Thermo Fisher Scientific, Wilmington, MA, USA). A260/A280≥1.6 and
A260/A230≥1 indicated acceptable RNA purity, while an acceptable
RNA integrity number ≥5 was obtained using the Agilent RNA 6000
Nano assay (Agilent Technologies, Inc., Santa Clara, CA, USA). gDNA
contamination was evaluated by gel electrophoresis.
Microarray profiling
Human genome-wide miRNA micro-array analysis was
performed by Phalanx Biotech with the Human miRNA
OneArray® V4 (Phalanx Biotech Group, Hsinchu, Taiwan,
R.O.C.). Each microarray contained 144 experimental control probes
and 1,884 unique human miRNA probes. Each unique probe had three
features, and probes contained 100% Sanger miRBase V18 miRNA
content. Briefly, fluorescent targets were prepared from 2.5
µg total RNA samples using the miRNA ULSTM Labeling kit
(Leica Biosystems Nussloch GmbH, Nussloch, Germany). Labeled miRNA
targets were hybridized to the micro-array with Phalanx
hybridization buffer in the OneArray® Hybridization
Chamber. Following overnight hybridization at 37°C, non-specific
binding was removed by three washing steps (wash one, 37°C, 5 min;
wash two, 37°C, 5 min and 25°C, 5 min; and wash three: rinse 20
times) with 2X saline sodium citrate solution (Loogene, Beijing,
China). The slides were dried by centrifugation at 70 × g for 5 min
and scanned using an Axon GenePix® 4000B scanner
(Molecular Devices, Sunnyvale, CA, USA). The Cy5 fluorescence
intensity of each spot was analyzed using GenePix, version 4.1
software (Molecular Devices). The signal intensity of each spot was
processed using the R project, version 2.12.1 (Bell Laboratories,
Murray Hill, NJ, USA). Spots for which the flag was <0 were
filtered out, whereas spots that passed the criteria were
normalized using the 75% scaling normalization method. Normalized
spot intensities were then converted into log2 ratios of gene
expression between the control and treatment groups. Spots with a
log2 ratio of ≥1 or ≤−1, in addition to a P<0.05 were analyzed
further. These differentially expressed miRNAs were subjected to
hierarchical cluster analysis using average linkage and Pearson's
correlation in order to measure the similarity.
Validation of microRNAs by reverse
transcription-quantitative polymerase chain reaction (RT-qPCR)
analysis
The differentially expressed miRNAs selected from
the microarray were validated by RT-qPCR analysis. cDNA was
generated with the PrimeScript™ RT-qPCR kit (Takara Bio, Inc.,
Otsu, Japan) in accordance with the manufacturer's instructions.
PCR was performed in a 7900HT fast RT-qPCR instrument (Applied
Biosystems Life Technologies, Singapore). The primers used were
purchased from Ribobio (Guangzhou, China) and the sequences for the
control were as follows: U6, forward 5′-GCGCGTCGTGAAGCGTTC-3′ and
reverse 5′-GTGCAGGGTCCGAGGT-3′. All reactions involved initial
denaturation at 95°C for 3 min, followed by 40 cycles of 95°C for
10 sec, 60°C for 20 sec and 70°C for 1 sec. miRNA quantification
was performed by using the miDETECT A TrackTM miRNA qRT-PCR Starter
Kit (Ribobio) in a 7900HT fast RT-qPCR instrument (Applied
Biosystems, Foster City, CA, USA). All quantitative reactions,
including the no-template controls, were run in duplicate. The
small nuclear RNA U6 was used as the normalization control.
Gene Ontology (GO) and Kyoto Encyclopedia
of Genes and Genomes (KEGG) pathway analyses
In order to identify possible mRNA targets of the
differentially expressed miRNAs, GO and KEGG were performed using
miRBase (http://www.mirbase.org/). Enrichment
analysis of miRNA target genes was performed using Cytoscape
(http://www.cytoscape.org/). Using
P<0.05 as a cutoff, three different annotations were analyzed:
Molecular function, biological process and cellular component.
The DAVID database (http://david.abcc.ncifcrf.gov/) was utilized in order
to analyze the significant pathways that involved genes that were
differentially expressed in HUVECs undergoing stimulation by
mechanical stretch. Selection was based on the results of Fisher's
exact test and the threshold of significance was defined by the
P-value.
Statistical analysis
Data are expressed as the mean ± standard error.
Student's t-test was used to determine the significance of
differences in miRNA expression between groups. All statistical
analyses were performed using SPSS, version 17.0 (SPSS, Inc.,
Chicago, IL, USA). P<0.05 was considered to indicate a
statistically significant difference. R project was used to
normalize data for the selection of differentially expressed
miRNAs.
Results
miRNA signature in HUVECs following
mechanical stretch stimulation
A total of 5,796 probes were included in the miRNA
microarray that was used in the current study. miRNA microarray
hybridization revealed that a total of 38 miRNAs were significantly
differentially expressed following mechanical stretch stimulation.
Of these, 20 were upregulated and 18 were downregulated (based on a
fold change ≥0.8 and P<0.05; Table
I). The differential miRNA expression patterns of HUVECs
stimulated by mechanical stretch were accurately separated from
those of the unstimulated controls by hierarchical cluster analysis
(Fig. 1).
 | Table IDifferentially expressed miRNAs in
human umbilical vein endothelial cells undergoing mechanical
stretch stimulation, selected at log2 |Fold change|≥0.8 and
P<0.05. |
Table I
Differentially expressed miRNAs in
human umbilical vein endothelial cells undergoing mechanical
stretch stimulation, selected at log2 |Fold change|≥0.8 and
P<0.05.
| Name | Fold change | P-value |
|---|
| has-miR-363-5p | −1.3723 | 0.0148 |
| has-miR-668 | −1.3153 | 0.0030 |
| has-miR-4638-5p | −1.2859 | 0.0196 |
| has-miR-16-5p | −1.1988 | 0.0177 |
| Has-miR-3676-3p | −1.1492 | 0.0029 |
| Has-miR-3175 | −1.0751 | 0.0007 |
| Has-miR-106a-5p | −1.0446 | 0.0395 |
| Has-miR-20a-5p | −1.0020 | 0.0097 |
| Has-miR-3156-5p | 1.1413 | 0.0041 |
| Has-miR-4660 | 1.1456 | 0.0036 |
| Has-miR-4725-3p | 1.1457 | 0.0498 |
| Has-miR-4271 | 1.1810 | 0.0134 |
| Has-miR-1273e | 1.1995 | 0.0297 |
| Has-miR-1273f | 1.2424 | 0.0023 |
| Has-miR-551b-5p | 1.2642 | 0.0030 |
| Has-miR-6514-5p | 1.3973 | 0.0011 |
| Has-miR-3691-5p | 1.5138 | 0.0033 |
| hsa-miR-584-5p | 1.5218 | 0.0041 |
| Has-miR-4430 | 2.3862 | 0.0071 |
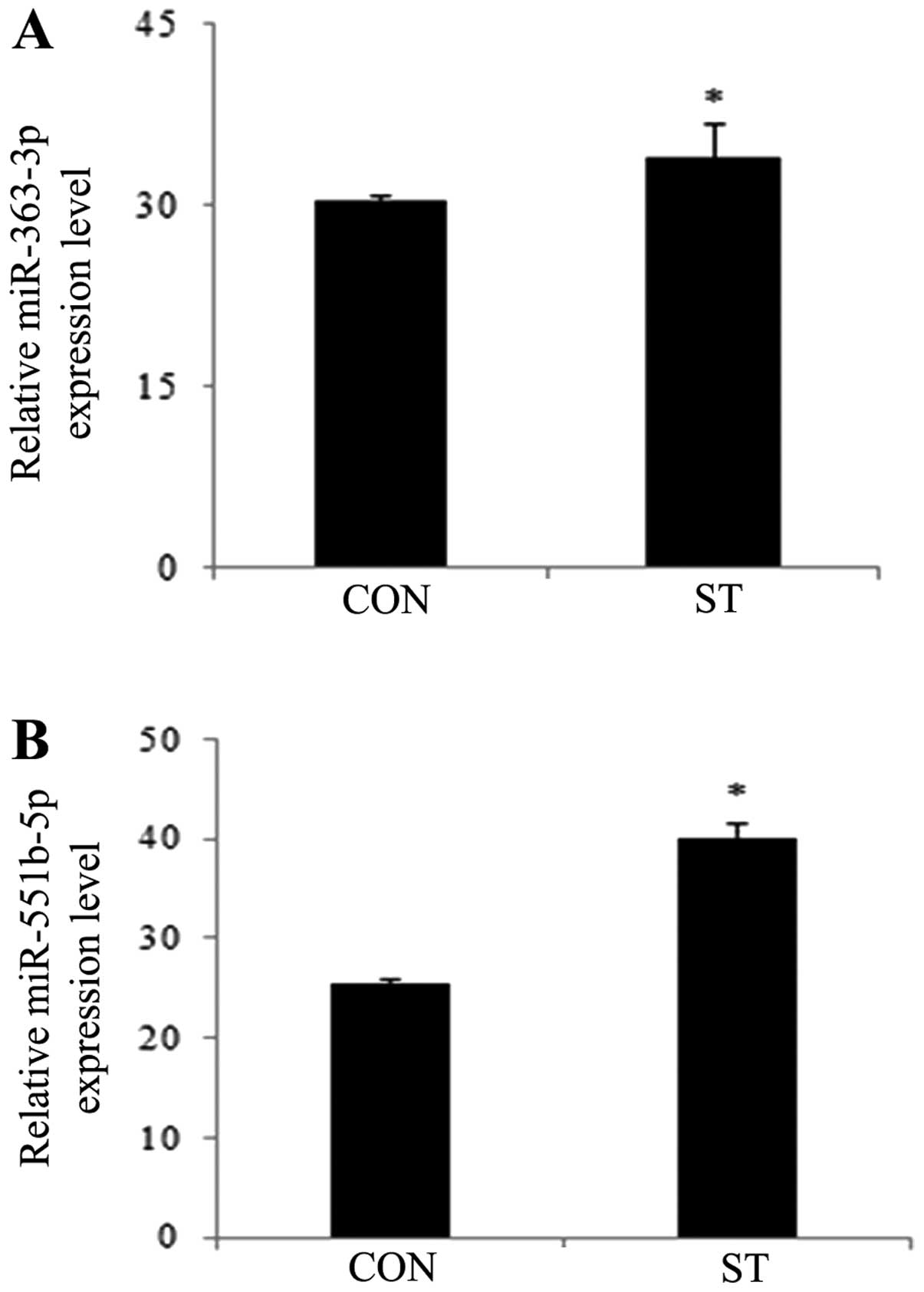
Validation of miRNA by RT-qPCR
RT-qPCR was used to validate the results of two
miRNAs that were identified through the microarray analysis. The
results demonstrated that the expression levels of miR-363-3p and
miR-551b-5p were significantly increased in ECs with mechanical
stretch stimulation, compared with the control ECs (Fig. 2). This result supported the miRNA
expression signatures identified in a previous experiment (Zheng
et al, unpublished data). However, no novel miRNAs were
identified.
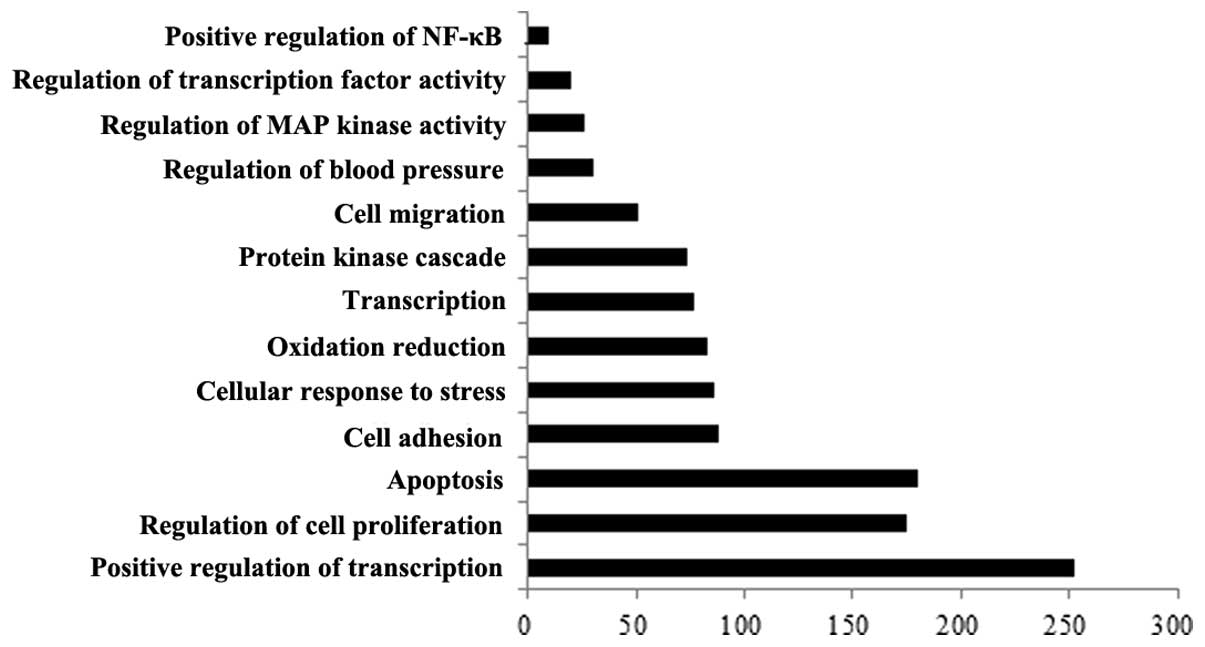
GO analysis
Using GO analysis of biological processes, the
differentially expressed miRNAs were classified into different
functional categories. As presented in Fig. 3, these miRNAs were identified to be
involved in various vessel-associated processes, including positive
regulation of nuclear factor-κB transcription, and regulation of
transcription factor activity, mitogen-activated protein kinase
(MAPK) activity and blood pressure.
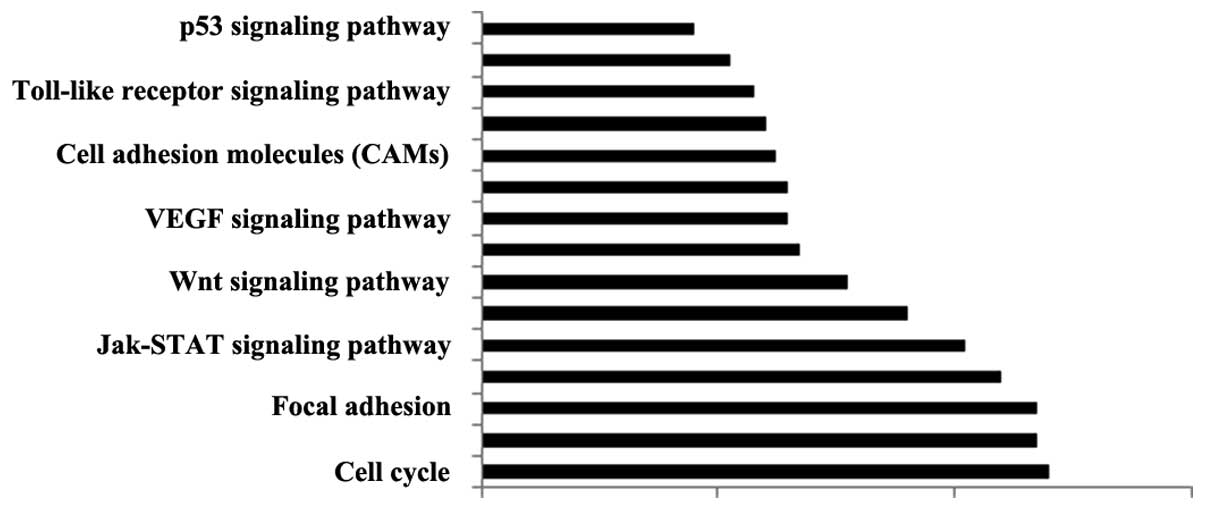
miRNA-associated regulatory networks
KEGG pathway analysis was conducted in order to
determine the pathways associated with the differentially expressed
miRNAs. It was observed that the miRNAs were involved in various
pathways and proteins associated with vessel diseases, including
the p53 signaling pathway, apoptosis, expression of cell adhesion
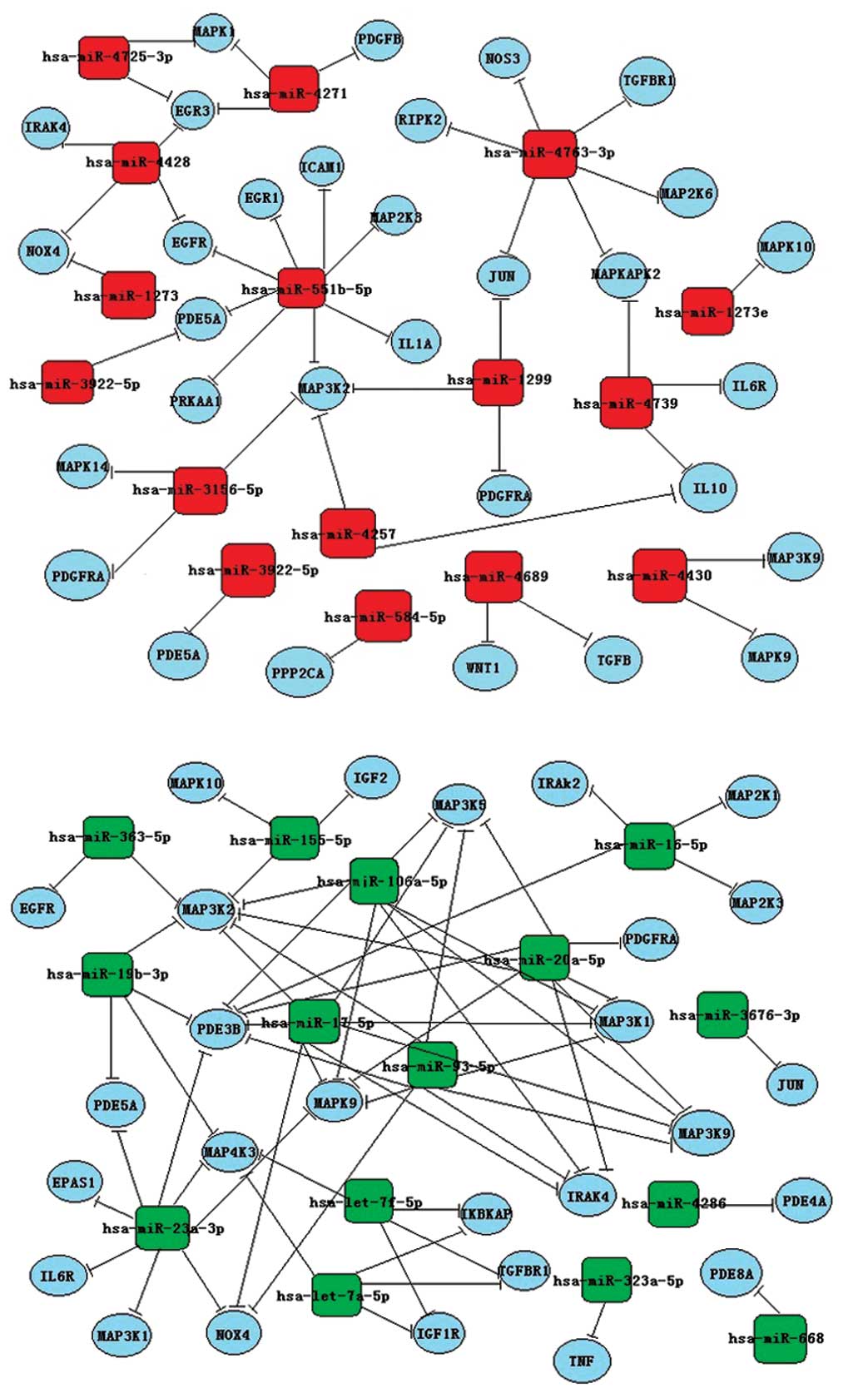
molecules and chemokine signaling pathways (Fig. 4). As demonstrated in Fig. 5, the differentially expressed
miRNAs had a variety of potential targets.
Discussion
VGF is a serious complication that requires
investigation in order to improve prognosis in cardiac surgery.
Previous studies have demonstrated that subsequent to CABG surgery,
VGF occurred in 25% of patients within 1 year and 50% of patients
within 5 years (12). Compared
with those undergoing their first CABG surgery, the mortality risk
has been reported to increase 13-fold in patients undergoing repeat
CABG surgery due to VGF (13).
Furthermore, diagnosing and treating VGF at early stages greatly
improves patient prognosis (14).
miRNAs regulate gene expression by acting on the
3′-untranslated regions of target genes, and serve a central role
in various physiological and pathological processes. Compared with
normal controls, differentially expressed miRNAs may be used as
diagnostic biomarkers and therapeutic targets. For the purpose of
elucidating the mechanism of endothelial dysfunction, the current
study focused on an miRNA signature present in ECs stimulated by
mechanical stretch. In total, 38 differentially expressed miRNAs
were identified in these cells, including 20 that were upregulated
and 18 that were downregulated. In order to ensure the quality of
the array, certain miRNAs were validated by RT-qPCR. GO and KEGG
pathway analysis was undertaken in order to predict the target
genes and functional pathways of the differentially expressed
miRNAs. GO analysis revealed that the target genes of these miRNAs
were involved in pathological processes associated with VGF,
including cell adhesion, cellular responses to stress and
regulation of transcription. Of these genes, the most notable was
EGR1. EGR1 is a gene that is expressed acutely at early VGF stages,
with no sustained expression. Various previous studies have
suggested that EGR1 is a critical transcription factor involved in
the regulation of intracellular signaling, cell growth,
differentiation and wound healing (15–17).
In a previous study, it was observed that EGR1 is involved in VGF
via regulating the promotion of EC proliferation, in addition to
the expression of inflammatory factors following mechanical stretch
stimulation (3). Additional
studies have demonstrated that EGR1 expression may upregulate
monocyte chemoattractant protein-1 expression, promote vascular
inflammation and induce thrombosis and atherosclerosis (18). Additionally, EGR1 has been
indicated to be capable of stimulating vascular smooth muscle cell
(VSMC) growth, migration and apoptosis, and inducing VGF via the
IGF-1 pathway (19).
KEGG analysis in the current study demonstrated that
the transforming growth factor-β (TGF-β) and MAPK signaling
pathways were involved in VGF. In the early stages of vascular
injury, TGF-β is released by inflammatory cells and platelets,
which attracts inflammatory factors and monocytes that secrete
fibroblast growth factor, tumor necrosis factor (TNF) and
interleukins. In turn, this promotes VSMC proliferation and
migration and increases extracellular matrix production. In
addition, TGF-β may promote collagen type I and III composition,
and induce intimal hyperplasia and vascular wall fibrosis (20). The MAPK signaling pathway is
strongly associated with autogenous vein grafts and vein
arterializations (21,22). The MAPK family includes Jun
N-terminal kinase, p38 and extracellular signal-regulated kinase,
and these molecules are involved in cell growth, differentiation
and apoptotic processes (22–24).
Cornelissen et al (25)
reported that mechanical stretch is an important factor for
inducing p38-MAPK activation in pressure distension of human
saphenous veins, while also identifying that p38-MAPK is associated
with apoptosis. TNF-α stimulates TNF receptor 1, activates MAPK
expression and induces VSMC migration (26). Due to the fact that VSMC migration
and apoptosis are important processes in VGF, Ge et al
(27) were able to successfully
use an MAPK inhibitor to limit intimal hyperplasia.
In the present study, 38 differentially expressed
miRNAs were identified, a number of which had been previously
reported to be associated with VGF. Zhi et al (28) identified a reduction of miR-106a-5p
expression in astrocytoma, and its target gene Fas-activated
serine/threonine kinase was upregulated, leading to poor survival
outcomes in patients with astrocytoma. Costa et al (29) reported that bone marrow endothelial
cells express miR-363-5p. It was observed that the miRNA was able
to directly modulate the expression of the angiocrine factors
tissue inhibitor of metalloprotein-ases-1 and thrombospondin 3 at
the post-transcriptional level. miR-363-5p downregulation
additionally was observed to result in a significant reduction in
EC tube formation on Matrigel, while increasing the adhesion of
hematopoietic precursor cells to ECs (29).
VGF is a complex disease caused by multiple genes,
factors and signaling pathways. An miRNA signature may provide a
basis for investigating the mechanism of this disease, in addition
to aiding in the identification of novel biomarkers or targets for
diagnosis and therapy of early-stage VGF. In the current study, an
miRNA signature was identified in ECs stimulated by mechanical
stretch. The results of the present study suggest that miRNAs are
vital in the process of endothelial dysfunction and are involved in
the pathogenesis of VGF. Thus, using miRNAs as biomarkers and
therapeutic targets has the potential to improve early diagnosis
and prognosis of patients with VGF.
Acknowledgments
The current study was supported by the National
Natural Science Foundation of China (grant no. 81070202).
Abbreviations:
|
CABG
|
coronary artery bypass graft
|
|
VGF
|
vein graft failure
|
|
EC
|
endothelial cell
|
|
miRNAs
|
microRNAs
|
|
EGR1
|
early growth response 1
|
|
RT-qPCR
|
reverse transcription-quantitative
polymerase chain reaction
|
|
GO
|
Gene Ontology
|
|
KEGG
|
Kyoto Encyclopedia of Genes and
Genomes
|
|
VSMC
|
vascular smooth muscle cell
|
|
MAPK
|
mitogen-activated protein kinase
|
References
|
1
|
Deb S, Wijeysundera HC, Ko DT, Tsubota H,
Hill S and Fremes SE: Coronary artery bypass graft surgery vs
percutaneous interventions in coronary revascularization: a
systematic review. JAMA. 310:2086–2095. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Li FD, Sexton KW, Hocking KM, et al:
Intimal thickness associated with endothelial dysfunction in human
vein grafts. J Surg Res. 180:e55–e62. 2013. View Article : Google Scholar
|
|
3
|
Zhang K, Cao J, Dong R and Du J: Early
growth response protein 1 promotes restenosis by upregulating
intercellular adhesion molecule-1 in vein graft. Oxid Med Cell
Longev. 2013:4324092013. View Article : Google Scholar
|
|
4
|
Okada M, Matsumori A, Ono K, et al: Cyclic
stretch upregulates production of interleukin-8 and monocyte
chemotactic and activating factor/monocyte chemoattractant
protein-1 in human endothelial cells. Arterioscler Thromb Vasc
Biol. 18:894–901. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Wu MH, Kouchi Y, Onuki Y, et al: Effect of
differential shear stress on platelet aggregation, surface
thrombosis, and endothelialization of bilateral carotid-femoral
grafts in the dog. J Vasc Surg. 22:382–392. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Jinek M and Doudna JA: A three-dimensional
view of the molecular machinery of RNA interference. Nature.
457:405–412. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Small EM and Olson EN: Pervasive roles of
microRNAs in cardiovascular biology. Nature. 469:336–342. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Moazed D: Small RNAs in transcriptional
gene silencing and genome defence. Nature. 457:413–420. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Lanford RE, Hildebrandt-Eriksen ES, Petri
A, et al: Therapeutic silencing of microRNA-122 in primates with
chronic hepatitis C virus infection. Science. 327:198–201. 2010.
View Article : Google Scholar
|
|
10
|
McDonald RA, White KM, Wu J, et al:
miRNA-21 is dysregulated in response to vein grafting in multiple
models and genetic ablation in mice attenuates neointima formation.
Eur Heart J. 34:1636–1643. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Stellos K and Dimmeler S: Vascular
microRNAs: from disease mechanisms to therapeutic targets. Circ
Res. 114:3–4. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Parang P and Arora R: Coronary vein graft
disease: pathogenesis and prevention. Can J Cardiol. 25:e57–e62.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Harskamp RE, Lopes RD, Baisden CE, de
Winter RJ and Alexander JH: Saphenous vein graft failure after
coronary artery bypass surgery: pathophysiology, management and
future directions. Ann Surg. 257:824–833. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Lopes RD, Mehta RH, Hafley GE, et al:
Relationship between vein graft failure and subsequent clinical
outcomes after coronary artery bypass surgery. Circulation.
125:749–756. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Rekhviashvili A and Abashidze R: The
relationship between endothelial dysfunction and 24-hour blood
pressure rhythm in patients with arterial hypertension. Georgian
Med News. 155:13–17. 2008.PubMed/NCBI
|
|
16
|
Harja E, Bucciarelli LG, Lu Y, Stern DM,
Zou YS, Schmidt AM and Yan SF: Early growth response-1 promotes
atherogenesis: mice deficient in early growth response-1 and
apolipoprotein E display decreased atherosclerosis and vascular
inflammation. Circ Res. 94:333–339. 2004. View Article : Google Scholar
|
|
17
|
Fujita T, Asai T, Andrassy M, et al:
PKCbeta regulates ischemia/reperfusion injury in the lung. J Clin
Invest. 113:1615–1623. 2004. View
Article : Google Scholar : PubMed/NCBI
|
|
18
|
Maekawa T, Takahashi N, Honda T, et al:
Porphyromonas gingivalis antigens and interleukin-6 stimulate the
production of monocyte chemoattractant protein-1 via the
upregulation of early growth response-1 transcription in human
coronary artery endothelial cells. J Vasc Res. 47:346–354. 2010.
View Article : Google Scholar
|
|
19
|
Wu X, Cheng J, Li P, Yang M, Qiu S, Liu P
and Du J: Mechano-sensitive transcriptional factor Egr-1 regulates
insulin-like growth factor-1 receptor expression and contributes to
neointima formation in vein grafts. Arterioscler Thromb Vasc Biol.
30:471–476. 2010. View Article : Google Scholar
|
|
20
|
Marchesi C, Paradis P and Schiffrin EL:
Role of the renin-angiotensin system in vascular inflammation.
Trends Pharmacol Sci. 29:367–374. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Pintucci G, Saunders PC, Gulkarov I, et
al: Anti-proliferative and anti-inflammatory effects of topical
MAPK inhibition in arterialized vein grafts. FASEB J. 20:398–400.
2006.
|
|
22
|
Saunders PC, Pintucci G, Bizekis CS, et
al: Vein graft arterialization causes differential activation of
mitogen-activated protein kinases. J Thorac Cardiovasc Surg.
127:1276–1284. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Pearson G, Robinson F, Beers Gibson T, Xu
BE, Karandikar M, Berman K and Cobb MH: Mitogen-activated protein
(MAP) kinase pathways: regulation and physiological functions.
Endocr Rev. 22:153–183. 2001.PubMed/NCBI
|
|
24
|
Wang XZ and Ron D: Stress-induced
phosphorylation and activation of the transcription factor CHOP
(GADD153) by p38 MAP Kinase. Science. 272:1347–1349. 1996.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Cornelissen J, Armstrong J and Holt CM:
Mechanical stretch induces phosphorylation of p38-MAPK and
apoptosis in human saphenous vein. Arterioscler Thromb Vasc Biol.
24:451–456. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Goetze S, Xi XP, Kawano Y, Kawano H, Fleck
E, Hsueh WA and Law RE: TNF-alpha-induced migration of vascular
smooth muscle cells is MAPK dependent. Hypertension. 33:183–189.
1999. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Ge JJ, Zhao ZW, Zhou ZC, Wu S, Zhang R,
Pan FM and Abendroth DK: p38 MAPK inhibitor, CBS3830 limits
vascular remodelling in arterialised vein grafts. Heart Lung Circ.
22:751–758. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Zhi F, Zhou G, Shao N, et al: miR-106a-5p
inhibits the proliferation and migration of astrocytoma cells and
promotes apoptosis by targeting FASTK. PLoS One. 8:e723902013.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Costa A, Afonso J, Osório C, et al:
miR-363-5p regulates endothelial cell properties and their
communication with hematopoietic precursor cells. J Hematol Oncol.
6:872013. View Article : Google Scholar : PubMed/NCBI
|