Introduction
A promising future for the use of bone marrow
mesenchymal stem cells (BMSCs) in regenerative medicine is
anticipated, due to their ability to self-replicate and
differentiate into various functional cells. Stem cells are the
most widely used seed cells in the field of neural regeneration and
bone tissue engineering, due to their easy availability, reduced
side effects, biological scaffold adhesion abilities, and promising
performances in vitro, as well as their strong proliferative
capacity in culture and passaging (1–6).
Therefore, stem cells are currently being widely investigated and
applied in the clinic (1). BMSCs
possess powerful differentiation potential and immunological
regulatory properties, and therefore have potential for use in the
field of cell injury. Numerous studies have demonstrated that BMSCs
are able to differentiate into neuron-like cells in vitro
via genetic manipulation, where various factors and chemical agents
are adopted to induce BMSC differentiation into neuron-like or
Schwann cells (1–3,7).
However, due to ethical considerations and the various methods used
to induce adult stem cell neural differentiation, how to produce
and identify the source of neuron-like cells for clinical cell
therapy is currently a significant problem.
BMSCs transplanted in adult Sprague-Dawley (SD) rat
may respond to microenvironmental cues and differentiate into
neuron-like cells. In addition, BMSCs have been shown to migrate
towards the source of lesions in the brain (4), which may enhance the repairing
capacity of injured tissue in animal models. A previous study
demonstrated that BMSC transplantation into the central nervous
system was able to impede Alzheimer's disease-like pathology and
upregulate δNp73 expression in the hippocampus of APP/PS1
transgenic mice (5). In addition,
Mohammadi et al (2,8) used undifferentiated bone marrow
stromal cells to induce sciatic nerve regeneration in rats.
Administration of BMSCs via the central nervous system and
peripheral system is considered to be safe in human subjects
(8–10). BMSCs may become a clinical choice
for cell therapy of the central nervous and peripheral systems,
since BMSCS have the advantage of reduced ethical regulation and do
not often induce tissue rejection. The rapid development of nerve
engineering technology has enabled many investigators to examine
the use of natural and artificial biomaterials. Constructed grafts
may be used to connect and repair in neurological regeneration
(9–11); however, the new nerve must possess
biocompatibility. Conversely, stem cells have the ability to
secrete neurotropic factors to repair injured neurons. BMSCs are
not prone to ethical and tissue rejection-related concerns;
however, further studies on the use of human BMSCs are
required.
Basic fibroblast growth factor (bFGF) and nerve
growth factor (NGF) are powerful mitogens that promote the
nutrition of neural stem cells and precursor cells present in the
mature nervous system. Through the expression of nerve-related
proteins, bFGF promotes cell proliferation and mitosis, and
enhances neuronal axon regeneration and spinal cord injury repair
(12). NGF is a homodimeric
peptide. By supporting the survival and growth of neural cells in
the nervous system, it is able to regulate cell growth and promote
neural differentiation. Furthermore, NGF exhibits nerve injury
healing ability in clinical therapy (13). BMSCs may be stably transfected in
order to overexpress exogenous genes. According to a previous
experiment, transfected BMSCs are capable of differentiating into
endodermal and ectodermal cells (14). It has also been reported that BMSCs
transplanted into neonatal mice brain may differentiate into
neurons and glial cells (15–19).
However, the differentiation rate of BMSCs into neuron-like cells
is much lower, as compared with other types of differentiated
cells; therefore, the present study aimed to increase the
efficiency of BMSC neural differentiation in vitro (14–17).
There are numerous chemical reagents and cytokines
widely used to induce the differentiation of neural BMSCs in
vitro, such as dimethyl sulfoxide, β-mercaptoethanol, vitamin
C, insulin, valproic acid, forsklin, hydrocortisone and NT-3
retinoic acid (18). Epidermal
growth factor (EGF) and bFGF are powerful mitogens and cell fate
drivers for neural precursor cells (19,20),
whereas bFGF may maintain a neuro-genic microenvironment in
vivo (21).
NGF is a type of neurotrophin, which exerts an
anti-apoptotic function in premature neurons (13). Based on effective biological
activation, NGF is associated with the neural differentiation and
migration of neural cells. In addition, NGF can protect axons and
myelin from inflammatory damage in order to modulate the immune
system, as well as protect and enhance excitotoxicity during
inflammatory activation. It has been demonstrated that NGF can
induce BMSC differentiation into neural cells, via generating
neuropeptide signals and receptors (6). These findings suggest that NGF is
essential for BMSC neural differentiation, which may be beneficial
for the treatment of injured nerves. The present study used NGF and
bFGF recombinant lentiviral vectors to transfect BMSCs in
vitro, and the neural differentiation efficiency of the BMSCs
was investigated. The aim of the present study was to observe the
differentiation of the co-transfected BMSCs into neuron-like cells
in vitro.
Materials and methods
Cell isolation and harvest
BMSCs were isolated and harvested according to a
previous method (22–24). Briefly, male and female SD rats
(age, 2–3 weeks, weighing 80–100 g) were sacrificed. Their bone
marrows were harvested by flushing the marrow cavities with 5 ml
Dulbecco's modified Eagle's medium (DMEM)/F12 supplemented with 10%
fetal bovine serum (FBS; Hyclone, GE Healthcare Life Sciences,
Logan, UT, USA) and 1% penicillin/streptomycin. The marrow cavities
were then rinsed two to three times until white. The sample was
transferred to 15 ml sterile centrifuge tubes, which were
centrifuged at 300 × g for 5 min. The precipitate was resuspended
in fresh DMEM/F12 (Hyclone) medium containing 10% FBS (Hyclone),
penicillin (100 U/ml (Hyclone), and streptomycin (100 g/l;
Hyclone). Subsequently, the cells were seeded at a density of
1×104 in 10 cm2 flasks (Falcon Plastics,
Oxnard CA, USA) and cultured at 37°C in a humidified atmosphere
containing 5% CO2. After 3 days, the medium was replaced
and non-adherent cells were removed. Spindle-shaped adherent MSCs
were expanded and purified with 3–5 passages after initial plating.
The medium was then replaced and the cells were observed under an
Olympus 81 inverted phase contrast microscope (Olympus Corporation,
Tokyo, Japan) every 3 days. After 12 days, spindle-shaped adherent
BMSCs were harvested with 0.25% trypsin (Hyclone, GE Healthcare
Life Sciences) and subcultured once they reached 80–90% confluence.
The third or fourth generation BMSCs were digested using 0.25%
EDTA-trypsin and seeded in six well plates containing poly-L-lysine
coverslips at a density of 1×106 cells/well. This study
was approved by the ethics committee of the Institutional Animal
Care and Use Committee of Dalian Medical University (Dalian,
China).
Flow cytometric analysis
For flow cytometry of the isolated BMSCs, BMSC cell
markers, including CD29, CD34, CD44, CD45, CD71, CD90 and CD106 (BD
Biosciences, Franklin Lakes, NJ, USA) were considered positive,
whereas CD45 was considered negative. The following mouse
monoclonal CD antibodies were conjugated with fluorescein
isothiocyanate (FITC; BD Biosciences): Anti-CD29 (eBioscience, San
Diego, CA, USA; cat. no. 12–0291); -CD34 (eBioscience; cat. no.
12–0341); -CD44 (eBioscience; cat. no. 12–0441) -CD45 (eBioscience;
cat. no. 11–0451), -CD71 (eBioscience; cat. no. 11–0711), -CD90
(eBioscience; cat. no. 11–0900); and -CD106 (eBioscience; cat. no.
11–4321). The surfaces of the fourth passage BMSCs were stained;
BMSC suspensions (1×105) were mixed with 10 µl
FITC, and incubated in a dark room at 4°C for 30 min. The cells
were then analyzed using a flow cytometer (BD FACSDiva Software
version 6.1.3; BD ARIA II and Diva software; BD Biosciences).
Plasmid construction and
transduction
Lentiviral-plasmids encoding human NGF (NM_002506.2)
and human bFGF2 (NM_002006.4) (Cyagen Biotechnology, Co., Ltd.,
Guangzhou, China) were generated by PCR amplification and subcloned
into the pLV.Des3d.P/puro expression vector (Invitrogen Life
Technologies, Carlsbad, CA, USA). The plasmids pLV.
ExSi.P/Puro-CMV-NGF-T2A, pLV.ExSi.P/Puro-CMV-FGF2-P2A,
pLV.ExSi.P/Puro- CMV-NGF-T2A-FGF2-P2A and pLV.ExSi.P/Puro-CMV were
constructed as previously described (25). Expression plasmids were transfected
into the 293T cells (Cyagen Biosciences, Sunnyvale, CA, USA) using
Lipofectamine® 2000 (Invitrogen Life Technologies),
according to the manufacturer's instructions. Briefly,
overexpression of the specific delivered gene was generated using a
lentiviral system (Sigma-Aldrich, St. Louis, MO, USA), according to
the manufacturer's instructions. Briefly, to generate the specific
NGF- and bFGF-containing lentiviruses, 1,077×105 293T
cells were co-transfected with pC9MV-VSV-G (15 µg), PRSV-Rev
(15 µg) and pMDLg/pRRE (15 µg). Transduction of 293T
cells with compatible packaging plasmids and 15 µg plv
plasmids was conducted in 10-cm2 flasks for 24 h using
Lipofectamine® 2000 transduction reagent (100 µl)
diluted in OPTI-MEM (1.5 ml; Thermo Fisher Scientific, Inc.,
Waltham, MA, USA). Following 16 h of transduction, the media was
replaced with OPTI-MEM containing 10 mM sodium butyrate. At 24 h
post-transduction, the media was replaced with viral harvesting
medium. Subsequently, the viruses were harvested and the media was
replaced with viral harvesting medium.
Harvesting of viruses and titration of
lentiviral vectors
BMSCs (3×105 cells) were seeded in a 6 cm
plate and incubated at 37°C for 24–48 h (5% CO2) until
the cells reached 30–50% confluence. The medium was then replaced
with serum-free medium, and transduction was performed 2 h later.
Briefly, the medium was removed and 1 ml virus-containing
supernatant and 1 µl polybrene (8 mg/ml; Santa Cruz
Biotechnology Inc., Dallas, TX, USA) was added with 3 ml complete
medium, and the cells were incubated for 2 h. The cells were then
allowed to grow for 2 days.
Western blotting
In order to detect the protein expression levels of
the two genes western blotting was performed. The transfected BMSCs
differentiated at various passages were lysed in
radioimmunoprecipitation buffer for 72 h. The proteins were then
extracted from SDS-PAGE and transferred to polyvinylidene
difluoride membranes, which were incubated with 5% skim-milk in
phosphate-buffered saline (PBS) for 1 h at 37°C to reduce
non-specific binding. The membranes were washed with PBS containing
0.1% Tween-20, and were incubated with antibodies targeting NGF
(1:500; cat. no. sc-33602; Santa Cruz Biotechnology, Inc.), bFGF
(1:1,000; cat. no. sc-79; Santa Cruz Biotechnology, Inc.),
β-tubulin III (1:500; cat. no. MFCD01323870; Sigma-Aldrich), GAPDH
(1:5,000; cat. no. sc-25778; Santa Cruz Biotechnology, Inc.), Akt
(1:1,000; cat. no. 9272; Cell Signaling Technology, Inc., Danvers,
MA, USA), phosphorylated (p)-Akt (1:1,000; cat. no. 9271; Cell
Signaling Technology, Inc.), extracellular-signal regulated kinases
(ERK; 1:1,000; cat. no. 9102; Cell Signaling Technology, Inc.), and
p-ERK (1:1,000; cat. no. 9101; Cell Signaling Technology) overnight
at 4°C. The membranes were then incubated with the corresponding
secondary antibodies. The blots were developed using enhanced
chemiluminescence western blotting reagents, and were visualized
using the Bio-Rad Image Lab system (Bio-Rad Laboratories, Inc.,
Hercules, CA, USA).
BMSC groups
The BMSCs were divided into five groups: The control
group, which consisted of untransfected BMSCs; the
plv-blank-transfected BMSCs group; the plv-bFGF-trans-fected BMSCs
group; the plv-NGF-transfected BMSCs group; and the plv-NGF-bFGF
co-transfected BMSCs group. All of the groups were cultured in
DMEM/F12 (Thermo Fisher Scientific, Inc.) supplemented with 20%
FBS, 1% penicillin and streptomycin (100 U/ml). When the cells had
proliferated and reached 80% confluency, the differentiation time
was between 1 and 7 days until the next passage. Subsequently, each
group was fixed for immuno cytochemistry.
Detection of neuron-like cells induced
from transfected BMSCs using immunofluorescence
Transfected fourth passage BMSCs were digested with
0.25% EDTA-trypsin. The cells (1×106) were then seeded
in 6-well plates containing poly-D-lysine-coated cover-glass, until
all of the cells were induced to neuronally differentiate. After 72
h, the cells were washed with PBS. The plates were then treated
with 1 ml 4% paraformaldehyde for 15 min, washed with PBS and fixed
with 3.7% formaldehyde in PBS for 25 min. Subsequently, the plates
were rinsed with PBS, and permeated with 0.1% Triton X-100
(Solarbio Beijing China) in 1 ml PBS for 20 min. The plates were
then washed with PBS for 1 h, blocked with 1% bovine serum albumin
in PBS for 1 h, and rinsed three times in PBS, each for 10 min. The
plates were incubated overnight with primary anti-nestin (1:200;
Santa Cruz Biotechnology, Inc.), anti-glial fibrillary acidic
protein (GFAP; 1:200; Wuhan Boster Biological Technology, Ltd.,
Wuhan, China), anti-neuron specific enolase (NSE; 1:200; Wuhan
Boster Biological Technology, Ltd.), and anti-β-tubulin III (1:500;
Sigma-Aldrich) in blocking solution for 2 h. The slides were washed
with blocking solution three times for 10 min each time, and the
plates were incubated for 2 h with secondary antibodies conjugated
to Alexa Fluor 488 (2 µg/ml, Invitrogen Life Technologies)
in blocking solution. Subsequently, the plates were washed with
blocking solution three times for 10 min each, and were incubated
with 1 µg/ml DAPI (Invitrogen Life Technologies) in blocking
solution for 2 min. The membranes were washed a further three times
with PBS and dried, and the cells were observed under a
fluorescence microscope (DP73; Olympus Corporation). β-tubulin III
was observed using a confocal laser scanning microscope (TCS SP5
II; Leica Microsystems GmbH, Wetzlar, Germany).
Statistical analysis
Data are presented as the mean ± standard deviation.
Standard analyses were carried out using an independent-samples
t-test. Experimental data were analyzed by SPSS version 19.0 (IBM,
Armonk, NY, USA). P<0.05 was considered to indicate a
statistically significant difference.
Results
Identification of BMSCs isolated from SD
rats
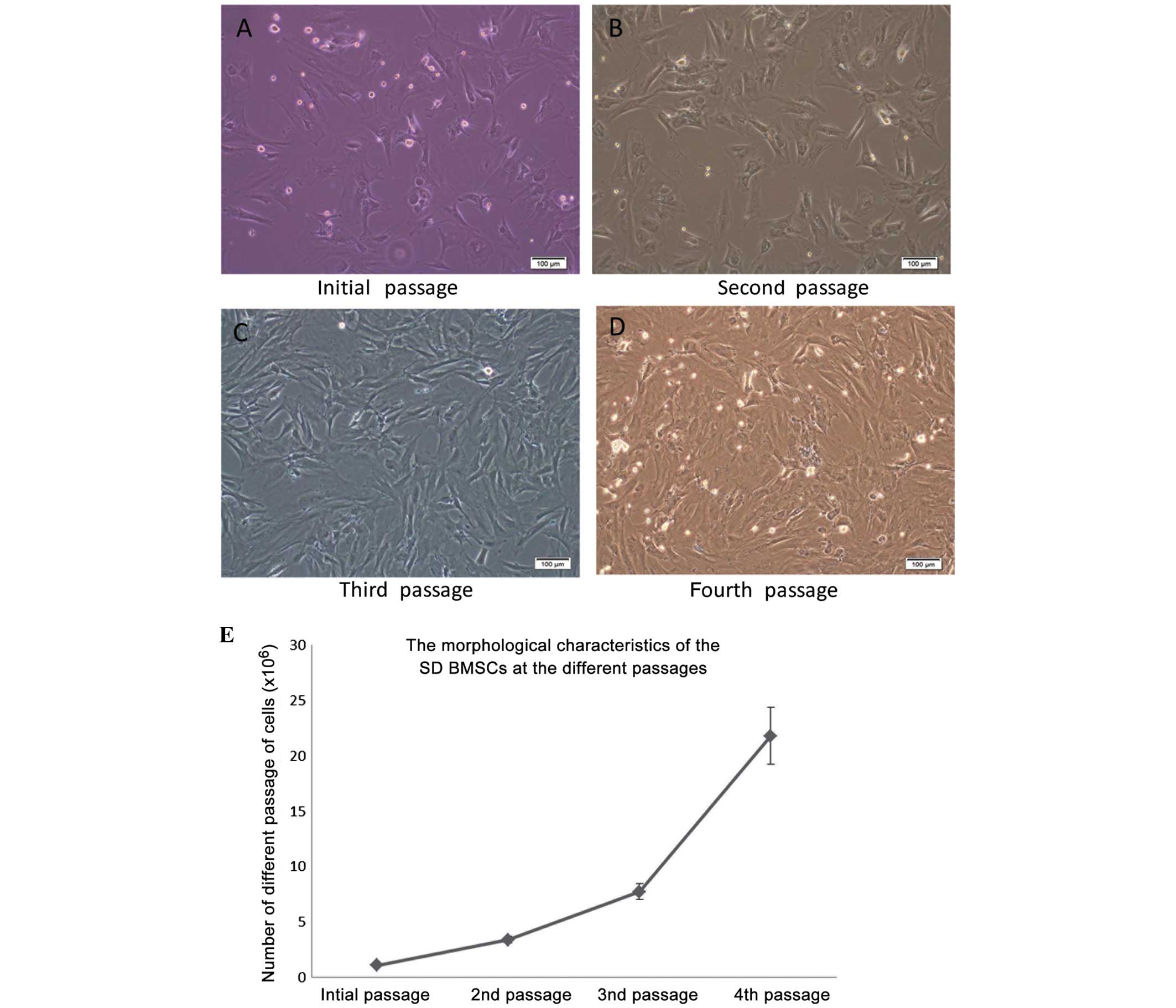
The morphology of BMSCs isolated from SD rats was
altered after the initial passage, as observed under an inverted
microscope (Fig 1). As shown in
Fig 1, the primary BMSCs were
oval-like and spindle-shaped, and some adherent fibroblast cells
were observed. Synaptic-like morphological changes occurred in the
cells, which gradually fused with each other following the
replacement of culture medium over the following days. As compared
with the initial cells, the second passage cells appeared larger,
and as they proliferated they extended from a triangle to polygon
shape. During the second passage, the rate of proliferation was
increased and the cells were shown to contain 1–2 visible round and
large nucleoli. While the cells are found to be integrated
minimally with the surrounding for shape and distribution, the
proliferation rate was significantly increased, as compared with in
the original cells. The BMSCs were especially easy to subculture
during the third passage. At the fourth passage increased cell
proliferation, polygonal cell morphology and long fusiform growth
were observed.
Identification of BMSCs
In previous studies, numerous antigenic markers have
been reported to be expressed in BMSCs rather than a single
specific antigenic marker (24).
According to previous studies (24), CD29 and CD44 are regarded as the
common positive antigen markers that may be used to identify mouse
BMSCs. In the present study, even more CD markers were analyzed, in
order to identify fourth passage BMSCs. The results of the flow
cytometric analysis on fourth passage BMSCs using CD29, CD44, CD90,
CD105, CD34, CD45, and CD106 monoclonal cell surface antibodies are
shown in Fig. 2A. Fig. 2B presents the percentage of
positively-stained cells for each antigen marker: CD29
(99.25±0.82%), CD34 (1.00±0.20%), CD44 (83.73±4.95%), CD45
(0.98±0.19%), CD71 (89.86±2.39%), CD90 (99.21±0.79%) and CD106
(86.42±5.67%). These results suggested that the extracted BMSCs
were high-purity, easily harvested, and stable.
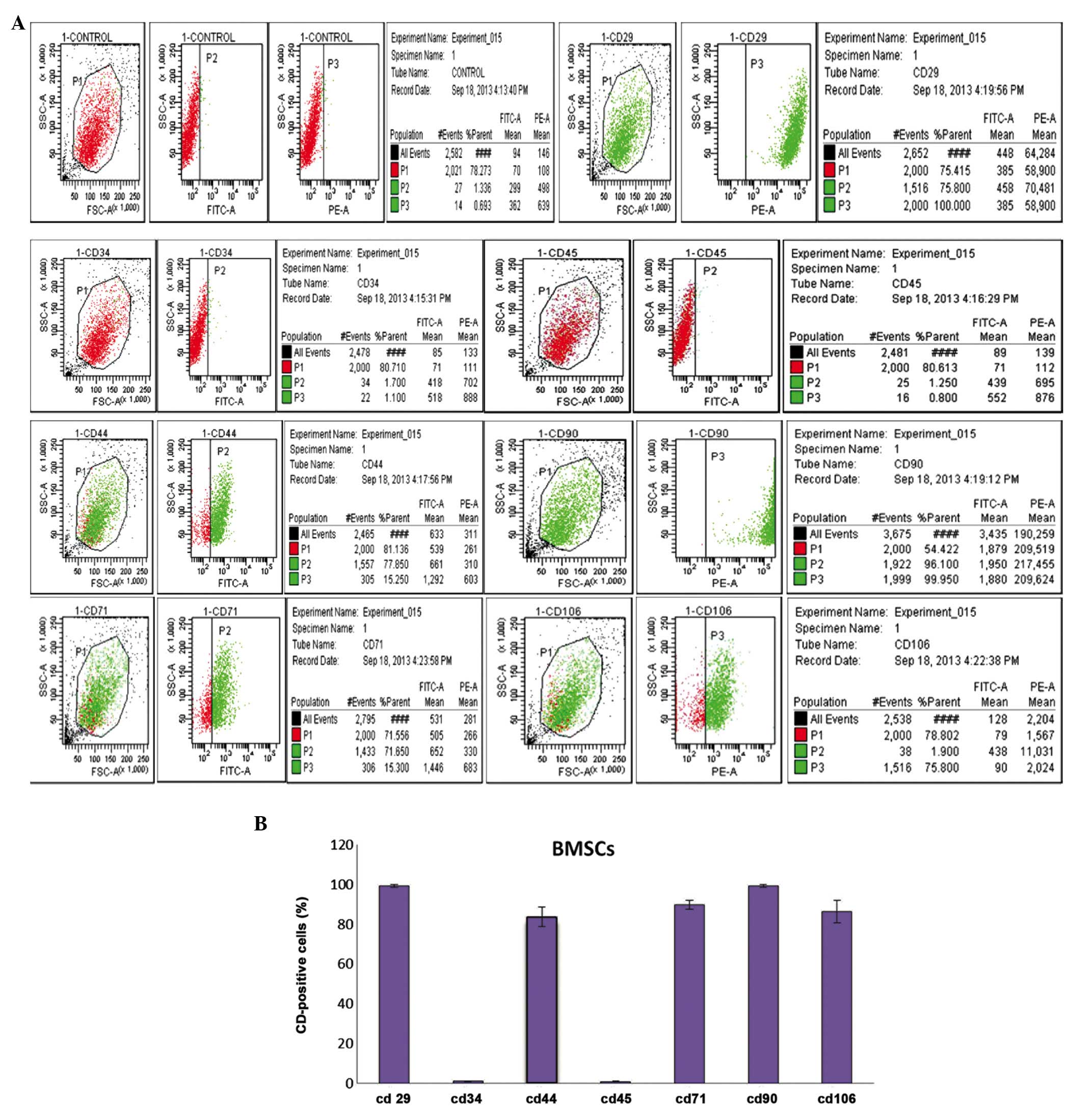 | Figure 2Identification and immunophenotypic
characterization of bone marrow mesenchymal stem cells (BMSCs)
isolated from Sprague Dawley rats, as determined by flow cytometric
analysis of surface antigens. (A) The cells were harvested and
stained with fluorescein isothiocyanate-conjugated anti-rat CD29,
CD34, CD44, CD45, CD71, CD90, CD106. The BMSCs were negative for
CD34, CD45, but positive for mesenchymal markers CD29, CD44, CD71,
CD90 and CD106. (B) Statistical analysis of numerous samples.
Fourth passage BMSCs CD marker expression was as follows: CD29
(99.25±0.82%), CD34 (1.00±0.20%), CD44 (83.73±4.95%), CD45
(0.98±0.19%), CD71 (89.86±2.39%), CD90 (99.21±0.79%), CD106
(86.42±5.67%). |
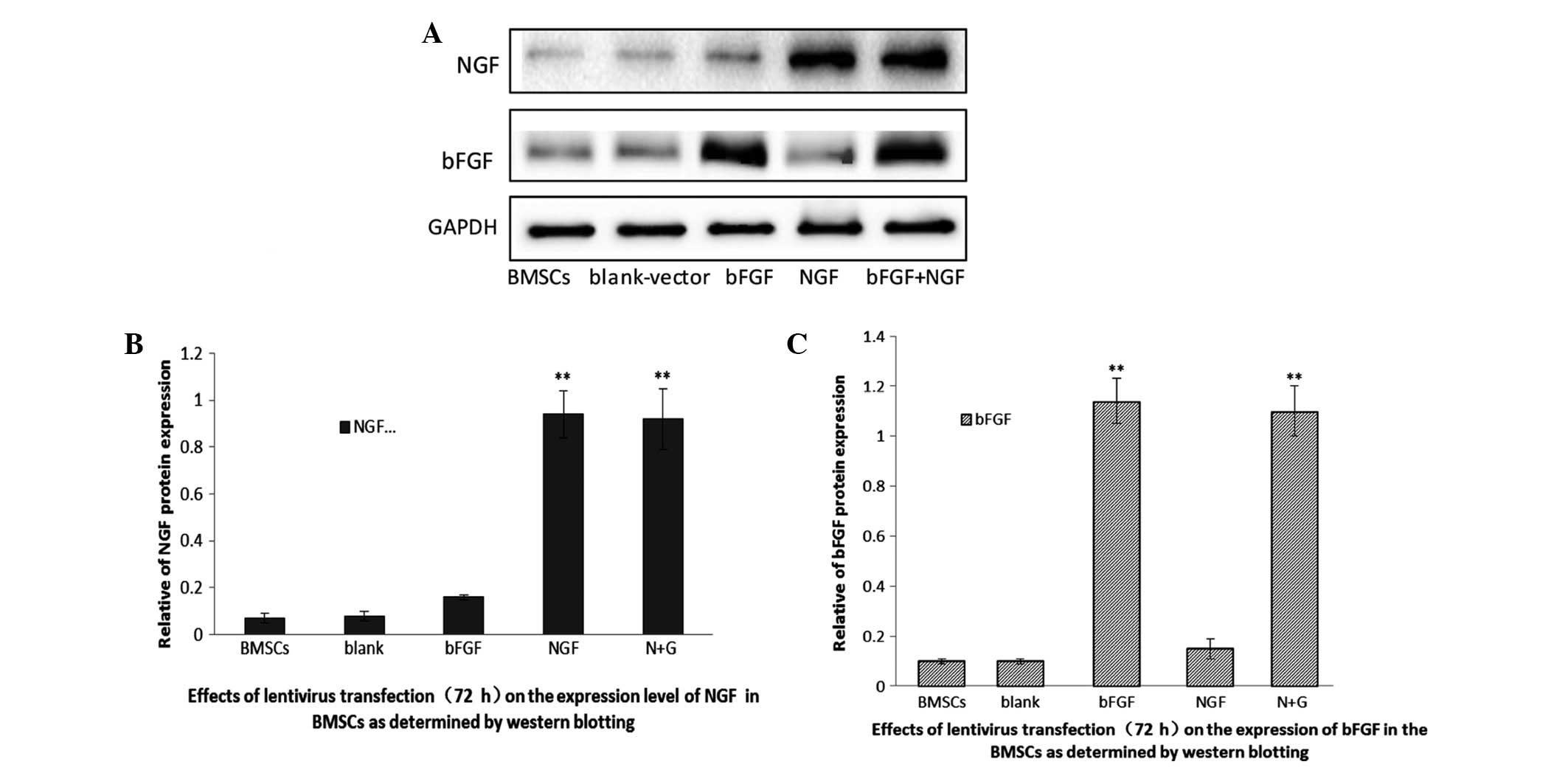
Transduction efficiency of NGF and bFGF
lentiviral vectors into BMSCs
A total of 72 h post-transduction, the transduction
efficiency of bFGF and NGF into BMSCs was determined. Western
blotting was used to detect the protein expression levels of the
two genes in the BMSCs. As shown in Fig. 3, untransfected BMSCs express little
NGF and bFGF, thus suggesting that BMSCs rarely secrete the two
proteins. Conversely, the two proteins were significantly
overexpressed in the BMSCs transfected with the lentiviral vectors.
These results indicated that the expression levels of NGF and bFGF
may be increased in BMSCs transfected with specific lenti-viral
vectors.
Differentiation of co-transfected
BMSCs
BMSC morphology was observed 72 h post-transduction.
The cells were retracted towards their flat cell nucleus forming a
compressed multipolar spherical cell body, producing membranous,
process-like extension peripherally. These cells have a
significantly higher refractive index difference than the neuronal
cell bodies involved in the process, similar to the termination
structure of the growth cone. The expansion of putative filopodia
demonstrated that the transfected BMSCs have the potential to
differentiate into neural-like cells. The number of cells and the
neuronal morphology were both increased, and the cell synapse
gradually formed over the 72 h. Conversely, the untransfected BMSCs
remained spindle-shaped. Subsequently, nestin (a neural stem cell
marker), NSE (a neuronal marker), and GFAP (an astrocyte marker)
were used to identify differentiated neural cells (Fig. 4A). Green immunofluorescence
intensity was used to detect the various neuron markers. NGF and
bFGF co-transfected BMSCs exhibited higher intensities of green
fluorescence. Western blotting detected the protein expression
levels of NSE, nestin and GFAP in the various groups (Fig. 4B). Notably, relative protein
expression levels were higher in the bFGF and NGF co-transfected
group, as compared with the NGF-transfected group or the
bFGF-transfected group (Fig. 4C).
Positive staining rates were 8.1±1.7%, 28.57±6.29% and 54.29±5.71%
for nestin; 11±2.66%, 40.14±5.81% and 69.29±5.82% for NSE; and
6.43±2.15% and 15.29±2.81%, for GFAP. In addition, the
co-transfected BMSCs and NGF-transfected BMSCs possessed increased
numbers of differentiating neuron-like cells, as compared with the
bFGF-transfected group (Fig.
4D).
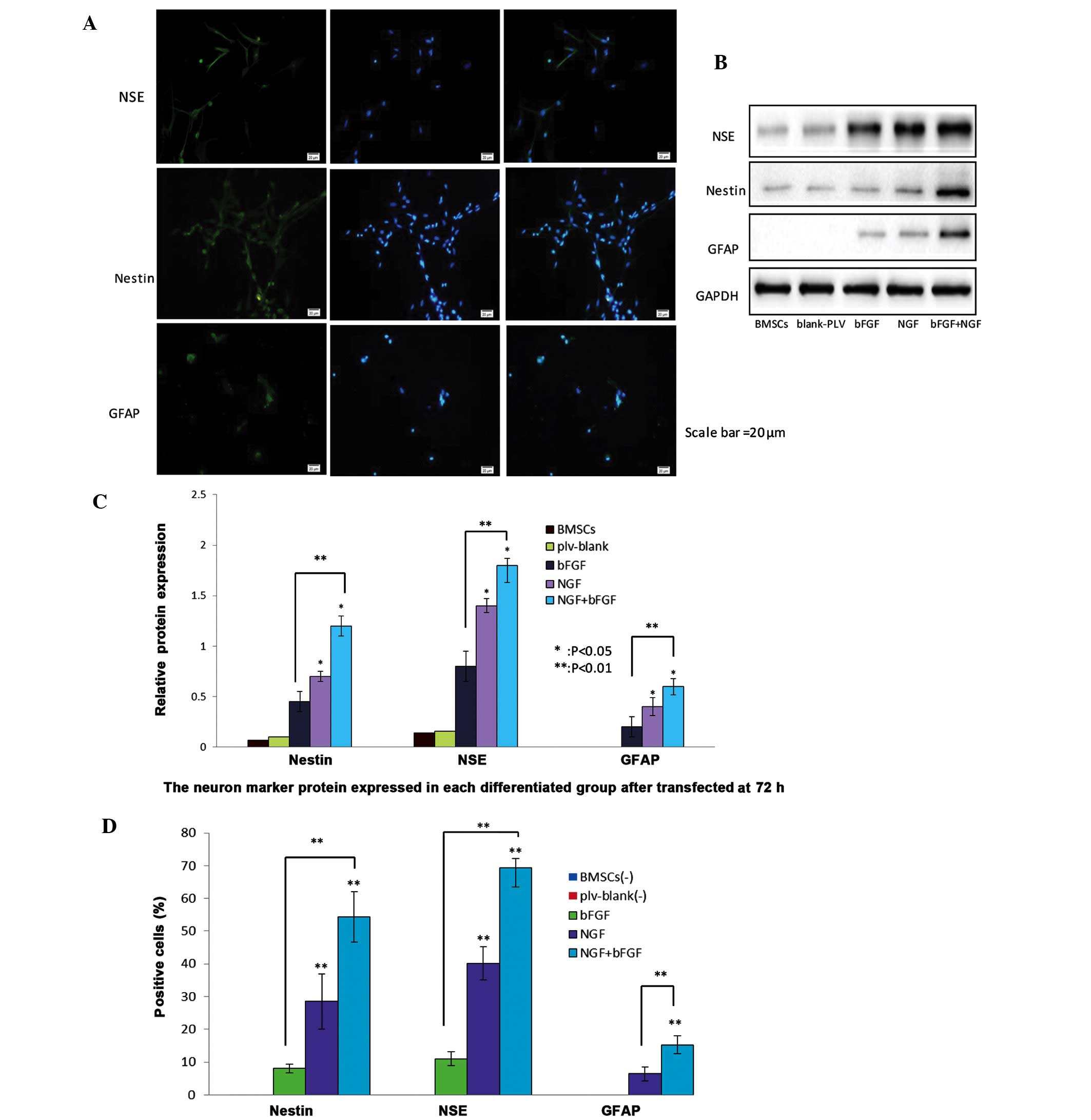 | Figure 4Protein expression levels of nestin,
neuron-specific enolase (NSE) and glial fibrillary acidic protein
(GFAP) neuron-markers in nerve growth factor (NGF)- and basic
fibroblast factor (bFGF)-transfected bone marrow mesenchymal stem
cells (BMSCs) 72 h post-transfection. (A) Differentiated cells were
stained with nestin, NSE and GFAP antibodies and counterstained
with DAPI (blue). Scale bar=20 µm. The NGF and bFGF
co-transfected BMSCs exhibited increased protein expression of
nestin, NSE and GFAP, as demonstrated by a marked increase in green
fluorescence after 72 h, detected by fluorescence microscopy. (B)
Western blotting detected the protein expression levels of neuron
markers in the transfected BMSCs after 72 h. The expression levels
of the neuron markers were higher in the co-transfected BMSCs
group, as compared with the other groups. The untransfected BMSCs
and plv-blank-transfected cells exhibited a slight expression of
neuron markers. (C) Western blot analysis of the protein expression
levels of nestin, NSE and GFAP in the untransfected BMSCs group,
plv-blank-transfected BMSCs group, bFGF-transfected BMSCs group,
NGF-transfected BMSCs group, and NGF and bFGF co-transfected BMSCs
group. Data show the protein levels normalized to GAPDH, and are
presented as the mean ± standard deviation (SD) (n=5).
*P<0.05, co-transfected group vs. NGF-transfected
group, and NGF-transfected group vs. bFGF-transfected group;
**P<0.01, co-transfected group vs bFGF-transfected
group.(D) Percentage of neural cells positive for nestin, NSE and
GFAP in the bFGF-transfected BMSCs group, NGF-transfected BMSCs
group, and co-transfected BMSCs group. Data are presented as the
mean ± SD (n=5). **P<0.01, each group was
significantly different, as compared with the other groups. |
Expression levels of β-tubulin III in
differentiated neuron-like cells at various passages
β-tubulin III, which is a component of the
cytoskeleton, is a neuronal specific tubulin considered to be an
indicator of early neuronal development (23–26).
Under laser scanning confocal microscopy, early neuronal-like cells
were identified in the initial passage of the transfected cells, as
they began to differentiate. When the cells had differentiated to
the second passage, weak green fluorescence, indicating β-tubulin
III expression, gathered in the perinuclear area and extended to
form a network structure. When the cells had differentiated to the
fourth passage, the presence of green fluorescent reactants was
significantly increased, indicating a higher expression of
β-tubulin III. In addition, strong peri-nuclear expression of
β-tubulin III was also detected. After 6 passages, the cell
structure was clearer, and the fluorescence intensity surrounding
the nucleus at the synapse was stronger. At the eighth passage,
nuclear green fluorescence was decreased, and extrusion was
detected (Fig. 5A). In the various
transfected BMSC groups, from passage 2–8, the protein expression
of β-tubulin III was initially increased, and was then decreased.
In the control BMSCs and plv-blank-trans-fected BMSCs the protein
expression of β-tubulin III was absent, whereas in the
bFGF-transfected group, the protein expression of β-tubulin III was
detected, but at small levels. The protein expression levels of
β-tubulin III were higher in the fourth and sixth passages, as
compared with in the second and fourth passages, respectively, of
BMSCs. Although in the NGF- and bFGF- transfected BMSCs, the
protein expression levels increased from passage 2–6, at passage 8,
the protein expression levels were markedly lower. The differences
between the two groups were statistically significant (Fig. 5B).
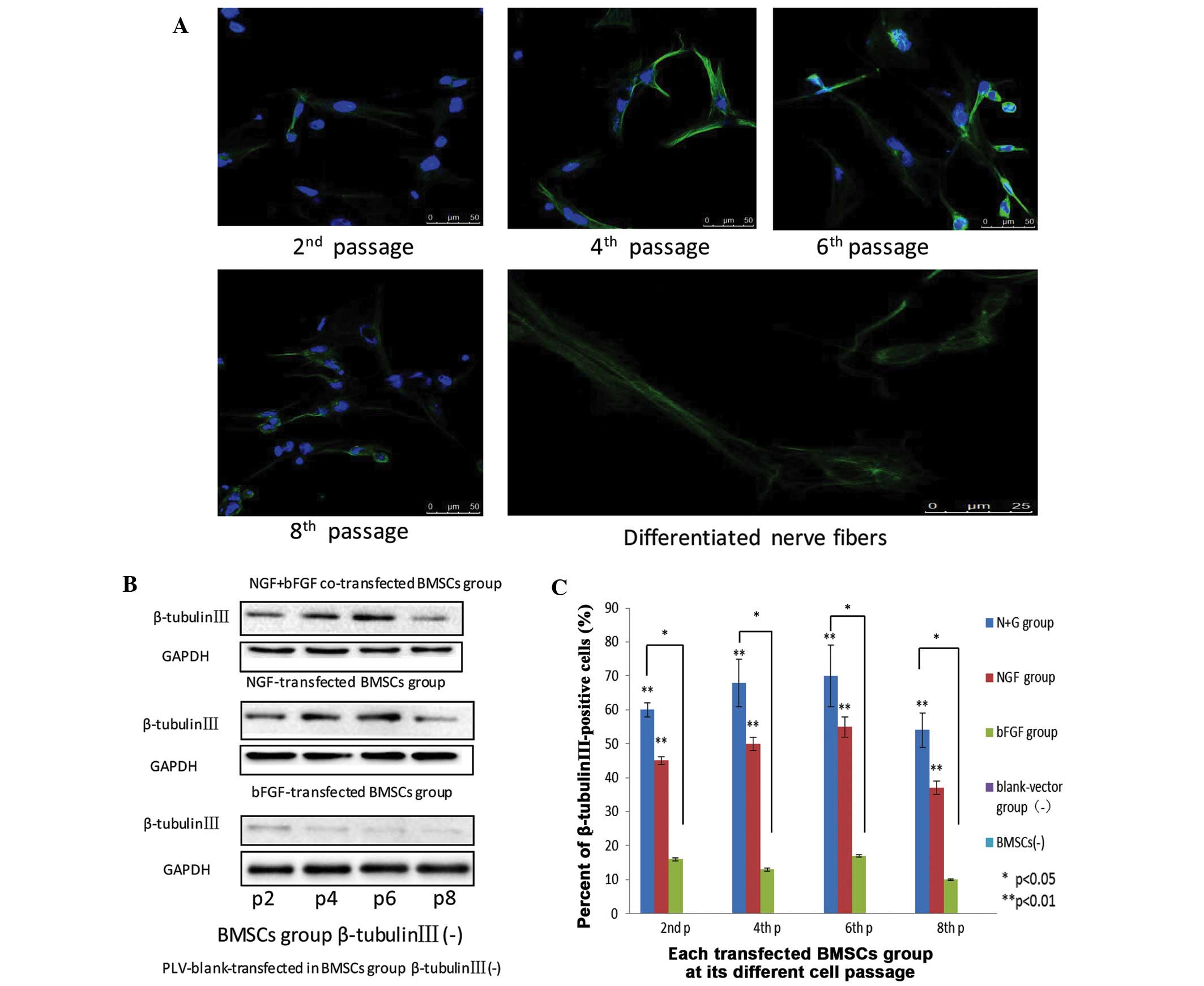 | Figure 5(A) Nerve growth factor (NGF) and
basic fibroblast growth factor (bFGF) co-transfected bone marrow
mesenchymal stem cells (BMSCs) at passages 2, 4, 6, and 8 express
β-tubulin III, as detected by the presence of green fluorescent
dye. Scale bar=50 mm. Neuron-like extensions and inter-connections
of NGF and bFGF were depicted. Nerve fibers scale bar=25 mm. All
images were obtained with a laser scanning confocal microscope
(Zeiss LSM 510). (B) The expression of β-tubulin III in the various
groups of differentiated cells at passages 2, 4, 6, and 8. (C) Flow
cytometric analysis of β-tubulin III expression in the three groups
at passages 2, 4, 6, and 8. Data are presented as the mean ±
standard deviation (n=3). *P<0.05,
**P<0.01. |
At the second passage, in the NGF and bFGF
co-transfected BMSCs group, the reaction rate with β-tubulin III
was 61.40±1.89%; in the NGF-transfected BMSCs group the reaction
rate was 45±1.2%; and in the bFGF-transfected BMSCs group the
reaction rate was 16.10±0.5%. At the fourth passage, the reaction
rate with β-tubulin III was 68±7.20%, 50±2.10%, and 13±0.50% in the
co-transfected, NGF-transfected, and bFGF-transfected cells,
respectively. At the sixth passage, the reaction rate with
β-tubulin III was 70.40±9.24%, 55±3.60% and 17±3.80% in the
co-transfected, NGF-transfected, and bFGF-transfected cells,
respectively. In addition, at the eighth passage, the reaction rate
with β-tubulin III was 54.57±5.15%, 37.18±1.27% and 11.28±0.26% in
the co-transfected, NGF-transfected, and bFGF-transfected cells,
respectively (Fig. 5C). These
results suggested that β-tubulin III expression was highest in the
co-transfected BMSCs, as compared with the other groups.
Furthermore, co-transfected BMSCs may differentiate into more
sophisticated neuron-like cells, as compared with the remaining
groups.
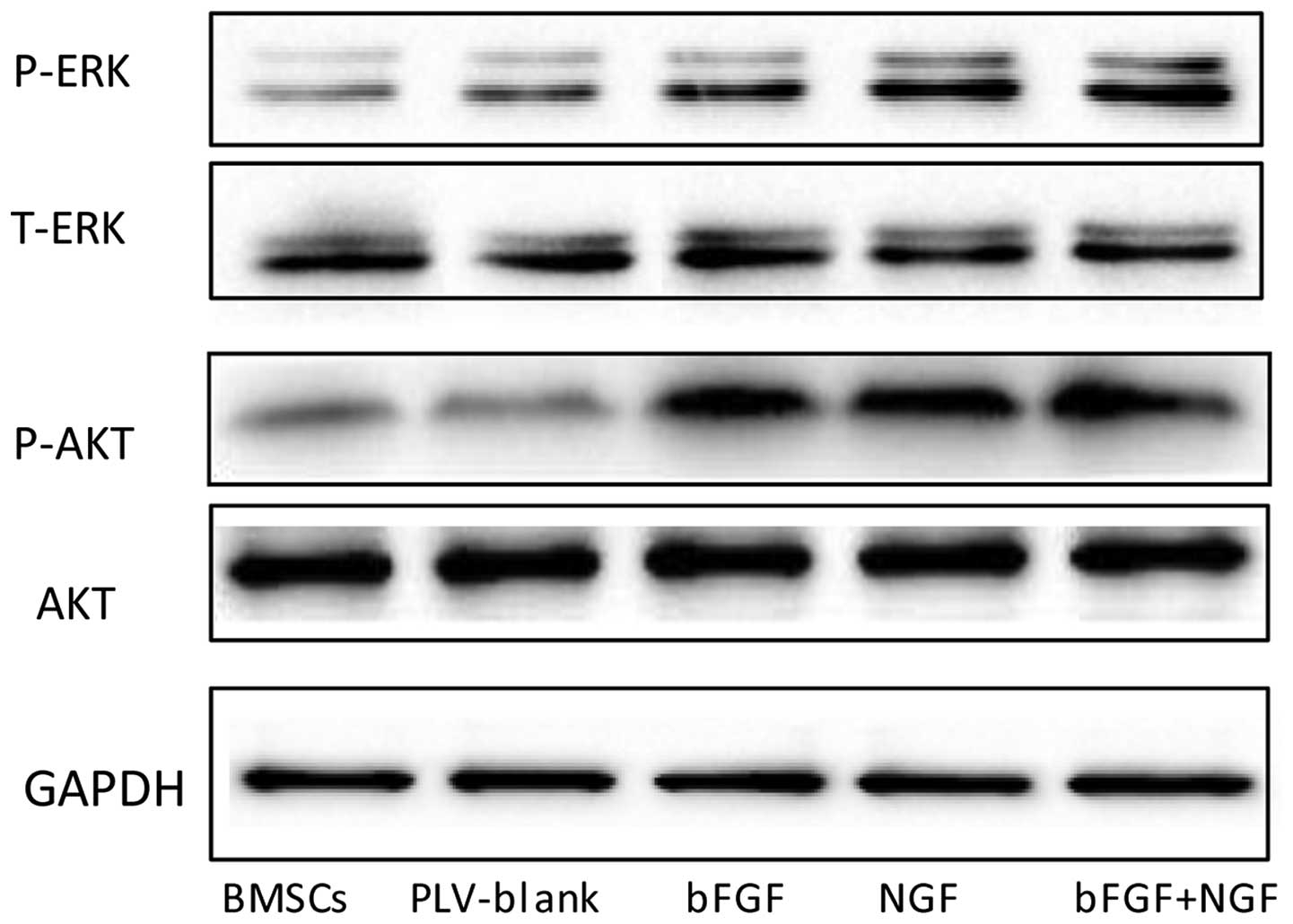
AKT and ERK signaling pathways are
involved in the neural differentiation of transfected BMSCs
MAPK has a significant role in the growth and
differentiation of BMSCs. In the present study, western blotting
was used to examine the protein expression levels of ERK in the
various groups. As shown in Fig.
6, the expression levels of p-ERK differed between the groups.
The expression levels of p-ERK were increased in the
NGF-transfected and co-transfected groups, and no significant
changes were detected between the expression of p-ERK in the BMSCs
and plv-blank-transfected BMSCs. These results indicated that the
MAPK pathway may manipulate the neuronal cell differentiation of
BMSCs. Transduction of BMSCs with bFGF and NGF enhanced the
phosphorylation of ERK (Fig. 6).
In addition, the western blot analysis demonstrated that NGF- and
bFGF-transfected BMSCs exhibited increased expression levels of AKT
phosphorylation. However, no significant differences in the
expression levels of AKT were detected between the control BMSCs
and blank-vector-transfected BMSCs. These results suggested that
AKT may also be involved in BMSCs neural differentiation.
Discussion
BMSCs have the potential of multi-directional
differentiation (22). BMSCs have
been reported to exert beneficial effects for use in clinical
treatment and cell regeneration, and may be regarded as an advanced
and promising therapeutic tool (23,27,28).
Neurotrophins have a predominant role in development and
differentiation, and stimulate the tightly regulated process of
BMSC neural differentiation. In order to take advantage of BMSCs
and neurotrophins for nerve cell regeneration, it is necessary to
research the specific mechanisms underlying neural differentiation.
Delcroix et al (27)
demonstrated that pre-treatment with bFGF was able to enhance
neural specification, and Fan et al (28) reported that NGF and vascular
endothelial growth factor enhance angiogenic effects in
vivo.
It is convenient to use BMSCs for the treatment of
injured tissue. Conversely, neurotrophic factors secreted by
transfected BMSCs are beneficial for the restoration of injured
tissue. BMSCs are able to promote survival of grafted cells, and
also secrete a sufficient amount of mature neurotrophic
factors.
bFGF is expressed in the embryonic and adult central
and peripheral nervous systems, and maintains the survival of
neuronal and glial cells, promotes sympathetic and parasympathetic
nervous axon growth, and promotes the repair of damaged nerves and
neurite outgrowth (19). In
addition, bFGF is able to induce differentiation of adrenal
pheochromocytoma cells-12 (29),
and extend human neural progenitor cell nerve regeneration and
neural differentiation (7).
Therefore, bFGF is considered an important growth factor. NGF is a
significant neuropeptide signal, which can induce BMSC neural
differentiation, and regulate cell proliferation and cell survival
growth (7). Lentiviral vectors may
be used to stably transfect cells, and genes integrated into the
genome of target cells using lentiviral vectors exhibit long-term
expression and immune response.
The present study constructed NGF and bFGF
co-transfected SD rat BMSCs, in order to determine the effects of
overexpression of these two genes on differentiation into
neuron-like cells. The results of the present study demonstrated
that NGF and bFGF co-transfection was able to induce the
differentiation of BMSCs into neuron-like cells; the co-transfected
group had the highest proportion of neuron-like cell
differentiation, as compared with the other groups.
In the present study, BMSCs of stable growth were
selected as cells to be transfected. Once the NGF and bFGF genes
were successfully recombined with the lentiviral vector in
vitro, western blotting was used to detect the expression
levels of each of the proteins. In addition to spindle-shaped and
polygonal cell morphology, a series of positive CD-antigenic
markers was used for identification of successfully transfected
BMSCs through flow cytometry. In the control BMSCs and blank
vector-transfected cells a low expression of NGF was detected. This
finding indicated that, the lentivirus-vector transfection and bFGF
may enhance the BMSCs, while BMSCs can themselves secrete a small
quantity of NGF. These results indicated that transfection with the
two genes effectively increased the bFGF and NGF protein expression
levels in BMSCs.
The results of the present study also demonstrated
that the NGF and bFGF co-transfected cells exhibited much higher
neuron-marker expression, as compared with the other groups. In the
fourth and sixth passages, β-tubulin III protein expression was
detected, as mature neuronal microtubule structures were formed and
migration increased, due to the maturation of the cytoskeleton.
Intracellular microtubules of β-tubulin III consist of a long line
of aligned fibrils formed by a hollow tubular structure. The
results of the present study indicated that transfected BMSCs were
able to differentiate from the polar cells of spindle-shaped and
polygonal morphology into neuron-like cells. This process occurred
alongside increasing β-tubulin III expression.
β-tubulin III, which is a neuron-specific tubulin,
is a component of the cytoskeleton, and has been considered a sign
of early neuronal development. β-tubulin III has an important role
in the development of the neuron cyto skeleton during neural
differentiation and maturation (26,30).
In the present study, in the second passage BMSCs, β-tubulin III
expression was weak, and was mainly distributed in the nucleus,
during the continual development of neurons. When the cells reached
passages 4 and 6, β-tubulin III expression increased and extended
outwards. It is possible that the synthesized intracellular rough
endoplasmic reticulum gradually begins to participate in the
formation of microtubules, thus extending the projections of the
nucleus. However, as the neuron cell matured to the eighth
generation, β-tubulin III expression was reduced, and extension out
of the cells was decreased. Western blotting also demonstrated that
in early neural differentiation, β-tubulin III expression gradually
increased, and then decreased at the eighth passage. Due to the
intracellular distribution and content of β-tubulin III, it is
believed that β-tubulin III has an important role in the neural
differentiation of axonal growth. These results suggest that
neuronal development may be associated with microtubule
polymerization. In the early stages of neural differentiation, it
may be hypothesized that β-tubulin III promotes the formation of
microtubules, thereby affecting neural differentiation and neuron
outgrowth. In addition, NGF and bFGF may enhance the expression of
β-tubulin III.
The results of the present study provide compelling
evidence that NGF and bFGF have a critical role in the
high-efficiency differentiation of BMSCs into neuron-like cells. It
has been previously reported exogenous factor-induced
differentiation of MSCs, suggesting that the ERK pathway is
involved in the neural differentiation of BMSCs (27,31).
The present study demonstrates that NGF and BFGF co-transfected
BMSCs exhibited an increased expression of ERK phosphorylation in
BMSCs, as well as increased proliferation and differentiation.
These results indicated that MAPKs may regulate BMSCs, thus
enhancing phosphorylation in NGF and bFGF-transfected cells.
Concurrently, the AKT pathway may also be involved, which is able
to inhibit cell apoptosis. The results of the present study
demonstrated that NGF- and bFGF-transfected BMSCs exhibited
increased expression levels of p-AKT, whereas the control BMSCs and
blank-vector-transfected BMSCs exhibited lower expression of AKT
phosphorylation, thus suggesting that that AKT pathway is involved
in regulation of BMSC neural differentiation.
In conclusion, bFGF and NGF co-transfected BMSCs
exhibited enhanced neural specification and response to neuronal
commitment of stem cells, resulting in the induction of
differentiation and maturation of neuron cells. Furthermore, ERK
and AKT signaling pathways were shown to be involved in the neural
differentiation of NGF and bFGF co-transfected BMSCs. The present
study may influence the use of cell therapy to treat adult nervous
system diseases in the future.
Acknowledgments
The authors of the present study would like to thank
Quentin Liu Lab at the Dalian Medical University for their critical
demands and technical support. The present study was supported by
the National Natural Science Foundation of China (grant no.
81270052).
References
|
1
|
Turgeman G, Pittman DD, Müller R, Kurkali
BG, Zhou S, Pelled G, Peyser A, Zilberman Y, Moutsatsos IK and
Gazit D: Engineered human mesenchymal stem cells: A novel platform
for skeletal cell mediated gene therapy. J Gene Med. 3:240–251.
2001. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Mohammadi R, Azizi S, Delirezh N,
Hobbenaghi R, Amini K and Malekkhetabi P: The use of
undifferentiated bone marrow stromal cells for sciatic nerve
regeneration in rats. Int J Oral Maxillofac Surg. 41:650–656. 2012.
View Article : Google Scholar
|
|
3
|
Pereira Lopes FR, Camargo de Moura Campos
L, Dias Corrêa J Jr, Balduino A, Lora S, Langone F, Borojevic R and
Blanco Martinez AM: Bone marrow stromal cells and resorbable
collagen guidance tubes enhance sciatic nerve regeneration in mice.
Exp Neurol. 198:457–468. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Wu J, Yu W, Chen Y, Su Y, Ding Z, Ren H,
Jiang Y and Wang J: Intrastriatal transplantation of
GDNF-engineered BMSCs and its neuroprotection in
lactacystin-induced Parkinsonian rat model. Neurochem Res.
35:495–502. 2010. View Article : Google Scholar
|
|
5
|
Wen SR, Qi HP, Ren YJ, Liu GJ, Gong FC,
Zhong H and Bi S: Expression of δNp73 in hippocampus of APP/PS1
transgenic mice following GFP-BMSCs transplantation. Neurol Res.
33:1109–1114. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Ding J, Cheng Y, Gao S and Chen J: Effects
of nerve growth factor and Noggin-modified bone marrow stromal
cells on stroke in rats. J Neurosci Res. 89:222–230. 2011.
View Article : Google Scholar
|
|
7
|
Zhu H1, Yang A, Du J, Li D, Liu M, Ding F,
Gu X and Liu Y: Basic fibroblast growth factor is a key factor that
induces bone marrow mesenchymal stem cells towards cells with
Schwann cell phenotype. Neurosci Lett. 559:82–87. 2014. View Article : Google Scholar
|
|
8
|
Mohammadi R, Azizi S, Delirezh N,
Hobbenaghi R and Amini K: Comparison of beneficial effects of
undifferentiated cultured bone marrow stromal cells and omental
adipose-derived nucleated cell fractions on sciatic nerve
regeneration. Muscle Nerve. 43:157–163. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Wang S, Yaszemski MJ, Knight AM,
Gruetzmacher JA, Windebank AJ and Lu L: Photo-crosslinked
poly(epsilon-capro-lactone fumarate) networks for guided peripheral
nerve regeneration: Material properties and preliminary biological
evaluations. Acta Biomater. 5:1531–1542. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Crouzier T, McClendon T, Tosun Z and
McFetridge PS: Inverted human umbilical arteries with tunable wall
thicknesses for nerve regeneration. J Biomed Mater Res A.
89:818–828. 2009. View Article : Google Scholar
|
|
11
|
Cuevas P, Carceller F, Garcia-Gómez I, Yan
M and Dujovny M: Bone marrow stromal cell implantation for
peripheral nerve repair. Neurol Res. 26:230–232. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Liu WG, Wang ZY and Huang ZS: Bone
marrow-derived mesen-chymal stem cells expressing the bFGF
transgene promote axon regeneration and functional recovery after
spinal cord injury in rats. Neurol Res. 33:686–693. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Colafreancesco V and Villoslada P:
Targeting NGF pathway for developing neuroprotective therapies for
multiple sclerosis and other neurological diseases. Arch Ital Biol.
149:183–192. 2011.
|
|
14
|
Kopen GC, Prockop DJ and Phinney DG:
Marrow stromal cells migrate throughout forebrain and cerebellum,
and they differentiate into astrocytes after injection into
neonatal mouse brains. Proc Natl Acad Sci USA. 96:10711–10716.
1999. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Li Y, Chen J and Chopp M: Adult bone
marrow transplantation after stroke in adult rats. Cell Transplant.
10:31–40. 2001.PubMed/NCBI
|
|
16
|
Li Y, Chopp M, Chen J, Wang L, Gautam SC,
XU YX and Zhang Z: Intrastriatal transplantation of bone marrow
nonhematopoietic cells improves functional recovery after stroke in
adult mice. J Cereb Blood Flow Metab. 20:1311–1319. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Chopp M, Zhang XH, Li Y, Wang L, Chen J,
Lu D, Lu M and Rosenblum M: Spinal cord injury in rat: Treatment
with bone marrow stromal cell transplantation. Neuroreport.
11:3001–3005. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Maden M: Retinoic acid in the development,
regeneration and maintenance of the nervous system. Nat Rev
Neurosci. 8:755–765. 2007. View
Article : Google Scholar : PubMed/NCBI
|
|
19
|
Bithell A, Finch SE, Hornby MF and
Williams BP: Fibroblast growth factor 2 maintains the neurogenic
capacity of embryonic neural progenitor cells in vitro but changes
their neuronal subtype specification. Stem Cells. 26:1565–1574.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Sanchez-Ramos J, Song S, Cardozo-Pelaez F,
Hazzi C, Stedeford T, Willing A, Freeman TB, Saporta S, Janssen W,
Patel N, et al: Adult bone marrow stromal cells differentiate into
neural cells in vitro. Exp Neurol. 164:247–256. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Mudò G, Bonomo A, Di Liberto V, Frinchi M,
Fuxe K and Belluardo N: The FGF-2/FGFRs neurotrophic system
promotes neurogenesis in the adult brain. J Neural Transm.
116:995–1005. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Zhu H, Guo ZK, Jiang XX, Li H, Wang XY,
Yao HY, Zhang Y and Mao N: A protocol for isolation and culture of
mesen-chymal stem cells from mouse compact bone. Nat Protoc.
5:550–560. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Katsetos CD, Legido A, Perentes E and Mörk
SJ: Class III beta-tubulin isotype: A key cytoskeletal protein at
the crossroads of developmental neurobiology and tumor
neuropathology. J Child Neurol. 18:851–867. 2003. View Article : Google Scholar
|
|
24
|
Wang Z, Deng Q, Zhang X and Zhang J:
Treatment of injured neurons with bone marrow stem cells
cotransfected by hTERT and Ad-BDNF in vitro. J Mol Neurosci.
38:265–272. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Fan BS and Lou JY: Enhancement of
angiogenic effect of co-transfection human NGF and VEGF genes in
rat bone marrow mesenchymal stem cells. Gene. 485:167–171. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Tao YX, Xu HW, Zheng QY and FitzGibbon T:
Noggin induces human bone marrow-derived mesenchymal stem cells to
differentiate into neural and photoreceptor cells. Indian J Exp
Biol. 48:444–452. 2010.PubMed/NCBI
|
|
27
|
Delcroix GJ, Curtis KM, Schiller PC and
Montero-Menei CN: EGF and bFGF pre-treatment enhances neural
specification and the response to neuronal commitment of MIAMI
cells. Differentiation. 80:213–227. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Fan BS and Lou JY: Enhancement of
angiogenic effect of co-transfection human NGF and VEGF genes in
rat bone marrow mesenchymal stem cells. Gene. 485:167–171. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Tobin JE, Xie M, Le TQ, Song SK and
Armstrong RC: Reduced axonopathy and enhanced remyelination after
chronic demyelination in fibroblast growth factor 2 (Fgf2)-null
mice: Differential detection with diffusion tensor imaging. J
Neuropathol Exp Neurol. 70:157–165. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Fanarraga ML, Avila J and Zabala JC:
Expression of unphosphorylated class III beta-tubulin isotype in
neuroepithelial cells demonstrates neuroblast commitment and
differentiation. Eur J Neurosci. 11:516–527. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Lam HJ, Patel S, Wang A, Chu J and Li S:
In vitro regulation of neural differentiation and axon growth by
growth factors and bioactive nanofibers. Tissue Eng Part A.
16:2641–2648. 2010. View Article : Google Scholar : PubMed/NCBI
|