Introduction
The ubiquitin-proteasome (UPP) and autophagy
pathways are the two major pathways involved in the degradation of
intracellular proteins in eukaryotic cells. The UPP pathway
predominantly degrades short half-life proteins, whereas autophagy
degrades long half-life proteins and organelles (1). The proteasome pathway is key in
maintaining the homeostasis of intracellular protein metabolism,
thereby regulating cell cycle progression, cell differentiation and
DNA damage repair (2). Targeting
the proteasome pathway is a promising strategy for cancer therapy,
as evidenced by the effects of several proteasome inhibitors in
preclinical and clinical trials (3–5).
However, clinical trials have demonstrated that developing
resistance to proteasome-targeting drugs is a significant problem
(6).
Autophagy degrades proteins and organelles via the
lysosome, which enables cells to digest their intrinsic components
to obtain the energy and nutrients required to survive in adverse
microenvironments (7). Under
normal physiological conditions, autophagy is limited to a base
level, however, under stressful conditions, autophagy is enhanced
(8). Autophagy can be regarded as
a 'double-edged sword'; sustained and excessive activation of
autophagy may lead to autophagic cell death, while moderate
induction of autophagy promotes cell survival (9). Accumulating evidence has demonstrated
that autophagy affects the sensitivity of tumor cells to
radiotherapy, chemotherapy and immunotherapy in several types of
cancer, including prostate and colon cancer, glioma and melanoma
(10–14). It has been previously reported that
inhibiting autophagy enhances the sensitivity of tumor cells to
radiotherapy (15,16). In addition, the activation of
autophagy attenuates the cytotoxicity of anti-HER2 monoclonal
antibody in breast cancer cells (17). Furthermore, autophagy has been
found to significantly improve the tolerance of hepatoma cells to
chemotherapy (18).
Previous studies have demonstrated that proteasome
inhibitors not only induce apoptosis, but they also activate
autophagy in tumor cells (9). In
our previous study, inhibition of the proteasome induced apoptosis
and autophagy (9). These data
indicated that autophagy also contributed to the resistance of
cancer cells to proteasome inhibitors. However, the underlying
mechanism remains to be fully elucidated.
The endoplasmic reticulum (ER) synthesizes proteins
and stores calcium, in addition to detecting and responding to
alterations in the microenvironment (19). Conditions of stress result in ER
stress, which in turn triggers the unfolded protein response
(20). ER chaperones mediate ER
associated protein degradation (ERAD) (21). ERAD detects and removes misfolded
proteins in the ER through the UPP and autophagy-lysosomal
pathways. Studies have demonstrated that abnormal protein
aggregation within the ER can activate the protein kinase RNA-like
endoplasmic reticulum kinase pathway and induce eukaryotic
translation initiation factor 2α (eIF2α) phosphorylation, which
promotes autophagy (22). Previous
studies have shown that the treatment of cancer cells with
proteasome inhibitors leads to autophagy accompanied by ER stress
(23–26). Therefore, the present study
hypothesized that ER stress may contribute to the induction of
autophagy by proteasome inhibitors, and that the inhibition of ER
stress or autophagy may sensitize cancer cells to
proteasome-targeting drugs.
In the present study, MCF-7 human breast cancer
cells were treated with the proteasome inhibitor, MG-132, in the
presence or absence of the autophagy inhibitor, 3-methyladenine
(3-MA), or the ER stress inhibitor, salubrinal. Subsequently, the
effects of treatment on cell viability, apoptosis and the induction
of autophagy were determined. The aim of the current study was to
investigate the promoting role of autophagy inhibitor 3-MA and
endoplasmic reticulum stress inhibitor salubrinal on the MCF-7
cell-growth inhibition effect of proteasome inhibitor MG-132, and
to provide a theoretical basis for the future development of breast
cancer treatments.
Materials and methods
Reagents
Gibco Dulbecco's modified Eagle's medium (DMEM) was
purchased from Thermo Fisher Scientific, Inc. (Waltham, MA, USA);
fetal bovine serum (FBS) was purchased from TBD Science (Hangzhou,
China); Calbiochem MG-132 was obtained from EMD Millipore (San
Diego, CA, USA) and was dissolved in phosphate-buffered saline
(PBS) to a storage concentration of 20 μmol/l. 3-MA was
purchased from Sigma-Aldrich (St. Louis, MO, USA) and was dissolved
in PBS to a storage concentration of 100 mmol/l. Salubrinal was
purchased from EMD Millipore and was dissolved in PBS to a storage
concentration of 100 μmol/l. The polyclonal rabbit
anti-microtubule-associated protein 1 light chain 3 (LC3; cat. no.
4108) antibody, polyclonal rabbit anti-caspase-3 (cat. no. 9662)
and polyclonal rabbit anti-caspase-12 (cat. no. 2202) antibodies
were purchased from Cell Signaling Technology, Inc. (Danvers, MA,
USA), and the monoclonal mouse anti-B cell lymphoma-2 (BCL-2; cat.
no. ab692), monoclonal mouse anti-Bcl-2-associated X protein (BAX;
cat. no. ab5714), monoclonal mouse anti-glucose-regulated protein
78 (Grp-78; ab151269) and monoclonal mouse anti-GADD153 (cat. no.
ab11419) antibodies were purchased from Abcam (Cambridge, MA, USA).
The Cell Cycle and Apoptosis Analysis kits were obtained from
Beyotime Institute of Biotechnology (cat. nos. C1052 and C1063;
Shanghai, China).
Cell line and cell culture
Human MCF-7 breast cancer cells were obtained from
the Gene Treatment Center of the China-Japan Union Hospital of
Jilin University (Changchun, China). The MCF-7 cells were cultured
in DMEM supplemented with 10% FBS, and were maintained at 37°C and
5% CO2. Cells (80–90% confluence) in the mid-log phase
were used in the subsequent experiments.
MTT assay to determine cell
viability
The MCF-7 cells were seeded onto 96-well plates at a
concentration of 1×105 cells/well and cultured overnight
at 37°C. The cells were divided into three groups. The first group
was treated with 2.5 μmol/l MG-132; the second group was
co-treated with 1.0 mmol/l 3-MA and MG-132; and the third group was
co-treated with 10 μmol/l salubrinal and MG-132. All groups
were treated for 12, 24 and 48 h at 37°C. Subsequent to treatment,
each group of cells was incubated with 5 mg/ml MTT for 4 h,
followed by detection using an enzyme-labeling measuring instrument
(Model 680; Bio-Rad Laboratories, Inc., Hercules, CA, USA) at 570
nm. Each experiment was repeated three times in triplicate.
Detection of apoptosis and cell
cycle
Annexin V-fluorescein isothiocyanate (FITC) and
propidium iodide (PI) (Beyotime Institute of Biotechnology, Haimen,
China) were used to evaluate the rates of apoptosis of the cells.
Subsequent to washing twice with PBS and adjusting the cells to a
density of 5×105 cells/ml, one group of MCF-7 cells were
fixed in 70% ethanol at 4°C overnight, treated with 100 mg/l RNase
(Beyotime Institute of Biotechnology) at 37°C for 30 min and
stained with 50 mg/l PI at 4°C for 30 min. Another group was
stained with 150 mg/l FITC at 20–25°C for 10 min and with 50 mg/l
PI at 4°C for 30 min. Following starvation overnight at 37°C in
serum-free DMEM, the MCF-7 cells (1×105 cells/ml) were
either treated with 2.5 μmol/l MG-132 or were co-treated
with 1.0 mmol/l 3-MA or 10 μmol/l salubrinal for 48 h at
37°C. Following treatment, the attached and floating cells were
harvested with 0.25% trypsin (Amresco LLC, Solon, OH, USA), washed
twice with PBS and adjusted to a density of 5×105
cells/ml. Flow cytometric analysis was then performed using the
FACScan flow cytometer (BD Biosciences, San Jose, CA, USA),
according to the manufacturer's protocol of the Cell Cycle and
Apoptosis Analysis kit (Beyotime Institute of Biotechnology).
Western blotting
Protein was extracted from cells with lysis buffer
(Beijing Chemicals Company Limited, Beijing, China) containing 1%
Nonidet P-40, 150 mM NaCl, 50 mM Tris-HCl (pH 7.5), 1 mM NaF, 1 mM
phenylmethyl-sulfonyl fluoride, 4 mg/ml leupeptin and 1 mg/ml
aprotinin for 30 min. Proteins were quantified with Coomassie Blue
staining assay using the Bradford Protein Assay kit (Beyotime
Institute of Biotechnology). Proteins (50 μg; 5
μg/μl) were resolved on a 12% sodium dodecyl
sulfate-polyacrylamide gel (Beijing Chemicals Company Limited) and
transferred to polyvinylidene difluoride membranes (Beyotime
Institute of Biotechnology) followed by blocking with 5% fat free
dry milk in Tris-buffered saline with Tween 20 (TBST; Beijing
Chemicals Company Limited), containing 20 mM Tris-HCl (pH 7.6), 150
mM NaCl and 0.02% Tween 20. The membranes were then incubated with
the following primary antibodies in TBST containing 0.1% Tween 20
and 1% fat-free milk, at 4°C overnight: LC3 (1:2,000); BCL-2
(1:3,000); BAX (1:500); polyclonal rabbit caspase-3 (1:1,000; cat.
no. 9662; Cell Signaling Technology, Inc.); caspase-12 (1:1,000);
Grp-78 (1:2,000). The membranes were then washed four times in TBST
and incubated at 20–25°C for 30 min with horseradish
peroxidase-conjugated secondary antibody (1:2,000; Bio-Rad
Laboratories, Inc.). Signals were developed using an ECL-Plus kit
(Beyotime Institute of Biotechnology). Protein expression levels
were quantified using ImageJ software (version 1.48; National
Institutes of Health, Bethesda, MD, USA)
RNA extraction and reverse
transcription-quantitative polymerase chain reaction (RT-qPCR)
The MCF-7 cells were plated onto a 6-well plate at a
concentration of 2×105 cells/well. Following 24 h
incubation, the cells were treated with MG-132 (2.5 μmol/l)
and co-treated with salubrinal (10 and 20 μmol/l) for an
additional 24 h. Total RNA was extracted using Invitrogen TRIzol
reagent (Thermo Fisher Scientific, Inc.), according to the
manufacturer's protocol. Total RNA (1 μg) was reverse
transcribed using a Reverse Transcription system (Select Cycler II
Gradient PCR Instrument; Takara Biotechnology, Co., Ltd., Dalian,
China). The single-stranded cDNA was amplified by qPCR with the
BeyoRT First Strand cDNA synthesis kit (RNase H minus; D7233;
Beyotime Institute of Biotechnology) using primer pairs specific
for GRP-78, growth arrest and DNA damage induced gene-153
(GADD153), caspase-12 and GAPDH. The reaction mixture was in a
total volume of 50 μl and contained cDNA (0.5 μg),
buffer (5 μl), primers (2 μl; Genewiz Biological
Technology Co., Ltd., Beijing, China), dNTP mixture (8 μl),
Taq (0.5 μl) and ddH2O (from the BeyoRT First
Strand cDNA synthesis kit). Cycling was conducted using a PX2
Thermal Cycler (Thermo Fisher Scientific, Inc.) and the qPCR
conditions were as follows: 95°C for 5 min, followed by 95°C for 40
sec, 55°C for 40 sec and 72°C for 1 min for 25 cycles, and 72°C for
10 min. The primers (GENEWIZ Biological Technology Company Limited,
Jiangsu, China) used in the present study were as follows: Grp-78,
forward 5′-GCAGCAGGACATCAAGTTCT-3′ and reverse 5′-CGCTGGTCAAAGTCT
TCTCC-3′; caspase-12, forward 5′-AATCTGTGGGACCAAGCAAC-3′ and
reverse 5′-GAGCCTTTG TAACAGCATCA-3′; and GADD153, forward
5′-AATGCTTGCTCTGATAGGCG-3′ and reverse 5′-CTGGAATCT GGAGAGTGAGG-3′.
The qPCR products were analyzed using a 1% agarose gel (Beijing
Chemicals Company Limited, Beijing, China).
Statistical analysis
Data were analyzed using SPSS software, version 17.0
(SPSS, Inc., Chicago, IL, USA). Experiments were repeated a minimum
of three times. Statistical comparisons were made using Student's
t-test. P<0.05 was considered to indicate a statistically
significant difference. Error bars represent the mean ± standard
deviation unless otherwise stated.
Results
Inhibition of autophagy or ER stress
sensitizes MCF-7 cells to MG-132
To examine the effect of inhibiting autophagy or ER
stress following inhibition of the proteasome on cell viability,
the MCF-7 cells were treated with MG-132, MG-132 and 3-MA, or
MG-132 and salubrinal for 12, 24 and 48 h. Cell viability was then
determined using an MTT assay. Compared with the untreated control
cells, MG-132 marginally inhibited cell viability. Notably,
inhibition of either autophagy with 3-MA or ER stress by salubrinal
enhanced the loss of cell viability by MG-132 (Fig. 1A). Cell apoptosis was detected
using annexin V-FITC and PI staining. Compared with the untreated
control, MG-132 induced apoptosis (Fig. 1B and C). Co-treatment of MG-132
with either 3-MA (Fig. 1D) or
salubrinal (Fig. 1E) resulted in
enhanced cell apoptosis, compared with that in the cells treated
with MG-132 alone. In addition, the percentage of MCF-7 cells in
the G2 phase was increased to 22.40, 24.80 and 25.76% by
MG-132, MG-132+3-MA, and MG-132+salubrinal, respectively.
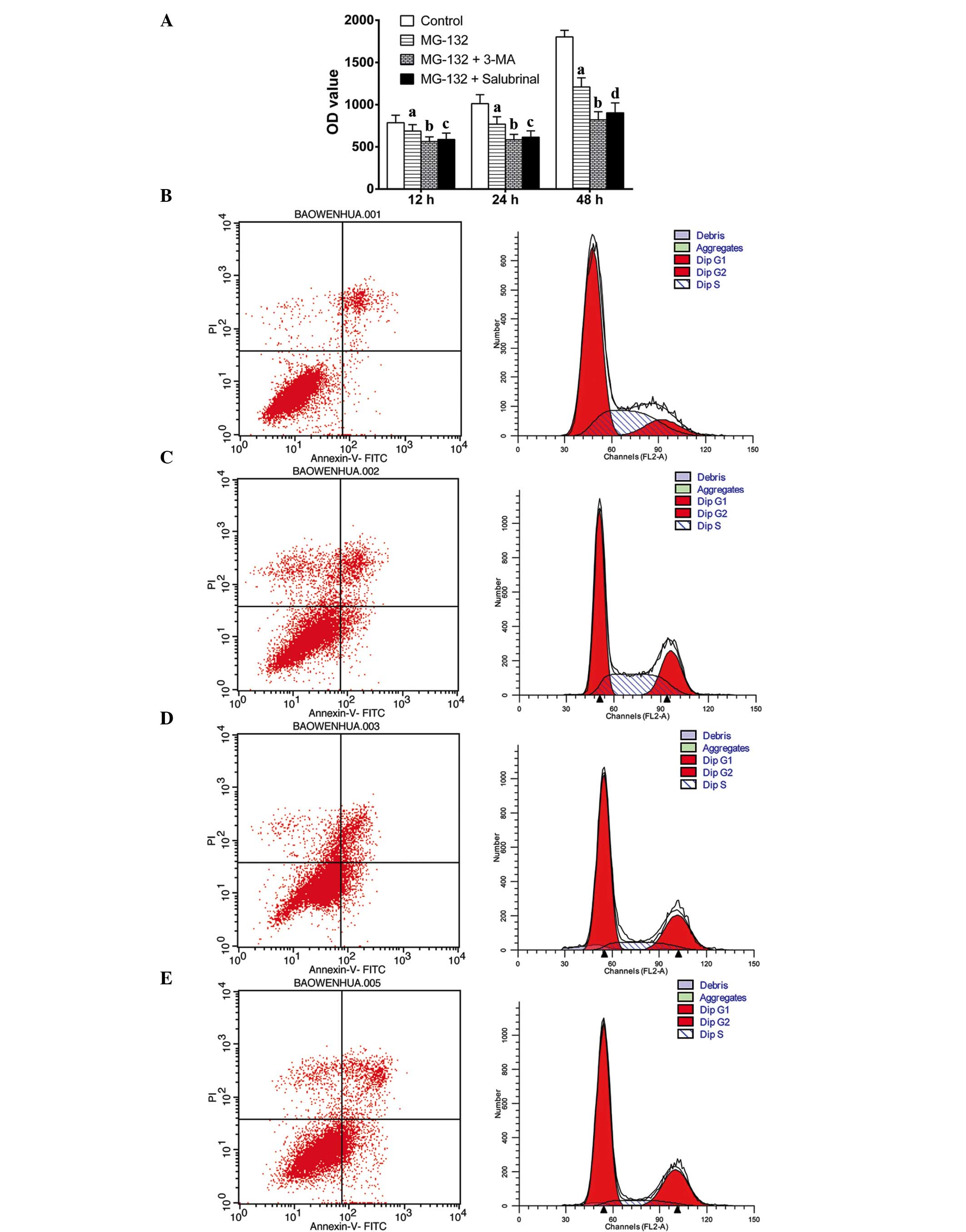 | Figure 13-MA and salubrinal enhance cell death
induced by MG-132 in MCF-7 cells. The MCF-7 cells were treated with
MG-132 (2.5 μmol/l), MG-132+3-MA (1 mmol/l) or
MG-132+salubrinal (10 μmol/l) for 12, 24 and 48 h. (A) Cell
proliferation was determined using a
3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide assay.
Apoptosis and the cell cycle were determined using flow cytometry
following annexin V-FITC and PI staining. (B) Control; (C) MG-132
(2.5 μmol/l); (D) MG-132 (2.5 μmol/l) + 3-MA (1
mmol/l); (E) MG-132 (2.5 μmol/l) + salubrinal (10
μmol/l). Error bars represent the mean ± standard deviation;
aP<0.05, vs. control group at each time point;
bP<0.05, vs. MG-132 group at each time point;
cP<0.05, vs. MG-132 group at 24 h and 48 h;
dP<0.01, vs. MG-132 group at 48 h. 3-MA,
3-methyladenine; FITC, fluorescein isothiocyanate; PI, propidium
iodide; OD, optical density. |
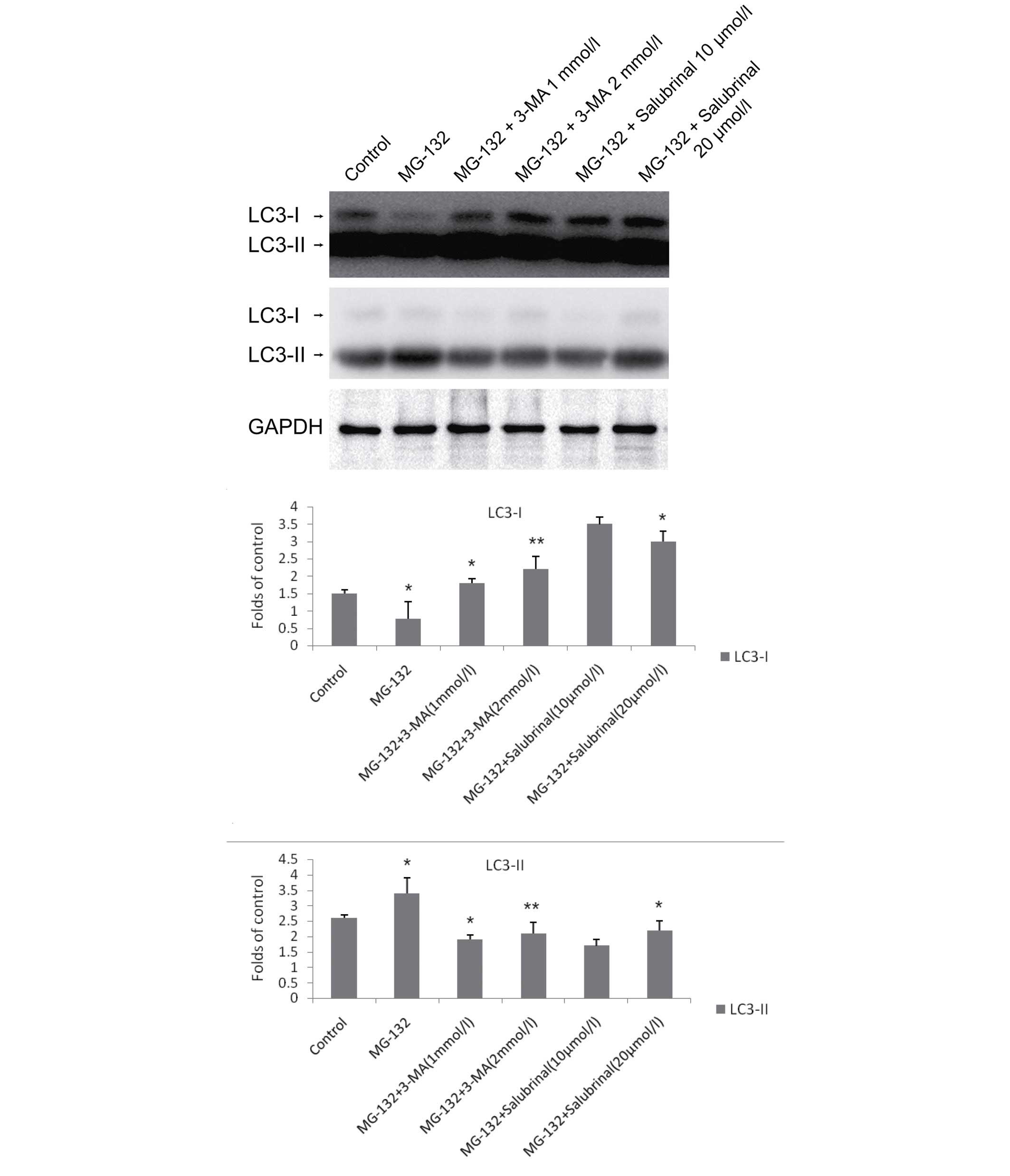
Inhibition of ER stress prevents the
induction of autophagy by MG-132
To determine the effects of MG-132, 3-MA, and
salubrinal on autophagy, the protein levels of LC3, a marker of
autophagy, were assessed using western blotting. LC3 has a
cytosolic form (LC3-I; 18-kDa) and a lipidated form (LC3-II; 16
kDa, and the conversion of LC3-I to LC3-II indicates the induction
of autophagy (27). As shown in
Fig. 2, the expression of LC3-I
was reduced, and the expression of LC3-II was increased in the
MG-132-treated cells. The expression of LC3-I was increased and the
conversion of LC3-I to LC3-II was reduced following the addition of
3-MA, which occurred in a dose-dependent manner. Similarly, in the
cells treated with MG-132 and salubrinal (10 and 20 μmol/l),
the expression of LC3-I was increased and the conversion of LC3-I
to LC3-II was reduced.
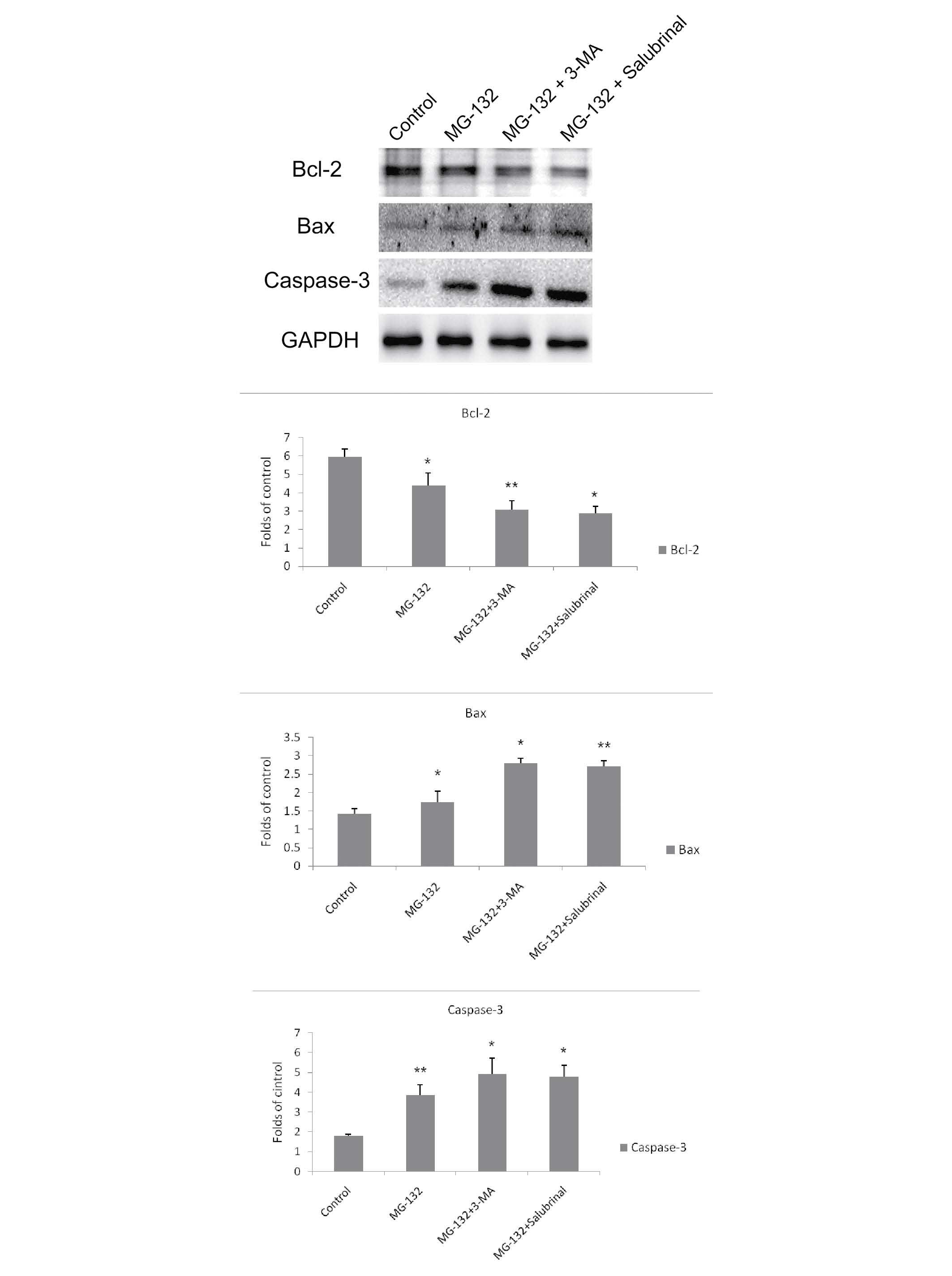
Inhibition of autophagy or ER stress
potentiates MG-132-induced apoptosis in MCF-7 cells
To investigate whether MG-132-induced apoptosis can
be upregulated by inhibiting autophagy or ER stress, the MCF-7
cells were treated with MG-132 in the presence or absence of 3-MA
or salubrinal for 48 h, following which the protein expression
levels of Bax, Bcl-2 and caspase-3 were determined. As shown in
Fig. 3, the protein expression of
anti-apoptotic Bcl-2 was reduced in the MG-132-treated cells, and
was downregulated further following co-treatment with 3-MA or
salubrinal. The expression levels of the apoptotic proteins, Bax
and caspase-3, were increased in the MG-132-treated cells, compared
with the control cells, and the levels were further upregulated
following co-treatment with 3-MA or salubrinal.
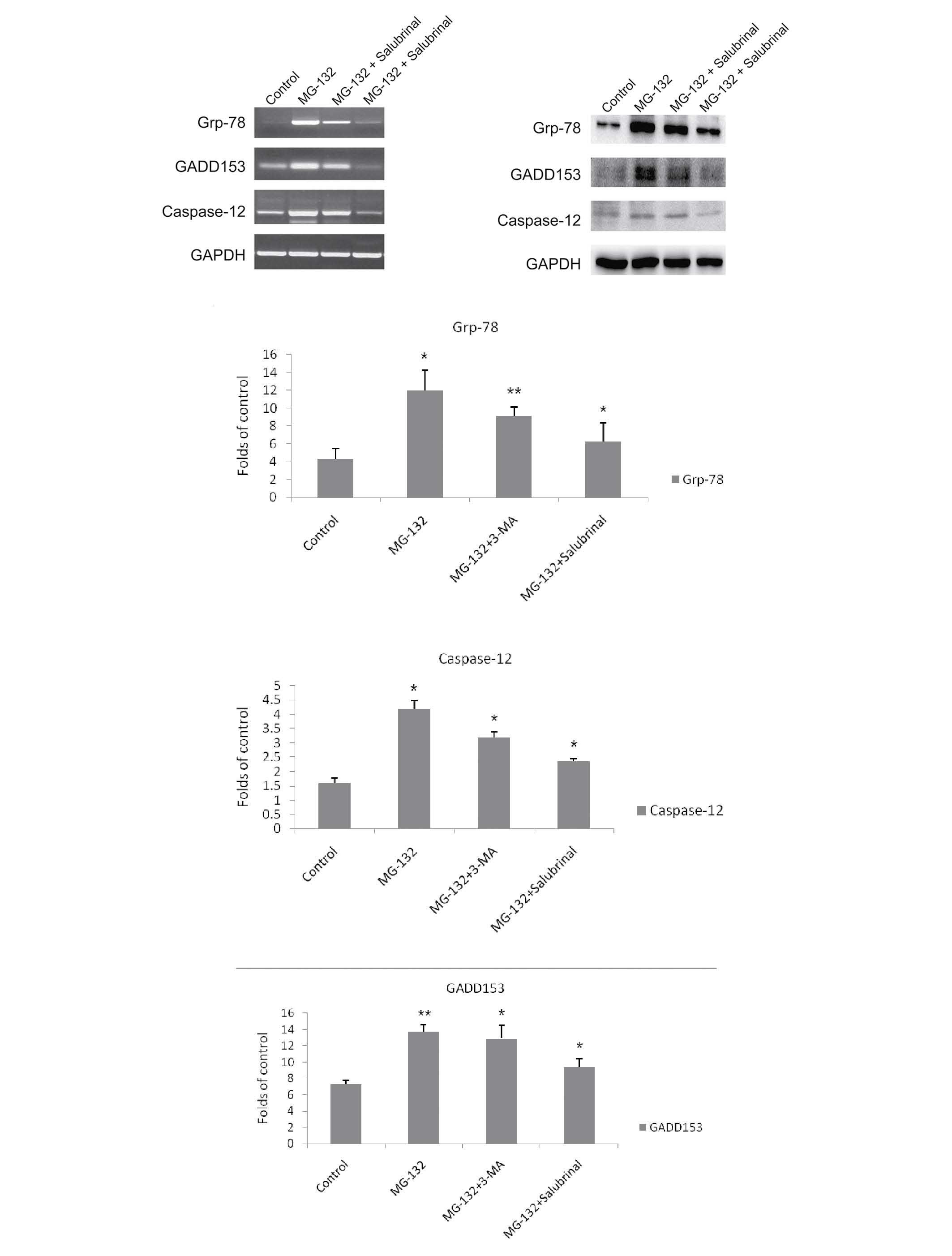
Salubrinal inhibits ER stress induced by
MG-132
Grp-78 and GADD153 proteins are associated with ER
stress, and caspase-12 is associated with ER stress-induced
apop-tosis (19,28,29).
To determine the effect of ER stress on MCF-7 cells following
treatment with MG-132, the MCF-7 cells were treated with MG-132 in
the presence or absence of salubrinal for 48 h, and the mRNA and
protein expression levels of Grp-78, GADD153 and caspase-12 were
determined using RT-qPCR and western blotting, respectively. MG-132
markedly increased the mRNA levels of Grp-78, GADD153 and
caspase-12. However, salubrinal inhibited the mRNA levels of
Grp-78, GADD153 and caspase-12, in a dose-dependent manner
(Fig. 4A). As expected, MG-132
increased the protein expression levels of Grp-78, GADD153 and
caspase-12, the effect of which was also eliminated by salubrinal
in a dose-dependent manner (Fig.
4B).
Discussion
In the present study, it was demonstrated that
inhibition of the proteasome by MG132 resulted in ER stress, and
inhibiting autophagy and ER stress following MG-132 treatment
inhibited cell viability. The inhibitory effect of 3-MA was
observed to be more effective than that of salubrinal. In addition,
the inhibition of ER stress prevented the induction of autophagy by
MG-132, suggesting that MG-132-induced autophagy may be associated
with ER stress. Notably, the inhibition of autophagy or ER stress
potentiated MG-132-induced apoptosis in the MCF-7 cells. The
present study demonstrated that ER stress may contribute to the
induction of autophagy by proteasome inhibition, and suggested that
inhibitors of ER stress and autophagy may be promising in the
development of novel combinatorial targeted cancer therapies using
proteasome inhibitors.
Autophagy can induce cancer cell death through an
apoptosis-dependent or an apoptosis-independent cascade (30), and can promote tumor cell survival
by degrading misfolded proteins and damaged organelles (31,32).
Previous studies have suggested that the inhibition of autophagy
may lead to apoptosis of tumor cells, which is associated with the
release of cytochrome c, the activation of caspase-3 and
poly ADP ribose polymerase, the phosphorylation of p53 and
downregulation of the anti-apoptotic Bcl-2 protein (33–35).
It was previously reported that MG-132-induced autophagy has a
protective role in breast cancer cells, whereas the autophagy
inhibitor, 3-MA, can enhance the cell death-inducing effect of
MG-132 in MCF-7 cells (9). In
agreement, the present study found that the inhibition of autophagy
sensitized the MCF-7 cells to MG-132 treatment. However, the
mechanism by which inhibition of the proteasome leads to the
autophagy of tumor cells remains to be fully elucidated.
ER stress is a highly conserved signal, which has
been investigated in several different types of cell and tissue
(36). Mild ER stress results in
an adaptive response by autophagy to avoid cell death, however,
excessive ER stress can lead to cell death (37). ER stress resulting from abnormal
protein aggregation in the ER can activate the PERK pathway, which
promotes the phosphorylation of eIF2α to induce the conversion of
LC3-I to LC3-II (38). The results
of the present study indicated that ER stress inhibited
MG-132-induced apoptosis in the MCF-7 cells. This supports the
finding of a previous observation that the accumulation of
misfolded proteins, resulting from inhibition of the UPP, trigger
ER stress, which in turn induces autophagy (39). Despite accumulating evidence
demonstrating that inhibition of the proteasome activates ER
stress, others have reoirted that excessive stress in the ER
increases ATF4 translation by the PERK pathway. This results in a
mitogen-activated protein kinase kinase kinase activation cascade,
via the inositol-requiring enzyme 1 pathway, leading to apoptosis
(19). The results of the present
study indicated that salubrinal increased apoptosis, decreased the
expression of ER stress-associated protein, enhanced MG-132-induced
cell death and increased the number of MCF-7 cells in the
G2/M phase. These results indicated that salubrinal
inhibited the activation of autophagy by inhibiting ER stress, thus
indirectly enhancing the cell death-inducing effect of the MG-132
proteasome inhibitor.
In conclusion, the results of the present study
suggested that the proteasome inhibitor, MG-132, induced autophagy
via the ER stress pathway, which ultimately resulted in apoptosis
of the MCF-7 cells. In addition, the inhibition of autophagy and ER
stress enhanced cell death and inhibited cell proliferation. These
findings may provide a theoretical basis for the application of
inhibitors of ER stress and autophagy in combination with
proteasome inhibitors for the development of novel combinatorial
treatment strategies in targeted cancer therapy.
Acknowledgments
The present study was funded by grants from the
National Natural Science Foundation of China (grant. no.
20130101150JC).
References
|
1
|
Rubinsztein DC: The roles of intracellular
protein-degradation pathways in neurodegeneration. Nature.
443:780–786. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Hershko A: The ubiquitin system for
protein degradation and some of its roles in the control of the
cell division cycle. Cell Death Differ. 12:1191–1197. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Momose I, Iijima M, Kawada M and Ikeda D:
A new proteasome inhibitor, TP-110, induces apoptosis in human
prostate cancer PC-3 cells. Biosci Biotechnol Biochem.
71:1036–1043. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Yang H, Landis-Piwowar KR, Chen D, Milacic
V and Dou QP: Natural compounds with proteasome inhibitory activity
for cancer prevention and treatment. Curr Protein Pept Sci.
9:227–239. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Fan WH, Hou Y, Meng FK, Wang XF, Luo YN
and Ge PF: Proteasome inhibitor MG-132 induces C6 glioma cell
apoptosis via oxidative stress. Acta Pharmacol Sin. 32:619–625.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Testa U: Proteasome inhibitors in cancer
therapy. Curr Drug Targets. 10:968–981. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Levine B and Kroemer G: Autophagy in the
pathogenesis of disease. Cell. 132:27–42. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Shintani T and Klionsky DJ: Autophagy in
health and disease: A double-edged sword. Science. 306:990–995.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Ge PF, Zhang JZ, Wang XF, Meng FK, Li WC,
Luan YX, Ling F and Luo YN: Inhibition of autophagy induced by
proteasome inhibition increases cell death in human SHG-44 glioma
cells. Acta Pharmacol Sin. 30:1046–1052. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Zhu K, Dunner K Jr and McConkey DJ:
Proteasome inhibitors activate autophagy as a cytoprotective
response in human prostate cancer cells. Oncogene. 29:451–462.
2010. View Article : Google Scholar :
|
|
11
|
Nishikawa T, Tsuno NH, Okaji Y, Shuno Y,
Sasaki K, Hongo K, Sunami E, Kitayama J, Takahashi K and Nagawa H:
Inhibition of autophagy potentiates sulforaphane-induced apoptosis
in human colon cancer cells. Ann Surg Oncol. 17:592–602. 2010.
View Article : Google Scholar
|
|
12
|
Davids LM, Kleemann B, Cooper S and Kidson
SH: Melanomas display increased cytoprotection to
hypericin-mediated cyto-toxicity through the induction of
autophagy. Cell Biol Int. 33:1065–1072. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Katayama M, Kawaguchi T, Berger MS and
Pieper RO: DNA damaging agent-induced autophagy produces a
cytoprotective adenosine triphosphate surge in malignant glioma
cells. Cell Death Differ. 14:548–558. 2007. View Article : Google Scholar
|
|
14
|
Li J, Qin Z and Liang Z: The prosurvival
role of autophagy in Resveratrol-induced cytotoxicity in human U251
glioma cells. BMC Cancer. 9:2152009. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Apel A, Herr I, Schwarz HP, Rodemann HP
and Mayer A: Blocked autophagy sensitizes resistant carcinoma cells
to radiation therapy. Cancer Res. 68:1485–1494. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Kim KW, Hwang M, Moretti L, Jaboin JJ, Cha
YI and Lu B: Autophagy upregulation by inhibitors of caspase-3 and
mTOR enhances radiotherapy in a mouse model of lung cancer.
Autophagy. 4:659–668. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Vazquez-Martin A, Oliveras-Ferraros C and
Menendez JA: Autophagy facilitates the development of breast cancer
resistance to the anti-HER2 monoclonal antibody trastuzumab. PloS
One. 4:e62512009. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Song J, Qu Z, Guo X, Zhao Q, Zhao X, Gao
L, Sun K, Shen F, Wu M and Wei L: Hypoxia-induced autophagy
contributes to the chemoresistance of hepatocellular carcinoma
cells. Autophagy. 5:1131–1144. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Moretti L, Cha YI, Niermann KJ and Lu B:
Switch between apoptosis and autophagy: Radiation-induced
endoplasmic reticulum stress? Cell Cycle. 6:793–798. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
DeGracia DJ and Montie HL: Cerebral
ischemia and the unfolded protein response. J Neurochem. 91:1–8.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Kapoor A and Sanyal AJ: Endoplasmic
reticulum stress and the unfolded protein response. Clin Liver Dis.
13:581–590. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Fujita E, Kouroku Y, Isoai A, Kumagai H,
Misutani A, Matsuda C, Hayashi YK and Momoi T: Two endoplasmic
reticulum-associated degradation (ERAD) systems for the novel
variant of the mutant dysferlin: Ubiquitin/proteasome ERAD(I) and
autophagy/lysosome ERAD(II). Hum Mol Genet. 16:618–629. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Yorimitsu T, Nair U, Yang Z and Klionsky
DJ: Endoplasmic reticulum stress triggers autophagy. J Biol Chem.
281:30299–30304. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Gavilán MP, Pintado C, Gavilán E, Jiménez
S, Ríos RM, Vitorica J, Castaño A and Ruano D: Dysfunction of the
unfolded protein response increases neurodegeneration in aged rat
hippo-campus following proteasome inhibition. Aging Cell.
8:654–665. 2009. View Article : Google Scholar
|
|
25
|
Younce CW and Kolattukudy PE: MCP-1 causes
cardiomyoblast death via autophagy resulting from ER stress caused
by oxidative stress generated by inducing a novel zinc-finger
protein, MCPIP. Biochem J. 426:43–53. 2010. View Article : Google Scholar
|
|
26
|
Zhou F, van Laar T, Huang H and Zhang L:
APP and APLP1 are degraded through autophagy in response to
proteasome inhibition in neuronal cells. Protein Cell. 2:377–383.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Tanida I, Ueno T and Kominami E: LC3
conjugation system in mammalian autophagy. Int J Biochem Cell Biol.
36:2503–2318. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Heath-Engel HM, Chang NC and Shore GC: The
endoplasmic reticulum in apoptosis and autophagy: Role of the BCL-2
protein family. Oncogene. 27:6419–6433. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Oyadomari S and Mori M: Roles of
CHOP/GADDI53 in endoplasmic reticulum stress. Cell Death Differ.
11:381–389. 2004. View Article : Google Scholar
|
|
30
|
Yu L, Alva A, Su H, Dutt P, Freundt E,
Welsh S, Baehrecke EH and Lenardo MJ: Regulation of an ATG7-beclin
1 program of autophagic cell death by caspase-8. Science.
304:1500–1502. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Cuervo AM, Stefanis L, Fredenburg R,
Lansbury PT and Sulzer D: Impaired degradation of mutant
alpha-synuclein by chaperone-mediated autophagy. Science.
305:1292–1295. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Rubinsztein DC, DiFiglia M, Heintz N, Dutt
P, Freundt E, Welsh S, Baehrecke EH and Lenardo MJ: Autophagy and
its possible roles in nervous system diseases, damage and repair.
Autophagy. 1:11–22. 2005. View Article : Google Scholar
|
|
33
|
Li J, Hou N, Faried A, Tsutsumi S,
Takeuchi T and Kuwano H: Inhibition of autophagy by 3-MA enhances
the effect of 5-FU-induced apoptosis in colon cancer cells. Ann
Surg Oncol. 16:761–771. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Huang S and Sinicrope FA:
Celecoxib-induced apoptosis is enhanced by ABT-737 and by
inhibition of autophagy in human colorectal cancer cells.
Autophagy. 6:256–269. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Walls KC, Ghosh AP, Franklin AV, Klocke
BJ, Ballestas M, Shacka JJ, Zhang J and Roth KA: Lysosome
dysfunction triggers Atg7-dependent neural apoptosis. J Biol Chem.
285:10497–10507. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
McMillan DR, Gething MJ and Sambrook J:
The cellular response to unfolded proteins: Intercompartmental
signaling. Curr Opin Biotechnol. 5:540–545. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Gao B, Zhang XY, Han R, Zhang TT, Chen C,
Qin ZH and Sheng R: The endoplasmic reticulum stress inhibitor
salubrinal inhibits the activation of autophagy and neuroprotection
induced by brain ischemic preconditioning. Acta Pharmacol Sin.
34:657–666. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Hoyer-Hansen M and Jäättelä M: Connecting
endoplasmic reticulum stress to autophagy by unfolded protein
response and calcium. Cell Death Differ. 14:1576–1582. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Ding WX, Ni HM, Gao W, Yoshimori T, Stolz
DB, Ron D and Yin XM: Linking of autophagy to ubiquitin-proteasome
system is important for the regulation of endoplasmic reticulum
stress and cell viability. Am J Pathol. 171:513–524. 2007.
View Article : Google Scholar : PubMed/NCBI
|