Introduction
Medicinal herbs are a tremendous source of natural
products with anti-cancer activity. Administration of natural
products has become an integral part in the prevention and
treatment of cancer. These phytochemicals provide potential novel
leads for the development of anti-angiogenic drugs (1,2).
Quercetin (3,3′,4′,5,7-pentahydroxy-flavone) is a
polyphe-nolic substance, which has been shown to have beneficial
biological properties due to its anti-oxidative activity as well as
the capacity to modify eicosanoid biosynthesis, prevent
atherosclerotic plaque formation and platelet aggregation, and to
have relaxant effects on cardiovascular smooth muscles (3–5). Of
note, quercetin has been demonstrated to induce apoptosis in a
variety of tumor cell types, while preventing the apoptosis of
certain non-tumorous cell types (6,7).
Furthermore, quercetin inhibits angiogenesis of tumors as well as
the expression of enzymes that activate carcinogens (2,8). In
particular, quercetin has been indicated to inhibit the growth and
to induce apoptosis of human leukemia cells by inhibiting the Wnt
protein β-catenin (9,10). Leukemia is among the most common
types of cancer worldwide; furthermore, its prevalence has been
increasing due to the extended human life, radiation (11,12)
and pollution (13,14), as a result of the decrease in the
incidence of infectious diseases in developed countries. The
incidence of leukemia is higher in developed countries than in
developing countries (15). In
developing countries, the use of natural plant extracts as remedies
against leukemia is common (15,16).
Heat shock proteins (HSPs) are a class of
functionally associated proteins with regulatory roles comprising
protein synthesis and degradation, prevention of stress-associated
injury, apoptosis and the generation of immune responses (17,18).
Five types of HSP have been identified in mammalian cells, among
which HSP27 has the important biological function of protecting
cells from damage arising from various stress factors with
phosphorylated HSP27 being considered as a potential diagnostic
marker for cancer (19). In
leukemia cells, HSP27 is closely associated with proliferation,
differentiation and heat resistance (20). Overexpression of HSP27 increases
the tumorigenicity and protects malignant cells against apoptotic
cell death through several mechanisms (21–24).
Previous studies have reported that quercetin
down-regulates the mRNA and protein expression of HSP27 and
increases the sensitivity of tumors to hyperthermia (25–27);
however, to the best of our knowledge, the anti-cancer effects of
quercetin together with HSP27 interference have not been studied in
leukemia cells. The present study aimed to investigate the effects
of small hairpin (sh)RNA with specificity against HSP27 (shHSP27)
on the anti-tumor effects of quercetin, including the inhibition of
cell proliferation and adhesion, induction of apoptosis and cell
cycle arrest, in U937 human leukemia cells.
The mammalian target of rapamycin (mTOR) is a
central regulator of cell growth, proliferation, differentiation
and survival. Studies have shown that mTOR is frequently
hyper-activated in cancer and is a crucial regulator of cancer cell
motility, invasion and metastasis (28,29).
Quercetin has been found to inhibit mTOR signaling during cancer
treatment and prevention (30,31).
Therefore, the present study also evaluated the regulatory activity
of quercetin plus shHSP27 on mTOR signaling in human leukemia U937
cells.
Materials and methods
Cell culture and reagents
The U937 human leukemia cell line was purchased from
the American Type Culture Collection (Manassas, VA, USA) and
cultured in RPMI-1640 medium (Hyclone Corp., Logan, UT, USA),
supplemented with 10% fetal bovine serum (Gibco; Thermo Fisher
Scientific, Inc., Waltham, MA, USA). Quercetin was purchased from
Sigma-Aldrich (St. Louis, MO, USA) and dissolved in dimethyl
sulfoxide to a concentration of 50 µM and stored at −20°C
for use. A Cell Counting Kit 8 (CCK-8) was obtained from JRDUN
Biotechnology Co. Ltd (Shanghai, China). An Annexin V-fluorescein
isothiocyanate (FITC) apoptosis detection kit and Matrigel were
purchased from BD Biosciences (Franklin Lakes, NJ, USA).
Lentiviral vector construction
For knockdown of HSP27 expression, the
pCMV-G-NR-U6-shRNA vector (Genechem, Shanghai, China) was used.
Three sequences of the human HSP27 gene HSPB1 (GenBank
accession no. NM001540) were selected as targets for RNA
interference: shHSP27-1 (start, 585 bp); 5′-GCTGCAAAATCCGATGAGA-3′;
shHSP27-2 (start, 293 bp), 5′-CCTGGATGTCAACCACTTC-3′; and shHSP27-3
(start, 322 bp), 5′-AGCTGACGGTCAAGACCAA-3′. In all subsequent
experiments, shHSP27-3 was used if not indicated otherwise.
For construction of the lentiviruses, 293T cells
(Enzyme-Linked Biological Technology Co., Ltd., Shanghai, China)
were transfected using Lipofectamine 2000 (Invitrogen; Thermo
Fisher Scientific) with plasmids expressing retroviral proteins
Gag-Pol and VSV-G (Addgene, Cambridge, MA, USA). At 48 h after
transfection, supernatants containing the retrovirus were collected
and frozen at −70°C until use.
RNA quantification by
reverse-transcription quantitative polymerase chain reaction
(RT-qPCR)
The expression of HSP27 in U937 cells was quantified
by real-time PCR. Total RNA was extracted using the TRIzol reagent
(Invitrogen) and 1 µg total RNA was reverse-transcribed into
cDNA using M-MLV reverse transcriptase in the presence of oligo
(dT)12–18. RT-PCR was performed in triplicate with SYBR Green
master mix (Toyobo, Osaka, Japan) for 10 min at 95°C for initial
denaturation, followed by 40 cycles of 95°C for 15 sec, 58°C for 30
sec and 72°C for 30 sec in the StepOne™ Real-Time PCR System
(Genuine Biosystem, Chennai, India). The following primers (Generay
Biotech Co., Ltd., Shanghai, China) were used: HSP27 forward,
5′-CCAGAGCAGAGTCAGCCAGCAT-3′ and reverse,
5′-CGAAGGTGACTGGGATGGTGA-3′; GAPDH forward,
5′-ACCACAGTCCATGCCATCAC-3′ and reverse, 5′-TCCACCACCCTGTTGCTGTA-3′.
GAPDH was used as a reference gene.
Cell proliferation assay
U937 cells were seeded in triplicate into 96-well
plates at a density of 2×104 cells/well. Cells were
treated with medium (control group), empty vector (50 µM/l),
shHSP27 vector (50 µM/l), quercetin (50 µM/l), empty
vector plus quercetin or shHSP27 vector plus quercetin. Following
incubation at 37°C for 0, 24 or 48 h, 10 µl CCK-8 solution
was added to each well and subsequent to further incubation for 3
h, the absorbance was detected at a wavelength of 450 nm. The cell
proliferation rate (relative to control) of the five treated groups
was calculated as follows: Proliferation rate (relative to control)
= (absorbance of experimental group/absorbance of control group)
×100%. All experiments were performed in triplicate.
Flow cytometry
U937 cells at 2×106 per well were seeded
into a six-well plate and treated with medium, empty vector,
shHSP27 vector, quercetin, empty vector + quercetin or shHSP27
vector + quercetin and then cultured for 48 h. Cells were then
collected, washed twice with phosphate-buffered saline (PBS) and
prepared for analysis of apoptosis and the cell cycle. For the
apoptosis assay, cell suspensions were adjusted to a density of
1×106/ml with binding buffer (Thermo Fisher Scientific,
Inc.). Annexin V-FITC and propidium iodide (PI) were added to the
cell suspension according to the manufacturer's instructions and
apoptosis was detected using a BD C6 flow cytometer (BD
Biosciences). For cell cycle analysis, the cells were fixed in 2 ml
pre-cooled 70% ethanol at 4°C overnight. The cells were washed
twice, stained with 400 µl PI (50 µg/ml) and 50
µg/ml RNase, and then subjected to flow cytometric
analysis.
Cell adhesion assay
An adhesive artificial basement membrane was
prepared by adding a mixture of serum-free RPMI-1640 medium and
Matrigel at a density of 100 µg/2,500 µl into a
12-well plate at 250 µl/well and drying overnight. U937
cells were seeded into the 12-well plate at a density of
2×104 cells/well and treated with the respective vectors
and/or quercetin as described above. Following 48 h of incubation
at 37°C, the culture medium was discarded and non-adherent cells
were removed by rinsing with PBS. Adherent cells were then fixed
with methanol for 15 min at 37°C and then stained with crystal
violet (JRDUN Biotechnology Co., Ltd., Shanghai, China) for 20 min.
The numbers of adherent cells were counted under a microscope
(BX51; Olympus, Tokyo, Japan).
Western blot analysis
Cells were lysed using radioimmunoprecipitation
assay lysis buffer (Keygentec, Nanjing, China) to extract total
proteins. The protein concentration was determined using a
bicinchoninic acid assay (Keygentec). Protein samples (30
µg) were separated by 10% SDS-PAGE and transferred onto
polyvinylidene fluoride membranes (Millipore, Billerica, MA, USA).
Membranes were blocked with 5% non-fat milk at room temperature for
1 h and incubated overnight at 4°C with the following antibodies:
B-cell lymphoma 2 (Bcl-2; cat. no. sc-492; 1:100) and
Bcl-2-associated X protein (Bax; cat. no. sc 492; 1:150), from
Santa Cruz Biotechnology, Inc. (Santa Cruz, CA, USA); cyclin B1
(cat. no. 12231; 1:1,000), cyclin D1 (cat. no. 2978; 1:1,000), AKT
(cat. no. 9272S; 1:1,000), phosphorylated (p)-AKT (cat. no. 4058S;
1:1,000), mammalian target of rapamycin (mTOR; cat. no. 2983;
1:1,000), p-mTOR (cat. no. 5536; 1:800) and hypoxia-inducible
factor (HIF)1α (cat. no. 3176; 1:1,000) from Cell Signaling
Technology, Inc. (Danvers, MA, USA); vascular endothelial growth
factor (VEGF; cat. no. ab46154; 1:1,000) and Notch1 (cat. no.
ab52627; 1:1,000), from Abcam (Cambridge, UK); and GAPDH (cat. no.
5471; 1:1,500; Cell Signaling Technology, Inc.). Membranes were
washed three times with Tris-HCl (pH 7.6; 20 mM) containing 137 mM
NaCl and 0.01% Tween-20 and then incubated with horseradish
peroxidase-conjugated goat anti-rabbit/mouse secondary antibody
(cat. no. A0208/A0216; 1:1,000; Beyotime Institute of
Biotechnology, Inc., Shanghai, China) at 25°C for 2 h. Membranes
were visualized using enhanced chemiluminescence solution
(Millipore) and x-ray film. Protein levels were determined by
densitometric analysis with normalization to GAPDH.
Statistical analysis
Values are expressed as the mean ± standard
deviation. All statistical analyses were performed using GraphPad
Prism 5.00 software (GraphPad Software, La Jolla, CA, USA).
Differences among groups were tested by one-way analysis of
variance followed by the Neuman-Keuls post-hoc test. A two-sided
P-value <0.05 was considered to indicate a statistically
significant difference.
Results
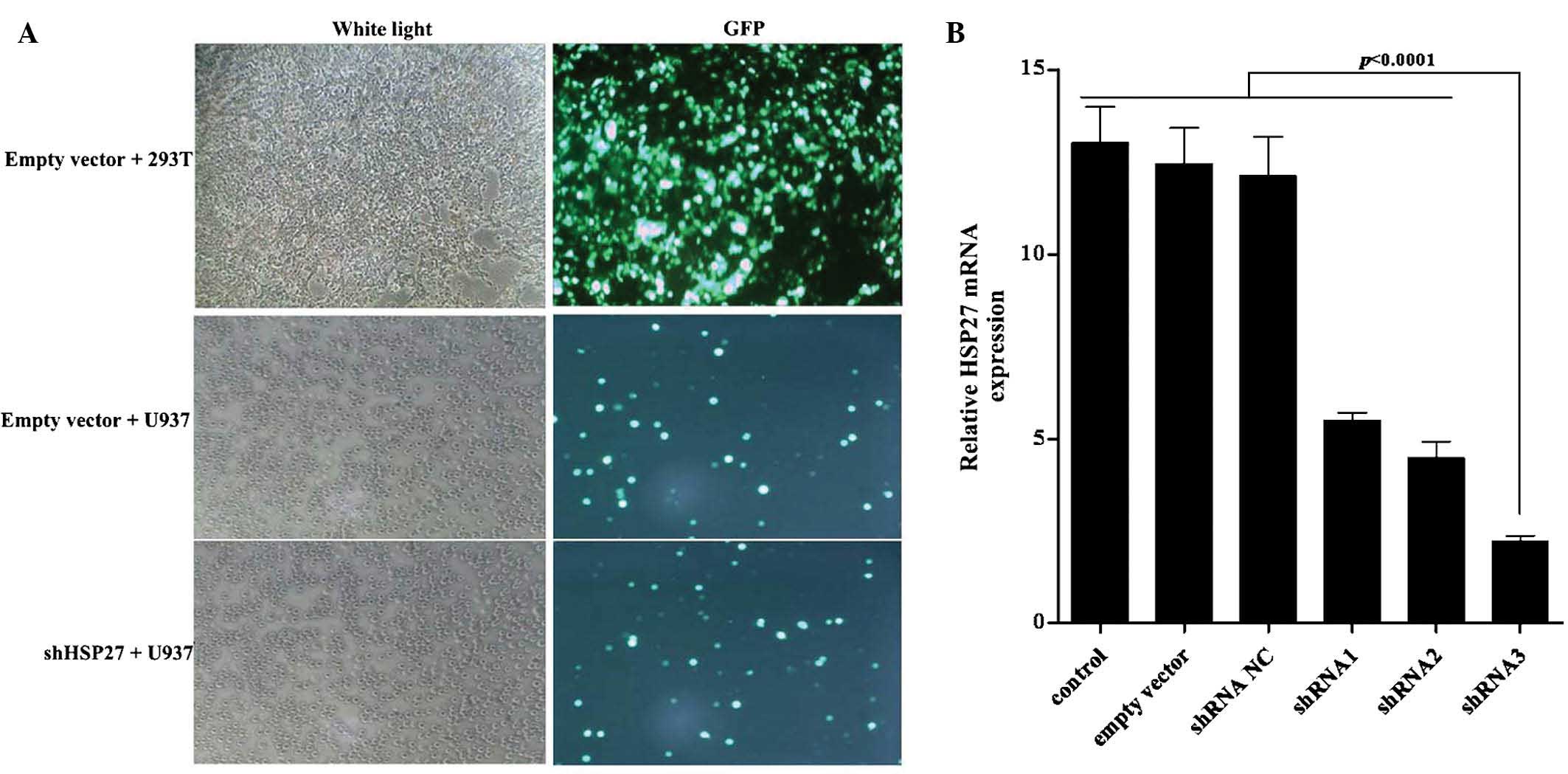
shHSP27 decreases HSP27 expression in
U937 cells
Numerous types of cancer cell constitutively express
HSP27 at elevated levels, which is associated with drug resistance
(32). To inhibit HSP27 expression
in U937 human leukemia cells, three pairs of shRNA were designed to
specifically target HSP27 and then ligated into the
pCMV-G-NR-U6-shRNA vector, which contains a green fluorescent
protein (GFP) expression gene for identification. Lentiviruses for
the expression of shHSP27 were harvested from the supernatant of
293T packaging cells. U937 cells were transfected with this
supernatant containing shHSP27 lentivirus and mRNA expression of
HSP27 was detected following 48 h of incubation. As shown in
Fig. 1A, a proportion of the U937
cells were successfully transfected with shHSP27-3 lentivirus and
HSP27 mRNA expression was considerably decreased by shHSP27-3
(Fig. 1B).
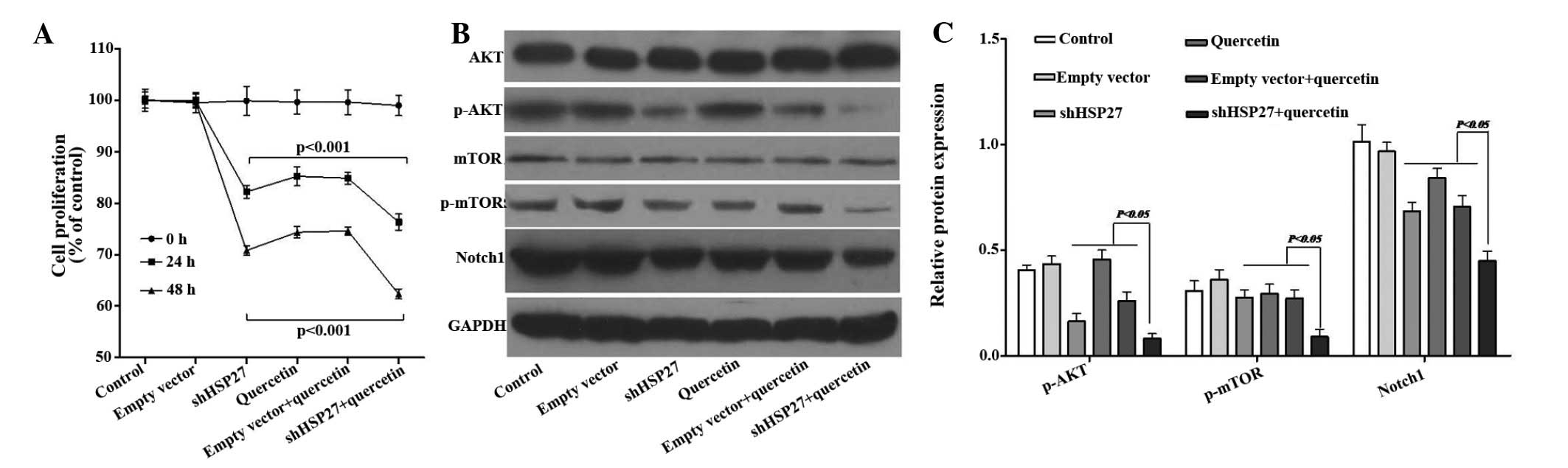
shHSP27 and quercetin jointly inhibit
U937-cell proliferation
Quercetin has been reported to inhibit the
proliferation of human leukemia cell lines, including U937
(33,34). At a concentration of 2 µM,
quercetin inhibited the proliferation of U937 cells by 15%
following 24 h of incubation, which was consistent the findings of
a previous study (34). When cells
were simultaneously treated with shHSP27, cell proliferation
further decreased by ~10% (Fig.
2A).
mTOR signaling has been reported to stimulate
cellular proliferation and aggressive tumor growth in numerous
cancer models (35). To
investigate the potential role of mTOR in the mechanism of action
of quercetin and shHSP27, the expression of mTOR signaling proteins
in U937 cells was assessed following treatment with quercetin
and/or shHSP27. Western blot analysis showed that following
treatment with quercetin, shHSP27 or their combination, the protein
levels of p-AKT and p-mTOR were markedly decreased, while levels of
total AKT were not obviously affected and those of total mTOR were
only marginally reduced; furthermore, the protein expression of
Notch1 was significantly decreased compared with that in the
control or empty vector-treated groups (Fig. 2B and C).
HSP27 knockdown enhances
quercetin-induced apoptosis of U937 cells
Following incubation of U937 cells with quercetin or
shHSP27 plus quercetin for 48 h, the percentage of apoptotic cells
significantly increased from 25.4 to 47.2% (Fig. 3A). To investigate the underlying
mechanism of this phenomenon, the protein expression of Bcl-2 and
Bax, two genes that regulate cell apoptosis, was assessed. Bcl-2 is
known inhibit apoptosis and to increase resistance to various
apoptosis-stimulating factors, while not being associated with cell
division and proliferation (36).
Bax, a homologous gene of the Bcl-2 family, antagonizes the
anti-apoptotic effect of the Bcl-2 by forming a heterodimer with
Bcl-2. It has been reported that the Bcl-2/Bax ratio is negatively
correlated with the apoptotic rate (37). While treatment with quercetin or
shHSP7 alone significantly reduced the Bcl-2/Bax ratio, their
combination synergistically decreased the Bcl-2/Bax ratio by ~90%
and almost totally blocked Bcl-2 expression in U937 cells (Fig. 3B).
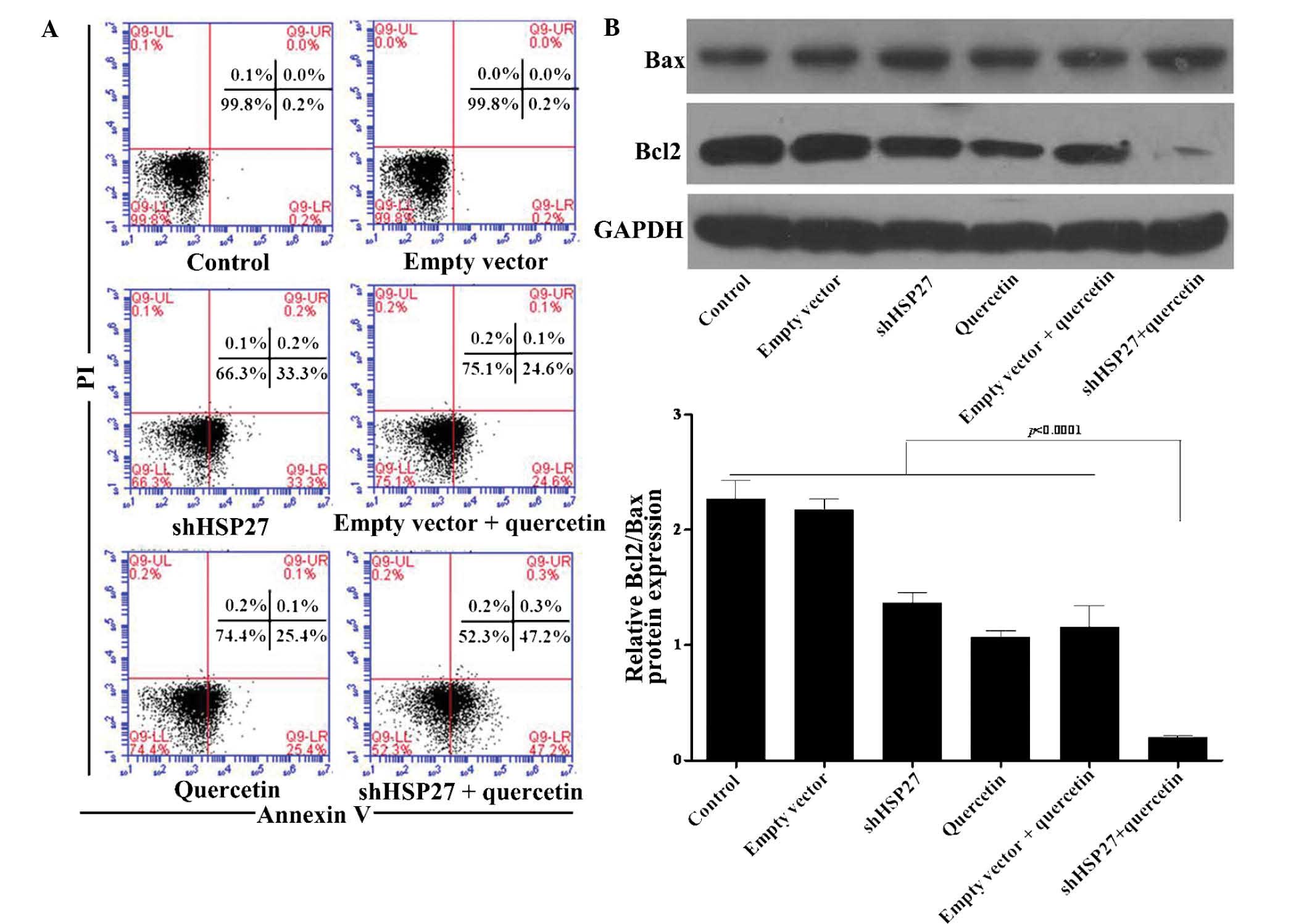 | Figure 3Effect of shHSP27 and quercetin on
apoptotic rates were analyzed in U937 cells. (A) The apoptotic rate
of U937 cells was analyzed by Annexin V/PI double staining and flow
cytometric analysis. Cell populations in the quadrants were defined
as follows: LL, viable cells; LR, early apoptotic cells; UR, late
apoptotic cells; UL, necrotic cells. (B) Expression of Bcl2 and Bax
in the experimental groups was assessed using western blot
analysis. A representative blot is shown and the Bcl2/Bax ratio
relative to GAPDH was determined by densitometric analysis. Values
are expressed as the mean ± standard deviation. P<0.001 vs. the
control group. PI, propidium iodide; shHSP27, small hairpin RNA
specific for heat shock protein 27; Bcl2, B-cell lymphoma 2; Bax,
Bcl2-associated X protein; UL, upper left; LR, lower right. |
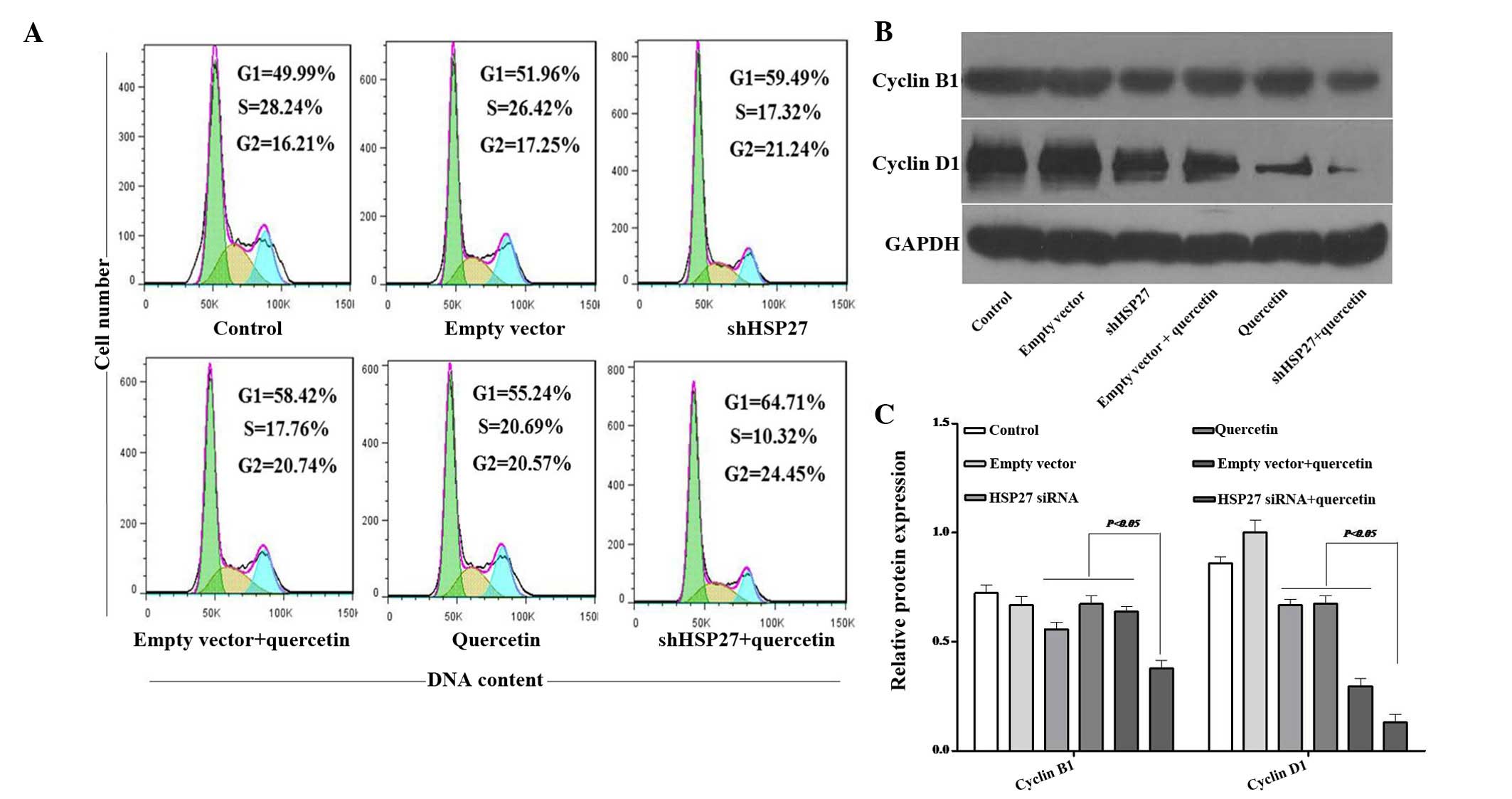
Quecetin plus shHSP27 induces U937-cell
accumulation in G1 phase
Cell proliferation is controlled by the progression
of the cell cycle (38). After
treatment with quercetin alone or shHSP27 with quercetin for 48 h,
the fraction of cells in G1 phase significantly increased from
49.99 to 55.24 and 64.71%, and the fraction of cells in G2 phase
significantly increased from 16.21 to 20.57 and 24.45%, however the
fraction of cells in S phase decreased from 28.24 to 20.69 and
10.32%, respectively (Fig. 4A).
This result indicated that shHSP27 further enhanced the G1-phase
arrest mediated by quercetin. Cyclins have an essential role in the
regulation of the cell cycle (38). Cyclin B1 was the first cell
cycle-associated protein to be identified (39); overexpression of cyclin B1 promotes
cell cycle progression to G2/M-phase and may lead to uncontrolled
cell proliferation and malignant transformation (40,41).
It has been reported that inhibition of cyclin B1 expression
decreases the G2/M-phase population, thereby suppressing cell
growth and inducing apoptosis (42). Cyclin D1 is closely associated with
the proliferation of cancer cells (43). By contrast to cyclin B1, cyclin D1
has an important role in the G1/S-phase transition, which may,
however, promote the occurrence of tumors; it is therefore
considered to be an oncoprotein (44). Quercetin significantly
downregulated the protein expression of cyclin D1; furthermore,
when combined with shHSP27, cyclinD1 expression was almost totally
diminished. In addition, quercetin and shHSP27 reduced the
expression of cyclin B1 (Fig. 4B and
C). These results may explain for the reduced cell-cycle
progression observed by flow cytometry.
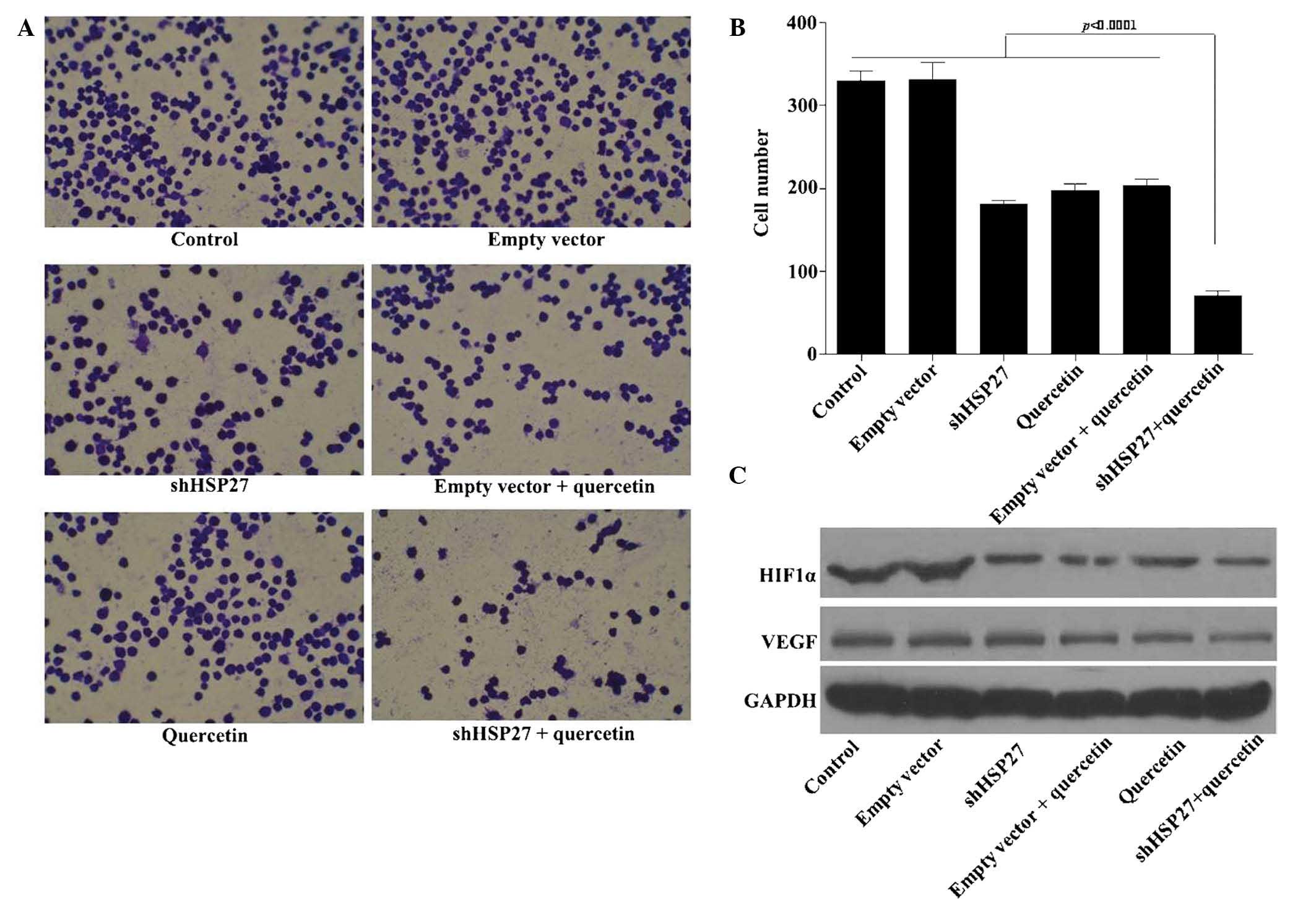
Suppression of HSP27 enhances
quercetin-induced inhibition of U937-cell adhesion
Adhesion of acute myeloid leukemia cells is linked
with resistance to chemotherapy (45,46),
and adhesion of U937 cells to fibronectin via β1 integrins has been
shown to inhibit etoposide- and mitoxantrone-induced apoptosis
(47).
Following treatment with quercetin, the adhesive
capacity of U937 cells dropped by ~30%, while simultaneous
transfection with shHSP27 vector further reduced cell adhesion to
~80% (Fig. 5A and B). This result
indicated that quercetin may serve as an adjuvant for
chemotherapeutics.
Besides cellular adhesion, angiogenesis is required
for invasive tumor-cell metastasis and represents an important
target in the control of cancer progression (48). The HIF1α/VEGF signaling pathway,
which regulates angiogenesis (49), is activated by upstream mTOR
signaling. As shown in Fig. 5C,
shHSP27 and quercetin markedly decreased the protein expression of
HIF1α and VEGF in U937 cells.
Previous studies have indicated that HIF1α increases
the expression of anti-apoptotic Bcl-2 and reduces the expression
of Bax (50); furthermore, in
ovarian cancer cells, phosphoinositide-3 kinase (PI3K) was reported
to mediate G1-to-S-phase progression and cyclin D1 expression
through activation of AKT/mTOR/p70S6K1 signaling (51). In line with this, the results of
the present study suggested that quer-cetin suppresses leukemia
progression by inhibiting mTOR signaling and that this suppressive
function can be further enhanced by shHSP27.
Discussion
Quercetin a major flavonoid contained in foods
including apples, onions, tea, red wine. Several in vitro
studies have shown that quercetin has activity against certain
types of cancer cell (27,52,53).
Furthermore, a clinical study on patients with an inherited
tendency to develop colorectal cancer found that combined dietary
intake of quercetin and curcumin decreased the number and size of
pre-cancerous rectal tumors (54).
Leukemia is a common condition worldwide and affects all age
groups; it is also the most common cancer type in children
(55) and adolescents. In recent
years, the incidence of leukemia has significantly increased
(56).
Studies have found that quercetin inhibits the
proliferation and induces apoptosis in human leukemia cells
(9,34); however, to the best of our
knowledge, the anti-tumor effects of quercetin combined with
shHSP27, as well as the underlying molecular mechanisms, have not
been reported. The present study evaluated the anti-tumor effects
of quercetin on the U937 acute myeloid leukemia cell line with
HSP27 knockdown.
The results showed that the time-dependent
inhibition of the proliferation of U937 cells by quercetin was
enhanced with simultaneous transfection of shHSP27. Furthermore,
cell cycle analysis showed that quercetin plus shHSP27
significantly induced the accumulation of U937 cells in G1 phase
with a minor increase in G2 phase, which indicated that quercetin
plus shHSP27 may inhibit the proliferation of U937 cells by
blocking the cell cycle. In addition, the expression of cell
cycle-associated proteins cyclin D1 and cyclin B1 in U937 cells
treated with quercetin plus shHSP27 was decreased following
treatment with quercetin plus shHSP27. Cyclin D1 is mainly involved
in G1/S-phase transition (44),
while cyclin B1 is associated with progression to G2/M phase. These
results indicated that shHSP27 plus quercetin blocked cell cycle
progression by inhibiting the expression of the cell-cycle proteins
cyclin D1 and -B1.
Evasion of apoptosis is a key factor during
carcinogenesis, cancer progression and drug resistance, while
induction of apoptosis is a desirable property of anti-cancer
treatments (57). The present
study found that the percentage of apoptotic U937 cells
significantly increased after treatment with quercetin plus
shHSP27. Furthermore, the expression of apoptotic signaling
proteins was detected, which revealed that the expression of
anti-apoptotic protein Bcl-2 decreased, while that of pro-apoptotic
protein Bax in U937 cells increased when treated with shHSP27 and
quercetin. This result indicated that shHSP27 plus quercetin
induced cell apoptosis by reducing the Bcl-2/Bax ratio in U937
cells.
Cell adhesion has a vital role in tumor metastasis
and the adhesion assay performed in the present study revealed a
significant inhibitory effect of shHSP27 plus quercetin compared
with that of quercetin only. In addition, angiogenesis is
associated with the occurrence and prognosis of leukemia (58,59).
VEGF, the primary factor stimulating blood-vessel growth, has been
confirmed to be elevated in leukemia and is vital for its
pathogenesis and progression (60). The present study revealed that
quercetin plus shHSP27 inhibited VEGF expression in U937 cells.
The Akt and mTOR (PI3K/Akt/mTOR) signaling network
is important in leukemia, where it regulates cell growth and
survival (61). Proteins
demonstrated to be involved in regulating apoptosis, cell cycle and
angiogenesis of U937 cells in the present study have all been
confirmed as parts of the mTOR signaling network in other diseases
(28,30,35,51,61,62),
while they have not been previously determined in leukemia. Levels
of p-AKT and p-mTOR were decreased following treatment with shHSP27
plus quercetin; furthermore, Notch1, Bcl-2, cyclin D1, HIF1α and
VEGF were decreased. These proteins constitute a huge network
regulated by mTOR expression, and mTOR signaling has a critical
role in the regulation of tumor-cell motility, invasion and cancer
metastasis (63). In order to
further determine the therapeutic potential of quercetin plus
shHSP27 in leukemia, the effects of quercetin on the mTOR-regulated
signaling network require further elucidation.
In conclusion, the present study demonstrated that,
compared with quercetin or shHSP27 alone, combined treatment with
quercetin plus shHSP27 significantly inhibited the proliferation
and adhesion as well as induced apoptosis and cell-cycle arrest in
U937 human leukemia cells. These anti-tumor effects may mainly
depend on the inhibition of the mTOR signal network. Quercetin plus
shHSP27 synergistically decreased the cellular adhesion capacity of
leukemia cells and HSP27 knockdown may therefore potentiate the
efficiency of chemotherapies of human leukemia.
Acknowledgments
This study was supported by the Scientific Research
Project of Heilongjiang Province Department of Education (no.
12541530).
References
|
1
|
Kuttan G, Pratheeshkumar P, Manu KA and
Kuttan R: Inhibition of tumor progression by naturally occurring
terpenoids. Pharm Biol. 49:995–1007. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Pratheeshkumar P, Sreekala C, Zhang Z,
Budhraja A, Ding S, Son YO, Wang X, Hitron A, Hyun-Jung K, Wang L,
et al: Cancer prevention with promising natural products:
Mechanisms of action and molecular targets. Anticancer Agents Med
Chem. 12:1159–1184. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Larson AJ, Symons JD and Jalili T:
Therapeutic potential of quercetin to decrease blood pressure:
Review of efficacy and mechanisms. Adv Nutr. 3:39–46. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Okamoto T: Safety of quercetin for
clinical application (Review). Int J Mol Med. 16:275–278.
2005.PubMed/NCBI
|
|
5
|
Formica JV and Regelson W: Review of the
biology of Quercetin and related bioflavonoids. Food Chem Toxicol.
33:1061–1080. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Lee JC, Kim J, Park JK, Chung GH and Jang
YS: The antioxidant, rather than prooxidant, activities of
quercetin on normal cells: Quercetin protects mouse thymocytes from
glucose oxidase-mediated apoptosis. Exp Cell Res. 291:386–397.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Liesveld JL, Abboud CN, Lu C, McNair C,
Menon A, Smith A, Rosell K and Rapoport AP: Flavonoid effects on
normal and leukemic cells. Leuk Res. 27:517–527. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Cheng S, Gao N, Zhang Z, Chen G, Budhraja
A, Ke Z, Son YO, Wang X, Luo J and Shi X: Quercetin induces
tumor-selective apoptosis through downregulation of Mcl-1 and
activation of Bax. Clin Cancer Res. 16:5679–5691. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Kang TB and Liang NC: Studies on the
inhibitory effects of quercetin on the growth of HL-60 leukemia
cells. Biochem Pharmacol. 54:1013–1018. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Román-Gómez J, Cordeu L, Agirre X,
Jiménez-Velasco A, San José-Eneriz E, Garate L, Calasanz MJ,
Heiniger A, Torres A and Prosper F: Epigenetic regulation of
Wnt-signaling pathway in acute lymphoblastic leukemia. Blood.
109:3462–3469. 2007. View Article : Google Scholar
|
|
11
|
Kimura A: Radiation associated leukemia
and myelodysplastic syndrome. Nihon Rinsho. 70:431–435. 2012.In
Chinese. PubMed/NCBI
|
|
12
|
Noshchenko AG, Bondar OY and Drozdova VD:
Radiation-induced leukemia among children aged 0–5 years at the
time of the Chernobyl accident. Int J Cancer. 127:412–426.
2010.
|
|
13
|
Malagoli C, Malavolti M, Costanzini S,
Fabbri S, Tezzi S, Palazzi G, Arcolin E and Vinceti M: Increased
incidence of childhood leukemia in urban areas: a population-based
case-control study. Epidemiol Prev. 39:102–107. 2015.PubMed/NCBI
|
|
14
|
Garcia-Perez J, López-Abente G,
Gómez-Barroso D, Morales-Piga A, Romaguera EP, Tamayo I,
Fernández-Navarro P and Ramis R: Childhood leukemia and residential
proximity to industrial and urban sites. Environ Res. 140:542–553.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Zeeshan R, Sultan S, Irfan SM, Kakar J and
Hameed MA: Clinico-hematological profile of patients with B-chronic
lymphoid leukemia in Pakistan. Asian Pac J Cancer Prev. 16:793–796.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Khalafalla MM, Abdellatef E, Daffalla HM,
et al: Antileukemia activity from root cultures of Vernonia
amygdalina. J Med Plants Res. 3:556–562. 2009.
|
|
17
|
Bukau B and Horwich AL: The Hsp70 and
Hsp60 chaperone machines. Cell. 92:351–366. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Polla BS, Kantengwa S, Gleich GJ, Kondo M,
Reimert CM and Junod AF: Spontaneous heat shock protein synthesis
by alveolar macrophages in interstitial lung disease associated
with phagocytosis of eosinophils. Eur Respir J. 6:483–488.
1993.PubMed/NCBI
|
|
19
|
Taba K, Kuramitsu Y, Ryozawa S, Yoshida K,
Tanaka T, Maehara S, Maehara Y, Sakaida I and Nakamura K:
Heat-shock protein 27 is phosphorylated in gemcitabine-resistant
pancreatic cancer cells. Anticancer Res. 30:2539–2543.
2010.PubMed/NCBI
|
|
20
|
Carper SW, Duffy JJ and Gerner EW: Heat
shock proteins in thermotolerance and other cellular processes.
Cancer Res. 47:5249–5255. 1987.PubMed/NCBI
|
|
21
|
Ricci JE, Maulon L, Battaglione-Hofman V,
Bertolotto C, Luciano F, Mari B, Hofman P and Auberger P: A Jurkat
T cell variant resistant to death receptor-induced apoptosis.
Correlation with heat shock protein (Hsp) 27 and 70 levels. Eur
Cytokine Netw. 12:126–134. 2001.PubMed/NCBI
|
|
22
|
Pandey P, Farber R, Nakazawa A, Kumar S,
Bharti A, Nalin C, Weichselbaum R, Kufe D and Kharbanda S: Hsp27
functions as a negative regulator of cytochrome c-dependent
activation of procaspase-3. Oncogene. 19:1975–1981. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Garrido C, Ottavi P, Fromentin A, Hammann
A, Arrigo AP, Chauffert B and Mehlen P: HSP27 as a mediator of
confluence-dependent resistance to cell death induced by anticancer
drugs. Cancer Res. 57:2661–2667. 1997.PubMed/NCBI
|
|
24
|
Garrido C, Bruey JM, Fromentin A, Hammann
A, Arrigo AP and Solary E: HSP27 inhibits cytochrome c-dependent
activation of procaspase-9. FASEB J. 13:2061–2070. 1999.PubMed/NCBI
|
|
25
|
Piantelli M, Tatone D, Castrilli G, Savini
F, Maggiano N, Larocca LM, Ranelletti FO and Natali PG: Quercetin
and tamoxifen sensitize human melanoma cells to hyperthermia.
Melanoma Res. 11:469–476. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Jakubowicz-Gil J, Rzymowska J and Gawron
A: Quercetin, apoptosis, heat shock. Biochem Pharmacol.
64:1591–1595. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Chen SF, Nieh S, Jao SW, Liu CL, Wu CH,
Chang YC, Yang CY and Lin YS: Quercetin suppresses drug-resistant
spheres via the p38 MAPK-Hsp27 apoptotic pathway in oral cancer
cells. PLoS One. 7:e492752012. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Zhou H and Huang S: Role of mTOR signaling
in tumor cell motility, invasion and metastasis. Curr Protein Pept
Sci. 12:30–42. 2011. View Article : Google Scholar
|
|
29
|
Efeyan A and Sabatini DM: mTOR and cancer:
Many loops in one pathway. Curr Opin Cell Biol. 22:169–176. 2010.
View Article : Google Scholar :
|
|
30
|
Pratheeshkumar P, Budhraja A, Son YO, Wang
X, Zhang Z, Ding S, Wang L, Hitron A, Lee JC, Xu M, et al:
Quercetin inhibits angiogenesis mediated human prostate tumor
growth by targeting VEGFR- 2 regulated AKT/mTOR/P70S6K signaling
pathways. PLoS One. 7:e475162012. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Bruning A: Inhibition of mTOR signaling by
quercetin in cancer treatment and prevention. Anticancer Agents Med
Chem. 13:1025–1031. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
O'Callaghan-Sunol C, Gabai VL and Sherman
MY: Hsp27 modulates p53 signaling and suppresses cellular
senescence. Cancer Res. 67:11779–11788. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Niu G, Yin S, Xie S, Li Y, Nie D, Ma L,
Wang X and Wu Y: Quercetin induces apoptosis by activating
caspase-3 and regulating Bcl-2 and cyclooxygenase-2 pathways in
human HL-60 cells. Acta Biochim Biophys Sin (Shanghai). 43:30–37.
2011. View Article : Google Scholar
|
|
34
|
Mahbub AA, Le Maitre CL, Haywood-Small SL,
McDougall GJ, Cross NA and Jordan-Mahy N: Differential effects of
polyphenols on proliferation and apoptosis in human myeloid and
lymphoid leukemia cell lines. Anticancer Agents Med Chem.
13:1601–1613. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Hansel DE, Platt E, Orloff M, Harwalker J,
Sethu S, Hicks JL, De Marzo A, Steinle RE, Hsi ED, Theodorescu D,
et al: Mammalian target of rapamycin (mTOR) regulates cellular
proliferation and tumor growth in urothelial carcinoma. Am J
Pathol. 176:3062–3072. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Osorio LM, Jondal M and Aguilar-Santelises
M: Regulation of B-CLL apoptosis through membrane receptors and
Bcl-2 family proteins. Leuk Lymphoma. 30:247–256. 1998.PubMed/NCBI
|
|
37
|
Podhorecka M, Halicka D, Klimek P, Kowal
M, Chocholska S and Dmoszynska A: Simvastatin and purine analogs
have a synergic effect on apoptosis of chronic lymphocytic leukemia
cells. Ann Hematol. 89:1115–1124. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Malumbres M and Barbacid M: Cell cycle,
CDKs and cancer: A changing paradigm. Nat Rev Cancer. 9:153–166.
2009. View
Article : Google Scholar : PubMed/NCBI
|
|
39
|
Evans T, Rosenthal ET, Youngblom J, Distel
D and Hunt T: Cyclin: A protein specified by maternal mRNA in sea
urchin eggs that is destroyed at each cleavage division. Cell.
33:389–396. 1983. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Hwang A, Maity A, McKenna WG and Muschel
RJ: Cell cycle-dependent regulation of the cyclin B1 promoter. J
Biol Chem. 270:28419–28424. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Hartwell LH and Kastan MB: Cell cycle
control and cancer. Science. 266:1821–1828. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Yuan J, Kramer A, Matthess Y, Yan R,
Spänkuch B, Gätje R, Knecht R, Kaufmann M and Strebhardt K: Stable
gene silencing of cyclin B1 in tumor cells increases susceptibility
to taxol and leads to growth arrest in vivo. Oncogene.
25:1753–1762. 2006. View Article : Google Scholar
|
|
43
|
Nakata Y, Shetzline S, Sakashita C, Kalota
A, Rallapalli R, Rudnick SI, Zhang Y, Emerson SG and Gewirtz AM:
c-Myb contributes to G2/M cell cycle transition in human
hematopoietic cells by direct regulation of cyclin B1 expression.
Mol Cell Biol. 27:2048–2058. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Donnellan R and Chetty R: Cyclin D1 and
human neoplasia. Mol Pathol. 51:1–7. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Becker PS: Dependence of acute myeloid
leukemia on adhesion within the bone marrow microenvironment.
Scientific World Journal. 2012:8564672012. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Koukoulis GK, Patriarca C and Gould VE:
Adhesion molecules and tumor metastasis. Hum Pathol. 29:889–892.
1998. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Hazlehurst LA, Valkov N, Wisner L, Storey
JA, Boulware D, Sullivan DM and Dalton WS: Reduction in
drug-induced DNA double-strand breaks associated with beta1
integrin-mediated adhesion correlates with drug resistance in U937
cells. Blood. 98:1897–1903. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Folkman J: Role of angiogenesis in tumor
growth and metastasis. Semin Oncol. 29(Suppl 6): S15–S18. 2002.
View Article : Google Scholar
|
|
49
|
Dai Y, Xu M, Wang Y, Pasha Z, Li T and
Ashraf M: HIF-1alpha induced-VEGF overexpression in bone marrow
stem cells protects cardiomyocytes against ischemia. J Mol Cell
Cardiol. 42:1036–1044. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Sasabe E, Tatemoto Y, Li D, Yamamoto T and
Osaki T: Mechanism of HIF-1alpha-dependent suppression of
hypoxia-induced apoptosis in squamous cell carcinoma cells. Cancer
Sci. 96:394–402. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Gao N, Flynn DC, Zhang Z, Zhong XS, Walker
V, Liu KJ, Shi X and Jiang BH: G1 cell cycle progression and the
expression of G1 cyclins are regulated by PI3K/AKT/mTOR/p70S6K1
signaling in human ovarian cancer cells. Am J Physiol Cell Physiol.
287:C281–C291. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Rong Y, Yang EB, Zhang K and Mack P:
Quercetin-induced apoptosis in the monoblastoid cell line U937 in
vitro and the regulation of heat shock proteins expression.
Anticancer Res. 20(6B): 4339–4345. 2000.
|
|
53
|
Scambia G, Ranelletti FO, Panici PB, De
Vincenzo R, Bonanno G, Ferrandina G, Piantelli M, Bussa S, Rumi C
and Cianfriglia M: Quercetin potentiates the effect of adriamycin
in a multidrug-resistant MCF-7 human breast-cancer cell line:
P-glycoprotein as a possible target. Cancer Chemother Pharmacol.
34:459–464. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Volate SR, Davenport DM, Muga SJ and
Wargovich MJ: Modulation of aberrant crypt foci and apoptosis by
dietary herbal supplements (quercetin, curcumin, silymarin, ginseng
and rutin). Carcinogenesis. 26:1450–1456. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Zwaan CM, Kolb EA, Reinhardt D,
Abrahamsson J, Adachi S, Aplenc R, De Bont ES, De Moerloose B,
Dworzak M, Gibson BE, et al: Collaborative Efforts Driving Progress
in Pediatric Acute Myeloid Leukemia. J Clin Oncol. 33:2949–2962.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Katz AJ, Chia VM, Schoonen WM and Kelsh
MA: Acute lymphoblastic leukemia: an assessment of international
incidence, survival, and disease burden. Cancer Causes Control.
26:1627–1642. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Gregoire M, Ligeza-Poisson C,
Juge-Morineau N and Spisek R: Anti-cancer therapy using dendritic
cells and apoptotic tumour cells: pre-clinical data in human
mesothelioma and acute myeloid leukaemia. Vaccine. 21:791–794.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Schmidt T and Carmeliet P: Angiogenesis: A
target in solid tumors, also in leukemia? Hematology Am Soc Hematol
Educ Program. 2011:1–8. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Trujillo A, McGee C and Cogle CR:
Angiogenesis in acute myeloid leukemia and opportunities for novel
therapies. J Oncol. 2012:1286082012. View Article : Google Scholar
|
|
60
|
Kampen KR, Ter Elst A and de Bont ES:
Vascular endothelial growth factor signaling in acute myeloid
leukemia. Cell Mol Life Sci. 70:1307–1317. 2013. View Article : Google Scholar
|
|
61
|
Neri LM, Cani A, Martelli AM, Simioni C,
Junghanss C, Tabellini G, Ricci F, Tazzari PL, Pagliaro P, McCubrey
JA and Capitani S: Targeting the PI3K/Akt/mTOR signaling pathway in
B-precursor acute lymphoblastic leukemia and its therapeutic
potential. Leukemia. 28:739–748. 2014. View Article : Google Scholar
|
|
62
|
Wang K, Liu R, Li J, Mao J, Lei Y, Wu J,
Zeng J, Zhang T, Wu H, Chen L, et al: Quercetin induces protective
autophagy in gastric cancer cells: Involvement of Akt-mTOR- and
hypoxia-induced factor 1α-mediated signaling. Autophagy. 7:966–978.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Alayev A and Holz MK: mTOR signaling for
biological control and cancer. J Cell Physiol. 228:1658–1664. 2013.
View Article : Google Scholar : PubMed/NCBI
|