Introduction
Parkinson's disease (PD) is the second most common
progressive neurodegenerative disorder worldwide, which is well
characterized by the degeneration of dopaminergic neurons in the
substantia nigra pars compacta (SNpc). The diagnosis of PD is
associated with various symptoms of motor dysfunction, including
progressive resting tremor, rigidity, bradykinesia and postural
instability (1). It has previously
been suggested that the progression of PD may be associated with
mitochondrial dysfunction (2), and
mitochondrial complex I inhibition is not limited to the brain in
PD. The brain is particularly susceptible to oxidative damage due
to its high expression of polyunsaturated fatty acids and
relatively low antioxidant activity (3); oxidative stress has been reported to
have a crucial role in the pathogenesis of PD, and is considered
the most important underlying mechanism (4). In addition, oxidative stress and
mitochondrial dysfunction have been identified as early events
during the process of neurodegeneration (5). The majority of research regarding
neurodegeneration in PD has focused on the brain, and most of the
resultant oxidative damage has been detected in the SNpc. A marked
decrease in glutathione (GSH) levels in the SNpc is considered to
be the earliest triggering event of neurodegeneration, which
precedes oxidative stress (6),
mitochondrial dysfunction and dopamine loss during early PD
(7). Therefore, restoration of
cellular GSH levels may prevent the initiation of PD.
Rotenone (ROT) is an active herbicide that can be
used to induce dopaminergic neuronal oxidative damage in
experimental animal models of PD (8). The polyphenolic flavonoid curcumin
[1,7-bis(4-hydroxy-3-methoxyphenyl)-1,6-heptadiene-3,5-dione],
which is a principle coloring agent extracted from the rhizomes of
Curcuma longa (9), is a
dietary spice that is often used in Indian cuisine and medicine
(10). Previous studies have
demonstrated that curcumin possesses various properties, including
antioxidant (11),
anti-inflammatory (12),
anticancer (13), iron-chelating
(14) and neuroprotective
activities (15). Notably,
previous research regarding various experimental models of PD
strongly supports the clinical application of curcumin in PD
(16). The specific underlying
mechanism is not fully understood; however, curcumin has exerted
promising antioxidant effects in various cell and animal models of
PD through modulating the generation of GSH, reactive oxygen
species (ROS) and malondialdehyde (MDA) (9,17,18).
Furthermore, our previous study demonstrated that curcumin was able
to attenuate ROT-induced cytotoxicity by exerting antioxidative and
antiapoptotic activity in PC12 rat pheochromocytoma cells, thus
suggesting that curcumin has neuroprotective effects on the
dopaminergic neurons of PD (19).
However, there is still a lack of data with regards to the
molecular mechanism underlying the protective effects of curcumin
in animal models of neurodegenerative diseases.
Previous studies have demonstrated that Akt
phosphorylation facilitates nuclear factor erythroid 2-related
factor 2 (Nrf2) translocation into the nucleus, which has an
important role in the defense against oxidative stress by
regulating the expression levels of antioxidant enzymes, including
heme oxygenase (HO)-1 and NAD(P)H:quinone oxidoreductase 1 (NQO1)
(20–22). However, the majority of studies
regarding the effects of ROT or curcumin on the progression of PD
have focused on in vitro cell models (18,19,23).
The present study aimed to investigate the effects of curcumin on
ROT-induced neurotoxicity and oxidative dysfunction, and to
determine the underlying mechanism associated with PD
progression.
Materials and methods
Animal care and cell culture
A total of 50 male Lewis rats (age, 7–8 weeks;
weight, 260–280 g) were purchased from Beijing Vital River
Laboratory Animal Technology Co., Ltd. (Beijing, China). The rats
were housed in a room maintained at 22°C with 40–70% humidity,
under a 12-h light/dark cycle with ad libitum access to
drinking water and a standard pellet diet. All scientific
procedures were carried out in accordance with the relevant
Guidelines in Neuroscience Research and the Care and Use of
Laboratory Animals of the Second Affiliated Hospital of Zhengzhou
University (Zhengzhou, China). The present study was approved by
the local ethics committee of Zhengzhou University (Zhengzhou,
China).
Human embryonic kidney (HEK)293T cells and SK-N-SH
human neuroblastoma cells were obtained from the American Type
Culture Collection (Manassas, VA, USA). The cells were cultured in
Dulbecco's modified Eagle's medium (Gibco; Thermo Fisher
Scientific, Inc., Waltham, MA, USA) supplemented with 4.5 g/l high
glucose, 2 mM L-glutamine, 1 mM sodium pyruvate (Thermo Fisher
Scientific, Inc.) and 10% heat-inactivated fetal bovine serum
(Gibco; Thermo Fisher Scientific, Inc.) at 37°C in an atmosphere
containing 5% CO2 and 95% air.
Experimental design
Rotenone (Sigma-Aldrich, St. Louis, MO, USA) was
dissolved in sunflower seed oil at 1.5 mg/ml. The rats were divided
randomly into four groups (n=6/group): (i) The CON group, in which
rats were injected with sunflower seed oil (0.5 ml/kg/day, twice
daily) as a vehicle for 50 days; (ii) the ROT + curcumin treatment
group (Sigma-Aldrich), in which rats were intragastrically
administered curcumin (100 mg/kg) twice a day for 50 days, and ROT
was subcutaneously administered twice a day at 1 ml/kg/day on day 4
of curcumin treatment; (iii) the ROT treatment group, in which the
rats received the same as the ROT + curcumin group beginning on day
1, except that gum arabica (Sigma-Aldrich) was administered instead
of curcumin; (iv) the curcumin treatment group, in which the rats
received the same as the ROT + curcumin group, except that
sunflower seed oil was administered instead of ROT. During the
experimental period, the food intake was monitored daily and body
weight was measured regularly. Behavioral assessments were carried
out among rats of all groups. Subsequently, the rats were
sacrificed by decapitation, and tissues were removed from the brain
on a Petri dish placed on ice under a microscope (Olympus BX50,
Tokyo, Japan). The SNpc regions were stored at −80°C, and the
cytosolic fraction was subsequently extracted and subjected to
various biochemical investigations and western blot analyses.
Apomorphine-induced rotations
A total of 24 h after the final injection, the
rotational behavior of the rats was determined following
intraperitoneal injection with freshly prepared apomorphine
hydrochloride (1 mg/kg, s.c.; Sigma-Aldrich) for 5–10 min.
Subsequently, full 360° unilateral turns were recorded in a 10 min
period. The rats that turned unilaterally >7 cycles/min, were
considered successful models of PD.
Catalepsy and open field tests
In the catalepsy test, rats were hung by the paws on
a vertical grid (25×45 cm, 1 cm space between each wire). The time
taken for the rats to move their paws or to perform any sort of
initial movement was noted. The rats were then placed with both
forepaws on a bar 10 cm above, parallel from the base in a
half-rearing position. The paw removal latency was noted, and the
maximum descent latency was fixed at 180 sec for both tests.
Behavioral tests were carried out in an open field
using a spontaneous activity instrument (Shanghai Jiliang Software
Technology Co., Ltd., Shanghai, China), including four boxes
(40×40×50 cm). Each box housed one rat at a time. An infrared
camera was attached to the top of the box to record the movement
time, movement distance and movement speed for 1 h. Subsequently,
the video images were sent to a computer and analyzed with the
JLBehv-LAR-4 software (Shanghai Jiliang Software Technology Co.,
Shanghai, China).
Biochemical measurements of GSH, ROS and
MDA
The obtained SNpc regions were immediately
homogenized using an ultrasonic cell disruptor (Model 150 V/T;
BioLogics, Manassas, VA, USA), and then subjected to centrifugation
at 14,000 × g for 15 min at 4°C. The supernatant was used for the
determination of the activity of various enzyme activities. GSH
content in the SNpc regions was measured as described previously
(24), and the fluorescence was
measured using a spectrofluorometer (Model RF-5000, Shimadzu
Corporation, Tokyo, Japan). GSH concentration was calculated from a
standard curve and expressed as µg GSH/mg protein. ROS
generation in the SNpc regions was assayed using
dihydrodichlorofluorescein diacetate, as described previously
(25). ROS formation was
quantified from a dichlorodihydrofluorescein (DCF) standard curve
and expressed as pmol DCF formed/min/mg protein. Lipid peroxidation
(LPO) levels in the cytosol of the SNpc were quantified by
measuring the formation of thiobarbituric acid reactive substances
(TBARS), as described previously (26). The absorbance of TBARS adducts was
measured at 532 nm, which was used to quantify MDA levels. The
commercial kits used to measure the levels of GSH, ROS and MDA were
purchased from Nanjing Jiancheng Bioengineering Institute (Nanjing,
China) according to the manufacturer's instructions.
Lentivirus preparation
HEK293T cells were co-transfected with envelope
plasmid, packaging plasmid and expression plasmid (pLKO.1;
Sigma-Aldrich) containing the shRNA using Lipofectamine®
2000 (Invitrogen; Thermo Fisher Scientific, Inc.) and the
Lentiviral Packaging Mix (Sigma-Aldrich), according to the
manufacturers' protocols. The short hairpin (sh)RNA sequence
targeting coding regions of the rat Nrf2 gene (shNrf2) and the
control shScramble plasmid (GenePharma Co., Ltd., Shanghai, China)
were used. The cell supernatant containing the lentivirus was
collected, and filtered through membrane filters (0.45-µm
pore size, Millipore Corporation, Bedford, MA, USA). Then, viral
supernatant was concentrated by ultracentrifugation for 2 h at
50,000 × g and 4°C and stored at −80°C until further use. The
number of relative infection units (RIU) was determined by
infecting SK-N-SH cells with a serial dilution.
Infusion with a shNrf2 lentivirus or
phosphoinositide 3-kinase (PI3K) inhibitor LY294002
In the shNrf2 lentivirus experiment, nine groups
(n=6/group) were designed: CON group; shScramble (control) group;
shScramble + ROT group; shScramble + curcumin group; shScramble +
ROT + curcumin group; shNrf2 group; shNrf2 + ROT group; shNrf2 +
curcumin group; shNrf2 + ROT + curcumin group. Lentiviruses
containing shScramble or shNrf2 (2×107 RIU; 10
µl) were infused into the SNpc regions with an infusion rate
of 0.5 µl/min. After 48 h recovery, the rats were treated
with ROT and/or curcumin for 50 days as described previously.
In the LY294002 experiment, five groups (n=6/group)
were designed: CON group; LY294002 (control) group; LY294002 + ROT
group; LY294002 + curcumin group; LY294002 + ROT + curcumin group.
ALZET® osmotic mini-pumps (0.5 µl/h; model 1007D;
DURECT Corporation, ALZET Osmotic Pumps, Cupertino, CA, USA) were
subcutaneously implanted on the back of the rats over the
latissimus dorsi muscles, and were connected by catheters to the
infusion cannula for intracerebroventricular infusion. The
inhibitor-treated rats received osmotic minipumps filled with
artificial cerebral spinal fluid (aCSF; Gibco; Thermo Fisher
Scientific, Inc.) alongside 0.1 µg/µl LY294002
(Sigma-Aldrich). The CON rats received equal volumes of
phosphate-buffered saline in aCSF. The pumps were implanted 5 min
after the first injection of ROT or curcumin, and remained active
for 7 days. The rats were treated with ROT and curcumin for 50
days, as described previously. Subsequently, all rats were
sacrificed and the SNpc regions of the brains were collected for
western blot analysis.
Western blot analysis
Tissues were homogenized on ice using
radioimmunoprecipitation assay lysis buffer (Beyotime Institute of
Biotechnology, Shanghai, China) and were centrifuged at 14,000 × g
for 15 min at 4°C. Protein concentration was determined using the
Pierce Bicinchoninic Acid Protein Assay kit (Pierce Biotechnology,
Inc., Rockford, IL, USA). Equal amounts of protein (25 µg
per lane) were loaded onto 10% sodium dodecyl
sulfate-polyacrylamide gel (Shanghai Sangon Biotechnology Co. Ltd.,
Shanghai, China), prior to being transferred onto polyvinylidene
difluoride membranes (PVDF; Millipore Corporation). Subsequently,
the membranes were blocked with 5% non-fat milk in 0.1%
Tris-buffered saline with Tween-20 (TBST; Shanghai Sangon
Biotechnology Co. Ltd.) for 1 h at room temperature, followed by
incubation with the primary antibodies overnight at 4°C. The
primary antibodies used were rabbit anti-tyrosine hydroxylase (TH)
polyclonal antibody (cat. no. T8700; Sigma-Aldrich), mouse
anti-β-actin monoclonal antibody (cat. no. A1978) and rabbit
anti-HO-1 polyclonal antibody (cat. no. H4535) (dilution, 1:1,000;
Sigma-Aldrich); rabbit anti-NQO1 polyclonal antibody (cat. no.
sc-25591) and rabbit anti-Nrf2 polyclonal antibody (cat. no.
sc-722) (dilution, 1:1,000; Santa Cruz Biotechnology, Inc., Dallas,
TX, USA); rabbit anti-phosphorylated (p)-Nrf2 (Ser40) monoclonal
antibody (cat. no. ab76026, dilution, 1:5,000; Abcam, Cambridge,
MA, USA); rabbit anti-Akt polyclonal antibody (cat. no. #9272) and
rabbit anti-p-Akt (Ser473) polyclonal antibody (cat. no. #9271)
(dilution, 1:1,000; Cell Signaling Technology, Inc., Beverly, MA,
USA). After incubation with appropriate goat anti-rabbit or
anti-mouse immunoglobulin G conjugated to horseradish peroxidase
(dilution, 1:5,000; Beyotime Institute of Biotechnology) for 1 h at
room temperature, the resultant protein bands were visualized by
enhanced chemiluminescence (Beyotime Institute of Biotechnology).
The absorbance values of target proteins were determined using
Gel-Pro Analyzer version 4.0 software (Media Cybernetics, Inc.,
Rockville, MD, USA).
Statistical analysis
Data are presented as the mean ± standard deviation
representative of three individual experiments performed in
triplicate. Statistical analyses were conducted using one-way
analysis of variance followed by the Student Newman-Keuls test.
Statistical analyses were performed using GraphPad Prism software
5.0 (GraphPad Software Inc., La Jolla, CA, USA). P<0.05 was
considered to indicate a statistically significant difference.
Results
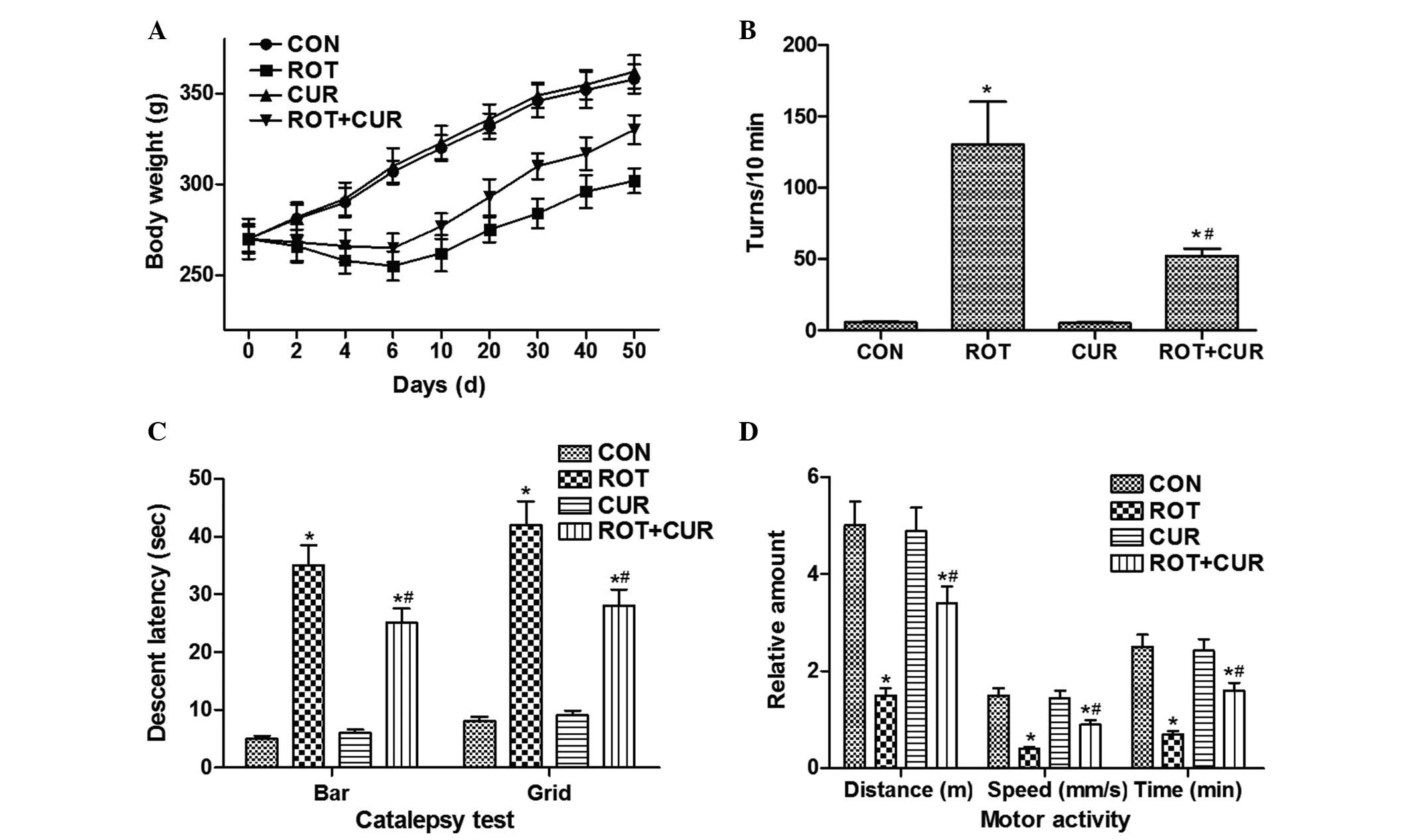
Curcumin partly alleviates ROT-induced
symptoms of PD
Almost all ROT-treated rats displayed significant
weight loss during the first 10 days of ROT treatment, which was
reversed after curcumin pretreatment. In the CON and
curcumin-treated rats, body weight increased slowly and no
significant difference was observed (Fig. 1A). In the rotational behavior test,
the curcumin-treated rats exhibited similar numbers of rotation
(5.1±0.26 turns/10 min), as compared with the CON group (5.6±0.24
turns/10 min; P>0.05); however, the ROT-treated rats exhibited
markedly higher rotational numbers (130±20.16 turns/10 min;
P<0.05). Notably, curcumin administration markedly suppressed
the ROT-induced enhanced rotational behavior (52±8.8 turns/10 min;
P<0.05) (Fig. 1B).
ROT-treated rats developed typical disordered
behavioral features, including catalepsy, unsteady movement and
hunched posture to varying degrees. ROT-treated rats exhibited
prolonged descent latency in the grid and bar test (Fig. 1C) and were considered hypokinetic;
the movement time, movement distance and movement speed of the
ROT-treated rats were markedly reduced, as compared with the CON
group (Fig. 1D). Furthermore,
curcumin pretreatment alleviated these symptoms, and no obvious
PD-associated postural or behavioral features were observed in the
CON and curcumin-treated groups. These data suggest that PD may be
induced in rats via chronic treatment with ROT, and the
parkinsonian symptoms may be partly alleviated following curcumin
pretreatment.
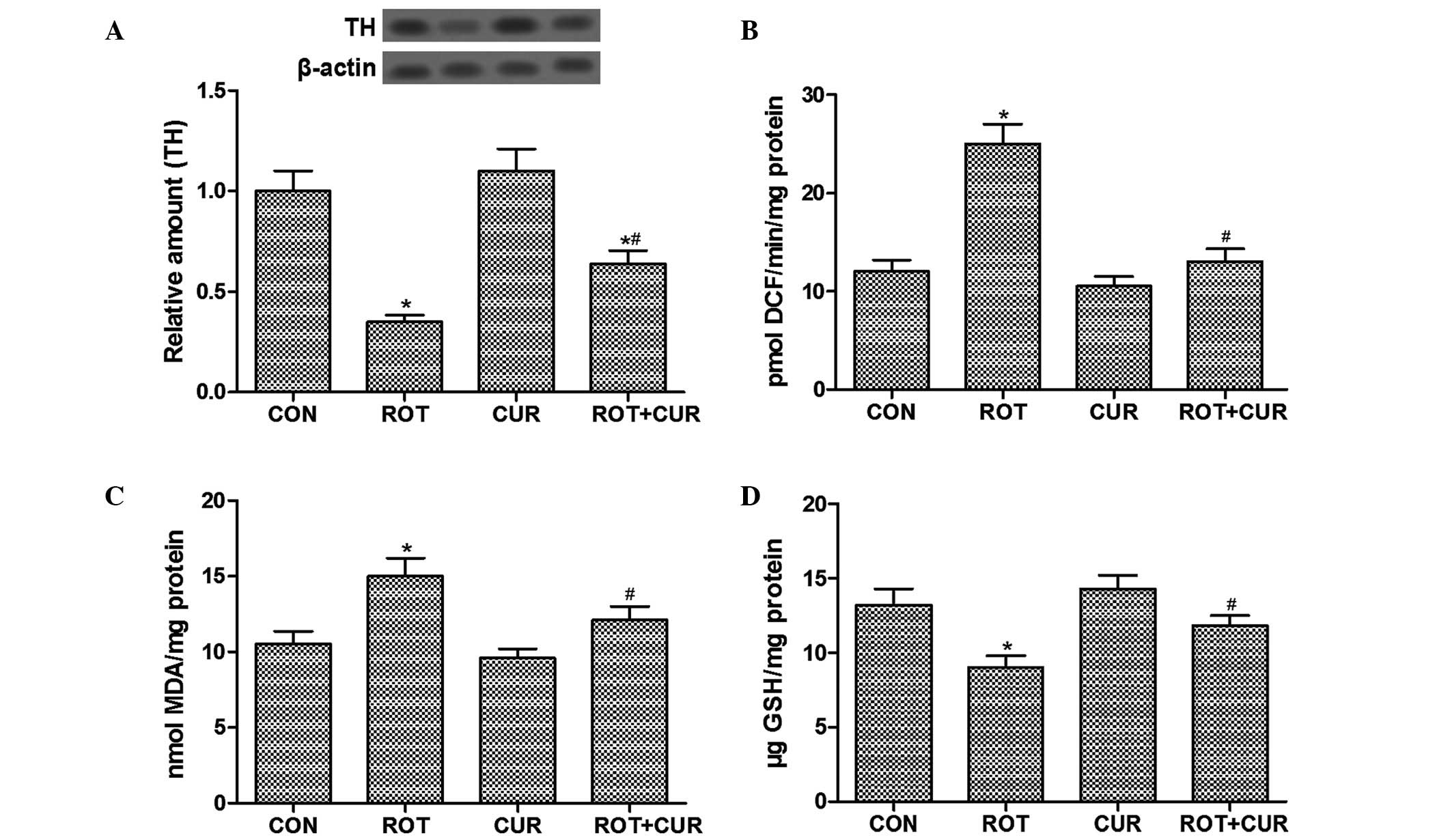
Curcumin ameliorates ROT-induced
dopaminergic neuronal damage and oxidative injury
The present study aimed to determine the protective
effects of curcumin on the neurochemical features of dopaminergic
neurons in ROT-treated rats. Western blot analysis revealed that
curcumin pretreatment restored the ROT-induced loss of TH protein,
which is considered a marker of dopaminergic neurons, in the SNpc
(Fig. 2A). ROS generation in the
SNpc was significantly enhanced (~105%) following treatment with
ROT exposure, as compared with in the CON group (Fig. 2B). Similarly, ROT-treated rats
displayed higher levels of LPO (MDA levels) with a 43% elevation,
as compared with the CON rats (Fig.
2C). Furthermore, GSH content was reduced following treatment
with ROT with a 32% reduction, as compared with the CON group
(Fig. 2D). Conversely, treatment
with curcumin resulted in an increase in GSH levels, and a decrease
in the accumulation of ROS and MDA levels induced by ROT treatment.
These results suggest that ROT-induced oxidative stress may exert
significant damage on dopaminergic neurons in the SNpc, and
curcumin pretreatment may be partially neuroprotective against ROT
exposure.
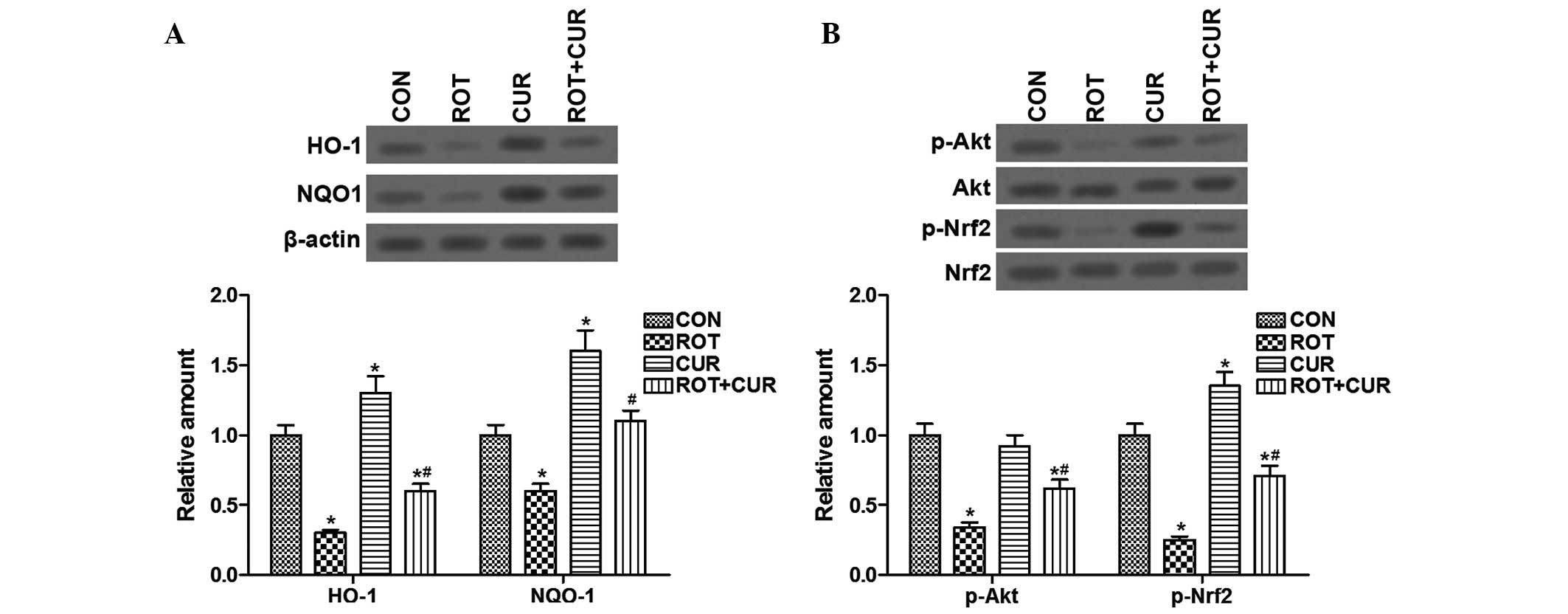
Curcumin activates the Akt/Nrf2-regulated
antioxidant system
The protective effects of Akt/Nrf2 signaling against
oxidative stress have previously been reported in PD (26); therefore, the present study aimed
to explore whether the Akt/Nrf2 antioxidant system was associated
with the protective effects of curcumin in ROT-treated rats.
Curcumin pretreatment prevented the ROT-induced reductions of HO-1
and NQO1 protein expression, and Akt/Nrf2 phosphorylation (Fig. 3A and B). Collectively, these data
suggest that curcumin may exhibit antioxidant and neuroprotective
effects via activation of the Akt/Nrf2 signaling pathway in the
SNpc dopaminergic neurons of ROT-induced parkinsonian rats.
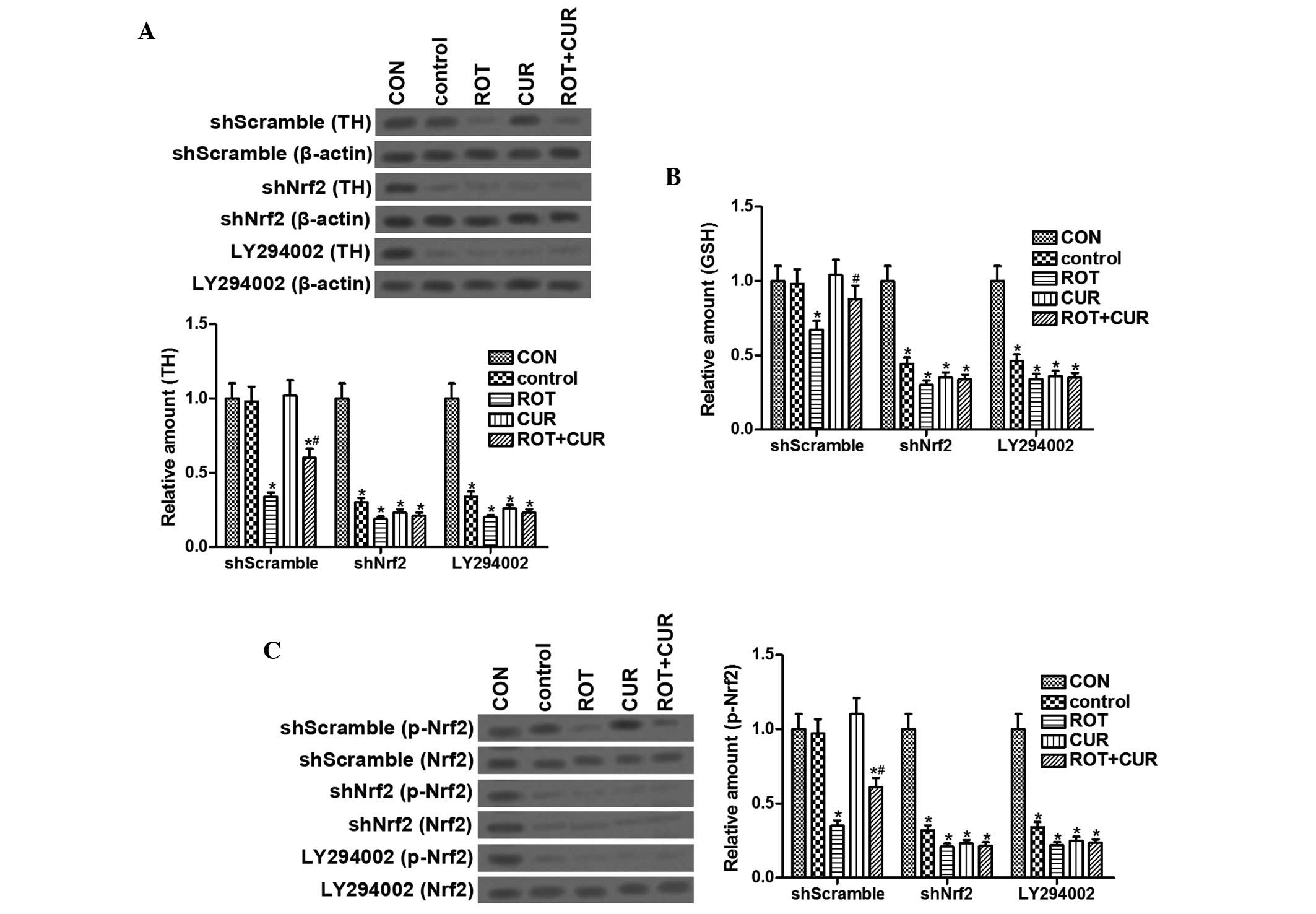
shNrf2 lentivirus and LY294002 suppress
curcumin-mediated neuroprotection
Transfection with a shNrf2 lentivirus or sublethal
chronic exposure to LY294002 prior to treatment with ROT/curcumin,
resulted in a failure of curcumin to prevent ROT-induced reductions
in TH and GSH expression (Fig. 4A and
B). In addition, Nrf2 phosphorylation was significantly reduced
in cells treated with shNrf2 lentivirus or LY294002 (Fig. 4C). These results indicate that the
neuro-protective effects of curcumin against ROT-induced SNpc
dopaminergic neuronal oxidative damage may be inhibited by
treatment with a shNrf2 lentivirus or LY294002.
Discussion
PD is regarded as a neurodegenerative disease, which
is characterized by the age-associated accumulation of oxidative
damage in human brain tissue. ROT, which is a commonly used natural
substance obtained from the roots of certain tropical plants
including Derris elliptica, is capable of causing
mitochondrial perturbation and oxidative damage (27). ROT is a lipophilic compound that
can freely cross cell membranes and gain access to cytoplasm and
mitochondria (28). Curcumin,
which is a major active polyphenol component of turmeric, exerts
various biological effects, and has been reported to possess
antioxidative activity against oxidative damage in cell and animal
models of PD (15,17). The aim of the present study was to
examine whether, and how, curcumin exerted protective effects
against ROT-induced dopaminergic neuronal oxidative damage in a rat
model of PD.
To better understand the therapeutic effects of
drugs specific to certain disease processes, animal models are
often used. Rats have been reported to develop motor and postural
deficits characteristic of PD following ROT minipump administration
for 35 days (29). Consistent with
previous research, the present study successfully established a
ROT-based rat model of PD, the chronic exposure of which was
administered by subcutaneous injection. In addition, the effects of
ROT could be significantly ameliorated with curcumin pretreatment.
The majority of studies regarding ROT have concentrated on the
neurotoxicity and behavioral deficits of neurodegenerative
diseases, such as PD (8,30–32).
For instance, ROT has been shown to result in motor impairment
(32), behavioral deficits
(31) and loss of TH
immunoreactivity in the striatum or SNpc (8). Furthermore, curcumin has previously
exhibited potential therapeutic protection by alleviating
neurotoxic behavioral changes in mice (22), and notably exhibited a protective
role in a 6-hydroxydopamine rat model of PD, where it suppressed
the degeneration of nigral dopaminergic neurons (9). In the present study, parkinsonian
symptoms were assessed using various behavioral tests, including
rotational behavior, catalepsy and open field tests; the results
indicated that curcumin pretreatment was able to partly alleviate
PD symptoms in the rats, and upregulate the expression levels of
TH, which were disordered by ROT.
A previous study demonstrated that the levels of
endogenous oxidative markers (ROS and MDA), redox status (GSH), and
response of antioxidant enzymes were associated with the state of
oxidative stress in rat brain tissues (33). ROT has been shown to induce
oxidative stress by stimulating the production of ROS (34) and MDA (35), and by reducing GSH levels in
vitro and in vivo (30,36).
In addition, curcumin is able to cross the blood-brain barrier and
has been reported to exert neuroprotective effects in neurological
disorders (10), which could
enhance the release of GSH (17),
and decrease the levels of MDA (17) and ROS (18), in order to protect against
oxidative damage in vitro and in vivo (16). The results of the present study
demonstrated that curcumin was able to reverse ROT-induced effects,
resulting in an elevation in GSH generation, and a reduction in the
levels of ROS and MDA.
It is well documented that Nrf2 is a basic leucine
zipper transcription factor that protects various tissues and cells
from ROS via the antioxidant response element (ARE)-mediated
induction of diverse antioxidant proteins and phase II detoxifying
enzymes, including HO-1 and NQO1 (21,37).
GSH depletion results in oxidative stress and TH activity loss,
which are considered symbolic events in neurodegeneration,
including in the progression of PD (17,34).
Curcumin may protect cortical neurons and reduce oxidative stress
levels, and it has been hypothesized that these neuroprotective
effects are regulated by the Akt/Nrf2 antioxidant system (38,39).
In addition, curcumin has been shown to affect the gene expression
of phase II enzymes (40),
inducing the antioxidant defense mechanism by activating the Nrf2
pathway and upregulating the expression levels of Nrf2/ARE
pathway-related genes HO-1 and NQO1 (38,41).
The present study indicated that the beneficial effects of curcumin
were due to changes in antioxidant enzyme activities, including
HO-1 and NQO1, via the activation of PI3K/Akt and transcription of
Nrf2. In addition, continuous infusion with shNrf2 lentivirus or
pharmacological inhibition with LY294002 suppressed Nrf2
phosphorylation and suppressed the expression levels of TH and GSH
in the SNpc, indicating that the Nrf2-associated cytoprotective and
antioxidative effects of curcumin are dependent on the PI3K/Akt
pathway.
In conclusion, the present study established a rat
model of PD using ROT, and examined the behavioral performance,
endogenous markers of oxidative stress, the activities of
antioxidant enzymes, and the activation of the related signaling
pathways in the SNpc following curcumin pretreatment. The results
indicated that the protective effects of curcumin against
ROT-induced dopaminergic neuronal oxidative damage were mediated by
activation of the Akt/Nrf2 signaling pathway. These results
demonstrated that curcumin exerts potential neuroprotective
therapeutic effects, and may be considered a novel therapeutic
agent for the treatment of PD.
Abbreviations:
|
PD
|
Parkinson's disease
|
|
SNpc
|
substantia nigra pars compacta
|
|
Nrf2
|
nuclear factor erythroid 2-related
factor 2
|
|
PI3K
|
phosphoinositide 3-kinase
|
|
NQO1
|
NAD(P)H:quinone oxidoreductase 1
|
|
LPO
|
lipid peroxidation
|
|
MDA
|
malondialdehyde
|
|
ROS
|
reactive oxygen species
|
|
HO-1
|
heme oxygenase-1
|
|
ROT
|
rotenone
|
|
TH
|
tyrosine hydroxylase
|
|
GSH
|
glutathione
|
|
shRNA
|
short hairpin RNA
|
References
|
1
|
Nutt JG and Wooten GF: Clinical practice.
Diagnosis and initial management of Parkinson's disease. N Engl J
Med. 353:1021–1027. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Camilleri A and Vassallo N: The centrality
of mitochondria in the pathogenesis and treatment of parkinson's
disease. CNS Neurosci Ther. 20:591–602. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Mariani E, Polidori MC, Cherubini A and
Mecocci P: Oxidative stress in brain aging, neurodegenerative and
vascular diseases: An overview. J Chromatogr B Analyt Technol
Biomed Life Sci. 827:65–75. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Tolleson CM and Fang JY: Advances in the
mechanisms of Parkinson's disease. Discov Med. 15:61–66.
2013.PubMed/NCBI
|
|
5
|
Sayre LM, Smith MA and Perry G: Chemistry
and biochemistry of oxidative stress in neurodegenerative disease.
Curr Med Chem. 8:721–738. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Perry TL and Yong VW: Idiopathic
Parkinson's disease, progressive supranuclear palsy and glutathione
metabolism in the substantia nigra of patients. Neurosci Lett.
67:269–274. 1986. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Bharath S, Hsu M, Kaur D, Rajagopalan S
and Andersen JK: Glutathione, iron and Parkinson's disease. Biochem
Pharmacol. 64:1037–1048. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Saravanan KS, Sindhu KM, Senthilkumar KS
and Mohanakumar KP: L-deprenyl protects against rotenone-induced,
oxidative stress-mediated dopaminergic neurodegeneration in rats.
Neurochem Int. 49:28–40. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Du XX, Xu HM, Jiang H, Song N, Wang J and
Xie JX: Curcumin protects nigral dopaminergic neurons by
iron-chelation in the 6-hydroxydopamine rat model of Parkinson's
disease. Neurosci Bull. 28:253–258. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Mythri RB and Bharath MM: Curcumin: A
potential neuroprotective agent in Parkinson's disease. Curr Pharm
Des. 18:91–99. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Dutta S, Padhye S, Priyadarsini KI and
Newton C: Antioxidant and antiproliferative activity of curcumin
semicarbazone. Bioorg Med Chem Lett. 15:2738–2744. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Biswas SK, McClure D, Jimenez LA, Megson
IL and Rahman I: Curcumin induces glutathione biosynthesis and
inhibits NF-kappaB activation and interleukin-8 release in alveolar
epithelial cells: Mechanism of free radical scavenging activity.
Antioxid Redox Signal. 7:32–41. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Singh S and Khar A: Biological effects of
curcumin and its role in cancer chemoprevention and therapy.
Anticancer Agents Med Chem. 6:259–270. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Daniel S, Limson JL, Dairam A, Watkins GM
and Daya S: Through metal binding, curcumin protects against lead-
and cadmium-induced lipid peroxidation in rat brain homogenates and
against lead-induced tissue damage in rat brain. J Inorg Biochem.
98:266–275. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Ataie A, Sabetkasaei M, Haghparast A,
Moghaddam AH and Kazeminejad B: Neuroprotective effects of the
polyphenolic antioxidant agent, Curcumin, against
homocysteine-induced cognitive impairment and oxidative stress in
the rat. Pharmacol Biochem Behav. 96:378–385. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Jagatha B, Mythri RB, Vali S and Bharath
MM: Curcumin treatment alleviates the effects of glutathione
depletion in vitro and in vivo: Therapeutic implications for
Parkinson's disease explained via in silico studies. Free Radic
Biol Med. 44:907–917. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Harish G, Venkateshappa C, Mythri RB,
Dubey SK, Mishra K, Singh N, Vali S and Bharath MM: Bioconjugates
of curcumin display improved protection against glutathione
depletion mediated oxidative stress in a dopaminergic neuronal cell
line: Implications for Parkinson's disease. Bioorg Med Chem.
18:2631–2638. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Chen J, Tang XQ, Zhi JL, Cui Y, Yu HM,
Tang EH, Sun SN, Feng JQ and Chen PX: Curcumin protects PC12 cells
against 1-methyl-4-phenylpyridinium ion-induced apoptosis by
bcl-2-mitochondria-ROS-iNOS pathway. Apoptosis. 11:943–953. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Cui QLSS: Curcumin antagonizes
rotenone-induced injury of PC12 Cells by antioxidant activity. Acta
Med Univ Sci Technol Huazhong. 39:37–46. 2010.
|
|
20
|
Tsou YH, Shih CT, Ching CH, Huang JY, Jen
CJ, Yu L, Kuo YM, Wu FS and Chuang JI: Treadmill exercise activates
Nrf2 antioxidant system to protect the nigrostriatal dopaminergic
neurons from MPP+ toxicity. Exp Neurol. 263:50–62. 2015. View Article : Google Scholar
|
|
21
|
Lee JM and Johnson JA: An important role
of Nrf2-ARE pathway in the cellular defense mechanism. J Biochem
Mol Biol. 37:139–143. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Kakkar V and Kaur IP: Evaluating potential
of curcumin loaded solid lipid nanoparticles in aluminium induced
behavioural, biochemical and histopathological alterations in mice
brain. Food Chem Toxicol. 49:2906–2913. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Mokrasch LC and Teschke EJ: Glutathione
content of cultured cells and rodent brain regions: A specific
fluorometric assay. Anal Biochem. 140:506–509. 1984. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Shinomol GK and Muralidhara: Prophylactic
neuroprotective property of Centella asiatica against
3-nitropropionic acid induced oxidative stress and mitochondrial
dysfunctions in brain regions of prepubertal mice. Neurotoxicology.
29:948–957. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Ohkawa H, Ohishi N and Yagi K: Assay for
lipid peroxides in animal tissues by thiobarbituric acid reaction.
Anal Biochem. 95:351–358. 1979. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Lim JH, Kim KM, Kim SW, Hwang O and Choi
HJ: Bromocriptine activates NQO1 via Nrf2-PI3K/Akt signaling: Novel
cytoprotective mechanism against oxidative damage. Pharmacol Res.
57:325–331. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Lin TK, Cheng CH, Chen SD, Liou CW, Huang
CR and Chuang YC: Mitochondrial dysfunction and oxidative stress
promote apoptotic cell death in the striatum via cytochrome
c/caspase-3 signaling cascade following chronic rotenone
intoxication in rats. Int J Mol Sci. 13:8722–8739. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Coulom H and Birman S: Chronic exposure to
rotenone models sporadic Parkinson's disease in Drosophila
melanogaster. J Neurosci. 24:10993–10998. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Betarbet R, Sherer TB, MacKenzie G,
Garcia-Osuna M, Panov AV and Greenamyre JT: Chronic systemic
pesticide exposure reproduces features of Parkinson's disease. Nat
Neurosci. 3:1301–1306. 2000. View
Article : Google Scholar : PubMed/NCBI
|
|
30
|
Madathil SK, Karuppagounder SS and
Mohanakumar KP: Sodium salicylate protects against rotenone-induced
parkinsonism in rats. Synapse. 67:502–514. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Santiago RM, Barbieiro J, Lima MM,
Dombrowski PA, Andreatini R and Vital MA: Depressive-like behaviors
alterations induced by intranigral MPTP, 6-OHDA, LPS and rotenone
models of Parkinson's disease are predominantly associated with
serotonin and dopamine. Prog Neuropsychopharmacol Biol Psychiatry.
34:1104–1114. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Swarnkar S, Singh S, Mathur R, Patro IK
and Nath C: A study to correlate rotenone induced biochemical
changes and cerebral damage in brain areas with neuromuscular
coordination in rats. Toxicology. 272:17–22. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Shinomol GK and Muralidhara: Bacopa
monnieri modulates endogenous cytoplasmic and mitochondrial
oxidative markers in prepubertal mice brain. Phytomedicine.
18:317–326. 2011. View Article : Google Scholar
|
|
34
|
Madathil KS, Karuppagounder SS, Haobam R,
Varghese M, Rajamma U and Mohanakumar KP: Nitric oxide synthase
inhibitors protect against rotenone-induced, oxidative stress
mediated parkinsonism in rats. Neurochem Int. 62:674–683. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Liu CB, Wang R, Pan HB, Ding QF and Lu FB:
Effect of lycopene on oxidative stress and behavioral deficits in
rotenone induced model of Parkinson's disease. Zhongguo Ying Yong
Sheng Li Xue Za Zhi. 29:380–384. 2013.In Chinese. PubMed/NCBI
|
|
36
|
Zhang Q, Zhang J, Jiang C, Qin J, Ke K and
Ding F: Involvement of ERK1/2 pathway in neuroprotective effects of
pyrroloquinoline quinine against rotenone-induced SH-SY5Y cell
injury. Neuroscience. 270:183–191. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Nguyen T, Sherratt PJ and Pickett CB:
Regulatory mechanisms controlling gene expression mediated by the
antioxidant response element. Annu Rev Pharmacol Toxicol.
43:233–260. 2003. View Article : Google Scholar
|
|
38
|
Wu J, Li Q, Wang X, Yu S, Li L, Wu X, Chen
Y, Zhao J and Zhao Y: Neuroprotection by curcumin in ischemic brain
injury involves the Akt/Nrf2 pathway. PLoS One. 8:e598432013.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Qin XY, Cheng Y and Yu LC: Potential
protection of curcumin against intracellular amyloid beta-induced
toxicity in cultured rat prefrontal cortical neurons. Neurosci
Lett. 480:21–24. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Dickinson DA, Iles KE, Zhang H, Blank V
and Forman HJ: Curcumin alters EpRE and AP-1 binding complexes and
elevates glutamate-cysteine ligase gene expression. FASEB J.
17:473–475. 2003.PubMed/NCBI
|
|
41
|
Gao S, Duan X, Wang X, Dong D, Liu D, Li
X, Sun G and Li B: Curcumin attenuates arsenic-induced hepatic
injuries and oxidative stress in experimental mice through
activation of Nrf2 pathway, promotion of arsenic methylation and
urinary excretion. Food Chem Toxicol. 59:739–747. 2013. View Article : Google Scholar : PubMed/NCBI
|