Introduction
Pseudomonas aeruginosa (PA), a Gram-negative
opportunistic bacterium, is a leading cause of hospital-acquired
pneumonia (1) and is responsible
for suppurative bacterial keratitis (2) and acute burn wound infection
(3). Clinically, antibiotics
including fluoroquinolones (4),
aminoglycosides (5) and
third-generation cephalexin (6)
are used to eradicate the bacteria. However, with increasing
difficulties in achieving satisfactory efficacy with current
antibiotic regimens, efforts have focused on investigating host
defense mechanisms against bacterial infections.
The host inflammatory response is a self-protective
strategy to control bacterial burden, and is characterized by the
accumulation of inflammatory cells and cytokines (7). During infection, inflammatory cells,
such as macrophages, are rapidly recruited to kill the invading
pathogens (7). Macrophages also
produce various pro-inflammatory cytokines including interleukin 6
(IL-6), IL-1β, tumor necrosis factor α (TNF-α) and macrophage
inflammatory protein 2 (MIP-2), which function to enhance the
antibacterial immune response (2,7,8).
These inflammatory mediators promote bacterial clearance, however,
if uncontrolled may result in tissue damage (2). Therefore, a combined strategy to
control both the bacterial burden and inflammation is critical for
the clinical treatment of PA infection.
Wingless-type MMTV integration site family, member
3A (Wnt3a) is a multifunctional molecule that has been reported to
have the ability to modulate host inflammation (9–11).
As a cysteine-rich secretory glycoprotein, Wnt3a secretes and binds
to its respective dimeric cell surface receptors, frizzled proteins
and low-density lipoprotein receptor-related protein 5/6,
inhibiting the activity of the 'destruction complex' and the
subsequent degradation of β-catenin (12). Stabilized β-catenin translocates
into the nucleus, binds to transcription factors such as T cell
factor and lymphoid-enhancing factor and promotes the expression of
target genes that are involved in proliferation, and carcinogenesis
(12).
Although studies have indicated a potential
association of Wnts with several inflammatory diseases, including
diabetic retinopathy (9),
inflammatory bowel disease (10)
and rheumatoid arthritis (11),
whether Wnt3a serves a positive or negative role in the
inflammatory response remains controversial. It has been reported
that Wnt3a increased the secretion of IL-1β and IL-6 in fibrotic
alveolar epithelia via activating Wnt/β-catenin signaling (13). However, other studies have
demonstrated that Wnt family members (such as Wnt2 and Wnt11)
suppress bacterial-induced IL-8 secretion in epithelial cells
(14,15). To date, little is known regarding
the activity of Wnt3a in PA-induced inflammatory responses.
Programmed cell death (apoptosis) is an important
strategy to control host inflammatory responses. However, the role
of Wnt3a in regulating apoptosis remains elusive. Zimmerman et
al (16) demonstrated that
Wnt3a enhanced apoptosis in melanoma cells, while Gui et al
(17) indicated that Wnt3a
stimulated β-cell proliferation and inhibited cytokine-induced
β-cell apoptosis. Thus, the role of Wnt3a in regulating PA-induced
macrophage apoptosis remains to be determined.
Liu et al (14) reported that Wnt family members have
different functions in pathogen clearance. Wnt2 had no effect on
inhibiting bacterial invasion (14), whereas Wnt11 significantly reduced
Salmonella invasion in epithelial cells (15). In addition, Wnt/β-catenin signaling
(canonical Wnt signaling) has been implicated in modulating the
generation of the major components in the oxygen-dependent
microbicidal system, such as reactive oxygen species (ROS)
(18) and nitric oxide (NO)
(19). Furthermore, studies have
demonstrated that antimicrobial peptides (the major components of
the oxygen-independent microbicidal system) such as
cathelicidin-related antimicrobial peptide (CRAMP) (20,21)
and β-defensin (BD) (22) are
required in the host resistance to PA infection, by controlling the
host inflammatory response and bacterial burden. However, to date,
little is known regarding the activity of Wnt3a in
macrophage-mediated bacterial killing ability.
In the present study, Wnt3a was demonstrated to
serve an anti-inflammatory role by suppressing the expression of
pro-inflammatory cytokines and promoting macrophage apoptosis.
Furthermore, bacterial clearance studies indicated that Wnt3a
enhanced intracellular bacterial killing via inducing the
expression of CRAMP and BD1.
Materials and methods
Cell culture
Conditioned media containing Wnt3a (Wnt3a-CM) was
prepared from mouse L cells stably expressing Wnt3a
(ATCC® CRL-2647™; American Type Culture Centre,
Manassas, VA, USA), and control conditioned media (Ctl-CM; American
Type Culture Centre) was obtained from parental L cells
(ATCC® CRL-2648™; American Type Culture Centre).
RAW264.7 murine macrophage-like cells (ATCC #TIB-71; American Type
Culture Centre) were cultured as described previously (23). Briefly, cells were cultured in
Dulbecco's modified Eagle's media (DMEM) supplemented with 10%
(v/v) fetal bovine serum (FBS), 1% penicillin-streptomycin and 1%
L-glutamine (all purchased from Invitrogen; Thermo Fisher
Scientific, Inc., Waltham, MA, USA) at 37°C in 5% CO2.
Cells were starved in DMEM containing 2% FBS overnight and then
exposed to Wnt3a-CM or Ctl-CM.
Western blotting
RAW264.7 cells were treated with various
concentrations (10, 20, 30, 40 or 50%) of Wnt3a-CM for 6 h or with
50% Wnt3a-CM for 6, 12, 24, 36 or 48 h. Subsequently, the cells
were washed three times with ice-cold PBS and then treated with
lysis buffer containing 1 mM phenylmethylsulfonyl fluoride, 1%
(v/v) protease inhibitor cocktail and 1 mM dithiothreitol (all
purchased from Sigma-Aldrich). Subsequently, the cell lysate was
obtained by centrifugation at 12,000 × g for 15 min at 4°C, after
which the protein concentration of the supernatant was determined
using the Quick Start Bradford protein assay (Bio-Rad Laboratories,
Inc., Hercules, CA, USA). 30 µg of each sample was separated
by 10% sodium dodecyl sulfate polyacrylamide gel electrophoresis
(Beijing Dingguo Changsheng Biotechnology Co., Ltd., Beijing,
China), and then transferred to nitrocellulose membranes (Pall
Corporation, Port Washington, NY, USA). The membranes were blocked
with 5% non-fat milk in PBS containing 0.1% Tween-20 and incubated
with the primary rabbit monoclonal antibodies against β-catenin
(1:1,000; cat. no. 8480), cleaved-caspase 3 (1:1,000; cat. no.
9664), cleaved-poly ADP ribose polymerase (PARP; 1:1,000; cat. no.
5625) and β-actin (1:5,000; cat. no. 3700; all Cell Signaling
Technology, Inc.) at 4°C overnight, followed by incubation with
IRDye 800CW donkey anti-rabbit IgG (H+L) antibodies (1:5,000;
DkxRb-003-D800NHSX; LI-COR, Inc., Lincoln, NE, USA) for 1 h at room
temperature. Subsequently, the bands were detected using an Odyssey
CLx Infrared Imaging System (LI-COR, Inc.), with an excitation
wavelength of 780 nm and an emission wavelength of 820 nm. Band
intensities were quantified using the Image Studio software,
version 4.0 (LI-COR, Inc.), according to the manufacturer's
protocol.
Immunofluorescent staining
A proportion of RAW264.7 cells were infected with PA
at a multiplicity of infection (MOI) of 1.0 for 12 h, after which
RAW264.7 cells (4×105 cells/ml) with or without PA
infection were exposed to 50% Wnt3a-CM or Ctl-CM for 6 h and then
seeded onto sterile glass cover slips, cultured overnight and fixed
in 4% formaldehyde (Sigma-Aldrich, St. Louis, MO, USA) at 4°C.
Subsequently, the sections were blocked with 5% bull serum albumin
in phosphate-buffered saline (PBS) at room temperature for 1 h,
after which they were incubated with rabbit anti-mouse β-catenin
(1:200; cat. no. 8480; Cell Signaling Technology, Inc., Danvers,
MA, USA) at 4°C overnight. The sections were then incubated with
Cy3-conjugated goat anti-rabbit IgG (1:1,000; cat. no. AP187C, EMD
Millipore, Billerica, MA, USA) at room temperature for 1 h.
Finally, sections were incubated with 4,6-diamino-2-phenyl indole
(DAPI, 1:10,000; Sigma-Aldrich) at room temperature for 5 min for
nuclear staining. The controls were similarly treated, although the
primary antibody was replaced with isotype matched IgG. The
sections were visualized using the Olympus BX41 microscope (Olympus
Corporation, Tokyo, Japan).
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Total RNA was isolated from the RAW264.7 cells using
TRIzol® reagent (Invitrogen; Thermo Fisher Scientific,
Inc.), and the purity of the RNA was determined by measuring the
ratio of absorbance at 260 and 280 nm using the NanoDrop™ 2000
Spectrophotometer (Thermo Fisher Scientific, Inc.). Total RNA (1
µg) was reverse transcribed into cDNA using the
High-Capacity RNA-to-cDNA kit (Applied Biosystems; Thermo Fisher
Scientific, Inc.), and then amplified using the SYBR Green Master
Mix (Bio-Rad Laboratories, Inc.), according to the manufacturer's
protocols. The primer sequences for IL-6, IL-1β, MIP-2, TNF-α,
mCRAMP, mBD1 and β-actin are listed in Table I. RT-qPCR reactions were performed
using the CFX96 Real-Time PCR System (Bio-Rad Laboratories, Inc.)
with the following cycling conditions: 95°C for 30 sec, followed by
40 cycles of 95°C for 5 sec and 60°C for 10 sec. Relative mRNA
expression levels were calculated following normalization to
β-actin using the 2−ΔΔCq method and the Prism 5.0
software (GraphPad Software, Inc., La Jolla, CA, USA).
 | Table INucleotide sequence of the primers
used in PCR amplification. |
Table I
Nucleotide sequence of the primers
used in PCR amplification.
| Gene | Primer sequence
(5′-3′) |
|---|
| β-actin | F:
GATTACTGCTCTGGCTCCTAGC |
| R:
GACTCATCGTACTCCTGCTTGC |
| IL-6 | F:
CACAAGTCCGGAGAGGAGAC |
| R:
CAGAATTGCCATTGCACAAC |
| IL-1β | F:
CGCAGCAGCACATCAACAAGAGC |
| R:
TGTCCTCATCCTGGAAGGTCCACG |
| MIP-2 | F:
TGTCAATGCCTGAAGACCCTGCC |
| R:
AACTTTTTGACCGCCCTTGAGAGTGG |
| TNF-α | F:
CACAGAAAGCATGATCCGCGAC |
| R:
TGCCACAAGCAGGAATGAGAAGAG |
| mCRAMP | F:
AGCTACAGGGATGCTGTGCT |
| R:
TCACTCGGAACCTCAGACT |
| mBD1 | F:
GGCATTCTCACAAGTCTTGGACGAAG |
| R:
AGCTCTTACAACAGTTGGGCTTATCTGG |
Flow cytometric analysis of
apoptosis
A proportion of RAW264.7 cells were infected with PA
at a MOI of 0.5 for 24 h. The apoptosis of RAW264.7 cells with or
without PA infection was assessed by flow cytometry using a
fluorescein isothiocyanate (FITC)-annexin V apoptosis detection kit
(BD Biosciences, Inc., Franklin Lakes, NJ, USA) according to the
manufacturer's protocol. Briefly, the cells (5×105) were
pooled, washed and resuspended in 500 µl binding buffer,
followed by the addition of 5 µl annexin V-FITC and 5
µl propidium iodide (PI). Subsequently, the cells were
incubated at room temperature in the dark for 15 min, and analyzed
by flow cytometry (Beckman Coulter EPICS XL/MCL; Beckman Coulter,
Brea, CA, USA), with an excitation wavelength of 488 nm and an
emission wavelength of 530 for FITC (green) and 575–610 nm for PI
(orange). Viable cells were unstained by annexin V or PI, early
apoptotic cells were stained by annexin V only, and late apoptotic
cells were stained by annexin V and PI.
Intracellular bacterial killing
assay
RAW264.7 cells were cultured in a 6-well plate and
then exposed to 50% Wnt3a-CM or Ctl-CM for 6 h, followed by PA
challenge at a MOI of 25 for 1 and 2 h, respectively. Subsequently,
the cells in one well were treated with 300 µg/ml gentamicin
(Sigma-Aldrich) for 30 min in order to kill the extracellular
bacteria, and then washed with PBS three times and lysed with 0.1%
Triton-X. Cells in the duplicate well were incubated for a further
1 h and then lysed in the same way. Serial 10-fold dilutions of
each sample were plated on Pseudomonas isolation agar (BD
Biosciences, Inc.) in triplicate and incubated overnight at 37°C.
For intracellular bacterial killing, the efficiency was calculated
using the following equation: Intracellular bacterial killing =
[colony forming units (CFU) (1 h) − CFU (2 h)]/CFU (1 h) ×
100%.
Phagocytosis assay
Phagocytosis was assayed by flow cytometry as
described previously (24).
Briefly, PA was incubated with Filmtracer Green Biofilm (FTGB,
1:50; Invitrogen; Thermo Fisher Scientific, Inc.) at room
temperature for 30 min in the dark, and then rinsed gently with
sterilized water. Subsequently, RAW264.7 cells were exposed to 50%
Wnt3a-CM or Ctl-CM for 6 h, and then infected with FTGB-stained PA
at a MOI of 25. Following 1 h of incubation, the cells were washed
three times with ice-cold PBS to remove nonadherent bacteria.
Extracellular fluorescence was quenched by the addition of 0.1%
trypan blue (Sigma-Aldrich) in PBS for 15 min. Cells were collected
and analyzed using a Beckman Coulter EPICS XL/MCL instrument, with
an excitation wavelength of 488 nm and an emission wavelength of
514 nm.
ROS measurement by flow cytometry
RAW264.7 cells were exposed to Wnt3a-CM or Ctl-CM
for 6 h. Following PA challenge, the cells were incubated with a
ROS-sensitive probe, 2′,7′-dichlorofluorecscin diacetate
(H2DCFDA; Invitrogen; Thermo Fisher Scientific, Inc.),
at a final concentration of 10 mm. Subsequently, the cells were
collected and analyzed using a Beckman Coulter EPICS XL/MCL flow
cytometer, with an excitation wavelength of 488 nm and an emission
wavelength of 530 nm. ROS levels were determined by the
fluorescence of dichlorofluorecscin (DCF), the deacetylated and
oxidized product of H2DCFDA.
Griess assay
RAW264.7 cells were exposed to Wnt3a-CM or Ctl-CM,
and the supernatant from each sample was collected at 12 and 24 h
following PA challenge by centrifugation at 1,000 × g for 10 min at
4°C. NO levels were determined by measuring the levels of the
stable end product, nitrite, using the Griess reagent assay
(Sigma-Aldrich), according to the manufacturer's protocol. Briefly,
50 µl culture supernatant mixed with 50 µl
sulfanilamide solution was added to wells in duplicate, and
incubated for 10 min at room temperature in the dark. Subsequently,
50 µl N-1-napthylethylenediamine dihydrochloride solution
was added to the wells and incubated for 10 min at room temperature
in the dark. Absorbance at 540 nm was measured using the iMarkTM
Microplate Absorbance Reader (Bio-Rad Laboratories, Inc.).
Statistical analysis
Data are presented as the mean ± standard error of
the mean of three independent experiments. Statistical analyses
were conducted using the Prism 5.0 software (GraphPad Software,
Inc., La Jolla, CA, USA). An unpaired, two-tailed Student's t-test
was used to analyze differences between the Wnt3a-CM and Ctl-CM
treated groups at the same time point following PA infection.
P<0.05 was considered to indicate a statistically significant
difference.
Results
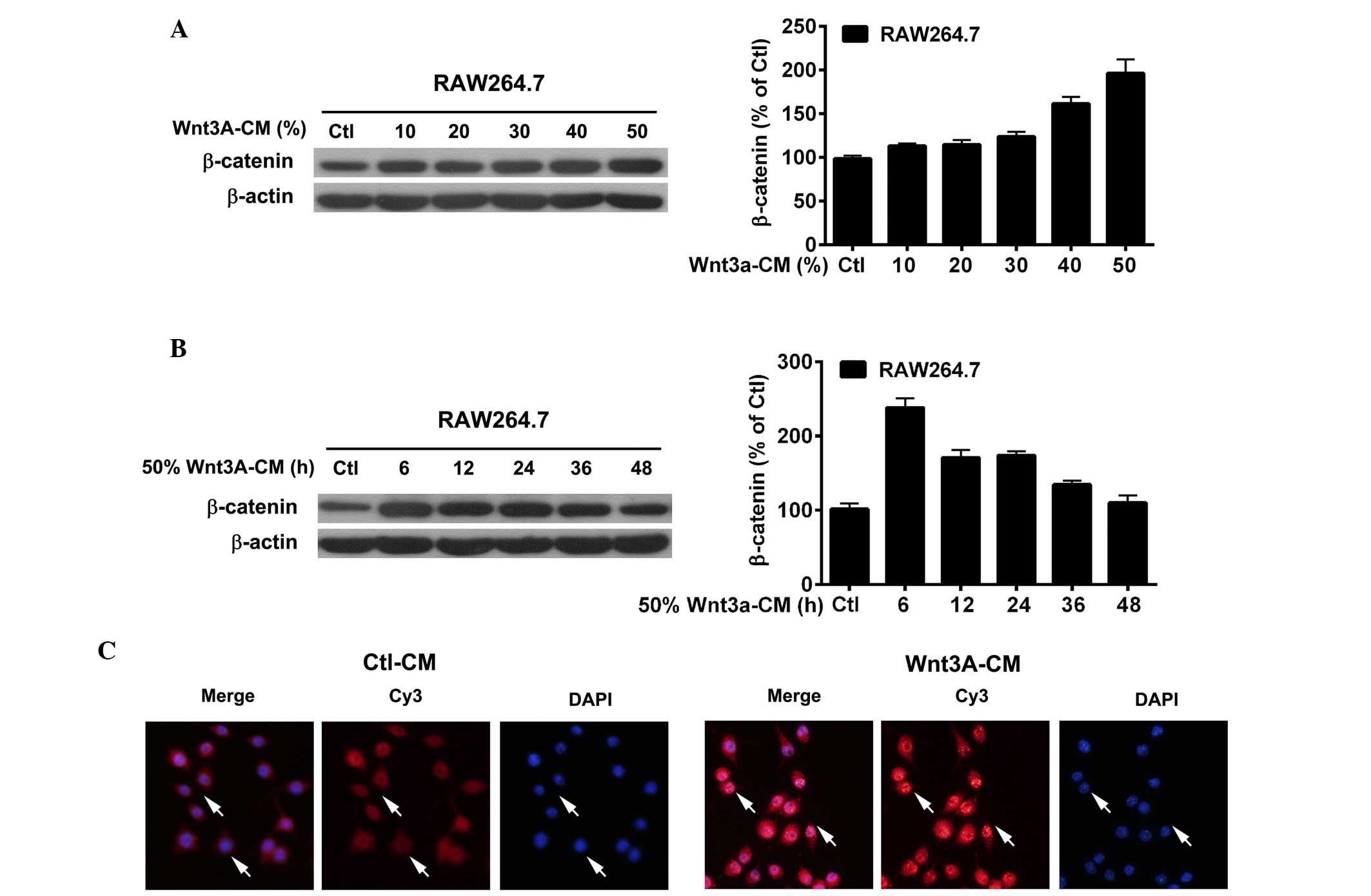
Efficacy of Wnt3a-CM exposure in
macrophages
The efficacy of Wnt3a-CM exposure was demonstrated
by the expression and distribution of β-catenin, a central molecule
of the canonical Wnt signaling pathway (12). RAW264.7 cells were exposed to
Wnt3a-CM at a range of concentrations (Fig. 1A) and to 50% Wnt3a-CM for various
durations (Fig. 1B). Western
blotting data showed that the protein levels of β-catenin were
markedly upregulated in RAW264.7 cells following exposure to 50%
Wnt3a-CM (Fig. 1A) and for 6 h
(Fig. 1B). Furthermore,
immunofluorescence data (Fig. 1C)
indicated that β-catenin (red staining, Cy3-labeled) was
upregulated in the cytoplasm and was translocated to the nucleus
(DAPI nuclear staining) following exposure to 50% Wnt3a-CM for 6
h.
Wnt3a inhibits pro-inflammatory cytokine
secretion in macrophages
To explore the inflammatory regulation of Wnt3a, the
expression of pro-inflammatory cytokines was measured by RT-qPCR in
Wnt3a-CM and Ctl-CM treated RAW264.7 cells before and after PA
infection. The RT-qPCR data showed that Wnt3a suppressed the mRNA
levels of IL-6 (P<0.05; Fig.
2A), IL-1β (P<0.01; Fig.
2B), MIP-2 (P<0.01; Fig.
2C) and TNF-α (P<0.05; Fig.
2D) following PA infection. In addition, no alteration in
pro-inflammatory cytokine expression was detected in Wnt3a-CM and
Ctl-CM treated groups prior to PA challenge.
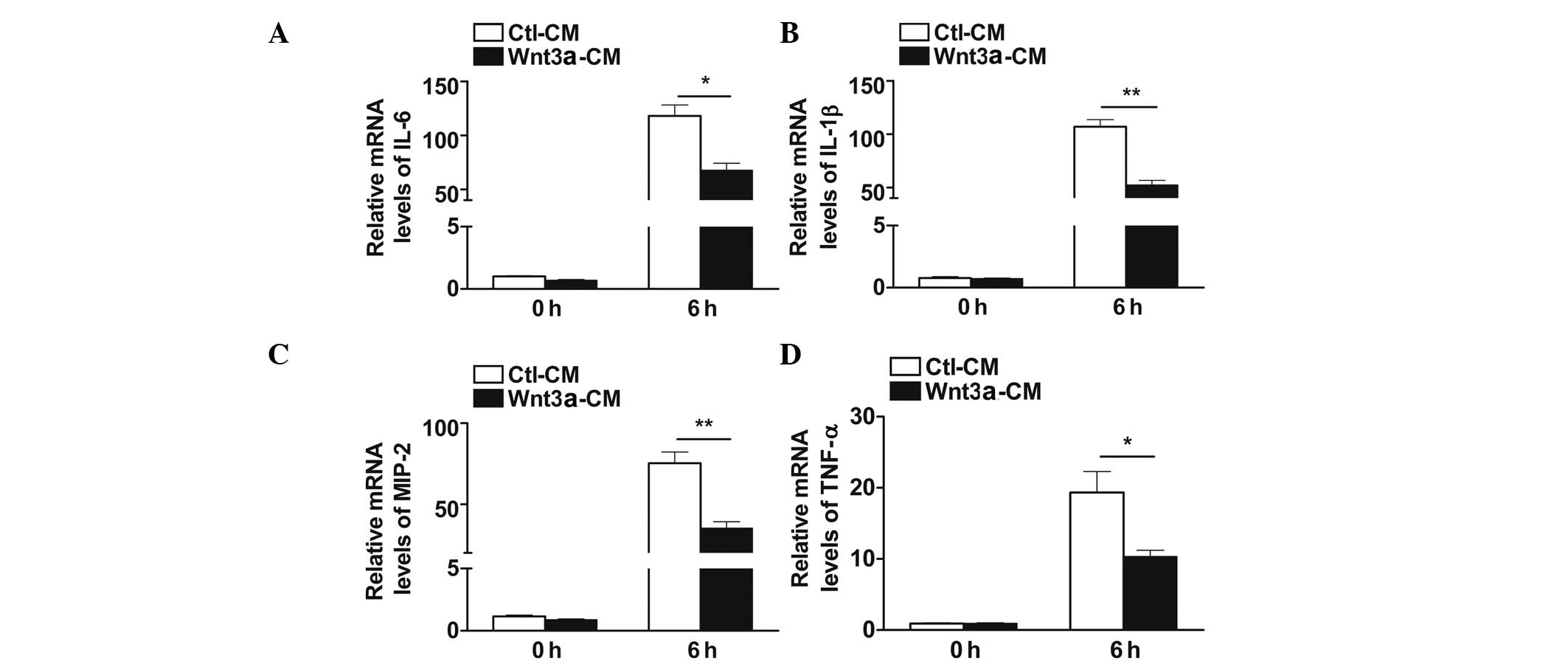 | Figure 2Wnt3a inhibited the expression of
pro-inflammatory cytokines in macrophages following infection. The
mRNA levels of (A) IL-6, (B) IL-1β, (C) MIP-2 and (D) TNF-α were
examined by reverse transcription-quantitative polymerase chain
reaction in 50% Wnt3a-CM and Ctl-CM treated RAW264.7 cells with (6
h) and without (0 h) PA infection. Data are presented as the mean ±
standard error and represent three individual experiments.
*P<0.05; **P<0.01. IL, interleukin;
MIP-2, macrophage inflammatory protein 2; TNF-α, tumor necrosis
factor-α; Wnt3a, wingless-type MMTV integration site family, member
3A; Wnt3a-CM, Wnt3a conditioned media; Ctl-CM, control conditioned
media; PA, Pseudomonas aeruginosa. |
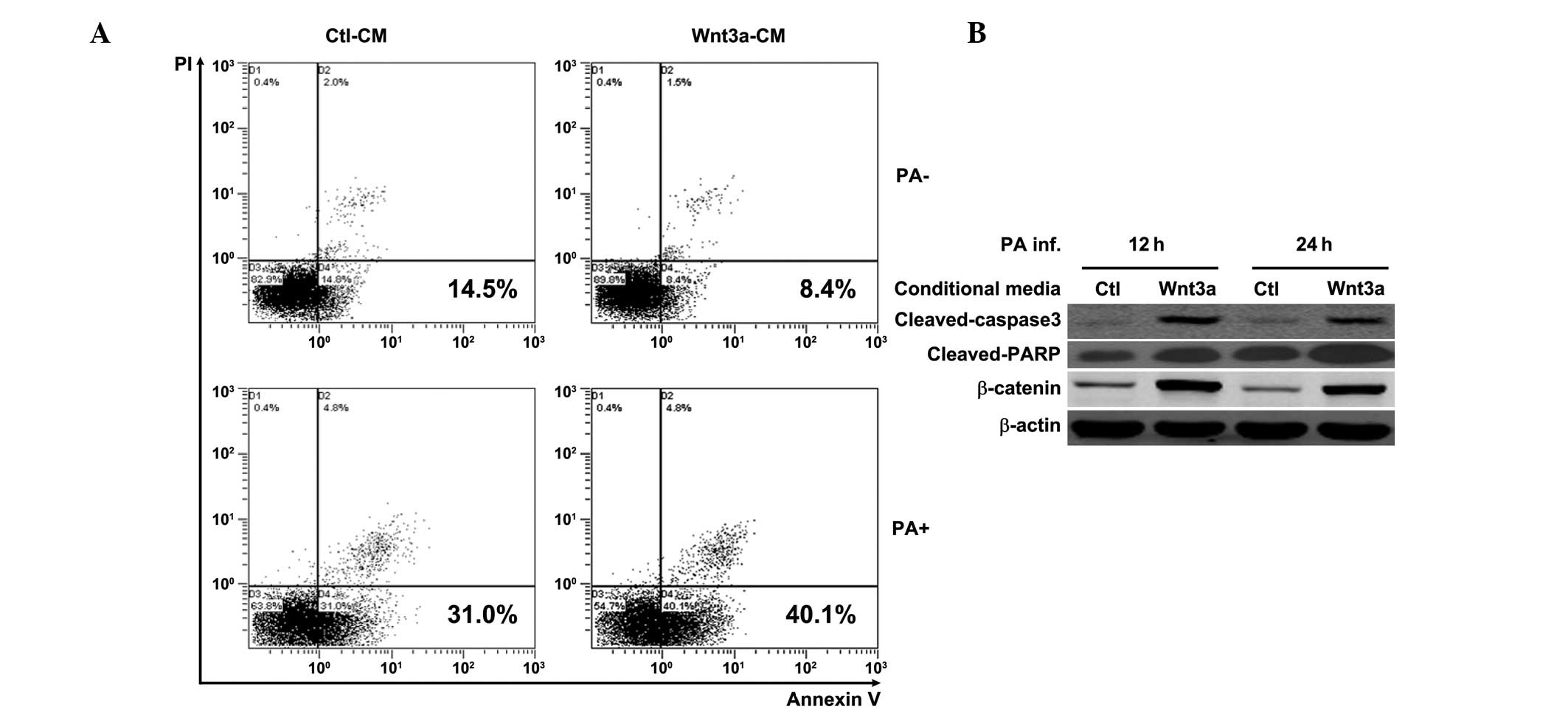
Wnt3a promotes macrophage apoptosis
following PA challenge
The role of Wnt3a in modulating macrophage apoptosis
was further examined by annexin-V/PI double staining and flow
cytometry. The flow cytometry data showed that Wnt3a increased the
number of annexin V positive cells following PA challenge (PA+;
Fig. 3A). However, Wnt3a slightly
reduced cell apoptosis in the absence of PA infection (PA-;
Fig. 3A), which was consistent
with the role of Wnt3a in cell proliferation. Additional evidence
for the occurrence of apoptosis was obtained by western blot
analysis, which measured cleaved caspase 3 and cleaved-PARP, two
hallmarks of apoptosis. The western blotting data showed that Wnt3a
induced caspase 3 and PARP cleavage in macrophages at 12 and 24 h
following PA stimulation (Fig.
3B), compared with the Ctl-CM treated cells. Furthermore, the
efficacy of Wnt3a-CM treatment in RAW264.7 cells (Fig. 3B) was confirmed by western
blotting, as indicated by the upregulation of β-catenin.
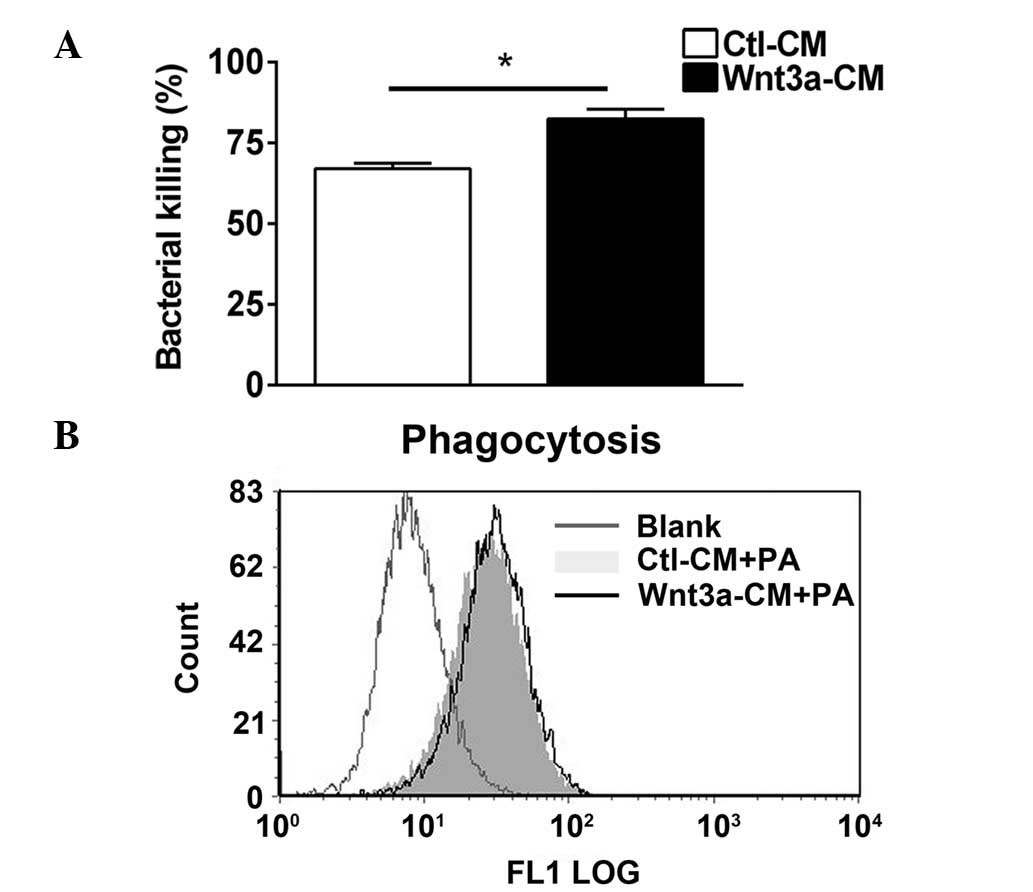
Wnt3a enhances intracellular bacterial
killing however, not phagocytosis following PA infection
It has been previously reported that Wnt11 prevents
bacterial invasiveness in intestinal epithelial cells (15), therefore, the present study
investigated whether Wnt3a modulates the process of bacterial
clearance. Bacterial clearance was assessed by using a bacterial
killing assay based on plate counts (Fig. 4A) and a phagocytosis assay using
flow cytometry (Fig. 4B).
Bacterial killing data indicated that Wnt3a promoted bacterial
killing following PA challenge in RAW264.7 cells (P<0.05;
Fig. 4A), and the phagocytosis
assay indicated alterations in the uptake of PA in Wnt3a-CM and
Ctl-CM treated RAW264.7 cells (Fig.
4B). These data demonstrated that Wnt3a enhances
macrophage-mediated intracellular killing of PA, however, was not
involved in the process of phagocytosis.
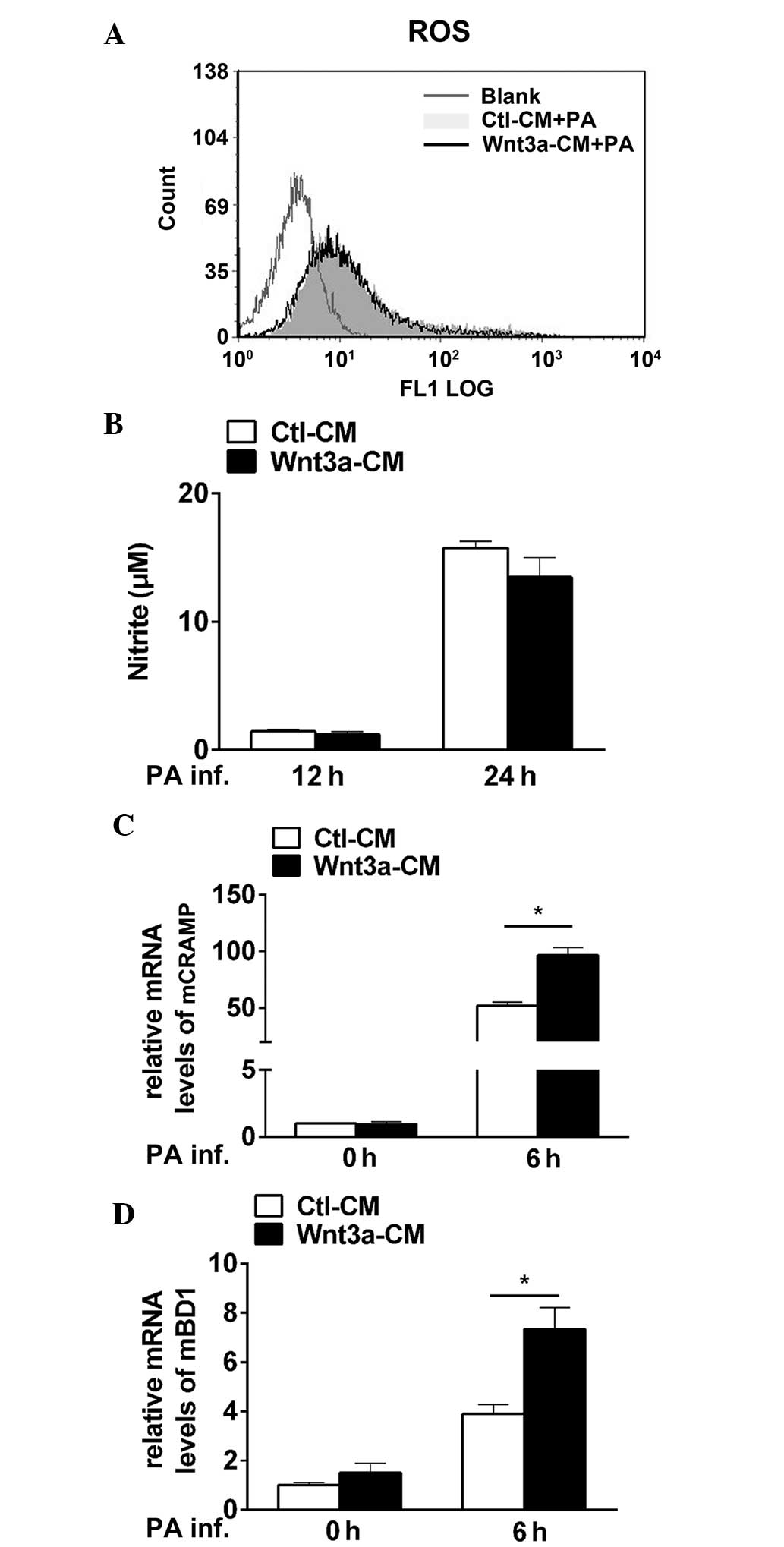
Wnt3a promotes macrophage-mediated
bacterial killing by elevating CRAMP and BD1 levels, however, not
ROS or NO production
To explore the microbicidal mechanisms involved
during PA infection, oxygen-dependent (ROS and NO) and
oxygen-independent microbicidal systems (antimicrobial peptides)
were measured in Wnt3a-CM and Ctl-CM treated RAW264.7 cells
following PA challenge. No alterations between the two groups were
observed in PA-induced ROS production, as indicated by the
percentage of DCF-positive cells (Fig.
5A), and NO levels, as indicated by the measurement of the
stable end product nitrate (Fig.
5B). However, the results showed that the mRNA levels of CRAMP
(P<0.05; Fig. 5C) and BD1
(P<0.05; Fig. 5D) were
significantly upregulated in Wnt3a-CM and Ctl-CM treated RAW264.7
cells following PA infection. These results demonstrated that Wnt3a
enhances macrophage-mediated intracellular killing via the
antimicrobial peptides CRAMP and BD1, but not by ROS and NO
production.
Discussion
As an upstream mediator in the Wnt/β-catenin
pathway, Wnt3a serves a critical role in modulating host
inflammation (9–11). However, the function of Wnt3a in
regulating PA-induced host inflammation and the associated
microbicidal mechanisms remain unclear. The present study
demonstrated that Wnt3a suppressed inflammation by reducing the
expression of pro-inflammatory cytokines and promoting apoptosis in
macrophages. In addition, it was observed that Wnt3a promoted
macrophage-mediated intracellular bacterial killing by elevating
the levels of the antimicrobial peptides CRAMP and BD1. Taken
together, these results shed light on the regulation of Wnt3a in
bacterial infectious diseases.
Previous studies have demonstrated that Wnt3a serves
a pro-inflammatory role, by enhancing the expression of IL-1β and
IL-6 in fibrotic alveolar epithelia (13). However, other studies have reported
that Wnt family members (Wnt2 and Wnt11) inhibited
Salmonella-induced IL-8 expression in intestinal epithelia
cells (14,15). Furthermore, Neumann et al
(25) reported that Wnt3a reduced
TNF release in Mycobacterium tuberculosis-infected
macrophages (25). These studies
suggest that Wnt3a may exert anti-inflammatory effects in
bacterial-induced inflammatory responses. However, the inflammatory
cytokine profiles orchestrated by bacteria stimulation may vary,
depending on the cell type and species of bacteria. The current
study indicated that Wnt3a served an anti-inflammatory role by
reducing the expression of pro-inflammatory cytokines including
IL-1β, IL-6, MIP-2 and TNF-α in PA-challenged macrophages. The role
of Wnt3a in regulating pro-inflammatory cytokines is consistent
with a previous study that demonstrated that β-catenin inhibited
the expression of pro-inflammatory cytokines in PA-infected
macrophages (23), which suggests
that Wnt3a may regulate PA-induced inflammatory cytokines via its
downstream mediator β-catenin. A potential mechanism underlying the
inflammatory regulation of Wnt3a may be that Wnt binding stabilizes
the transcription factor β-catenin, which in turn enters the
nucleus and negatively regulates nuclear factor-κB, leading to the
suppression of the expression of pro-inflammatory cytokines
(26,27).
Apoptosis is another strategy employed by the host
to control excessive inflammatory responses. Previous studies have
demonstrated that Wnt3a promotes the apoptosis of melanoma cells
(16) and Bacillus
Calmette-Guerin-infected RAW264.7 cells (28); however, it exhibited anti-apoptotic
activity in β-cells (17) and
increased the proliferation of heart valve interstitial cells
(29). The present study
demonstrated that Wnt3a enhanced PA-induced apoptosis in
macrophages, which is indispensable for the control of robust
inflammatory responses, contributing to the role of Wnt3a in
bacterial-induced anti-inflammatory effects.
Inflammatory mediators are critical for the host
defense, and lead to clearance of the infectious pathogen (7). However, in the present study, Wnt3a
was observed to promote bacterial killing, which appears to be in
conflict with its anti-inflammatory activity. Therefore, additional
bactericidal mechanisms that may be involved in the process of
macrophage-mediated bacterial killing triggered by Wnt3a were
investigated. Phagocytic clearance by macrophages is key to the
endogenous control of PA (1).
However, the present study indicated that Wnt3a had no influence on
phagocytosis. It has been reported that Wnt3/β-catenin signaling
promotes ROS (18) and NO
(19) production, with these being
major components in the oxygen-dependent microbicidal system.
However, the present study observed no alteration in the generation
of NO or ROS in Wnt3a-CM and Ctl-CM treated macrophages following
PA infection. In addition to the oxygen-dependent pathway (e.g.,
NO/ROS), immune cells possess an oxygen-independent pathway (e.g.,
antimicrobial peptide, lysozymes or lysosomal hydrolytic enzymes)
to fight against invading microorganisms (30,31).
It has been previously reported that CRAMP-deficient B6 mice are
more susceptible to PA ocular infection than wild-type mice
(20). Hazlett and Wu (22) demonstrated that defensins, as a
major family of antimicrobial peptides, exert widely antimicrobial
activity (22). The present study
observed that Wnt3a promoted CRAMP and BD1 expression, indicating
that the antimicrobial activity of Wnt3a may be associated with the
induction of CRAMP and BD1.
Inflammation and bacterial virulence contribute to
infectious diseases; however, treating bacterial infection with
antibiotics does not commonly prevent pathology caused by an
excessive immune response. The present study demonstrated that
Wnt3a suppressed inflammation and enhanced bacterial killing, thus
suggesting that Wnt3a may be considered a therapeutic strategy for
the treatment of PA infection.
Acknowledgments
The present study was supported by grants from the
National Natural Science Foundation of China (grant no. 81401645)
and Guangdong Medical Science Foundation (grant no. B2014447).
References
|
1
|
Lovewell RR, Patankar YR and Berwin B:
Mechanisms of phagocytosis and host clearance of Pseudomonas
aeruginosa. Am J Physiol Lung Cell Mol Physiol. 306:L591–L603.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Hazlett LD: Corneal response to
Pseudomonas aeruginosa infection. Prog Retin Eye Res. 23:1–30.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Bielecki P, Glik J, Kawecki M and Martins
dos Santos VA: Towards understanding Pseudomonas aeruginosa burn
wound infections by profiling gene expression. Biotechnol Lett.
30:777–790. 2008. View Article : Google Scholar
|
|
4
|
Kowalski RP, Romanowski EG, Mah FS, Shanks
RM and Gordon YJ: Topical levofloxacin 1.5% overcomes in vitro
resistance in rabbit keratitis models. Acta Ophthalmol.
88:e120–e125. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
McCormick C, Caballero A, Tang A, Balzli
C, Song J and O'Callaghan R: Effectiveness of a new tobramycin
(0.3%) and dexamethasone (0.05%) formulation in the treatment of
experimental Pseudomonas keratitis. Curr Med Res Opin.
24:1569–1575. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Mohammadpour M, Mohajernezhadfard Z,
Khodabande A and Vahedi P: Antibiotic susceptibility patterns of
Pseudomonas corneal ulcers in contact lens wearers. Middle East Afr
J Ophthalmol. 18:228–231. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Hazlett LD: Pathogenic mechanisms of P.
aeruginosa keratitis: A review of the role of T cells, Langerhans
cells, PMN and cytokines. DNA Cell Biol. 21:383–390. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Kernacki KA, Goebel DJ, Poosch MS and
Hazlett LD: Early cytokine and chemokine gene expression during
Pseudomonas aeruginosa corneal infection in mice. Infect Immun.
66:376–379. 1998.PubMed/NCBI
|
|
9
|
Zhou T, Hu Y, Chen Y, Zhou KK, Zhang B,
Gao G and Ma JX: The pathogenic role of the canonical Wnt pathway
in age-related macular degeneration. Invest Ophthalmol Vis Sci.
51:4371–4379. 2010. View Article : Google Scholar :
|
|
10
|
You J, Nguyen AV, Albers CG, Lin F and
Holcombe RF: Wnt pathway-related gene expression in inflammatory
bowel disease. Dig Dis Sci. 53:1013–1019. 2008. View Article : Google Scholar
|
|
11
|
Miao CG, Yang YY, He X, Li XF, Huang C,
Huang Y, Zhang L, Lv XW, Jin Y and Li J: Wnt signaling pathway in
rheumatoid arthritis, with special emphasis on the different roles
in synovial inflammation and bone remodeling. Cell Signal.
25:2069–2078. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Wodarz A and Nusse R: Mechanisms of Wnt
signaling in development. Annu Rev Cell Dev Biol. 14:59–88. 1998.
View Article : Google Scholar
|
|
13
|
Aumiller V, Balsara N, Wilhelm J, Günther
A and Königshoff M: WNT/β-catenin signaling induces IL-1β
expression by alveolar epithelial cells in pulmonary fibrosis. Am J
Respir Cell Mol Biol. 49:96–104. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Liu X, Lu R, Wu S, Zhang YG, Xia Y, Sartor
RB and Sun J: Wnt2 inhibits enteric bacterial-induced inflammation
in intestinal epithelial cells. Inflamm Bowel Dis. 18:418–429.
2012. View Article : Google Scholar :
|
|
15
|
Liu X, Wu S, Xia Y, Li XE, Xia Y, Zhou ZD
and Sun J: Wingless homolog Wnt11 suppresses bacterial invasion and
inflammation in intestinal epithelial cells. Am J Physiol
Gastrointest Liver Physiol. 301:G992–G1003. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Zimmerman ZF, Kulikauskas RM, Bomsztyk K,
Moon RT and Chien AJ: Activation of Wnt/β-catenin signaling
increases apoptosis in melanoma cells treated with trail. PLoS One.
8:e695932013. View Article : Google Scholar
|
|
17
|
Gui S, Yuan G, Wang L, Zhou L, Xue Y, Yu
Y, Zhang J, Zhang M, Yang Y and Wang DW: Wnt3a regulates
proliferation, apoptosis and function of pancreatic NIT-1 beta
cells via activation of IRS2/PI3K signaling. J Cell Biochem.
114:1488–1497. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Kim JS, Yeo S, Shin DG, Bae YS, Lee JJ,
Chin BR, Lee CH and Baek SH: Glycogen synthase kinase 3beta and
beta-catenin pathway is involved in toll-like receptor 4-mediated
NADPH oxidase 1 expression in macrophages. FEBS J. 277:2830–2837.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Du Q, Park KS, Guo Z, He P, Nagashima M,
Shao L, Sahai R, Geller DA and Hussain SP: Regulation of human
nitric oxide synthase 2 expression by Wnt beta-catenin signaling.
Cancer Res. 66:7024–7031. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Huang LC, Reins RY, Gallo RL and McDermott
AM: Cathelicidin-deficient (Cnlp −/−) mice show increased
susceptibility to Pseudomonas aeruginosa keratitis. Invest
Ophthalmol Vis Sci. 48:4498–4508. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Kumar A, Hazlett LD and Yu FS: Flagellin
suppresses the inflammatory response and enhances bacterial
clearance in a murine model of Pseudomonas aeruginosa keratitis.
Infect Immun. 76:89–96. 2008. View Article : Google Scholar :
|
|
22
|
Hazlett L and Wu M: Defensins in innate
immunity. Cell Tissue Res. 343:175–188. 2011. View Article : Google Scholar
|
|
23
|
Chen K, Yin L, Nie X, Deng Q, Wu Y, Zhu M,
Li D, Li M, Wu M and Huang X: β-Catenin promotes host resistance
against Pseudomonas aeruginosa keratitis. J Infect. 67:584–594.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Mariencheck WI, Savov J, Dong Q, Tino MJ
and Wright JR: Surfactant protein A enhances alveolar macrophage
phagocytosis of a live, mucoid strain of P. aeruginosa. Am J
Physiol. 277:L777–L786. 1999.PubMed/NCBI
|
|
25
|
Neumann J, Schaale K, Farhat K, Endermann
T, Ulmer AJ, Ehlers S and Reiling N: Frizzled1 is a marker of
inflammatory macrophages and its ligand Wnt3a is involved in
reprogramming Mycobacterium tuberculosis-infected macrophages.
FASEB J. 24:4599–4612. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Sun J, Hobert ME, Rao AS, Neish AS and
Madara JL: Bacterial activation of beta-catenin signaling in human
epithelia. Am J Physiol Gastrointest Liver Physiol. 287:G220–G227.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Sun J, Hobert ME, Duan Y, Rao AS, He TC,
Chang EB and Madara JL: Crosstalk between NF-kappaB and
beta-catenin pathways in bacterial-colonized intestinal epithelial
cells. Am J Physiol Gastrointest Liver Physiol. 289:G129–G137.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Wu X, Deng G, Hao X, Li Y, Zeng J, Ma C,
He Y, Liu X and Wang Y: A caspase-dependent pathway is involved in
Wnt/β-catenin signaling promoted apoptosis in Bacillus
Calmette-Guerin infected RAW264.7 macrophages. Int J Mol Sci.
15:5045–5062. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Xu S and Gotlieb AI: Wnt3a/β-catenin
increases proliferation in heart valve interstitial cells.
Cardiovasc Pathol. 22:156–166. 2013. View Article : Google Scholar
|
|
30
|
Thomas EL, Lehrer RI and Rest RF: Human
neutrophil antimicrobial activity. Rev Infect Dis. 10(Suppl 2):
S450–S456. 1988. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Wiesner J and Vilcinskas A: Antimicrobial
peptides: The ancient arm of the human immune system. Virulence.
1:440–464. 2010. View Article : Google Scholar : PubMed/NCBI
|