Introduction
Chemokines are a group of peptides of small
molecular weight (8–11 kDa). They induce the chemotaxis of various
leukocyte subtypes and regulate leukocyte trafficking to sites of
inflammation (1). The major
function of chemokines is the recruitment of leukocytes to sites of
inflammation (1). CXC chemokine
ligand 12 (CXCL12), or stromal cell derived factor (SDF-1), is a
68-amino-acid long CXC chemokine, which was originally identified
as a growth factor for mouse pre-B cells (2). CXCL12 is constitutively expressed by
various types of cell and tissue, and exhibits chemoattractive
activity for endothelial cells, epithelial cells, monocytes, bone
marrow neutrophils, dendritic cells and, in particularly, T cells
and their co-stimulators (3).
Following the connecting of the chemokine with CXC chemokine
receptor 4 (CXCR4) and its receptor, CXCL12 can exert series of
functions, including the transendothelial migration of inflammatory
cells and mobilization of leukocytes (4). Previous studies have shown that
certain types of immunomodulatory cytokines can affect the
expression levels of CXCR4 on T cells. It was reported that
interleukin (IL)10 and IL4 significantly upregulate or downregulate
the expression of CXCR4 on CD4+ T lymphocytes (5). In addition, the interferon (IFN)γ
cytokine induces prompt downregulation in the mRNA expression of
CXCR4 and attenuates endothelial cell migration towards CXCL12
(6). Increases in the expression
levels of CXCR4 or CXCL12 have been reported in several
inflammatory diseases, including rheumatoid arthritis, systemic
lupus erythematosus and inflammatory bowel disease (IBD) (4,7–9).
AMD3100 has been considered as a specific inhibitor of CXCR4
(10). In previous reports,
AMD3100 was found to specifically inhibit CXCL12-mediated
repercussions and have beneficial effects in several animal models
of immune diseases, including diabetes (11), asthma (12) and IBD (8). Thus, this antagonist may provide an
effective way of inhibiting thyroid damage in iodine-induced
autoimmune thyroiditis in NOD.H-2h4 mice, which express
I-A k in their NOD genetic background. These mice spontaneously
develop autoimmune thyroiditis (SAT) and produce anti-mouse
thyroglobulin (MTg) autoantibodies, and are a prototype murine
model of Hashimoto's thyroiditis (HT) in humans (13).
Autoimmune thyroid diseases (AITDs), including HT
and Graves' disease are chronic organ-specific autoimmune diseases.
They are characterized by the destruction of thyroid follicles by
infiltrating inflammatory cells and mononuclear cell infiltration
of the thyroid gland (14). It has
been shown that T helper (Th)1 cytokines are commonly prevalent in
HT, as well as in experimental autoimmune thyroiditis (EAT), by
analyzing of the expression of cytokines, including IL1, IL2, IL6,
IL10, IFNγ and TNFα, due to the infiltration of T cells and
macrophages (15–18). Furthermore, the thyroid follicular
cells produce several types of cytokines themselves (19). In previous years, chemokines have
been considered to be important in endocrine autoimmune disease,
and studies have demonstrated CC and CXC chemokine overexpression
in HT and EAT (20–22). It is currently accepted that
IFNγ-inducible chemokines (CXCL9, CXCL10, CXCL11, CXCL12, CXCL13
and CCL22) may be the most important mediating chemokines for
formation of the germinal center (GC) (23). Thyrocytes are the primary source of
CXCL12 in the thyroid, whereas CXCR4 is expressed by T and B cells
(13). Thyrocytes can also produce
CCL2, CXCL9 and CXCL10. Iodine may induce thyrocyte necrosis,
stimulating resident macrophages to produce IL1 and TNFα, which may
in turn induce CXCL12 synthesis by the adjacent thyrocytes
(24).
To the best of our knowledge, no previous studies
have evaluated CXCL12 and CXCR4 in an iodine-induced autoimmune
thyroiditis NOD.H-2h4 mouse model. The aim of the
present study was to measure the levels of CXCL12 in
NOD.H-2h4 mice, and to investigate the potential effects
of the CXCL12 and CXCR4 antagonist, AMD3100, on the inflammation
barrier in mouse models of SAT.
Materials and methods
Animals
Male NOD.H-2h4 mice (6-week-old; 18–21 g)
were purchased from Jackson Laboratory (Bar Harbor, ME, USA). Male
BALB/c mice (6-week-old; 18–21 g), were purchased from Vital River
Laboratories (Beijing, China). The animals were housed under
specific pathogen-free conditions in a controlled temperature and
humidity environment, with day-night light cycles in the animal
facility of China Medical University (Shenyanh, China). The present
study was performed in strict accordance with the recommendations
in the Guide for the Care and Use of Laboratory Animals of the
National Institutes of Health (25). The experimental protocol was
approved by the ethics committee of the Ethics of Animal
Experiments of China Medical University (Shenyang, China). All
surgery was performed under anesthesia using 10% chloral hydrate
(Melone Pharmaceutical Co., Ltd., Dailan, China), and all efforts
were made to minimize suffering.
Experimental design
For the induction of autoimmune thyroiditis, the
mice were administered with 0.05% (500 mg/l) sodium iodide (NaI) in
the drinking water (1,000 times higher than normal iodine intake)
during the study period. The CXCR4 antagonist, AMD3100, was
obtained from Sigma-Aldrich (St. Louis, MO, USA). For treatment, 10
mg/kg of AMD3100 dissolved in 200 µl phosphate-buffered
saline (PBS), or 200 µl PBS alone were administered
intraperitoneally three times a week during the study period.
Normal control mice received regular drinking water throughout the
experiment.
Experimental groups
Experiment one
A total of 12 NOD.H-2h4 mice and 12
BALB⁄c mice were randomly selected and divided into four groups:
NOD.H-2h4 mice provided with regular drinking water
(NOD.H-2h4-CON; n=6); NOD.H-2h4 mice provided
with 0.05% NaI drinking water (NOD.H-2h4-HI; n=6);
BALB⁄c mice provided with regular drinking water (BALB⁄c-CON; n=6);
and BALB⁄c mice provided with 0.05% NaI drinking water (BALB⁄c-HI;
n=6). Animals were anesthetized and sacrificed using an overdose of
10% chloral hydrate at week 8 of the experiment.
Experiment two
A total of 36 NOD.H-2h4 mice were
randomly-selected and divided into two groups: NOD.H-2h4
mice with regular drinking water (CON) group and
NOD.H-2h4 mice with 0.05% NaI drinking water (HI) group,
with four mice per group. A total of 4 mice per group were
anesthetized and sacrificed at weeks 0, 2, 4, 8, and 16 of the
experiment.
Experiment three
A total of 29 NOD.H-2h4 mice were
randomly selected and divided into three groups: NOD.H-2
h4 mice with regular drinking water (CON; n=10),
NOD.H-2h4 mice with 0.05% NaI drinking water (AIT; n=10) and
NOD.H-2h4 mice with 0.05% NaI drinking water and AMD3100
treatment (AMD3100; n=9). The animals were anesthetized and
sacrificed at week 8 of the experiment.
Assessment of autoimmune
thyroiditis
At the experimental end points, the mice were
weighed and anesthetized via intraperitoneal injection of 10%
chloral hydrate (26). Thyroid
tissues were removed and then washed with cold normal saline, dried
on a pad of filter paper and weighed on an electronic balance
(BS210S; Sartorius, Göttingen Germany). From each mouse, one
thyroid lobe was fixed in 10% para-formaldehyde (Beijing Solarbio
Science & Technology Co., Ltd., Beijing, China) for at least 24
h and embedded in paraffin (Huayong, Shanghai, China). Sections of
5-µm thickness were prepared and stained with hematoxylin
and eosin (HE; Beyotime Institute of Biotechnology, Hangzhou,
China). Histological changes in the thyroid tissue were observed
under light microscopy (BX51/BX52; Olympus Corporation, Tokyo,
Japan). The extent of lymphocytic infiltration was assessed, as
previously described (26,27). Briefly, HE-stained thyroid sections
were graded on the following scale, according to the approximate
area of lymphocytic infiltration: 0=normal; 1+=1–10%; 2+=10–30%;
3+=30–50%; 4+ >50%. The thyroiditis scores were expressed as the
mean of at least three non-contiguous sections from each thyroid
gland.
Serum thyroglobulin antibody (TgAb)
measurements using ELISA
Blood was collected from the orbital vein, incubated
at room temperature for at least 2 h and the serum was separated by
centrifugation at 1,006 × g for 20 min at room temperature and
stored at −80°C until further analysis. MTg was prepared as
previously described (26).
Thyroid gland tissues were homogenized in PBS and centrifuged at
1,000 × h at 4°C for 10 min. MTg was obtained from the supernatant
using by a salting out as previously described (28), and then purified by repeated gel
filtration using Sephadex G-20 (Pharmacia; Pfizer, Inc., New York,
NY, USA). Samples were stored at −7°C until analysis. The TgAb
titers were assessed in duplicate by indirect ELISA, using serum
from the individual mice, as described previously (26,28).
Briefly, sera were diluted to 1:100 with PBS and incubated at 37°C
for 1 h on 96-well EIA/RIA plates (Corning, St. Louis, MO, USA)
coated with 10 µg/ml mouse thyroglobulin [prepared from
frozen mouse thyroids as previously described (28)]. Peroxidase-labeled goat anti-mouse
immunoglobulin G (polyclonal; 1:250 dilution; A0168; Sigma-Aldrich)
was used as the secondary antibody. The color change of tetramethyl
benzidine was measured at 450 nm using a microtiter plate reader
(F200PRO; Tecan Group, Ltd., Zurich, Switzerland), with TgAb levels
expressed as optical density (OD) values.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR) analysis
Tissue was stored in RNAlater (Qiagen GmbH, Hilden,
Germany) at −80°C, and homogenized using Tissue Lyser II (Quiagen,
Hilden, Germany) at 20°C. Total RNA was extracted from the splenic
cells using TRIzol reagent (Invitrogen; Thermo Fisher Scientific,
Inc., Waltham, MA, USA), according to the manufacturer's protocol.
For RT, 1 µg total RNA was used to prepare cDNA (26). A reverse transcriptase kit
(PrimeScript RT reagent kit; Takara Biotechnology Co., Ltd.,
Dalian, China) and an ABI 9700 PCR meter (Applied Biosystems;
Thermo Fisher Scientific, Inc.) was used for cDNA synthesis (37°C
for 15 min, followed by 85°C for 5 sec). Transcripts were
quantified using a Roche LightCycler480 real-time PCR System
(version 1.5.0; Roche Diagnostics, Mannheim, Germany) using
SYBR® Premix Ex Taq TM II (Takara Biotechnology Co.,
Ltd.), according to the manufacturer's protocol. PCR parameters
were as follows: 95°C For 30 sec, followed by 40 cycles of 95°C for
5 sec, 60°C for 20 sec and 65°C for 15 sec. At the end of the PCR
cycles, a melting curve analysis was performed. Experiments were
performed in triplicate and data were quantified and analyzed using
LightCycler 480 (Roche Diagnostics) analysis software. The
reactions were performed using a total volume of 20 µl, in a
0.2-ml flat cap PCR tube (Axygen; Tewksbury, MA, USA). The primers
used are indicated in Table I.
 | Table ISequence of primers used for
amplification in reverse transcription-quantitative polymerase
chain reaction analysis. |
Table I
Sequence of primers used for
amplification in reverse transcription-quantitative polymerase
chain reaction analysis.
| Gene | Reference
sequence | Forward primer
(5′-3′) | Reverse primer
(5′-3′) |
|---|
| CXCL12 | NM_021704.3 |
GCTCTGCATCAGTGACGGTA |
ATCTGAAGGGCACAGTTTGG |
| IFNγ | NM_008337.3 |
CACGGCACAGTCATTGAAAG |
AATCTGGCTCTGCAGGATTT |
| IL10 | NM_010548.2 |
CCAAGCCTTATCGGAAATGA |
TTTTCACAGGGGAGAAATCG |
| β-actin | NM_007393.3 |
GGTCATCACTATTGGCAACG |
TCCATACCCAAGAAGGAAGG |
Flow cytometry
The spleens were harvested from the mice following
sacrifice at week 8 of the experiment, and pressed through a
200-gauge stainless steel mesh (Stainless Steel Nets Factory,
Shenyang, China) (26). The tissue
was suspended in 10 ml PBS and centrifuged at 111.8 × g for 10 min
at room temperature. The erythrocytes were resuspended in 3 ml
lysis solution (ammonium chloride; Beyotime Institute of
Biotechnology), incubated at room temperature for ~3 min, and
centrifuged at 111.8 × g for 10 min at room temperature. The cells
were then washed and immunostained with monoclonal fluorescein
isothiocyanate-conjugated rat anti-mouse CD4 (RM4-5; 553047),
monoclonal PerCP-Cy 5.5-conjugated rat anti-mouse CD19 (1D3;
551001), monoclonal PerCP-Cy 5.5-conjugated rat anti-mouse CD8
(53-6.7; 561109), monoclonal phycoerythrin-conjugated rat
anti-mouse IL10 (JES5-16E3; 554467), and monoclonal allophycocyanin
(APC)-conjugated rat anti-mouse IFNγ (XMG1.2; 562018). All
antibodies were purchased from BD Biosciences (San Jose, CA, USA)
and were diluted 1:100. All staining was performed according to
manufacturer protocols. For intracellular cytokine detection, the
cells were stimulated with a leukocyte activation cocktail in the
presence of GolgiPlug (BD Biosciences) for 6 h at 37°C prior to
staining with fluorophore-conjugated anti-IFNγ and anti-IL10 using
a Cytofix/Cyto perm plus kit (BD Biosciences) in accordance with
the manufacturer's instructions. The stained cells were then
analyzed using a FACSCalibur flow cytometer (BD Biosciences) and
FlowJo 7.6.1 software (Tree star Inc., Ashland, OR, USA).
Thyroid immunohistochemistry
The mouse thyroids were embedded in paraffin and
sectioned coronally with a microtome into 5-µm sections
(29). The sections were dewaxed
and rehydrated, and treated for endogenous peroxidase with 3%
methanol-hydrogen peroxide (Maixin, Fuzhou, China) for 10 min. All
sections were incubated with the mouse anti-CXCL12 primary
monoclonal antibody (dilution 1:50; MAB530; R&D Systems,
Minneapolis, MN, USA) at 4°C overnight and were then incubated for
40 min at room temperature. The tissue sections were then incubated
at room temperature with biotin-conjugated secondary antibodies
(Maixin) for 10 min, and in streptavidin-peroxidase complex
(Maixin) for 10 min. Subsequently, the sections were treated with a
solution of 3,3′-diaminobenzidine (DAB; Maixin) for 1–3 min,
depending upon the staining of the DAB reaction product observed
under a light microscope (BX51/BX52; Olympus Corporation). Finally,
the sections were counterstained with hematoxylin, dehydrated,
rinsed with distilled water, and mounted in neutral gum (China
National Medicines, Shanghai, China). All sections from each
thyroid were viewed at 400× magnification. The integral OD (IOD)
values, indicating the expression levels of the proteins were
measured using Image-Pro Plus 5.0 software (Media Cybernetics,
Inc., Silver Spring, MD, USA).
Statistical analysis
All results are expressed as the mean ± standard
error of the mean. One-way analysis of variance was performed to
determine whether there were statistically significant differences
among the groups, and post-test comparisons were performed using
the Bonferroni's test or Dunnett's T3 test using the SPSS 17.0
software package (SPSS, Inc., Chicago, IL, USA). Graphs were
analyzed using GraphPad Prism 5 software (GraphPad Software Inc.,
San Diego, CA, USA). P<0.05 was considered to indicate a
statistically significant difference.
Results
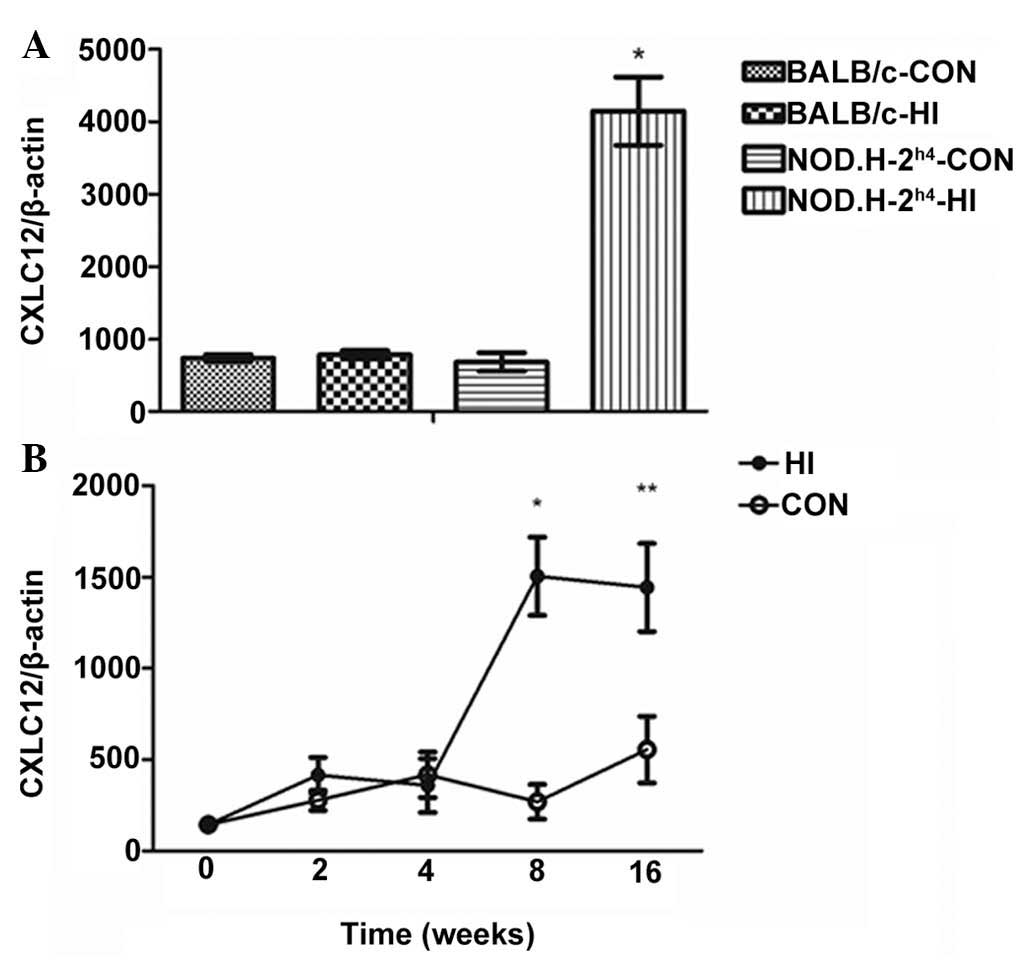
mRNA expression of CXCL12 is increased in
the thyroid of NOD.H-2 h4-HI mice
As shown in Fig.
1A, no difference in the CXCL12 chemokine was observed between
the thyroid tissues of the BAL/C-CON group and BAL/C-HI group.
However, the mRNA expression of CXCL12 in the thyroid was
significantly increased in the NOD.H-2h4-HI group,
compared with the NOD.H-2h4-CON group, BAL/C-CON group
and BAL/C-HI group.
Subsequently, dynamic changes in the mRNA expression
of CXCL12 in the thyroid of NOD.H-2h4 mice were examined
(Fig. 1B). No differences were
observed between the CON group and HI group at 0, 2 or 4 weeks;
however, significant differences were apparent at 8 and 16 weeks,
in which the mRNA expression levels of CXCL23 were significantly
higher, compared with those in the CON group.
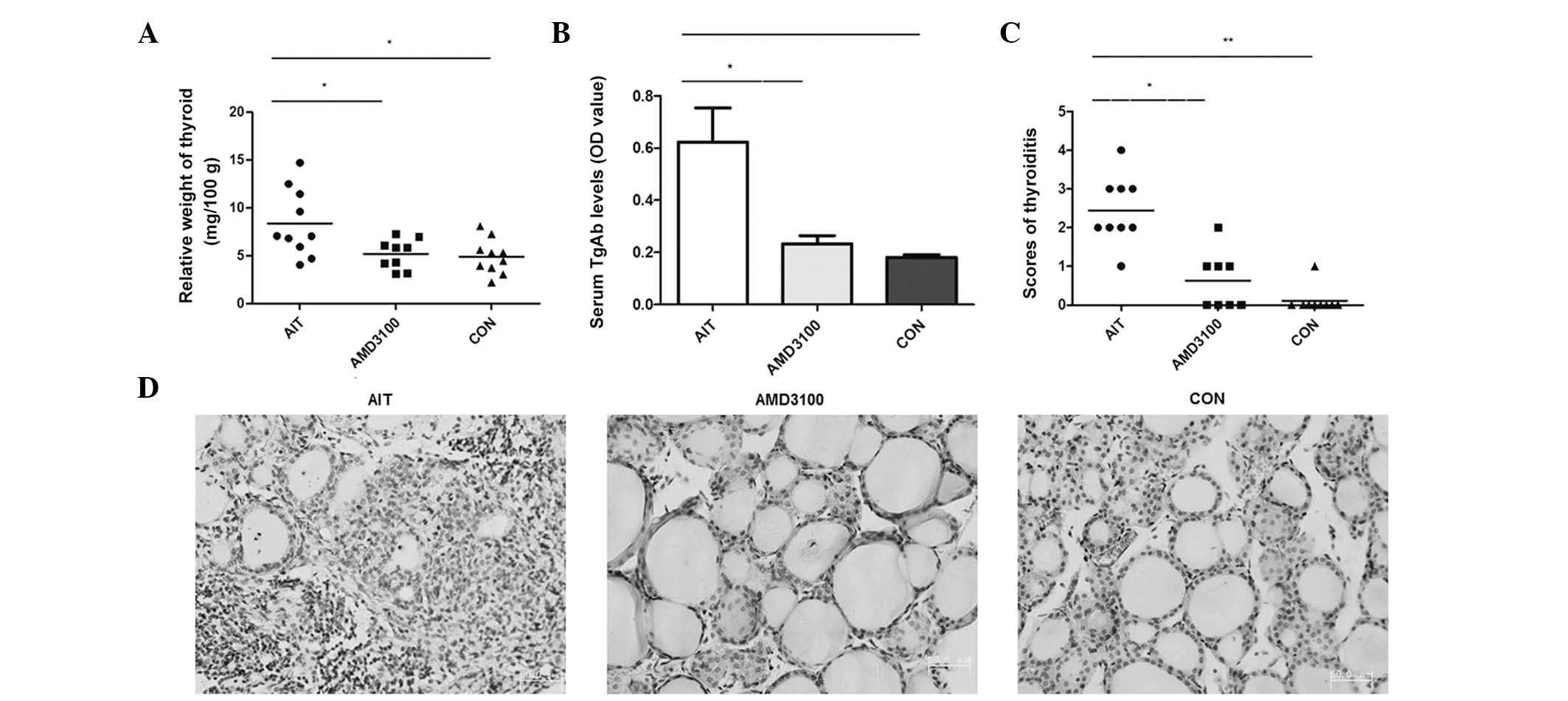
Relative thyroid tissue weight is reduced
in AMD3100-treated mice
As shown in Fig.
2A, the relative weights of the thyroids in the AIT group, in
which NOD.H-2h4 mice received NaI in their drinking
water for 8 weeks (8.86±1.12 mg/100 g), were increased
significantly, compared with those in the CON group (4.88±0.57
mg/100 g) and AMD3100 group (5.20±0.52 mg/100 g). The histological
changes of the enlarged thyroids included a lighter color and
tougher texture, compared with the normal thyroids. No significant
differences were identified between the CON group and AMD3100
group.
Serum TgAb titer is reduced in
AMD3100-treated mice
Serum TgAb titers were significantly increased in
the AIT group, compared with the CON group (0.625±0.13, vs.
0.18±0.009, respectively; P=0.022). By contrast, the AMD3100 group
had significantly reduced serum TgAb titers, compared with the AIT
group (0.232±0.03, vs. 0.625±0.13, respectively; P=0.043; Fig. 2B). No significant differences were
found between the CON group and AMD3100 group.
Lymphocytic infiltration is less severe
in the thyroid of AMD3100-treated mice
In the present study, lymphocytic infiltration was
observed in the thyroids in almost 100% of the mice in the AIT
group; and the scores of thyroiditis were significantly elevated in
the AIT group, compared with the CON group. In the AMD3100 group,
the severity of lymphocytic infiltration in the thyroid was lower,
compared with that in AIT group. The scores of thyroiditis were not
significantly reduced in the AMD3100 group, compared with the CON
group (Fig. 2C and D).
mRNA expression of IFNγ is reduced and
IL10 is increased in the spleen of AMD3100-treated mice
The mRNA expression of IL10 decreased in the AIT
group, compared with the CON group (2.41±0.61, vs. 5.34±0.38,
respectively; P=0.018). However, the mRNA expression of IL10 was
increased in AMD3100 group, compared with the AIT group (5.79±0.90,
vs. 2.41±0.61, respectively; P=0.039; Fig. 3A). The mRNA expression of IFNγ was
higher in the AIT group, compared with the CON group (5.67±0.076,
vs. 2.44±0.20, respectively; P=0.008). The mRNA expression of IFNγ
was reduced in the AMD3100 group, compared with the AIT group
(2.43±0.44, vs. 5.67±0.076, respectively; P=0.008; Fig. 3B). However, no significant
differences were observed in the expression levels of IFNγ and IL10
between the AMD3100 and NOD groups.
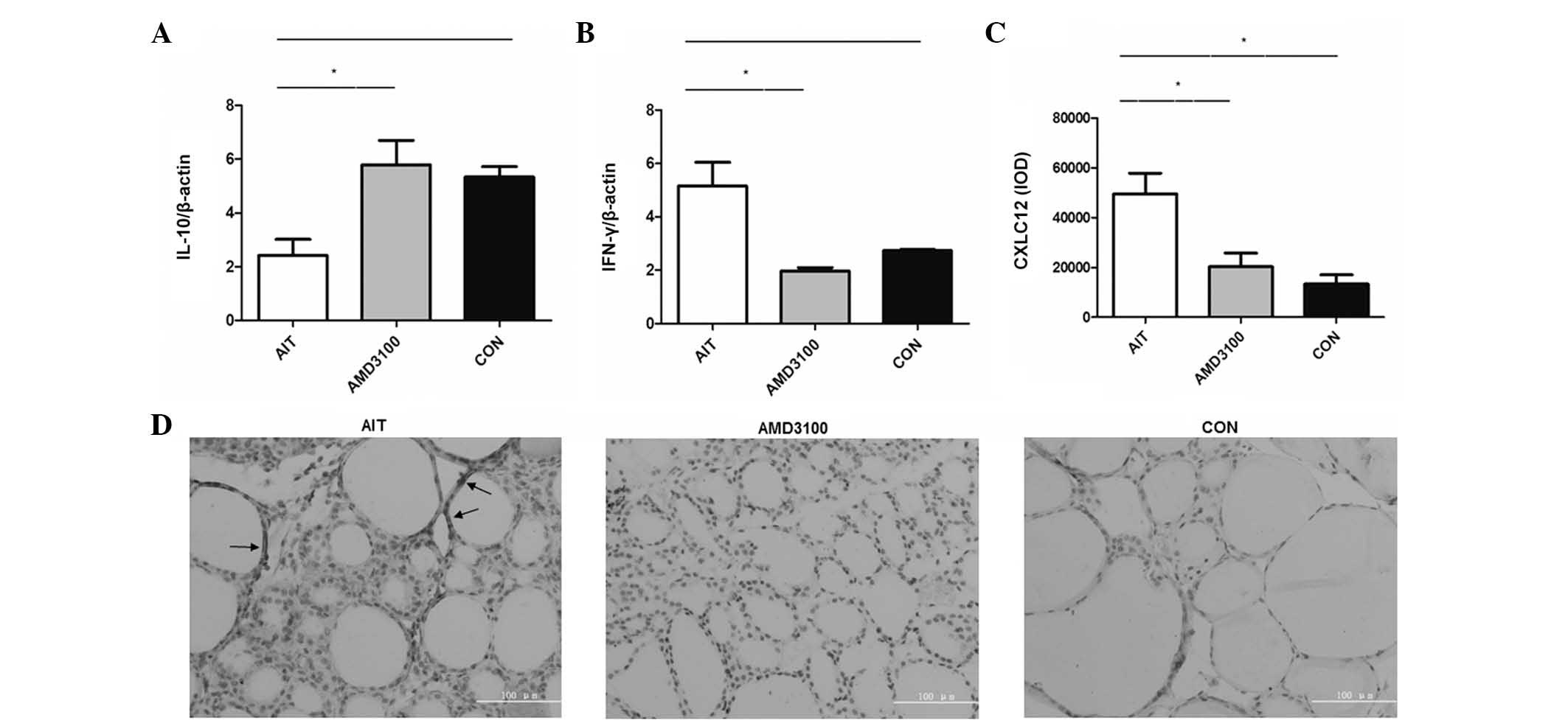 | Figure 3mRNA expression levels of IFNγ, IL10
and CXCL12 in AMD3100-treated mice. (A) CXCL12, expressed as the
IOD value; mRNA expression levels of (B) IFNγ and (C) IL-10. IFNγ
were determined in the spleen busing reverse
transcription-quantitative polymerase chain reaction analysis, and
expressed relative to β-actin. Data are presented as the mean ±
standard error of the mean of four-five mice per group. Statistical
analyses were performed consecutively with one-way analysis of
variance and Bonferroni's or Dunnett's T3 tests.
*P<0.05 and **P<0.001. (D)
Immunohistochemistry-stained thyroid sections from the different
groups, arrows indicate CXCL12. (magnification, ×400; scale bar=100
µm). CXCL12, CXC chemokine ligand 12; IFN, interferon γ;
IL-10, interleukin-10; AIT; autoimmune thyroiditis; CON, control;
IOD, integral optical density. |
CXCL12 is reduced in the thyroid of
AMD3100-treated mice
CXCL12 was distributed predominantly in the thyroid
cells. The IOD values of CXCL12 were increased in the AIT group,
compared with the CON group; however, the IOD values of CXCL12 were
reduced in the AMD3100-treated group, compared with the AIT group.
No significant difference was observed between the AMD3100 group
and CON group (Fig. 3C and D).
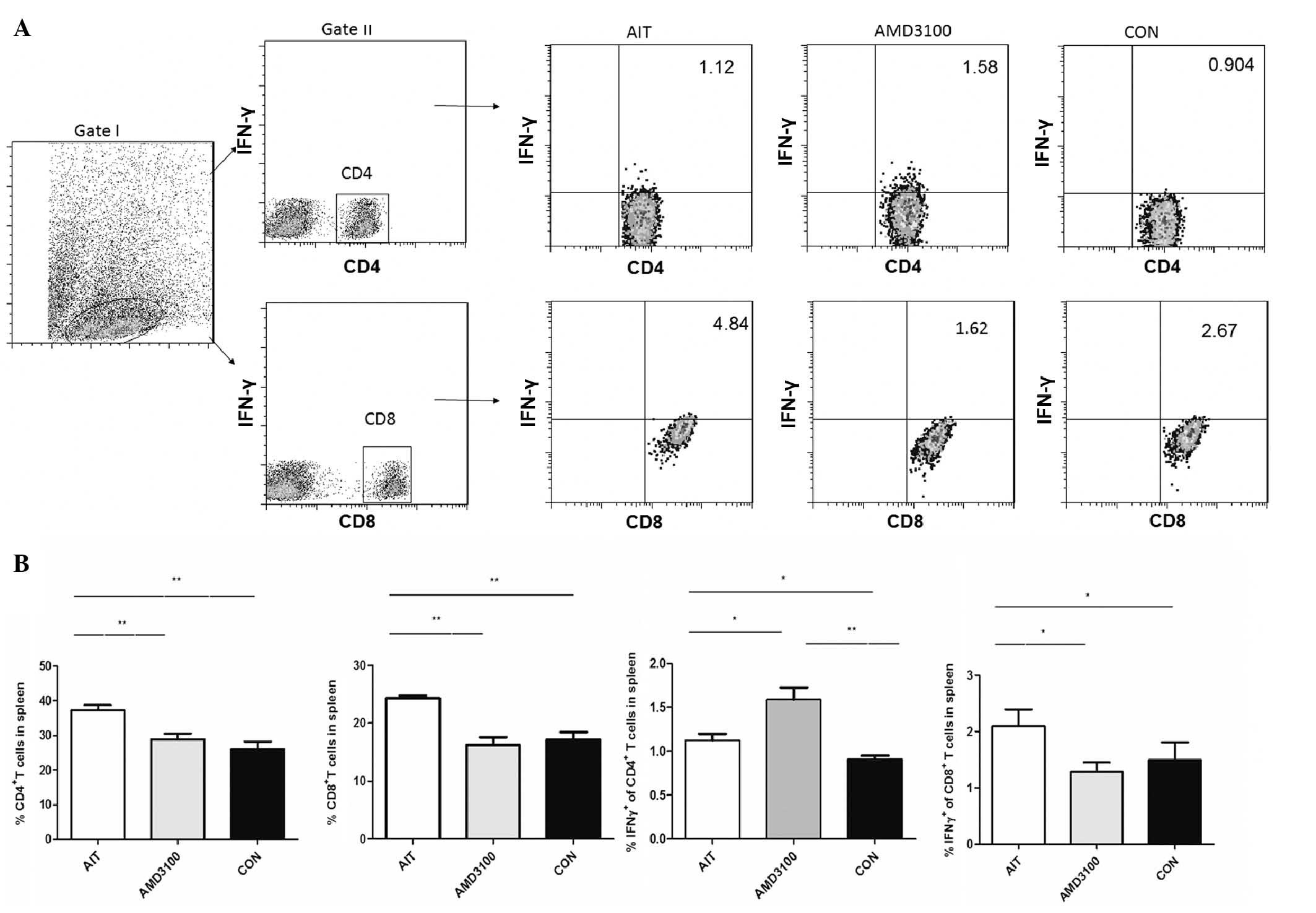
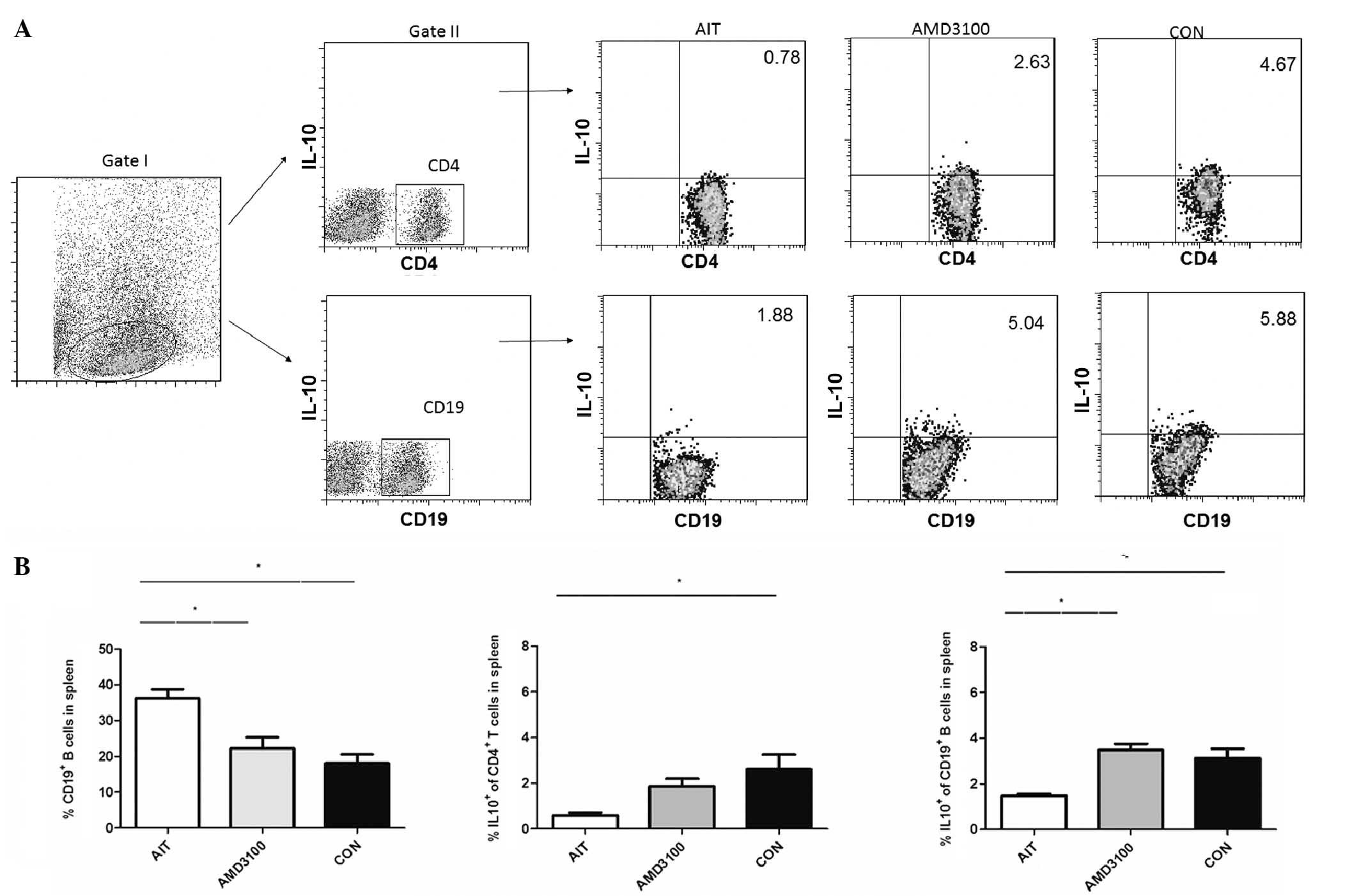
Changes in T cells and B cells in the
spleen of AMD3100-treated mice
The present study evaluated the percentages of
CD4+ T cells, CD8+ T cells, CD19+
B cells, CD4+IL10+ T cells,
CD19+IL10+ B cells,
CD4+IFNγ+ T cells and
CD8+IFNγ+ T cells in the splenocyte
population (Figs. 4 and 5). As summarized, the percentages of
CD4+ T cells, CD8+ T cells and
CD19+ B cells were significantly increased in the AIT
group, compared with the CON group; however, the percentages were
significantly decreased in the AMD3100 group, compared with the AIT
group. The percentages of CD4+IFNγ+ T cells
were significantly increased in the CON group, AIT group and
AMD3100 group, respectively. The percentages of
CD8+IFNγ+ T cells were significantly
increased in the AIT group, compared with the CON group, but were
significantly decreased in the AMD3100 group, compared with the AIT
group. For the percentages of CD4+IL10+ T
cells, significantly lower levels were observed in the AIT group,
compared with the CON group, but were marginally increased in the
AMD3100 group, compared with the AIT group. Similarly,
CD19+IL10+ B cells were reduced in the AIT
group, compared with the CON group, and were significantly elevated
in the AMD3100 group, compared with the AIT group. No significant
differences were observed in the percentages of CD4+ T
cells, CD8+ T cells, CD19+ B cells,
CD4+IFNγ+ T cells,
CD8+IFNγ+ T cells,
CD4+IL10+ T cells and
CD19+IL10+ B cells between the AMD3100 group
and CON group.
Discussion
Chemokines are involved in autoimmune diseases;
however, there are few reports regarding changes in the expression
of chemokines in iodine-induced autoimmune thyroiditis
NOD.H-2h4 mouse models. Following the provision of 0.05%
NaI in drinking water to NOD.H-2h4 mice, the incidence
of subacute thyroiditis (SAT) is almost 100% in female and male
mice at 6–8 weeks of age (13),
whereas the BALB/c mice strain is not an autoimmune
disease-susceptible species. In the present study, the thyroids of
NOD.H-2h4 mice and BALB/c mice were examined, and almost
100% of the NOD.H-2h4 mice developed thyroiditis after 8
weeks pf drinking high iodine water. However, no BAL/C mice
developed thyroiditis. The mRNA expression levels of CXCL12 were
increased in the NOD.H-2h4-HI group mice.
When the NOD.H-2h4 mice did not form
thyroid lymphocytic infiltration, there was no difference in the
mRNA expression of CXCL12 between the HI and CON groups. However,
when the NOD.H-2h4 mice developed thyroid lymphocytic
infiltration, the mRNA expression of CXCL12 was significantly
increased in the HI group, compared with the CON group. This
suggested an association between lymphocytic infiltration and
chemokine increase in the NOD.H-2h4-HI group. As CXCL12
appeared to be one of the important factors involved in recruiting
lymphocytes to the thyroid in NOD.H-2h4-HI group for 8
weeks. Therefore, the present study inhibited the binding of CXCL12
and CXCR4, to investigate the possible mechanisms of CXCL12 in
thyroiditis.
In the present study, elimination of CXCL12-mediated
effects by antagonization of CXCR4 through treatment with AMD3100
significantly decreased thyroiditis activity. Using
immunohistochemistry, it was found that CXCL12 was upregulated in
the AIT group, compared with the CON group, but was almost absent
in the AMD3100-treated group. In addition, as the levels of CXCL12
decreased, reduced inflammation of the thyroid gland was observed.
This suggested that CXCL12 is one of the important cytokines in
AIT, and circulating CXCR4+ leukocytes may be attracted
to inflamed tissues. In addition, the level of CXCL12 was increased
in the AIT thyroid, and all these mice had lymphocytic infiltration
in the thyroids with severity scores of between 1+ and 4+, had
increased thyroid relative weights and higher serum TgAb titers in
the relative to the CON group, which were decreased in the
AMD3100-treated group. This result was in accordance with those
observed in experimental colitis (30) and autoimmune collagen-induced
arthritis mouse models (31).
Several previous studies have been performed to
investigate the role of the CXCL12/CXCR4 chemokine axis in
autoimmunity thyroiditis. Specifically, Armengol et al
(24) previously demonstrated that
the mRNA and protein expression levels of CXCL12 in the thyroid
glands of patients with HT are higher than, compared with
non-autoimmune thyroid glands, and that thyrocytes are the
predominant source of CXCL12 in autoimmune thyroiditis. It was also
suggested that, as in non-obese diabetic mouse thyroiditis
(32), iodine overload may induce
thyrocyte necrosis, which stimulates resident macrophages to
produce Th1 cell-deriving cytokines, which may induce CXCL12
synthesis by adjacent thyrocytes. Thyrocytes can also produce CCL21
(33), CXCL9 and CXCL10 (20), and can induce lymphoid follicles,
which arise in the thyroid gland and even in multinodular goiter, a
clinical entity of uncertain etiology. In other animal model
autoimmunity diseases, including mouse models of dextran sulfate
sodium (DSS)-induced colitis (8),
the expression of CXCL12 in the colon of the mice were markedly
increased; and application of a CXCR4 antagonist, TF14016, remitted
the colonic inflammation in DSS-induced colitis and reduced TNFα
and IFNγ production in the mesenteric lymph node cells. This
suggested a possible role for the CXCL12/CXCR4 chemokine axis in
the pathophysiology of autoimmune diseases.
It has been confirmed that the CXCL12/CXCR4 axis is
an efficacious leukocyte chemoattractant, which can attract
lymphocytes and mononuclear cells from the bloodstream to sites of
inflammation (30). In the present
study, it was found that disease progression in
NOD.H-2h4 mice may be promoted by the elevated
expression of CXCL12 through its effect on T cell trafficking,
which is partly adjusted by the CXCL12-CXCR4 interaction. It has
been documented that CD4+, CD8+ T cells and
CD19+ B cells are involved in the destruction of thyroid
follicles in AIT (32,34). The present study showed that
increased numbers of CD4+, CD8+T cells and
CD19+ B cells were present in the spleen of the AIT
mice, compared with the CON mice. Following AMD3100 treatment of
NOD.H-2h4 mice, the percentages of CD4+,
CD8+ T cells and CD19+ B cells decreased
significantly in the spleen. This indicated that elevated
expression of CXCL12 may dysregulate lymphocyte trafficking. Thus,
CXCR4-expressing T cells may be enlisted to the immune sites of AIT
by enhanced expression of CXCL12, and CXCL12 is likely a crucial
contributing factor to the development of thyroiditis in
NOD.H-2h4 mice.
In the present study, the CXCR4 antagonist, AMD3100,
significantly reduced the mRNA expression of IFNγ, but increased
the mRNA expression of IL10. To further understand the reason for
the effect of AMD3100 on IFNγ and IL10 production, the present
study investigated its effects on T cells and B cells in the
spleen. It was found that NOD.H-2h4 AIT mice provided
with NaI had lower percentages of CD4+IL10+ T
cells and CD19+IL10+ B cells. Upon
administration of the AMD3100, the percentages of
CD4+IL10+ T cells and
CD19+IL10+ B cells increased. IL10-expressing
T cells (T regulatory-1 cells) are involved in the balance of
immune responses and the deletion of destructive tissue pathology
by limiting and stopping immune responses (35,36).
These results are in agreement with a previous report that
indicated IL10 can protect against the development of HT (37). IFNγ is a Th1 cytokine produced by
CD4+ Th1 cells and is critical for the development of
SAT in NOD.H-2h4 mice (38), and is also secreted by
CD4+ T cells, CD8+ T cells and natural killer
(NK) cells. IFNγ alone or in combination with other inflammatory
cytokines upregulates the expression of adhesion molecules, and
certain chemokines and chemokine receptors to recruit T cells to
the site of inflammation. Annunziato et al reported that
IFNγ downregulates the expression of CXCR4 on the surface of T
lymphocytes (39), suggesting that
this receptor may be connected with the Th cell. In the results of
the flow cytometric analysis in the present study,
CD4+IFNγ+ T cells and
CD8+IFNγ+ T cells were higher in number in
the AIT group, compared with the control group. Following AMD3100
treatment of the NOD.H-2h4 mice, the percentage of
CD4+IFNγ+ T cells increased, but the
percentage of CD8+IFNγ+ T cells decreased in
the spleen. The total mRNA expression of IFNγ was reduced in the
AMD3100 treatment group. CD8+IFNγ+ T cells
may be important for the decreased expression of IFNγ noted in the
AMD3100-treated group. The expression of IFNγ+
NK+ cells was not analyzed and, therefore, requires
further verification.
Therefore, recruiting IFNγ- and IL10-producing
CD4+ T cells and CD19+ B cells via CXCL12
into an inflamed thyroid site may be an important regulatory
mechanism. Taken together, the immune response in AIT is complex,
due to the involvement of Th 1 cells, the B cell immune response
and pro-inflammatory and regulatory cytokines. To the best of our
knowledge, the present study is the first suggesting that the
elimination of CXCL12-mediated effects may treat thyroiditis via
alterations in Th 1 cells, the B cell immune response, and
pro-inflammatory and regulatory cytokines.
AMD3100 was initially developed to treat human
immunodeficiency virus infections through the antagonism of the
CXCR4 (40). It is a slow,
tightly-binding, reversible inhibitor (41). There is a transfer from the
pathogenic T helper cytokine profile to the antagonistic T helper
cytokine profile in AMD3100-treated mice (42). The present study demonstrated that
AMD3100 treatment of AIT NOD.H-2h4 mice significantly
reduced the infiltration of lymphocytes in the thyroid, and
decreased the titer of TgAb. Therefore, CXCL12 may affect IL10 and
IFNγ production in AIT mice, and this may be inhibited by AMD3100.
Currently, the role of AMD3100 in autoimmune diseases is
controversial. For example, in mouse models of type I diabetes
melitus (11), collagen-induced
arthritis (9), DSS-induced colitis
(30) and asthma (12); the treatment of mice with AMD3100
significantly reduced the severity of the disease. By contrast,
several studies have reported that AMD3100 treatment promotes
disease development (8,43,44).
Thus, the results of treatment with AMD3100 have indicated that, in
addition to its pro-inflammatory role in autoimmune experimental
models and human autoimmune diseases, CXCL12 may also have
anti-inflammatory properties.
The precise mechanism underlying the differing
conclusions of previous studies remains to be elucidated, however,
the findings of the present study lay the foundation for further
investigation of AMD3100, for the prevention and/or treatment of
autoimmune thyroiditis and possibly other autoimmune diseases with
elevated expression levels of CXCL12.
In conclusion, the present study demonstrated that
the CXCR4 antagonist, AMD3100, decreased the severity of autoimmune
thyroiditis in mice, by inhibiting the production of cytokines
and/or the migration of cytokine-producing lymphocytes. This
revealed the therapeutic potential for the CXCR4 antagonist in the
treatment of thyroiditis.
Abbreviations:
|
NaI
|
sodium iodide
|
|
AIT
|
autoimmune thyroiditis
|
|
SAT
|
subacute thyroiditis
|
|
HT
|
Hashimoto's thyroiditis
|
|
TgAb
|
thyroglobulin antibody
|
Acknowledgments
The present study was supported by the Department of
Science and Technology at the Research Foundation of Key Laboratory
of Endocrine Diseases (grant no. F11-244-1-00) and the PhD Programs
Foundation of Ministry of Education of China (grant no.
20122104120001).
References
|
1
|
Rotondi M, Chiovato L, Romagnani S, Serio
M and Romagnani P: Role of chemokines in endocrine autoimmune
diseases. Endocrine Rev. 28:492–520. 2007. View Article : Google Scholar
|
|
2
|
D'Apuzzo M, Rolink A, Loetscher M, Hoxie
JA, Clark-Lewis I, Melchers F, Baggiolini M and Moser B: The
chemokine SDF-1, stromal cell-derived factor 1, attracts early
stage B cell precursors via the chemokine receptor CXCR4. Eur
Immunol. 27:1788–1793. 1997. View Article : Google Scholar
|
|
3
|
Murdoch C: CXCR4: Chemokine receptor
extraordinaire. Immunol Rev. 177:175–184. 2000. View Article : Google Scholar
|
|
4
|
Momcilović M, Mostarica-Stojković M and
Miljković D: CXCL12 in control of neuroinflammation. Immunol Res.
52:53–63. 2012. View Article : Google Scholar
|
|
5
|
Jinquan T, Quan S, Jacobi HH, Madsen HO,
Glue C, Skov PS, Malling HJ and Poulsen LK: CXC chemokine receptor
4 expression and stromal cell-derived factor-1alpha-induced
chemotaxis in CD4+ T lymphocytes are regulated by interleukin-4 and
interleukin-10. Immunology. 99:402–410. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Gupta SK, Lysko PG, Pillarisetti K,
Ohlstein E and Stadel JM: Chemokine receptors in human endothelial
cells. Functional expression of CXCR4 and its transcriptional
regulation by inflammatory cytokines. J Biol Chem. 273:4282–4287.
1998. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Wang A, Guilpain P, Chong BF, Chouzenoux
S, Guillevin L, Du Y, Zhou XJ, Lin F, Fairhurst AM, Boudreaux C, et
al: Dysregulated expression of CXCR4/CXCL12 in subsets of patients
with systemic lupus erythematosus. Arthritis Rheum. 62:3436–3446.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Mikami S, Nakase H, Yamamoto S, Takeda Y,
Yoshino T, Kasahara K, Ueno S, Uza N, Oishi S, Fujii N, et al:
Blockade of CXCL12/CXCR4 axis ameliorates murine experimental
colitis. J Pharmacol Exp Ther. 327:383–392. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
De Klerck B, Geboes L, Hatse S,
Kelchtermans H, Meyvis Y, Vermeire K, Bridger G, Billiau A, Schols
D and Matthys P: Pro-inflammatory properties of stromal
cell-derived factor-1 (CXCL12) in collagen-induced arthritis.
Arthritis Res Ther. 7:R1208–R1220. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Donzella GA, Schols D, Lin SW, Esté JA,
Nagashima KA, Maddon PJ, Allaway GP, Sakmar TP, Henson G, De Clercq
E and Moore JP: AMD3100, a small molecule inhibitor of HIV-1 entry
via the CXCR4 co-receptor. Nat Med. 4:72–77. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Leng Q, Nie Y, Zou Y and Chen J: Elevated
CXCL12 expression in the bone marrow of NOD mice is associated with
altered T cell and stem cell trafficking and diabetes development.
BMC Immunol. 9:512008. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Lukacs NW, Berlin A, Schols D, Skerlj RT
and Bridger GJ: AMD3100, a CxCR4 antagonist, attenuates allergic
lung inflammation and airway hyperreactivity. Am J Pathol.
160:1353–1360. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Braley-Mullen H, Sharp GC, Medling B and
Tang H: Spontaneous autoimmune thyroiditis in NOD. H-2h4 mice J
Autoimmun. 12:157–165. 1999. View Article : Google Scholar
|
|
14
|
Charreire J: Immune mechanisms in
autoimmune thyroiditis. Adv Immunol. 46:263–334. 1989. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Ajjan RA, Watson PF, McIntosh RS and
Weetman AP: Intrathyroidal cytokine gene expression in Hashimoto's
thyroiditis. Clin Exp Immunol. 105:523–528. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Roura-Mir C, Catálfamo M, Sospedra M,
Alcalde L, Pujol-Borrell R and Jaraquemada D: Single-cell analysis
of intrathyroidal lymphocytes shows differential cytokine
expression in Hashimoto's and Graves' disease. Eur J Immunol.
27:3290–3302. 1997. View Article : Google Scholar
|
|
17
|
Paschke R, Schuppert F, Taton M and Velu
T: Intrathyroidal cytokine gene expression profiles in autoimmune
thyroiditis. J Endocrinol. 141:309–315. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Salzano M, Russo E, Postiglione L, Guerra
A, Marotta V, Esposito S and Vitale M: Interferon-γ inhibits
integrin-mediated adhesion to fibronectin and survival signaling in
thyroid cells. J Endocrinol. 215:439–444. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Watson PF, Pickerill AP, Davies R and
Weetman AP: Semi-quantitative analysis of interleukin-1 alpha,
interleukin-6 and interleukin-8 mRNA expression by human
thyrocytes. J Mol Endocrinol. 15:11–21. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Garcia-Lopez MA, Sancho D, Sanchez-Madrid
F and Marazuela M: Thyrocytes from autoimmune thyroid disorders
produce the chemokines IP-10 and Mig and attract CXCR3+
lymphocytes. J Clin Endocrinol Metab. 86:5008–5016. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Goulvestre C, Batteux F and Charreire J:
Chemokines modulate experimental autoimmune thyroiditis through
attraction of autoreactive or regulatory T cells. Eur J Immunol.
32:3435–3442. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Antonelli A, Ferri C, Fallahi P, Ferrari
SM, Frascerra S, Carpi A, Nicolini A and Ferrannini E:
Alpha-chemokine CXCL10 and beta-chemokine CCL2 serum levels in
patients with hepatitis C-associated cryoglobulinemia in the
presence or absence of autoimmune thyroiditis. Metabolism.
57:1270–1277. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Campbell JJ and Butcher EC: Chemokines in
tissue-specific and microenvironment-specific lymphocyte homing.
Curr Opin Immunol. 12:336–341. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Armengol MP, Cardoso-Schmidt CB, Fernández
M, Ferrer X, Pujol-Borrell R and Juan M: Chemokines determine local
lymphoneogenesis and a reduction of circulating CXCR4+ T and CCR7 B
and T lymphocytes in thyroid autoimmune diseases. J Immunol.
170:6320–6328. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
National Research Council (US) Committee
for the Update of the Guide for the Care and Use of Laboratory
Animals: Guide for the Care and Use of Laboratory Animals. 8th
edition. Washington (DC): National Academies Press (US); 2011
|
|
26
|
Xue H, Wang W, Shan Z, Li Y, Li Y, Teng X,
Gao Y, Fan C and Teng W: Dynamic changes of CD4+CD25+ regulatory T
cells in NOD.H-2h4 mice with iodine-induced autoimmune thyroiditis.
Biol Trace Elem Res. 143:292–301. 2011. View Article : Google Scholar
|
|
27
|
Teng X, Shan Z, Teng W, Fan C, Wang H and
Guo R: Experimental study on the effects of chronic iodine excess
on thyroid function, structure and autoimmunity in autoimmune-prone
NOD.H-2h4 mice. Clin Exp Med. 9:51–59. 2009. View Article : Google Scholar
|
|
28
|
Imaizumi M, Pritsker A, Kita M, Ahmad L,
Unger P and Davies T: Pregnancy and murine thyroiditis:
Thyroglobulin immunization leads to fetal loss in specific
allogeneic pregnancies. Endocrinology. 142:823–829. 2001.PubMed/NCBI
|
|
29
|
Zhang L, Teng W, Liu Y, Li J, Mao J, Fan
C, Wang H, Zhang H and Shan Z: Effect of maternal excessive iodine
intake on neurodevelopment and cognitive function in rat offspring.
BMC Neurosci. 13:1212012. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Xia XM, Wang FY, Xu WA, Wang ZK, Liu J, Lu
YK, Jin XX, Lu H and Shen YZ: CXCR4 antagonist AMD3100 attenuates
colonic damage in mice with experimental colitis. World J
Gastroenterol. 16:2873–2880. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Matthys P, Hatse S, Vermeire K, Wuyts A,
Bridger G, Henson GW, De Clercq E, Billiau A and Schols D: AMD3100,
a potent and specific antagonist of the stromal cell-derived
factor-1 chemokine receptor CXCR4, inhibits autoimmune joint
inflammation in IFN-gamma receptor-deficient mice. J Immunol.
167:4686–4692. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Hutchings PR, Verma S, Phillips JM, Harach
SZ, Howlett S and Cooke A: Both CD4(+) T cells and CD8(+) T cells
are required for iodine accelerated thyroiditis in NOD mice. Cell
Immunol. 192:113–121. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Grant AJ, Goddard S, Ahmed-Choudhury J,
Reynolds G, Jackson DG, Briskin M, Wu L, Hübscher SG and Adams DH:
Hepatic expression of secondary lymphoid chemokine (CCL21) promotes
the development of portal-associated lymphoid tissue in chronic
inflammatory liver disease. Am J Pathol. 160:1445–1455. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Yu S, Medling B, Yagita H and
Braley-Mullen H: Characteristics of inflammatory cells in
spontaneous autoimmune thyroiditis of NOD.H-2h4 mice. J Autoimmun.
16:37–46. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Ganesh BB, Bhattacharya P, Gopisetty A and
Prabhakar BS: Role of cytokines in the pathogenesis and suppression
of thyroid autoimmunity. J Interferon Cytokine Res. 31:721–731.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Trinchieri G: Regulatory role of T cells
producing both interferon gamma and interleukin 10 in persistent
infection. J Exp Med. 194:F53–F57. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Shi L, Bi M, Yang R, Zhou J, Zhao S, Fan
C, Shan Z, Li Y and Teng W: Defective expression of regulatory B
cells in iodine-induced autoimmune thyroiditis in non-obese
diabetic H-2(h4) mice. J Endocrinol Invest. 37:43–50. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Fang Y, Yu S and Braley-Mullen H:
Contrasting roles of IFN-gamma in murine models of autoimmune
thyroid diseases. Thyroid. 17:989–994. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Annunziato F, Cosmi L, Galli G, Beltrame
C, Romagnani P, Manetti R, Romagnani S and Maggi E: Assessment of
chemokine receptor expression by human Th1 and Th2 cells in vitro
and in vivo. J Leukoc Biol. 65:691–699. 1999.PubMed/NCBI
|
|
40
|
De Clercq E: The bicyclam AMD3100 story.
Nat Rev Drug Discov. 2:581–587. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Fricker SP, Anastassov V, Cox J, Darkes
MC, Grujic O, Idzan SR, Labrecque J, Lau G, Mosi RM, Nelson KL, et
al: Characterization of the molecular pharmacology of AMD3100: A
specific antagonist of the G-protein coupled chemokine receptor,
CXCR4. Biochem Pharmacol. 72:588–596. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Momcilović M, Mostarica-Stojković M and
Miljković D: CXCL12 in control of neuroinflammation. Immunol Res.
52:53–63. 2012. View Article : Google Scholar
|
|
43
|
Aboumrad E, Madec AM and Thivolet C: The
CXCR4/CXCL12 (SDF-1) signalling pathway protects non-obese diabetic
mouse from autoimmune diabetes. Clin Exp Immunol. 148:432–439.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Brunn A, Utermöhlen O, Mihelcic M,
Sánchez-Ruiz M, Carstov M, Blau T, Ustinova I, Penfold M,
Montesinos-Rongen M and Deckert M: Differential effects of
CXCR4-CXCL12- and CXCR7-CXCL12-mediated immune reactions on murine
P0106-125-induced experimental autoimmune neuritis. Neuropathol
Appl Neurobiol. 39:772–787. 2013. View Article : Google Scholar : PubMed/NCBI
|