Introduction
Colon cancer is a common malignancy with a high
incidence worldwide, which is increasing each year in South Korea
as the population ages and adapts to westernized eating habits
(1–3). Most attention is focused on
post-operative cancer metastasis, as the post-operative survival
rate of colon cancer is only 50% (4). Cancer cells require large amounts of
energy and molecules, including lipids and amino acids, for cell
growth and proliferation and to protect themselves through the
regulation of pro-survival factors (5). The number of studies on the potential
use of natural plant extracts for the inhibition of cancer-cell
proliferation and metastasis through the regulation of pro-survival
factors has increased (4–6). A number of previous studies have
shown that various active components of herbal medicines, such as
curcumin, as well as selenium, induce apoptosis through the
regulation of AMP-activated protein kinase (AMPK) and Akt in colon
cancer cells (4,5,7). The
adjustment of eating habits may have beneficial effects on the AMPK
and Akt pathways and prevent the occurrence or the progression of
colon cancer.
Apoptosis is the programmed cell death as a response
to cell damage through various biological events, including DNA
damage, hypoxia, cell cycle arrest and metabolic stress (8). AMPK is activated by metabolic stress,
including hypoxia, heat shock and energy deprivation. In addition,
AMPK induces apoptosis through the inhibition of the mammalian
target of rapamycin (mTOR) pathway, which regulates the growth and
supply of nutrition to cancer cells (5,9,10).
Furthermore, AMPK has been demonstrated to be an upstream factor of
the phosphoinositide-3 kinase (PI3K)/Akt pathway and induce the
phosphorylation of forkhead box O3a (11). Akt, one of the serine/threonine
protein kinases, regulates cancer growth and proliferation by
phosphorylation of mTOR, and inhibits the expression of
pro-apoptotic proteins, including B-cell lymophoma 2
(Bcl-2)-associated X protein (Bax) and Bcl-2 homologous antagonist
killer (Bak), thereby suppressing apoptosis (12,13).
A previous study showed that a polysaccharide
extract from a medicinal mushroon induced apopotosis in colon
cancer cells via the suppression of Bcl-2 and activation of Bax/Bak
(14). The intrinsic apoptotic
pathway, which is mitochondria-dependent, is activated by the
release of apoptotic molecules into the cytosol from the
mitochondria through a change in mitochondrial membrane
permeability (MMP) caused by mitochondrial dysfunction (8,15–17).
The Bcl-2 family is well known to participate in changes of the
MMP; in particular, Bcl-2 has been found to suppress apoptosis
through inhibiting pro-apoptotic proteins, including Bax and Bak
(8,13,18).
The Bcl-2 family also has a major role in the
mitochondria-dependent activation of caspases (19). Therefore, regulation of the
pro-survival factor Bcl-2, and its upstream regulators may
represent an effective treatment for cancer.
Numerous studies have shown that Bax and Bax are
major pro-apoptotic proteins in mitochondria-mediated apoptosis.
Activation of Bax/Bak results in their translocation to
mitochondria from the cytosol, where they oligomerize with each
other (20). For this reason,
pores are formed in the mitochondrial outer membrane and cytochrome
C is released into the cytosol (21), where it activates caspase-9, which
in turn forms the apoptosome complex with apoptotic protease
activating factor 1 (19,22). The apoptosome complex activates
caspase-3 through the cleavage of pro-caspase-3 (22).
A large number of previous studies have detailed the
apoptotic effects of various plant extracts through the regulation
of AMPK and the Akt pathway in colon cancer cells (1,4,9).
Cnidium monnieri (L.) Cusson is an umbelliferous plant and
has been used for the treatment of impotence, pain in female
genitalia, suppurative dermatitis and carbuncle (23,24).
The fruit of Cnidium monnieri (L.) Cusson is widely used as
a medicinal herb (25); however,
the anti-cancer effects of Cnidium monnieri (L.) Cusson
extract (CME) have not been investigated. Accordingly, the present
study investigated the effects of CME on mitochondria-mediated
apoptotic proteins and apoptosis in HCT116 colon cancer cells.
Furthermore, AMPK and Akt inhibitors were employed to assess
whether the mechanism of action of CME is dependent or independent
of the associated signaling pathways.
Materials and methods
Reagent
Cnidium monnieri (L.) Cusson (CME) was
purchased from Dong Kyung PHARM (Seoul, Korea). A total of 100 g
CME was soaked in 99% ethanol (800 ml) stirred at room temperature
for 48 h. The extract was filtered through qualitative filter paper
no.1 (Toyo Roshi Kaisha, Ltd., Tokyo, Japan) and concentrated with
a rotary evaporator to remove the ethanol. CME was dissolved in
dimethyl sulfoxide (stock solution, 120–200 µg/ml) prior to
treatment and stored at −20°C. The final concentration of CME in
the culture medium was controlled at 120–200 µg/ml. The
fluorescein isothiocyanate-Annexin V apoptosis detection kit was
obtained from BD Biosciences (Franklin Lakes, NJ, USA). The Pierce
lactate dehydrogenase (LDH) Cytotoxicity Assay kit was purchased
from Thermo Fisher Scientific (Waltham, MA, USA). The Caspase-3
Activity Assay kit was obtained from Abcam (Cambridge, MA, USA).
Specific antibodies that recognized phosphorylated (p)mTOR
(Ser2448) (2971), (p)Akt (Ser473) (4051), (p)AMPKα1 (Thr172)
(2535), Bax (5023), Bak (6947), pro-caspase-3 (9665), Bcl-2 (2876)
and β-actin (4967) were obtained from Cell Signaling Technology
(Beverly, MA, USA) and P53-upregulated modulator of apoptosis
(PUMA) antibody (4976) was purchased from Santa Cruz Biotechnology
(Dallas, TX, USA). LY294002 (PI3K/Akt inhibitor) and Compound C
(AMPK inhibitor) were purchased from Calbiochem (San Diego, CA,
USA). Horseradish peroxidase (HRP)-conjugated goat anti-mouse
(PA1-30126) and goat anti-rabbit (166–2408) secondary antibodies
were purchased from Thermo Fisher Scientific, Inc., and Bio-Rad
Laboratories, Inc., (Tokyo, Japan), respectively.
Cell culture
HCT116 colon cancer cells were obtained from the
American Type Culture Collection (Rockville, MD, USA). The cells
were grown in RPMI-1640 medium (Hyclone Laboratories Inc., Logan,
UT, USA) containing 10% fetal bovine serum (Hyclone Laboratories
Inc.) and 1% antibiotics (100 mg/l streptomycin and 100 U/ml
penicillin; Hyclone Laboratories Inc.) at 37°C in a 5%
CO2 atmosphere. The cells were sub-cultured by
detachment with Trypsin-EDTA (Hyclone Laboratories Inc.) and
re-seeding at 1×106 cells/ml per 100-mm plate every 48
h.
LDH release assay
Cells were seeded at 2.5×105 cells/ml per
well in a 96-well plate and incubated for 24 h. The cells were then
treated with CME (120–200 µg/ml) and then incubated at 37°C
in a 5% CO2 atmosphere. Certain samples were pre-treated
with the respective inhibitor (10 µM Compound C or 40
µM LY294002) for 30 min prior to treatment with CME. After
24 h, the high control cells (maximum LDH release) control cells
were treated with Cell Lysis solution from the LDH Cytotoxicity
Assay kit for 45 min followed by centrifugation at 250 ×g for 3
min. The absorbance of the solu tion in each well was determined
using a microplate reader (Bio-Rad Laboratories, Inc.) at 490 and
655 nm.
Determination of apoptosis by Annexin
V/propidium iodide (PI) staining
Cells were seeded at 1×106 cells/ml in a
60-mm plate and incubated for 24 h. The cells were then treated
with CME (120–180 µg/ml) for 24 h at 37°C in a 5%
CO2 atmosphere. Certain samples were pre-treated with
the respective inhibitor (10 µM Compound C or 40 µM
LY294002) for 30 min prior to treatment with CME. Total cells were
harvested by trypsinization, collected by centrifugation, washed
with phosphate-buffered saline (PBS) and re-suspended in binding
buffer. Cells were stained with Annexin V and PI for 15 min.
Fluorescence intensity was analyzed using a FACS Canto flow
cytometer (BD Biosciences).
Identification of apoptosis by Hoechst
33342 staining
Cells were seeded at 1×104 cells/ml in a
12-well plate containing glass cover slips (Marienfeld-Superior
GmbH & Co., Lauda-Königshofen, Germany) and incubated for 24 h.
Following incubation, the cells were treated with the CME (120, 140
or 160 µg/ml) for 24 h at 37°C in a 5% CO2
atmosphere. The cells were then stained with 0.7 µM Hoechst
33342 and incubated for 30 min. Cells were fixed with 3.5%
formaldehyde (500 µl) for 20 min and then gently washed with
150 µl PBS for 5 min (thrice) The slips were mounted with 10
µl of mounting solution (50% glycerol). The stained
chromatin fragments indicative of apoptosis were observed using a
fluorescence microscope (magnification, ×200; Axioskop 50; Carl
Zeiss, Inc., Thornwood, NY, USA).
Western blot analysis
Cells were seeded at 1×105 cells/ml in a
six-well plate and incubated for 24 h. The cells were then treated
with CME (120, 140 or 160 µg/ml) for 24 h at 37°C in a 5%
CO2 atmosphere. Certain samples were pre-treated with
the respective inhibitor (10 µM Compound C or 40 µM
LY294002) for 30 min prior to treatment with CME. Cells were then
rinsed twice with ice-cold PBS and scraped with
radioimmunoprecipitation assay lysis buffer (50 mM Tris-HCl pH 8.0,
150 mM NaCl, 1% NP-40, 0.5% sodium deoxycholate and 1 mM
phenylmethanesulfonylfluoride) [all purchased from Cell Signaling
Technology, Inc. (Beverly, MA, USA)] and subjected to western blot
analysis. Protein quantification was performed using Bradford assay
and 30 µg protein was loaded per lane. Nitrocellulose
membranes (GE Healthcare Life Sciences, Chalfont, UK) were blocked
with 2% bovine serum albumin (Boyogen, Melbourne, Australia) in 1X
Tris-buffered saline with Tween 20 (TBST; 24.7 mM Tris-HCl, 137 mM
NaCl and 0.05% Tween-20; pH 8.0) and incubated overnight at 4°C
with following primary monoclonal antibodies: Rabbit (p)mTOR, mouse
(p)Akt, rabbit (p) AMPKα (all 1:2,000), rabbit Bax, rabbit Bak,
rabbit caspase-3; and rabbit PUMA (all 1:1,000), rabbit Bcl-2 and
rabbit β-actin (all 1:2,000) primary polyclonal antibodies.
Membranes were washed four time with 1X TBST for 5 min at room
temperature and subsequently incubated with HRP-conjugated goat
anti-mouse and goat anti-rabbit polyclonal secondary antibodies
(both 1:10,000) for 90 min at room temperature with gentle
agitation. Following washing four times with 1X TBST for 10 min at
room temperature, proteins were detected using SuperSignal West
Pico Chemiluminescent Substrate (PI34080; Thermo Fisher Scientific,
Inc., Waltham, MA, USA) and visualized on CP-BU new X-ray film
(Agfa HealthCare, Inc., Mortsel, Belgium).
Caspase-3 activity assay
Cells were seeded into a six-well plate at
1×105 cells/ml. Following incubation for 24 h, the cells
were pre-treated with or without Compound C (10 µM) or
LY94002 (40 µM) for 30 min prior to treatment with CME (160
µg/ml). Cells were harvested using Trypsin-EDTA (Hyclone
Laboratories, Inc.) and re-suspended in 50 µl cold Cell
Lysis Buffer on ice for 10 min. A total of 130 µg protein
lysate was added to a Reaction Buffer with 10 mM dithiothreitol.
After addition of 200 µM Glu-Val-Asp p-nitroanilide
(DEVD-p-NA), cells were incubated at 37°C for 1 h and 30 min. The
absorbance of the solution in each well was determined using a
microplate reader (Model 680; Bio-Rad Laboratories, Inc.) at 415
nm.
Statistical analysis
LDH release and caspase-3 activity were
statistically analyzed using an unpaired analysis of variance and
Duncan's multiple range test using SPSS 20.0 software (IBM Corp,
Armonk, NY, USA). A value of P<0.05 was considered to indicate a
statistically significant difference.
Results
CME has cytotoxic effects on HCT116 colon
cancer cells
The present study investigated the cytotoxic effects
of CME through the LDH release assay. The cells were treated with
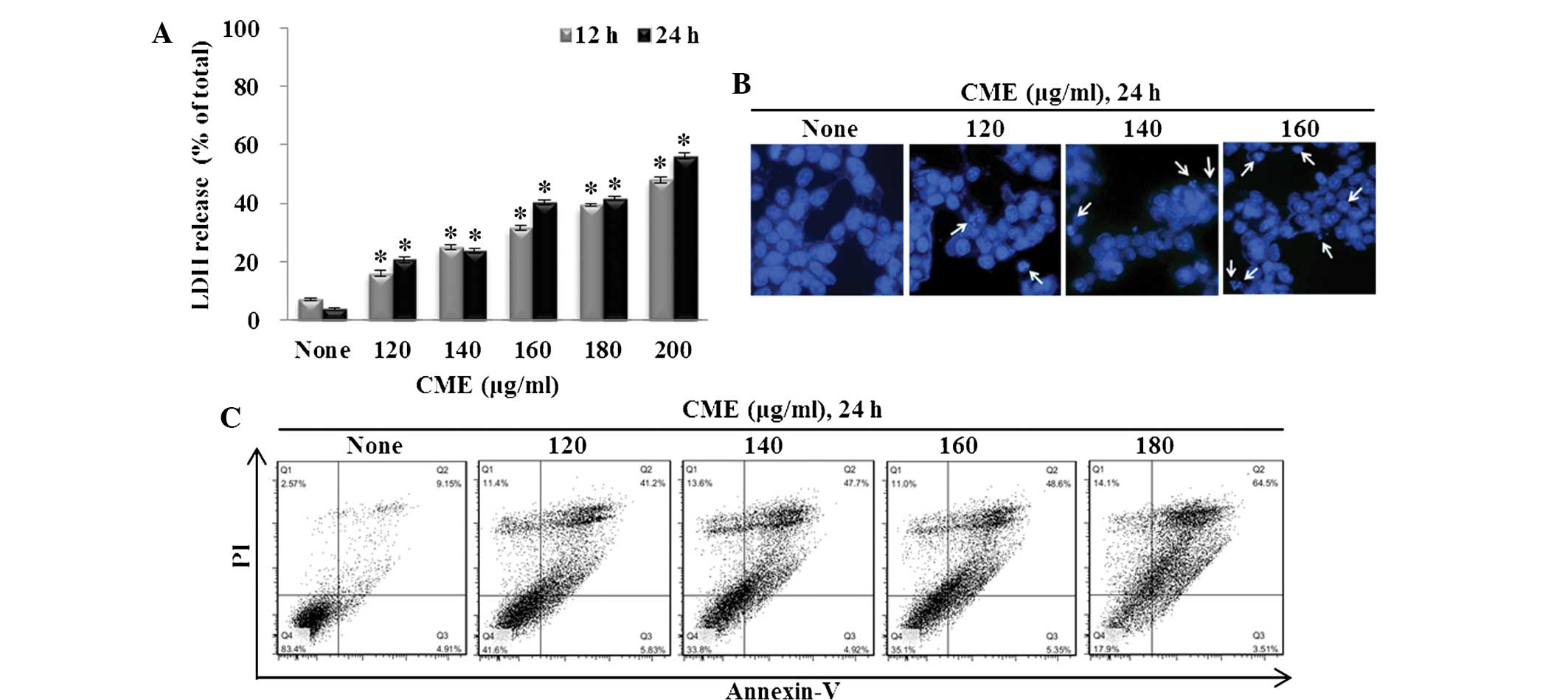
various concentrations of CME for 12 or 24 h. As shown in Fig. 1A, LDH release significantly
increased by 22.38, 32.16, 37.56, 41.38 and 50.77% following
treatment with 120, 140, 160, 180 or 200 µg/ml CME,
respectively, compared with that in the control group (24 h;
P<0.001). CME significantly induced the release of LDH in a
dose-dependent manner.
CME induces apoptosis in HCT116 colon
cancer cells
Annexin V-PI staining and Hoechst 33342 staining
were performed to identify CME-induced apoptosis. To measure the
appearance of apoptotic bodies, Hoechst 33342 staining was
performed. After treatment with CME (120, 140 or 160 µg/ml)
for 24 h, the apoptotic DNA fragmentation increased in a
dose-dependent manner (Fig. 1B).
The cells were cultured with the CME (120, 140, 160 or 180
µg/ml) for 24 h prior to Annexin V-PI staining. Then, the
percentage of Annexin V-positive cells was analyzed by flow
cytometry. As shown in Fig. 1C,
the ratio of Annexin V-positive cells was low in the control group
(14.0 6%). However, as the concentration of CME increased, the
percent of Annexin V-positive cells increased to 47.03% (120
µg/ml), 52.62% (140 µg/ml), 53.95% (160 µg/ml)
and 68.01% (180 µg/ml).
CME regulates the expression of
mitochondria-mediated apoptotic proteins
To examine the mechanisms by which CME regulates
signaling proteins during apoptosis, the expression of p-AMPK,
p-Akt and apoptosis-associated proteins was assessed by western
blot analysis. As shown in Fig.
2A, as the concentration of CME increased, the levels of p-AMPK
and PUMA increased, while the levels of p-Akt, p-mTOR,
pro-caspase-3 and Bcl-2 decreased. Furthermore, the
mitochondria-associated apoptotic proteins Bax and Bak were
increased in a dose-dependent manner (Fig. 2A).
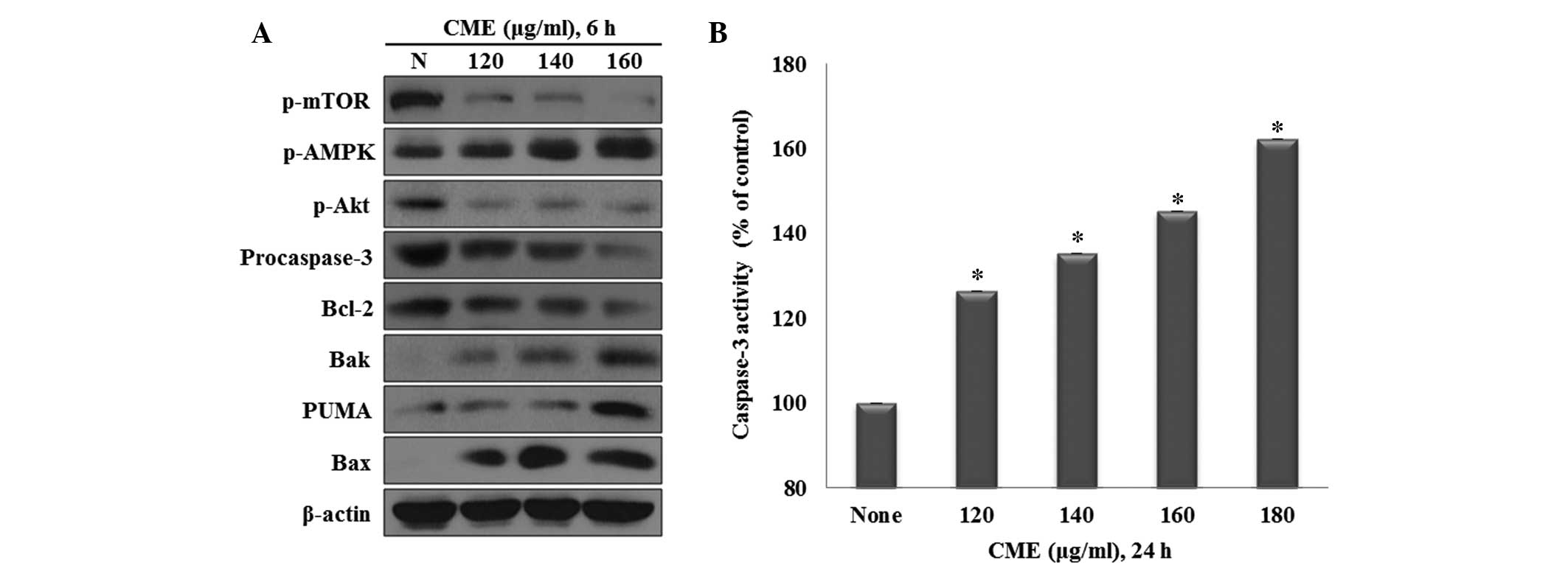 | Figure 2CME regulates the expression of
mitochondria-mediated apoptotic proteins and caspase-3 activation.
(A) Effects of CME on the levels of p-mTOR, p-AMPK, p-Akt,
procaspase-3, Bcl-2, Bak, PUMA and Bax in HCT116 colon cancer
cells. Cells were treated with 120–160 µg/ml CME for 6 h and
protein levels were determined by western blot analysis. β-actin
served as a protein-loading control. (B) CME induces caspase-3
activation in HCT116 colon cancer cells. Cells were treated with
CME (120–180 µg/ml) for 24 h. Statistical analysis was
performed using one-way analysis of variance.
*P<0.001 vs. the control, as determined by
independent sample t-test (each experiment, n=3). p,
phosphorylated; mTOR, mammalian target of rapamycin; AMPK,
adenosine monosphosphate-activated protein kinase; PUMA,
P53-upregulated modulator of apoptosis; CME, Cnidium
monnieri (L.) Cusson extract; Bcl-2, B-cell lymphoma 2; Bak,
Bcl-2-homologous antagonist killer; Bax; Bcl-2-associated X
protein. |
Effect of CME on activation of caspases-3
in HCT116 colon cancer cells
Western blot analysis demonstrated that CME
suppressed the expression of pro-caspase-3, indicating that
pro-caspase-3 was cleaved to caspase-3 (Fig. 2A). To further determine the
influence of CME on caspase-3 activation a caspase-3 activity assay
was performed using DEVD-p-NA, a substrate of caspase-3. The cells
were treated with various concentrations of CME (120, 140, 160 or
180 µg/ml) for 24 h, followed by the measurement of the
cleavage of DEVD-p-NA by caspase-3. The results showed that
CME induced caspase-3 activation through the cleavage of
pro-caspase-3 in a dose-dependent manner (Fig. 2B).
CME induces apoptosis through directly
regulating the de-phosphorylation of Akt via the AMPK-independent
pathway
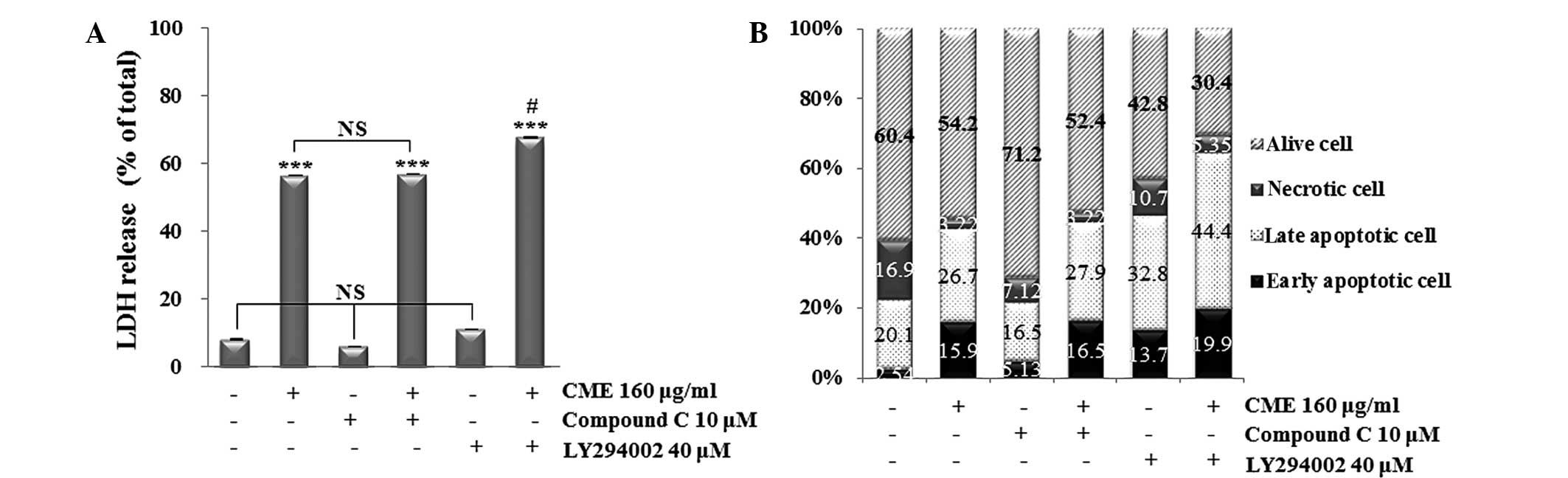
To confirm the association between CME-induced
cytotoxicity, apoptosis and the AMPK and Akt pathways, LDH release
assay and Annexin V-PI staining were performed after co-treatment
of the cells with Compound C (AMPK inhibitor) or LY294002 (Akt
inhibitor) prior to incubation with CME. The results showed that
the groups treated with Compound C or LY294002 only showed a
similar LDH release to that in the control group, while treatment
with CME alone or in combination with Compound C markedly increased
the LDH release to a similar extent (Fig. 3A). Of note, the LY294002 and CME
co-treated group showed a significant increase in LDH release
compared with that in the CME-treated group. Apoptosis was also
investigated by Annexin V/PI double staining (Fig. 3B). Treatment with Compound C alone
decreased the apoptotic rate, while treatment with LY294002 alone
increased the apoptotic rate of HCT116 cells compared with that in
the control group. Treatment with CME increased the apoptotic rate,
and pre-treatment with Compound C prior to CME treatment resulted
in a similar apoptotic rate. Of note, pre-treatement with LY294002
followed by incubation with CME led to the highest apoptotic rate.
All of these results indicated that CME induced apoptosis in HCT116
cells through the downregulation of Akt dephosphorylation.
To investigate whether CME treatment decreases the
dephosphorylation of Akt through the activation of AMPK or through
direct inhibition, the effects of Compound C, LY294002 alone or in
combination with CME on the levels of the upstream regulators
(p-AMPK and p-Akt) and downstream regulators (Bcl-2, Bax, Bak and
pro-caspase-3) were assessed by western blot analysis (Fig. 4A and B). When cells were treated
with CME alone or pre-treated with Compound C followed by
incubation with CME, the expression of p-Akt, p-mTOR, pro-caspase-3
and Bcl-2 was significantly decreased (P<0.001), whereas Bax and
Bak expression levels were significantly increased (P<0.001), as
compared with the control group. Although the expression of p-AMPK
was increased in the CME-treated group the Compound C and CME
co-treated groups, as compared with the control, a greater increase
was observed in the CME-treated group. LY294002 decreased the
expression of p-mTOR, p-Akt, procaspase-3 and Bcl-2, and increased
the expression of p-AMPK, Bax and Bak, as compared with the
control. These effects were aggravated when cells were pre-treated
with LY294002 and then incubated with CME.
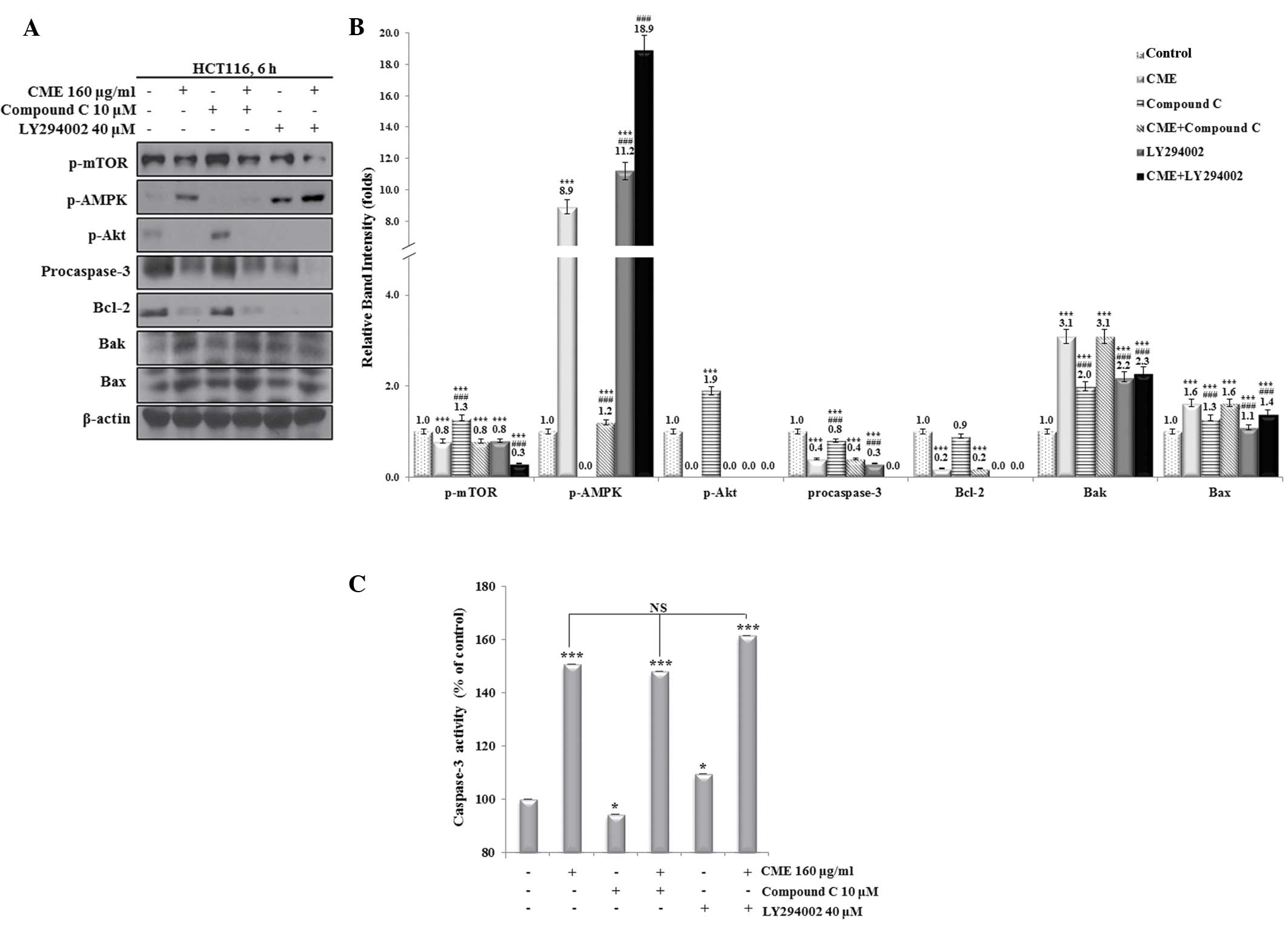 | Figure 4CME induces apoptosis through direct
regulation of the dephosphorylation of Akt. (A) Pre-treatment with
compound C or LY294002 prior to incubation with CME regulates
p-mTOR, p-AMPK and p-Akt in HCT116 colon cancer cells. Cells were
pre-treated with 10 µM compound C or 40 µM LY294002
for 30 min and then treated with 160 µg/ml CME for 6 h.
Protein levels were determined by western blot analysis and (B) the
relative band intensities were quantified. (C) Caspase-3 activity
in HCT116 colon cancer cells pre-treated with compound C or
LY294002 prior to incubation with CME. Caspase-3 activity was
measured using a Caspase-3 activity kit. Statistical analysis was
performed using an independent sample t-test. *P<0.05
and ***P<0.001 vs. the control; ###P<0.001 vs. the
CME-treated group, as determined by independent sample t-test (each
experiment, n=3). NS, not significant; p, phosphorylated; mTOR,
mammalian target of rapamycin; AMPK, adenosine
monosphosphate-activated protein kinase; PUMA, P53-upregulated
modulator of apoptosis; CME, Cnidium monnieri (L.) Cusson
extract; Bcl-2, B-cell lymphoma 2; Bak, Bcl-2-homologous antagonist
killer; Bax; Bcl-2-associated X protein. |
To determine whether the CME-induced apoptosis
proceeded via the mitochondrial pathway, a caspase-3 activity assay
was performed (Fig. 4C). Treatment
with Compound C alone decreased the caspase-3 activity, while
treatment with LY294002 alone increased the caspase-3 activity
compared with that in the control group. Caspase-3 activity was
significantly increased in the CME-treated and combined treatment
groups compared to those in the control group. Caspase-3 activity
was not significantly different between the CME-treated group and
the Compound C- or LY294002-pre-treatment + CME groups. However,
the group subjected to pre-treatment with LY294002 and CME showed a
slight increase in caspase-3 activation over that in the other
CME-treated groups. These results indicated that CME-induced
apoptosis of HCT116 colon cancer cells occurs through the
dephosphorylation of Akt and via an AMPK-independent pathway.
Discussion
Cancer is caused by the abnormal proliferation of
cells, and interference with cell proliferation as well as
induction of cancer cell death have been widely employed in the
treatment of cancer (6). Colon
cancer is of particular interest in South Korea, as its prevalence
has increased with the adaptation of Western eating habits
(1,2). Previous studies have shown that
various plant extracts have anti-cancer effects and cause apoptosis
of HCT116 colon cancer cells (4,26–28).
The present study investigated the apoptotic effects of an extract
of the fruit of Cnidium monnieri (L.) Cusson, based on
previous studies (23,25). Furthermore, it was confirmed that
CME induced apoptosis through downregulating Akt, and that
mitochondrial-mediated apoptotic proteins are involved via an
AMPK-independent pathway.
The present study first demonstrated the cytotoxic
effect of CME through the LDH release assay, as LDH release is a
common marker of cellular death (29). Cellular death occurs due to various
biological events, including DNA damage and metabolic stress, which
also activates pro-apoptotic proteins (8). The most important pro-apoptotic
proteins are Bax and Bak, which are translocated to the
mitochondria upon their activation (20). Activation of Bax and Bak leads to
the opening of the pore of the outer mitochondrial membrane, whichs
results in apoptosis through the release of LDH, cytochrome C and
other molecules into the cytosol (21). For these reasons, increased levels
of LDH release are indicative of cell death. The present study
found that CME treatment increased the release of LDH by colon
cancer cells in a dose-dependent manner.
A previous study found that curcumin treatment of
HCT116 colon cancer cells augmented the number of apoptotic bodies
in a dose-dependent manner (4).
The present study used Annexin V/PI and Hoechst 33342 staining to
quantify the occurrence of apoptosis and to observe the formation
of apoptotic bodies in HCT116 cells treated with CME. The results
showed that CME increased the number of Annexin V-positive,
apoptotic cells and induced apoptotic DNA fragmentation.
Apoptosis signaling is regulated by molecules
including AMPK and Akt in response to environmental changes
(5,8,12).
Cancer cells regulate the activation of several signaling
molecules, such as pro-survival factors, to stimulate cell growth
and proliferation, and to protect themselves (5,6). The
induction of apoptosis through the control of pro-survival and
pro-apoptotic factors been extensively studied (1,4,6).
Therefore, the present study investigated whether CME modulates
pro-survival factors and pro-apoptotic mitochondrial factors
through the regulation of upstream regulators, including AMPK and
Akt. According to a previous study, when HT-29 colon cancer cells
were treated with quercetin, the expression levels of p-AMPK, Bax,
and cleaved caspase-3 were increased, while the expression of Bcl-2
was decreased (5). In addition,
the activation of p53 by the inhibition of p-Akt induced an
increase in the expression of PUMA, which is a downstream protein
of p53, thereby inducing apoptosis through the inhibition of Bcl-2
and cleavage of procaspase-3 (4,30).
Caspase-3 is a downstream regulator of other caspases, such as
caspases-8 and 9, and can be activated by caspase-9 (15). Caspase-9, in turn, is activated
through cytochrome-C release due to the activation of Bax and Bak
(19,22). When HCT116 colon cancer cells were
treated with corosolic acid or Lepidium virginicum L.
extract, procaspase-3 and Bcl-2 expression were decreased and Bax
expression was increased (27,28).
Procaspase-3 expression was also shown to be decreased by curcumin
or Dorema glabrum seed extract (26,31).
The decline of procaspase-3 levels signifies an increase in
cleaved, activated caspase-3 levels and apoptosis (30). The present study showed that CME
treatment decreased the protein levels of p-Akt, p-mTOR,
procaspase-3 and Bcl-2, while increasing the levels of p-AMPK,
PUMA, Bax and Bak. In addition, the results clearly demonstrated
that CME treatment of HCT116 colon cancer cells activated
caspase-3. The present study found that CME regulated the
phosphorylation of AMPK and Akt as well as the expression of
apoptosis-associated proteins, including p-mTOR, PUMA, Bax and
Bak.
To confirm the role of AMPK and Akt in CME-induced
apoptosis, cells were pre-treated with AMPK and Akt inhibitors
prior to incubation with CME. Co-treatment with CME and Compound C
did not significantly affect the rate of LDH release compared to
that in the CME-treated group. However, in the LY294002 + CME
group, cell death was significantly increased compared with that in
the CME-treated group. The LY294002-treated group also increased
compared with than the control, although this difference was not
significant. Furthermore, Annexin V/PI staining and flow cytometric
analysis showed similar effects on the apoptotic rates. These
results indicated that CME exerts cytotoxic effects and induces
apoptosis via the downregulation of Akt.
p-AMPK and p-Akt are upstream regulators that
trigger apoptosis through modulation of individual signaling
pathways (2,12). To investigate whether CME
downregulates Akt through the activation of AMPK or through direct
inhibition, signaling proteins associated with AMPK- and
Akt-mediated apoptosis were assessed after treatment CME alone or
following pre-treatment with AMPK and Akt inhibitors. According to
the results, the expression levels of p-mTOR, p-Akt, pro-caspase-3,
Bcl-2, Bax and Bak were almost the same in the CME group and CME +
Compound C group. In addition, the protein levels of p-AMPK were
significantly higher in the CME group compared with those in the
CME + Compound C group. The levels of the signaling proteins
following treatment with LY294002 was similarly increased or
decreased compared to those following treatment with CME, and
combined treatment further enhanced these effects. A previous study
showed that AMPK suppresses Akt/mTOR signaling (32). However, in the present study,
combined treatment with Compound C and CME decreased the levels of
p-Akt compared to those in the groups treated with Compound C or
CME only. Therefore, the CME-induced suppression of p-Akt was not
caused by an increase of p-AMPK, but by the direct suppression of
the dephosphorylation of Akt. p-Akt suppresses the translocation of
Bax to the mitochondria, while LY294002 increases Bax translocation
(21). The present study showed
that Bax and Bak expression following various treatments was
increased compared to that in the control. LY294002 treatment led
to similar increases in Bax to those following CME treatment, and
the trend was further amplified following combined treatment of
LY294002 and CME. Activation of Bax and Bak induces caspase-3
activity (5). Therefore, the
present study performed a caspase-3 activity assay to determine
whether CME-induced apoptosis proceeded via the mitochondrial
pathway. There were no significant differences in caspase-3
activity between the group co-treated with Compound C and CME and
that treated with CME alone. This also supported the finding that
CME induced apoptosis through an AMPK-independent pathway. A
previous study has shown that LY294002 treatment is capable of
increasing caspase-3 activation in human umbilical vein endothelial
cells (33). In the present study,
LY294002 treatment of HCT116 colon cancer cells also significantly
increased caspase-3 activity compared to that in the control group.
In addition, in the LY294002 + CME group, caspase activation was
slightly, but not significantly increased compared to that in the
CME-treated group.
The present study was the first to report that CME
induced apoptosis by exerting metabolic stress through direct
regulation of p-Akt via an AMPK-independent pathway in HCT116 colon
cancer cells. CME directly downregulated Akt, which suppressed
Bcl-2, activated Bax and Bak, and led to apoptosis by caspase-3
activation and possibly through cytochrome C release.
Acknowledgments
The present study was supported by the Korea
Research Foundation Grant (grant no. KRF-2010-0021402) and the
Technological Innovation R&D Program (grant no. S2128176)
funded by the Small and Medium Business Administration (SMBA,
Korea).
References
|
1
|
Park SY, Kim IS, Lee SH, Lee SH, Jung DW,
Park OJ and Kim YM: Anti-proliferative effects of selenium in HT-29
colon cancer cells via inhibition of Akt. J Life Sci. 22:55–61.
2012. View Article : Google Scholar
|
|
2
|
Lee SH, Park SY, Kim IS, Park OJ and Kim
YM: Effects of resveratrol on migration and proliferation in HT-29
colon cancer cells. KSBB J. 27:289–294. 2012. View Article : Google Scholar
|
|
3
|
Grady WM and Markowitz SD: Genetic and
epigenetic alterations in colon cancer. Annu Rev Genomics Hum
Genet. 3:101–128. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Park SY, Lee SH, Park OJ and Kim YM:
Apoptotic effects of curcumin and EGCG via Akt-p53 signaling
pathway in HCT116 colon cancer cells. J Life Sci. 21:89–95. 2011.
View Article : Google Scholar
|
|
5
|
Lee SH, Jung DW, Kim GT, Park SY, Kim SY,
Park OJ and Kim YM: Quercetin of plants extracts regulates sestrin2
and induces apoptosis in HT-29 colon cancer cells. Cancer Prev Res.
17:244–250. 2012.
|
|
6
|
Lee SH, Kim GT, Kim JI, Lim EG, Kim IS and
Kim YM: The Extract from Lysimachia foenum-graecum induces
apoptosis in MCF-7 breast cancer cells. KSBB J. 28:303–309. 2013.
View Article : Google Scholar
|
|
7
|
Lee YK, Park SY, Kim YM, Kim DC, Lee WS,
Surh YJ and Park OJ: Suppression of mTOR via Akt-dependent and
-independent mechanisms in selenium-treated colon cancer cells:
Involvement of AMPKalpha1. Carcinogenesis. 31:1092–1099. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Park C, Jin CY, Choi TH, Hong SH and Choi
YH: Effect on proapoptotic Bcl-2 on naringenin-induced apoptosis in
human leukemia U937 cells. J Life Sci. 23:1118–1125. 2013.
View Article : Google Scholar
|
|
9
|
Park SY, Lee SH, Park OJ and Kim YM:
Apoptotic effects of selenium via AMPK-VASP signal pathway in
B16F10 melanoma cells. Cancer Prev Res. 15:313–319. 2010.
|
|
10
|
Seo BR, Min KJ, Cho IJ, Kim SC and Kwon
TK: Curcumin significantly enhances dual PI3K/Akt and mTOR
inhibitor NVP-BEZ235-induced apoptosis in human renal carcinoma
caki cells through down-regulation of p53-dependent Bcl-2
expression and inhibition of Mcl-1 protein stability. PLoS One.
9:e95582014. View Article : Google Scholar
|
|
11
|
Greer EL, Oskoui PR, Banko MR, Maniar JM,
Gygi MP, Gygi SP and Brunet A: The energy sensor AMP-activated
protein kinase directly regulates the mammalian FOXO3 transcription
factor. J Biol Chem. 282:30107–30119. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Kim D and Chung J: Akt: Versatile mediator
of cell survival and beyond. J Biochem Mol Biol. 35:106–115. 2002.
View Article : Google Scholar
|
|
13
|
Cory S, Huang DC and Adams JM: The Bcl-2
family: Roles in cell survival and oncogenesis. Oncogene.
22:8590–8607. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Lavi I, Friesem D, Geresh S, Hadar Y and
Schwartz B: An aqueous polysaccharide extract from the edible
mushroom Pleurotus ostreatus induces anti-proliferative and
pro-apoptotic effects on HT-29 colon cancer cells. Cancer Lett.
61–70. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Deng YB, Lin YH and Wu XW: TRAIL-induced
apoptosis requires Bax-dependent mitochondrial release of
Smac/DIABLO. Genes Dev. 16:33–45. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Jiang M, Wang CY, Huang S, Yang T and Dong
Z: Cisplatin-induced apoptosis in p53-deficient renal cells via the
intrinsic mitochondrial pathway. Am J Physiol Renal Physiol.
296:F983–F993. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Zhuang S, Yan Y, Daubert RA, Han J and
Schnellmann RG: ERK promotes hydrogen peroxide-induced apoptosis
through caspase-3 activation and inhibition of Akt in renal
epithelial cells. Am J Physiol Renal Physiol. 292:F440–F447. 2007.
View Article : Google Scholar
|
|
18
|
Jeong SY and Seol DW: The role of
mitochondria in apoptosis. BMB Rep. 41:11–22. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Chien SY, Wu YC, Chung JG, Yang JS, Lu HF,
Tsou MF, Wood WG, Kuo SJ and Chen DR: Quercetin-induced apoptosis
acts through mitochondrial- and caspase-3-dependent pathways in
human breast cancer MDA-MB-231 cells. Hum Exp Toxicol. 28:493–503.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Degli Esposti M and Dive C: Mitochondrial
membrane permeabilisation by Bax/Bak. Biochem Biophys Res Commun.
304:455–461. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Tsuruta F, Masuyama N and Gotoh Y: The
phosphatidylinositol 3-kinase (PI3K)-Akt pathway suppresses bax
translocation to mitochondria. J Biol Chem. 277:14040–14047. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Twiddy D and Cain K: Caspase-9 cleavage,
do you need it? Biochem J. 405:e1–2. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Yang LL, Wang MC, Chen LG and Wang CC:
Cytotoxic activity of coumarins from the fruits of Cnidium monnieri
on leukemia cell lines. Planta Med. 69:1091–1095. 2003. View Article : Google Scholar
|
|
24
|
Zhu YP: Tonifying herbs. Chinese Material
Medica: Chemistry, Pharmacology And Applications. Harwood Academic
Publishers; Amsterdam: pp. 6241998
|
|
25
|
Chou SY, Hsu CS, Wang KT, Wang MC and Wang
CC: Antitumor effects of osthol from Cnidium monnieri: An in vitro
and in vivo study. Phytother Res. 21:226–230. 2007. View Article : Google Scholar
|
|
26
|
Watson JL, Hill R, Lee PW, Giacomantonio
CA and Hoskin DW: Curcumin induces apoptosis in HCT-116 human colon
cancer cells in a p21-independent manner. Exp Mol Pathol.
84:230–233. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Sung B, Kang YJ, Kim DH, Hwang SY, Lee YJ,
Kim MJ, Yoon JH, Kim CM, Chung HY and Kim ND: Corosolic acid
induces apoptotic cell death in HCT116 human colon cancer cells
through a caspase-dependent pathway. Int J Mol Med. 33:943–949.
2014.PubMed/NCBI
|
|
28
|
Chae YH, Shin DY, Park C, Lee YT, Moon SG
and Choi YH: Induction of apoptosis in human colon carcinoma HCT116
cells using a water extract of Lepidium virginicum L. J Korean Soc
Food Sci Nutr. 40:649–659. 2011. View Article : Google Scholar
|
|
29
|
Moran JH and Schnellmann RG: A rapid
beta-NADH-linked fluorescence assay for lactate dehydrogenase in
cellular death. J Pharmacol Toxicol Methods. 36:41–44. 1996.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Lee WK and Kim SJ: Sulforaphane-induced
apoptosis was regulated by p53 and Caspase-3 dependent pathway in
human chondrosarcoma, HTB-94. J Life Sci. 21:851–857. 2011.
View Article : Google Scholar
|
|
31
|
Bannazadeh Amirkhiz M, Rashtchizadeh N,
Nazemiyeh H, Abdolalizadeh J, Mohammadnejad L and Baradaran B:
Investigating apoptotic effects of methanolic extract of Dorema
glabrum seed on WEHI-164 cells. ISRN Pharmacol. 2013:9498712013.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Pantovic A, Krsti A, Janjetovica K, Kocic
J, Harhaji-Trajkovic L, Bugarski D and Trajkovi V: Coordinated
time-dependent modulation of AMPK/Akt/mTOR signaling and autophagy
controls osteogenic differentiation of human mesenchymal stem
cells. Bone. 52:524–531. 2013. View Article : Google Scholar
|
|
33
|
Shi H and Feng JM: Aristolochic acid
induces apoptosis of human umbilical vein endothelial cells in
vitro by suppressing PI3K/Akt signaling pathway. Acta
Pharmacologica Sinica. 32:1025–1030. 2011. View Article : Google Scholar : PubMed/NCBI
|