Introduction
Platelets are anucleated blood cells released from
bone marrow megakaryocytes through a complex process that is not
fully understood (1,2). At sites of vascular injury, platelets
come into contact with exposed subendothelial components and form a
plug to prevent excessive blood loss. However, if this process is
not controlled, it may result in thrombotic events causing severe,
life-threatening diseases, including myocardial infarction or
ischemic stroke (3,4). Based on the above evidence,
thrombosis is almost exclusively treated by anti-platelet and
anti-coagulant compounds, which reduce thrombosis propagation.
However, current anti-platelet and anticoagulant drugs always have
poor efficacy and associated side effects, these limitations render
it imperative to develop more effective drugs with reduced side
effects (5–8).
A previous study by Northeast et al (9) using a rat model of venous thrombosis
demonstrated that during natural resolution of venous thrombi, the
activity of plasminogen activator urokinase (uPA) is upregulated
(9). Subsequent gene level studies
demonstrated that deletion of the gene encoding uPA markedly
inhibited normal thrombosis resolution (10), while the adenoviral expression of
uPA enhanced venous thrombosis resolution (11). These noteworthy results suggest
there may be clinical potential in upregulating the expression of
uPA in thrombosis resolution. uPA is a serine protease that is
important for cell migration and angiogenesis (12,13).
Several previous studies demonstrated that the secretion of uPA was
increased by the overexpression of mitogen-activated protein kinase
(MAPK) 14 (p38) and MAPK8 (JNK) (14–16),
which are important components of the MAPK signaling pathway.
Activation of the pathway is associated with the activation of
various transcription factors, resulting in the increased
expression of numerous genes involved in tumor cell proliferation,
apoptosis and angiogenesis (15).
Thus, it is possible that the p38 and JNK MAPK pathways participate
in the process of uPA-regulated thrombosis recanalization, and that
agents with an effect on this pathway may improve thrombosis
treatment.
The herb Paeonia lactiflora pall, which is
known as 'Shao Yao' in Chinese, has been used in traditional
Chinese medicine for over 1,000 years to treat cramp, pain,
giddiness and congestion (17).
Paeoniflorin, the major component of Paeonia lactiflora
pall, has previously been reported to exhibit various
pharmacological effects, including anti-inflammation, anti-allergy,
anti-hyperglycemia, enhanced cognition and thrombosis prevention
(18). Furthermore, paeoniflorin
was also reported to have an effect on MAPK pathways (18). These previous studies suggest that
paeoniflorin may regulate uPA to promote recanalization following
thrombosis.
To verify the hypothesis, the current study measured
the concentration of fibronectin (FN), fibrinogen (FIB), D-dimer
(D-D), 6-keto prostaglandin F1a
(6-Keto-PGF1a), thromboxane B2
(TXB2) and uPA in the serum of spontaneously
hypertensive rats (SHRs) using a sandwich enzyme-linked
immunosorbent assay (ELISA) to confirm the effect of paeoniflorin
on thrombosis recanalization. The cytotoxicity of paeoniflorin on
human umbilical vein endothelial cells (HUVECs) was estimated by
3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT)
assay and the potential link between paeoniflorin and uPA was
evaluated using western blot analysis. The present study aimed to
determine the mechanism by which the effects of paeoniflorin on
thrombosis are mediated and to improve the practical use of this
Chinese medicine compound in the clinic. The results of the present
study demonstrated the preliminary mechanism of paeoniflorin to
enhance the expression of uPA to recanalize thrombosis, which may
facilitate uPA as a potential therapeutic strategy against
thromobosis in the clinic.
Materials and methods
Materials
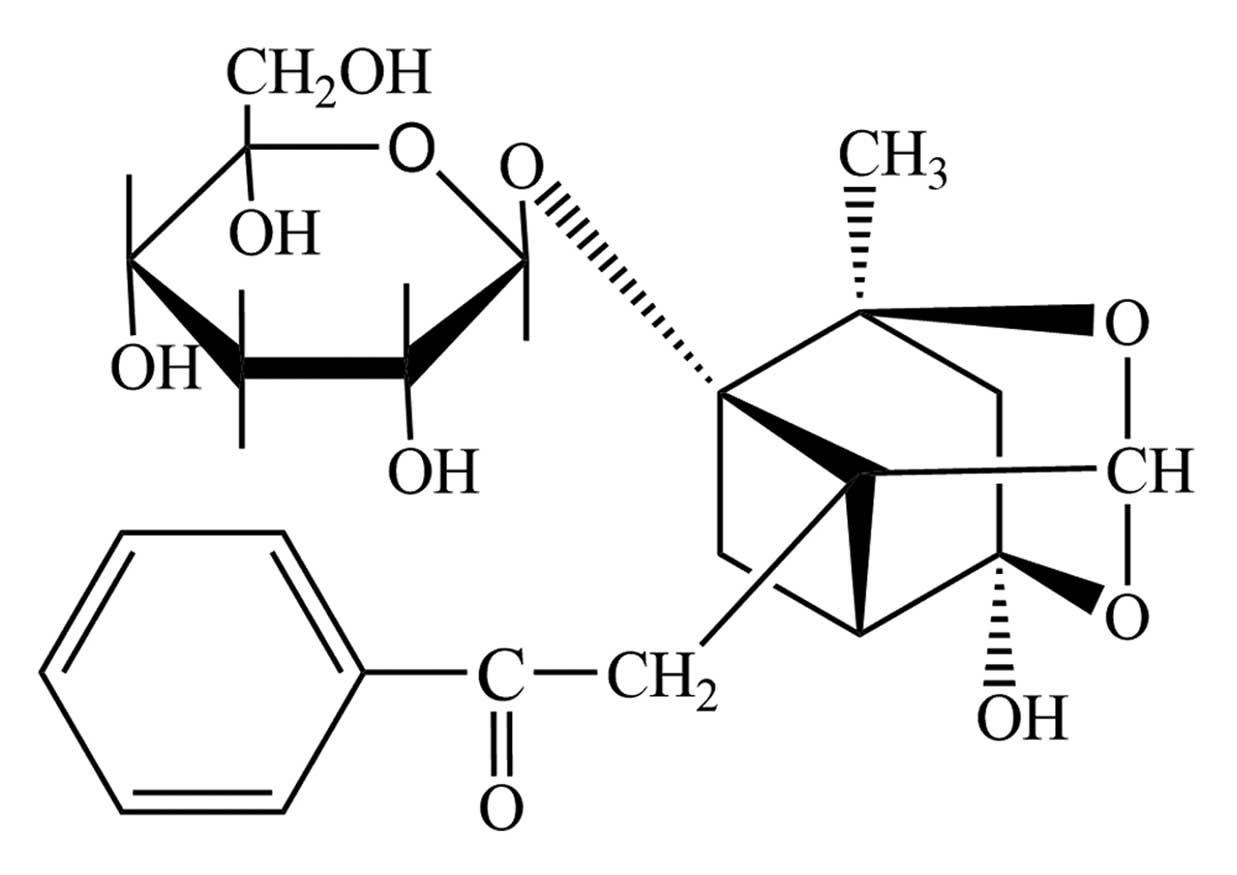
Paeoniflorin (purity, >99%; Fig. 1) was obtained from Sigma-Aldrich
(St. Louis, MO, USA). Antibodies obtained for use in the present
study are as follows: Rabbit monoclonal anti-JNK (cat. no.
ab110724), rabbit polyclonal anti-phosphorylated (p)-JNK (cat. no.
ab47337), rabbit monoclonal anti-p38 (cat. no. ab170099), rabbit
polyclonal anti-p-p38 (cat. no. ab47363), rabbit monoclonal
anti-uPA (cat. no. ab133563) and rabbit polyclonal
anti-glyceraldehyde 3-phosphate dehydrogenase (GAPDH; cat. no.
ab9485) were purchased from Abcam (Cambridge, MA, USA). JNK
inhibitor (SP600125) and p38 inhibitor (SB203580) were obtained
from EMD Millipore (Billerica, MA, USA). The HUVEC line was
purchased from the American Type Culture Collection (Manassas, VA,
USA). The cells were cultured in medium 199 (Gibco; Thermo Fisher
Scientific, Inc., Waltham, MA, USA) supplemented with 20% fetal
bovine serum (Gibco; Thermo Fisher Scientific, Inc.), 2 mM
L-glutamine (Sigma-Aldrich), 5 U/ml heparin (Sigma-Aldrich), 100
IU/ml penicillin (Sigma-Aldrich), 10 μg/ml streptomycin
(Sigma-Aldrich) and 50 μg/ml endothelial cell growth
supplement (American Type Culture Collection). Newly purchased
cells were cultured in a humidified incubator at 5% CO2
and 37°C. Cells between passage 3 and 6 were used for further
experiments.
Grouping of model animals and gavage
experiment
Male SHRs (n=30; age 18 weeks; weight, 350–400 g)
were provided by the Laboratory Animal Center of the First
Affiliated Hospital of Sun Yat-Sen University (Guangzhou, China)
for the thrombosis model and were housed in cages at room
temperature (20–25°C) and 80% humidity, with a 12-h light/dark
cycle. Food and water were available ad libitum. The animals
were randomly divided into control, paeoniflorin and aspirin groups
(n=10 per group). The control, paeoniflorin and aspirin group
animals received 2 ml/kg normal saline, 5 mg/kg paeoniflorin and 50
mg/kg aspirin (Sigma-Aldrich), respectively, every 2 days for 2
weeks by gavage method. All the animal experiments were conducted
in accordance with the Guide for the Care and Use of Laboratory
Animals published by the National Institutes of Health (19), and the present study was approved
by the ethics committee of Nanyang Institute of Technology
(Nanyang, Henan).
Collection of serum samples and
ELISA
Each rat was anesthetized with 10% chloral hydrate
(0.3 ml/100 g; Sigma-Aldrich) at the end of the treatment by
intraperitoneal injection, the rats were sacrificed and blood
samples were collected from the abdominal aorta by opening the
abdominal cavity. Serum was separated by centrifugation at 2,200 ×
g at 4°C for 20 min and preserved at −80°C. For each serum sample,
the concentration of FN, FIB, D-D, 6-Keto-PGF1a,
TXB2 and uPA, which were all closely associated with the
prothrombotic state (PTS), were measured using ELISA kits according
to the manufacturer's instructions (Nanjing Jiancheng
Bioengineering Institute, Nanjing, China). Briefly, plates were
coated and incubated overnight with 3.4 mg/ml non-biotinylated 3D5
(100 μl/well; Nanjing Jiancheng Bioengineering Institute) in
200 mM NaHCO3 at pH 9.6 at 4°C, and then washed 4 times
with phosphate-buffered saline (PBS) containing 0.05% Tween 20
(PBST). Following incubation with blocking buffer (150
μl/well PBST containing 2.5% gelatin) for 2 h at 37°C, the
plates were again washed 4 times with PBST, and a 100 μl
serum sample (diluted 1:1 with PBS) was added to each well. The
plates were then incubated at 37°C for 2 h. After washing 4 times
with PBST, 100 μl of 1 μg/ml biotinylated 3D5 in
blocking buffer was added to each well and incubated at 37°C for
another 2 h. The wells were then washed 4 times with PBST and
incubated with 100 μl/well ExtrAvidin-Alkaline phosphatase
(Sigma-Aldrich) in blocking buffer (1:5,000) and incubated for 1 h
at 37°C. Following another 4 washes with PBST, the enzyme substrate
Yellow 'pNPP' (100 μl/well; Sigma-Aldrich) was added to each
well and incubated for 30 min at 37°C for color development.
Optical density (OD) values of different targeted molecules were
recorded with a microplate reader at a wavelength of 450 nm (Model
550; Bio-Rad Laboratories, Inc., Hercules, CA, USA). The
concentration of each protein was estimated from the standard curve
determined by serial dilution.
Cytotoxicity of paeoniflorin and cell
proliferation assay
To determine the cytotoxic effect of paeoniflorin on
HUVECs, a 3-(4,5dimethylthiazol-2-yl)-2,5-diphenyltetrazolium
bromide (MTT) assay was performed according to the standard
protocol. Briefly, 50 μl exponentially growing HUVECs
(2×105 cells/ml) were seeded into a 96-well plate in
triplicate. The cells were treated with 0 (control), 0.01, 0.05,
0.1, 0.5, 1, 5, 10, 20 and 50 μmol/l paeoniflorin for 24 h.
Following incubation with paeoniflorin, 5 mg/ml MTT (Sigma-Aldrich)
was added to each well and incubated for another 4 h at 37°C. The
OD values in different wells were recorded using a microplate
reader at 450 nm. The survival rates (%) of cells with the
different treatments were calculated as: (OD value in treatment
group - OD value in blank control group)/(OD value in negative
control group - OD value in blank control group) × 100.
The highest concentration with the majority cells
surviving was 0.5 μmol/l, thus, the cell proliferation assay
was conducted using this concentration. HUVECs were treated with 5
μmol/l paeoniflorin for 0, 24, 48, 72 and 96 h. Normal
HUVECs were used as a control to determine the effect of
paeoniflorin on the viability of HUVECs over the time course. The
detection of cell proliferation was conducted as described above.
The OD values in different wells were recorded using the microplate
reader at 450 nm.
Western blot analysis
HUVECs were divided into 4 treatment groups:
Untreated; incubated with 0.5 μmol/l paeoniflorin for 24 h;
pretreated with 50 μmol/l SB203580 for 1 h and incubated
with 0.5 μmol/l paeoniflorin for 24 h; and pretreated with
50 μmol/l SP600125 for 1 h and incubated with 0.5
μmol/l paeoniflorin for 24 h. The protein samples were
extracted from the cells after the 24-h incubation using SDS Lysis
Buffer on ice for 30 min. The protein concentration was determined
using the Pierce BCA Protein assay kit (Thermo Fisher Scientific,
Inc.). All the extracts were boiled with loading buffer for 5 min
prior to sodium dodecyl sulfate polyacrylamide gel electrophoresis
(SDS-PAGE) on 10% gels (EMD Millipore). Proteins then were
transferred onto polyvinylidene difluoride membranes (EMD
Millipore). The membranes were washed with Tris-buffered
saline-0.05% Tween 20 (Sigma-Aldrich) three times for 20 min each
time. The membranes were blocked with 5% non-fat milk prior to
incubation with specific antibodies [JNK (1:1,000), p-JNK (1:800),
p38 (1:800), p-p38 (1:1,000), uPA (1:1,000) and GAPDH (1:2,000)]
overnight at room temperature. Following three additional washes,
horseradish peroxidase-conjugated monoclonal goat anti-rabbit
secondary antibodies (1:1,500; Beyotime Institute of Biotechnology;
cat. no. A2080) were added and the membranes were incubated for
another 4 h. Following 3 final washes, the blots were developed
using Beyo ECL Plus reagent (Beyotime Institute of Biotechnology)
and the results were detected in the Chemi Doc™ MP system (Bio-Rad
Laboratories, Inc.). GAPDH was used as a loading control for
western blot analysis.
Statistical analysis
Data are expressed as the mean ± standard deviation.
Multiple comparisons were conducted using one-way analysis of
variance with least significant difference test method. Statistical
analysis was conducted using SPSS software version 19.0 (IBM SPSS,
Armonk, NY, USA). P<0.05 was considered to indicate a
statistically significant difference.
Results
Paeoniflorin improves the PTS and
increases uPA expression in SHRs
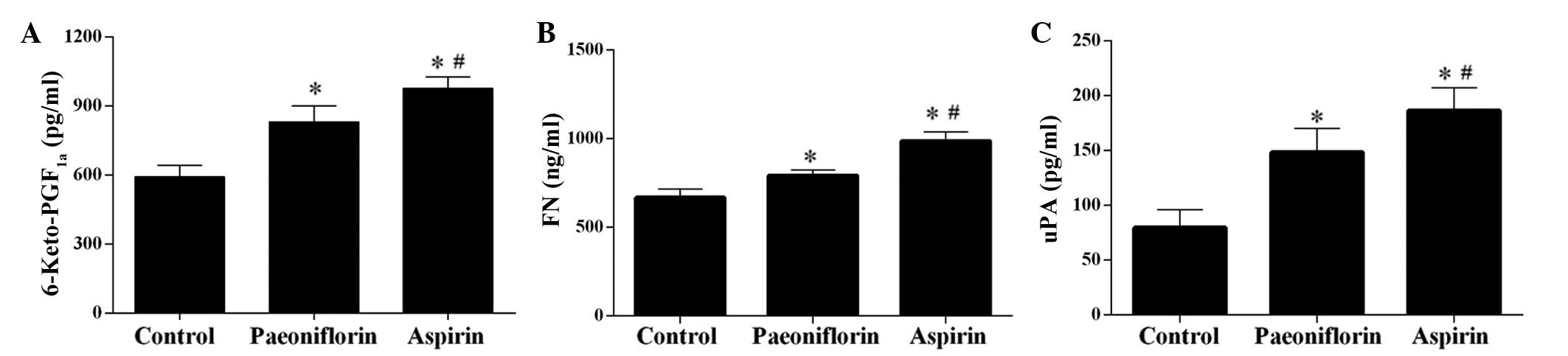
The ELISA results demonstrated that treatment with
paeoniflorin significantly increased the concentration of
6-Keto-PGF1a, FN and uPA in serum samples (P<0.05),
however, the effect was weaker than that of aspirin, which served
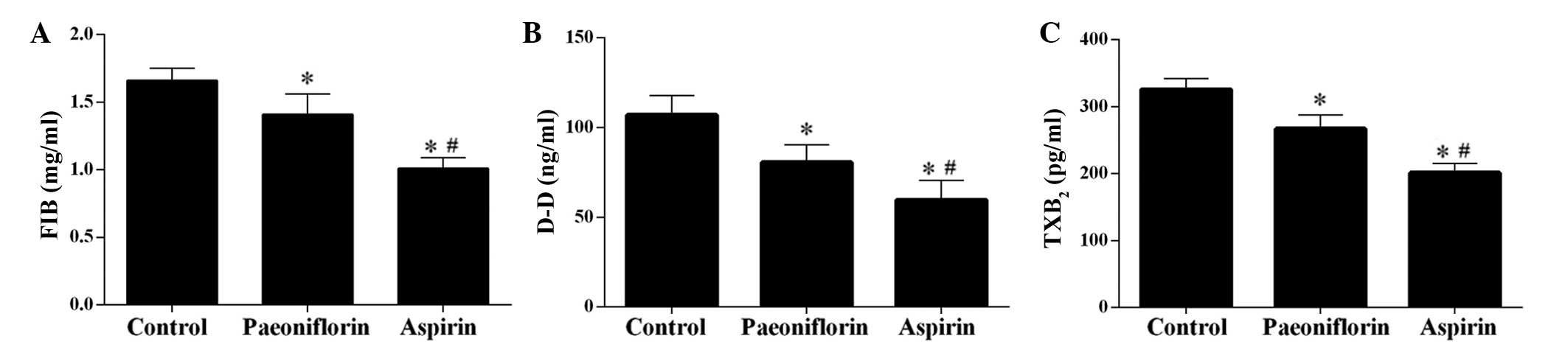
as a control as it is in anti-thrombotic drug (Fig. 2) (20). Furthermore, the levels of FIB, D-D
and TXB2 were significantly downregulated by
paeoniflorin, however, treatment with aspirin exerted a more potent
effect (Fig. 3).
Effect of paeoniflorin on HUVEC viability
and proliferation
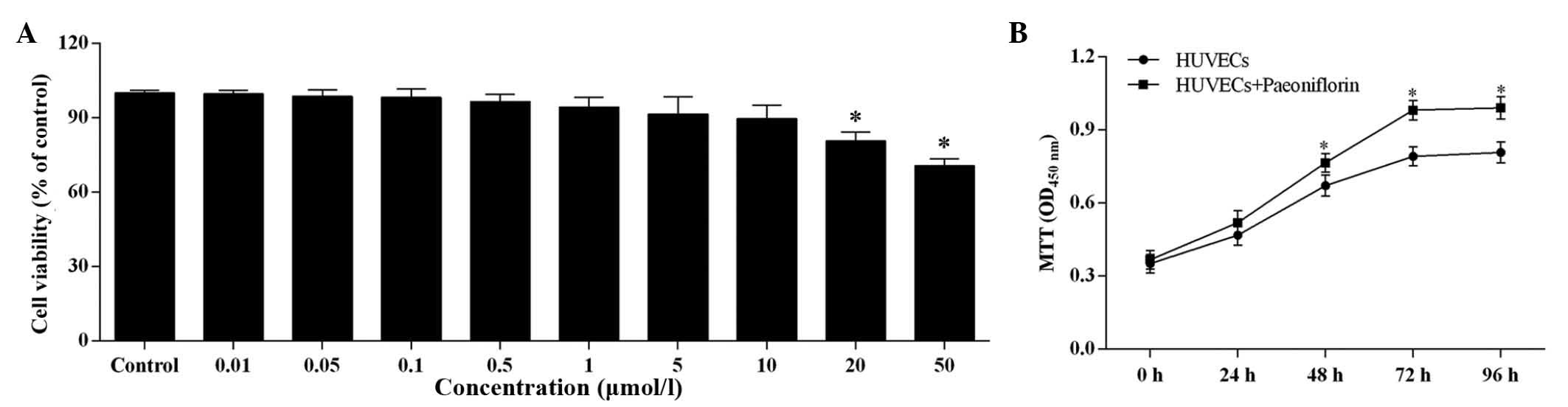
The increasing paeoniflorin concentration had a
negative effect on the cell viability of HUVECs (Fig. 4A). There was no significant
difference in cell viability compared with control when the
paeoniflorin concentration was <0.5 μmol/l, however, when
the concentration was >0.5 μmol/l, the cell viability was
significantly reduced. However, even at 50 μmol/l
paeoniflorin, the cell viability remained within the safe range
(>70%).
Based on the results of the MTT assay, 0.5
μmol/l was determined as the suitable paeoniflorin
concentration for use in further experiments. At this
concentration, paeoniflorin significantly increased cell
proliferation compared with untreated HUVECs after 48, 72 and 96-h
treatment (P<0.05; Fig.
4B).
Treatment of paeoniflorin activates the
phosphorylation of JNK and p38, and upregulates the expression of
uPA
To examine the effect of paeoniflorin on the MAPK
pathway and the subsequent influence on the expression of uPA, the
current study used western blot analysis to quantify the protein
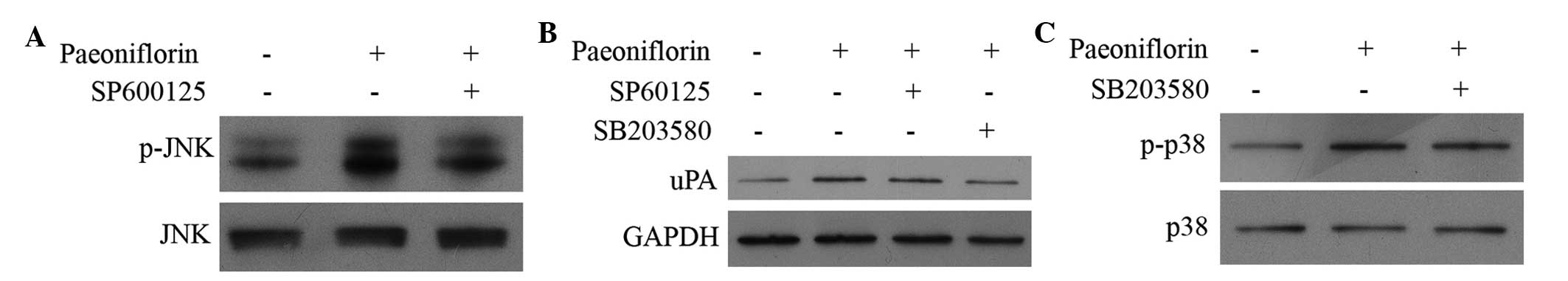
expression levels of JNK, p-JNK, p38, p-p38 and uPA. As
demonstrated in Fig. 5, the
expression levels of p-JNK, p-p38 and uPA in HUVECs were
upregulated by paeoniflorin treatment compared with control.
Treatment with SB203580 (p38 inhibitor) and SP600125 (JNK
inhibitor) demonstrated direct effects on the MAPK pathway and
synthesis of uPA. Treatment with paeoniflorin activates the
phosphorylation of JNK and p38 and upregulates the expression of
uPA (Fig. 5). Treatment with
SP600125, a JNK inhibitor, and SB203580, a p38 inhibitor, reduced
the phosphorylation of JNK and p38, and inhibited the expression of
uPA compared with paeoniflorin treatment (Fig. 5B and C).
Discussion
A previous study demonstrated that monocyte ingress
in thrombosis resolution is dependent on the expression of uPA
(21). It is hypothesized that uPA
promotes thrombosis resolution by assisting monocyte recruitment to
the thrombus; thus, facilitating revascularization and remodeling
(22). However, application of uPA
using a single adenoviral dose may not be an optimum treatment due
to the larger volume of thrombi found in human veins, and the
delivery of greater doses is currently expensive. In order to
address this problem, a more effective application of uPA to
recanalize thrombosis is required. Considering that the expression
of uPA is regulated through MAPK pathways, the current study used
agents that inhibit the pathway to investigate their effect on the
associated processes.
The present study examined the effect of
paeoniflorin, a compound used in Chinese medicine, on thrombosis
and the expression of uPA. The results demonstrated that
paeoniflorin improves the PTS in rat models of thrombosis and
upregulates the expression of uPA via the MAPK signaling pathways.
Ye et al (23) previously
reported the antithrombotic effect of paeoniflorin, however, no
further studies have been conducted to elucidate the underlying
mechanism of this effect. In the current study, ELISA results
demonstrated that the concentration of 6-Keto-PGF1a and
FN in serum samples was significantly increased, whereas the levels
of FIB, TXB2, and DD were significantly reduced.
6-Keto-PGF1a is the stable metabolite of prostaglandin
I2. FN is important for stabilizing extracellular matrix
material, and measuring the levels of FN may be used to assess
endothelial damage (24,25). FIB has previously been associated
with an increased risk of venous thrombosis (26). TXB2 indicates activation
of platelets and a decreased D-D level is hypothesized to reflect
reduced secondary fibrinolytic activity (27,28).
All the changes to these molecules observed in the present study
were indicated an improvement of thrombosis following paeoniflorin
treatment. Although the effect of paeoniflorin was not as potent as
aspirin, the practical concentration of paeoniflorin used in the
clinic could be adjusted to a level which may be comparable to or
stronger than that of aspirin given the low cytotoxicity of
paeoniflorin demonstrated by the MTT assay. To achieve this,
further comprehensive animal and clinical studies are planned to be
conducted in the future. The current study measured the cytotoxic
effect of paeoniflorin on HUVECs by an MTT assay. It was
demonstrated that the proliferative ability of the cells was
increased by paeoniflorin with low cytotoxicity. Although
paeoniflorin reduced cell viability in a concentration-dependent
manner when the concentration was >0.5 μmol/l, cell
viability of HUVECs remained >70%.
The effect of paeoniflorin on the MAPK signaling
pathway has been previously reported. Guo et al (18) demonstrated that chronic treatment
with paeoniflorin suppresses the activation of the MAPK pathway,
particularly p38 and JNK, and may reduce the production of
pro-inflammatory molecules and exert neuroprotective effects during
stroke. Although the study by Guo et al (18) and the current study investigated
the effects of paeoniflorin on the MAPK pathway, the results
demonstrated were quite different. In the present study, the effect
of paeoniflorin on MAPK signaling was positive; the phosphorylation
of p38 and JNK were increased. The paradox may be caused by the
differing concentrations used in the studies. The rats in the
previous study received 5 mg/kg paeoniflorin twice per day for 2
weeks, whereas in the present study, HUVECs received 0.5
μmol/l paeoniflorin for 24 h. This information is important
as it may be used to determine the treatment concentration of
paeoniflorin if used in the clinic. The dose of paeoniflorin used
to treat different diseases should be carefully considered to avoid
a negative effect. However, the treatment with JNK and p38
inhibitors exhibited complex effects on the MAPK signaling pathway
and expression of uPA. Treatment with SP600125 reduced the
phosphorylation of JNK and inhibited the expression of uPA.
SB203580, a p38 inhibitor, blocked the phosphorylation of p38 and
exhibited an inhibitory effect on the expression level of uPA.
These results indicate that the regulation of uPA by paeoniflorin
may be dependent on other factors and future comprehensive studies
should be conducted to determine the pathways associated with uPA
regulation.
In conclusion, paeoniflorin improves the PTS and
upregulates uPA via the MAPK signaling pathway. However, this
positive effect is dependent on the concentration of paeoniflorin
used in the experiment. Different concentrations of paeoniflorin
may improve certain diseases and exert a negative effect on others.
The results of the present study preliminarily demonstrated that
paeoniflorin acts by enhancing the expression of uPA to increase
recanlization following thrombosis, which may facilitate the
uPA-dependent treatment of thrombosis in the clinic.
References
|
1
|
Italiano JE, Patel-Hett S and Hartwig JH:
Mechanics of proplatelet elaboration. J Thromb Haemost. 5(Suppl 1):
S18–S23. 2007. View Article : Google Scholar
|
|
2
|
Bender M, Eckly A, Hartwig JH, Elvers M,
Pleines I, Gupta S, Krohne G, Jeanclos E, Gohla A, Gurniak C, et
al: ADF/n-cofilin-dependent actin turnover determines platelet
formation and sizing. Blood. 116:1767–1775. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Ruggeri ZM: Platelets in atherothrombosis.
Nature Med. 8:1227–1234. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Stoll G, Kleinschnitz C and Nieswandt B:
Molecular mechanisms of thrombus formation in ischemic stroke:
Novel insights and targets for treatment. Blood. 112:3555–3562.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Schrör K: Antiplatelet drugs. A
comparative review. Drugs. 50:7–28. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Ni H and Freedman J: Platelets in
hemostasis and thrombosis: Role of integrins and their ligands.
Transfus Apher Sci. 28:257–264. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Barrett NE, Holbrook L, Jones S, Kaiser
WJ, Moraes LA, Rana R, Sage T, Stanley RG, Tucker KL, Wright B and
Gibbins JM: Future innovations in anti-platelet therapies. Br J
Pharmacol. 154:918–939. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Bird JE, Giancarli MR, Allegretto N,
Barbera F, Wong P, Schumacher WA, Ogletree ML and Seiffert D:
Prediction of the therapeutic index of marketed anti-coagulants and
anti-platelet agents by guinea pig models of thrombosis and
hemostasis. Thromb Res. 123:146–158. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Northeast AD, Soo KS, Bobrow LG, Gaffney
PJ and Burnand KG: The tissue plasminogen activator and urokinase
response in vivo during natural resolution of venous thrombus. J
Vasc Surg. 22:573–579. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Singh I, Burnand KG, Collins M, Luttun A,
Collen D, Boel-houwer B and Smith A: Failure of thrombus to resolve
in urokinase-type plasminogen activator gene-knockout mice: Rescue
by normal bone marrow-derived cells. Circulation. 107:869–875.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Gossage JA, Humphries J, Modarai B,
Burnand KG and Smith A: Adenoviral urokinase-type plasminogen
activator (uPA) gene transfer enhances venous thrombus resolution.
J Vasc Surg. 44:1085–1090. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Aguirre Ghiso JA, Alonso DF, Farías EF,
Gomez DE and Bal de Kier Joffè E: Deregulation of the signaling
pathways controlling urokinase production. Its relationship with
the invasive phenotype. Eur J Biochem. 263:295–304. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Blasi F and Carmeliet P: uPAR: A versatile
signalling orchestrator. Nat Rev Mol Cell Biol. 3:932–943. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Lee KH, Hyun MS and Kim JR: Growth
factor-dependent activation of the MAPK pathway in human pancreatic
cancer: MEK/ERK and p38 MAP kinase interaction in uPA synthesis.
Clin Exp Metastasis. 20:499–505. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Liu SQ, Huang JA, Qin MB, Su YJ, Lai MY,
Jiang HX and Tang GD: Sphingosine kinase 1 enhances colon cancer
cell proliferation and invasion by upregulating the production of
MMP-2/9 and uPA via MAPK pathways. Int J Colorectal Dis.
27:1569–1578. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Benasciutti E, Pagès G, Kenzior O, Folk W,
Blasi F and Crippa MP: MAPK and JNK transduction pathways can
phosphorylate Sp1 to activate the uPA minimal promoter element and
endogenous gene transcription. Blood. 104:256–262. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Nizamutdinova IT, Jin YC, Kim JS, Yean MH,
Kang SS, Kim YS, Lee JH, Seo HG, Kim HJ and Chang KC: Paeonol and
paeoniflorin, the main active principles of Paeonia albiflora,
protect the heart from myocardial ischemia/reperfusion injury in
rats. Planta medica. 74:14–18. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Guo RB, Wang GF, Zhao AP, Gu J, Sun XL and
Hu G: Paeoni-florin protects against ischemia-induced brain damages
in rats via inhibiting MAPKs/NF-κB-mediated inflammatory responses.
PLoS One. 7:e497012012. View Article : Google Scholar
|
|
19
|
National Research Council: Guide for the
Care and Use of Laboratory Animals. 8th edition. National Academies
Press: Washington, D.C; 1996
|
|
20
|
Miner J and Hoffhines A: The discovery of
aspirin's antithrombotic effects. Tex Heart Inst J. 34:179–186.
2007.PubMed/NCBI
|
|
21
|
Lijnen HR: Matrix metalloproteinases and
cellular fibrinolytic activity. Biochemistry (Mosc). 67:92–98.
2002. View Article : Google Scholar
|
|
22
|
Burnand KG, Gaffney PJ, McGuinness CL,
Humphries J, Quarmby JW and Smith A: The role of the monocyte in
the generation and dissolution of arterial and venous thrombi.
Cardiovasc Surg. 6:119–125. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Ye J, Duan H, Yang X, Yan W and Zheng X:
Anti-thrombosis effect of paeoniflorin: Evaluated in a
photochemical reaction thrombosis model in vivo. Planta Med.
67:766–767. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Yoshida A, Nakao S, Kobayashi M and
Kobayashi H: Flow-mediated vasodilation and plasma fibronectin
levels in preeclampsia. Hypertension. 36:400–404. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Pulkkinen MO, Lehto M, Jalkanen M and
Näntö-Salonen K: Collagen types and fibronectin in the uterine
muscle of normal and hypertensive pregnant patients. Am J Obstet
Gynecol. 149:711–717. 1984. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Lowe GD and Rumley A: Use of fibrinogen
and fibrin D-dimer in prediction of arterial thrombotic events.
Thromb Haemost. 82:667–672. 1999.PubMed/NCBI
|
|
27
|
Bowen RS, Zhang Y, Gu Y, Lewis DF and Wang
Y: Increased phospholipase A2 and thromboxane but not prostacyclin
production by placental trophoblast cells from normal and
preeclamptic pregnancies cultured under hypoxia condition.
Placenta. 26:402–409. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Zhao S, Gu Y, Lewis DF and Wang Y:
Predominant basal directional release of thromboxane, but not
prostacyclin, by placental trophoblasts from normal and
preeclamptic pregnancies. Placenta. 29:81–88. 2008. View Article : Google Scholar
|