Introduction
Osteosarcomas are malignant tumors arising from
mesenchymal cells (1). They are
the second most common primary bone malignancy, after multiple
myeloma, and represent 15% of all bone tumors (1). Men are more commonly afflicted than
women (ratio, 1.5:1) and 75% of patients are aged 15–25 years
(1). Risk factors include
retinoblastoma, ionizing radiation and germline p53 mutations
(1). Treatment strategies include
combinations of surgery, chemotherapy and radiation; with
chemotherapy increasing the 2- and 25-year survival rates (1–3).
Osteosarcomas predominantly metastasize to the lungs, liver and
brain (1), and overall survival is
~56% at 5 years and ~52% at 10 years (4). Angiogenesis is important in the
progression of malignant tumors, evolution and metastasis (5,6).
Endothelial progenitor cells (EPCs) are pluripotent stem cells that
have the potential to differentiate into mature endothelial cells
(7). They possess a strong
capacity for multiplication and colony formation (7). EPCs are present in the bone marrow
and in the peripheral blood (derived from the bone marrow)
(8). Peripheral blood EPCs are
important in active revascularization by differentiating into
mature endothelial cells to form new blood vessels (9,10).
Mukai et al (11) used an
in vivo 3D model using Matrigel to demonstrate that EPCs
form tubular structures. In addition, previous studies have
indicated that osteosarcoma cells have the potential to enhance
angiogenesis (12–14).
Previous studies have demonstrated that the stromal
cell-derived factor-1α (SDF-1α) and its receptor, C-X-C chemokine
receptor type 4 (CXCR4) are important in stem cell homing and tumor
metastasis (15,16). The SDF-1α/CXCR4 signal transduction
pathway is also important in tumor angiogenesis (17,18).
SDF-1α is expressed in hypoxic tissues, such as tumors and damaged
tissues, and is the primary chemokine that mobilizes pro-angiogenic
cells (19). CXCR4 is expressed on
EPCs, where it mediates the specific homing of EPCs to hypoxic
tissues to initiate angiogenesis (19).
Melittin is the main compound found in bee venom
(20). Modern pharmacology studies
have observed that melittin exerts various antitumor effects by
inhibiting tumor cell growth (21,22),
promoting tumor cell apoptosis (23–25),
and inhibiting tumor angiogenesis (26) and migration (27,28).
However, the effect of melittin on EPCs remains unknown.
Therefore, the aim of the present study was to
examine the effects of melittin on EPCs and on osteosarcoma-induced
angiogenesis, and to investigate the underlying mechanisms of these
effects. The current study hypothesized that melittin exerts its
effects by modulating SDF-1α/CXCR4 signaling.
Materials and methods
Animals
Male BALB/c nu/nu nude mice (n=24), aged 4–6 weeks
and weighing 18–20 g, were bought from Shanghai SLAC Laboratory
Animal Co., Ltd. (Shanghai, China). Mice were housed in separate
cages at the Animal Experiment Centre of the First Affiliated
Hospital of Guangxi University of Traditional Chinese Medicine
(Guangxi, China) in a specific pathogen-free environment and were
maintained under a 12-h light/dark cycle, with access to food and
water ad libitum. Mice were acclimatized to their
environment for one week prior to experiments. All procedures and
animal experiments were approved by the Animal Care and Use
Committee of the First Affiliated Hospital of Guangxi University of
Traditional Chinese Medicine.
Cell culture
The UMR-106 rat osteoblastic osteosarcoma cell line
was obtained from the Institute of Orthopedics of the Fourth
Military Medical University (Xi'an, China). UMR-106 cells were
cultured in Gibco Dulbecco's modified Eagle's medium (DMEM; Thermo
Fisher Scientific, Inc., Waltham, MA, USA) supplemented with Gibco
10% fetal bovine serum (FBS; Thermo Fisher Scientific, Inc.) at
37°C in a humidified atmosphere of 5% CO2.
Isolation, culture and identification of
EPCs
EPCs were obtained using previously described
methods (29,30). The femur and tibia were removed
under sterile conditions. Metaphyseal bone forceps were used to
remove the bone ends to expose the medullary cavity. M199 medium
(Gibco; Thermo Fisher Scientific, Inc.) was injected from one end
of the bone, and the bone marrow washing out was collected on the
other end. Mononuclear cells were isolated by density gradient
centrifugation. Following two washes with phosphate-buffered saline
(PBS; Labest Biotechnology Co., Ltd., Beijing, China), mononuclear
cells were suspended in M199 medium with 10% FBS, 10 ng/ml vascular
endothelial growth factor (Prospec-Tany TechnoGene Ltd., East
Brunswick, NJ, USA) and 10 ng/ml basic fibroblast growth factor
(Prospec-Tany TechnoGene Ltd.). Mononuclear cells were inoculated
in Petri dishes pre-coated with rat fibronectin (RFN;
Sigma-Aldrich, St. Louis, MO, USA) and incubated at 37°C in a 5%
CO2 atmosphere. The medium was replaced after 4 days,
and subsequently every 3 days. Cells at passage 3 were stained with
rabbit anti-rat cluster of differentiation (CD)34 (Abcam,
Cambridge, MA, USA), rabbit anti-rat CD133 (Abcam), fluorescein
isothiocyanate (FITC)-conjugated goat anti-rabbit immunoglobulin
(Ig)G (Yingrun Biotechnology Co., Ltd., Changsha, China) and
Cy3-conjugated goat anti-rabbit IgG (Yingrun Biotechnology Co.,
Ltd.). The double-positive expression of CD34 and CD133, indicating
EPCs (31,32), was observed using a BX51
fluorescence microscope (Olympus Corporation, Tokyo, Japan).
Non-specific anti-rat antibodies served as an isotype control.
Effect of melittin on cell viability
UMR-106 cells and EPCs (1,000 cells/well) were
plated in 96-well plates for 24 h. Blank control wells contained
medium without cells. Following 24 h, the culture medium was
replaced with DMEM containing 5% FBS to synchronize the cells for 6
h. The cells were treated with various concentrations (0, 1, 2, 4,
8 and 16 µg/ml dissolved in 0.9% saline) of melittin
(>98% purity; Sigma-Aldrich) for 48 h. MTT (10 µl; 5
mg/ml; Ameresco, Inc., Framingham, MA, USA) was added and the
incubation continued for 4 h. The medium containing MTT was
discarded, 200 µl dimethyl sulfoxide (Haoran Biotechnology
Co., Ltd., Shanghai, China) was added and the plates were agitated
for 10 min. A Model 500 Microplate Reader (Bio-Rad Laboratories,
Inc., Hercules, CA, USA) was used to measure the optical density
(OD) at a wavelength of 490 nm. The percentage of cell viability
was calculated according to the following formula: Cell viability
(%) = (average OD of the experimental group)/(average OD of the
control group) × 100, where average is the optical density obtained
from three independent experiments performed in triplicate.
Adhesion of EPCs
EPCs (1×105 cells/well) were plated in
12-well RFN-coated plates. EPCs were divided into four groups:
Control (untreated); 10 ng/ml SDF-1α (Prospec-Tany TechnoGene
Ltd.); 1 µg/ml melittin + 10 ng/ml SDF-1α; and 3
µg/ml melittin + 10 ng/ml SDF-1α. At these concentrations
melittin was determined to have low toxicity in preliminary
experiments. Following 2 h, the culture medium was discarded, the
EPCs were washed twice with PBS, and fixed with 95% alcohol for 30
min. Cells were washed twice with PBS, stained with 0.1% crystal
violet (200 µl; Haoran Biotechnology Co., Ltd.) for 15 min
and washed twice with PBS. The plates were dried and observed using
the IX70 inverted optical microscope (Olympus Corporation). Images
of five randomly selected fields were captured (magnification,
×100). Adherent EPCs were counted by two independent observers who
were blinded to the experimental conditions.
Migration of EPCs
The migration ability of EPCs was assessed using
24-well Transwell plates (pore size, 8 µm; Corning
Incorporated, Corning, NY, USA). M199 medium containing 2% FBS
(control) and the treatments (10 ng/ml SDF-1α; 1 µg/ml
melittin + 10 ng/ml SDF-1α; and 3 µg/ml melittin + 10 ng/ml
SDF-1α) was placed in the lower chamber and served as a
chemoattractant. M199 culture medium (200 µl) containing
1×105 EPCs was placed in the upper chamber. Following 6
h, the cells on the upper surface of the filter were removed by
gently wiping with a cotton swab. The cells that had migrated were
fixed with 95% alcohol for 30 min and stained with 0.1% crystal
violet. Migrated cells were visualized using the inverted optical
microscope. Images of five randomly selected fields were captured
(magnification, ×100) and the number of migrated cells was counted
by two independent observers blinded to the experimental
conditions.
Tube formation assay
Matrigel (BD Biosciences, Franklin Lakes, NJ, USA)
was added to 48-well plates to a total volume of 120
µl/well. Wells were divided into four groups: Control
(untreated); 10 ng/ml SDF-1α, 1 µg/ml melittin + 10 ng/ml
SDF-1α; and 3 µg/ml melittin + 10 ng/ml SDF-1α. Gel was
allowed to polymerize for 30 min at 37°C and EPCs (5×104
cells/well) were inoculated on the Matrigel. Following 23 h of
incubation, at 37°C in 5% CO2, the area covered by the
tube network was determined using the inverted optical microscope
(magnification, ×100).
Western blot analysis of CXCR4, SDF-1α,
phosphorylated (p)-AKT, AKT, p-ERK1/2, ERK1/2 in UMR-106 cells and
EPCs
Cells were lysed using a total protein lysis buffer
(Shanghai Pufei Biotech Co., Ltd., Shanghai, China) and centrifuged
at 8000 × g for 15 min at 4°C. Protein concentrations were
determined using a BCA kit (Shanghai Pufei Biotech Co., Ltd.).
Equal quantities of proteins were isolated by 8% SDS-PAGE (run at
110 Volts for 90 min) and transferred onto nitrocellulose membranes
(Invitrogen; Thermo Fisher Scientific, Inc.). Membranes were
blocked for 2 h in 5% dried milk at room temperature, and washed in
Tris-buffered saline with Tween-20 (TBST) three times for 10 min
each time. Membranes were incubated overnight at 4°C with primary
antibodies as follows: rabbit polyclonal CXCR4 (1:200; Abcam; cat.
no. ab74012), SDF-1α (1:200; Yuan Mu Biotechnology Co., Ltd.,
Shanghai, China; cat. no YM-XQ4938P); p-AKT (1:200; Abcam; cat. no.
ab38449); AKT (1:200; Abcam; cat. no. ab79360); p-ERK1/2 (1:200;
Santa Cruz Biotechnology, Inc.; cat. no. sc-101760); ERK1/2 (1:200;
Santa Cruz Biotechnology, Inc.; cat. no. sc-292838); mouse
monoclonal β-actin (1:200; Abcam; cat. no. ab6276). Membranes were
washed with TBST three times for 10 min each time, and incubated
for 1 h at room temperature with polyclonal HRP-conjugated goat
anti-rabbit IgG (1:1,000; GenScript Co., Ltd., Nanjing, China; cat.
no. A00098) and polyclonal HRP-conjugated goat anti-mouse IgG
(1:1,000; Wuhan Boster Biological Technology, Ltd., Wuhan, China;
cat. no. BA1050) secondary antibodies. Following three further
washes in TBST, for 10 min each time, membranes that contained the
relevant protein bands were observed by enhanced chemiluminescence
using the BeyoECL Plus hypersensitive ECL chemiluminescence kit
(Beyotime Institute of Biotechnology, Nanjing, China). Images were
captured and analyzed.
UMR-106 osteosarcoma xenograft mouse
model
The mouse model of osteosarcoma was established
using a previously described method (33). Male nude mice were anesthetized by
intraperitoneal injection of 1% sodium pentobarbital (70 mg/kg) and
inoculated with UMR-106 cells (2×105/20 µl) in
the left hind leg tibial plateau. Intra-tumor multipoint local
injections were administered when the tumors grew to ~0.5×0.5 cm.
In the negative control group (n=6), tumors were injected with
normal saline in a total volume of 200 µl, once a day. For
the low, moderate and high melittin groups (n=6/group), mice
received a local injection of 160, 320 and 640 µg/kg
melittin, respectively, once a day, for 5 days. The mice were
treated for two 5-day periods with one day between the two periods.
Following each injection, the major and minor axes of the tumors
were measured. The size of the tumors was calculated using the
following formula: Tumor volume (mm3) = major axis ×
minor axis2/2 (34).
Mice were sacrificed on the day after the second period of
treatment.
Immunofluorescence staining for CD34 and
CD133
Mouse tumor tissue samples were fixed in 10%
neutralized formalin (Haoran Biotechnology Co., Ltd.), dehydrated,
embedded in paraffin and sliced into 4-µm sections. The
sections were dewaxed, endogenous peroxidase was neutralized with
3% H2O2 and antigen retrieval was performed
by incubating the sections in 0.01 M citrate buffer (pH 6.0; Labest
Biotechnology Co., Ltd.) for 40 min at 92°C, and in normal goat
serum (Wuhan Amyjet Scientific Co., Ltd., Wuhan, China) for 20 min
at 37°C. The sections were incubated with rabbit polyclonal CD34
(1:100; Abcam; cat. no. ab185732), rabbit polyclonal CD133 (1:100;
Otwo Biotech Co., Ltd., Shenzhen, China; cat. no. 251149),
FITC-conjugated goat anti-rabbit IgG (1:500; Walan Biotechnology
Co., Ltd., Shanghai, China; cat. no. AS011) and Cy3-conjugated goat
anti-rabbit IgG (1:500; Sanying Biotechnology Co., Ltd., Wuhan,
China; cat. no. 00009-2) in the dark, at 37°C for 30 min. Sections
were washed 3 times in PBS, glycerophosphoric acid buffer solution
(Labest Biotechnology Co., Ltd.) was added, and observed under IX70
inverted fluorescence microscope (Olympus Corporation). The
double-positive area was quantified as the percentage of the total
area using Image Pro Plus software, version 5.0 (Media Cybernetics,
Inc., Rockville, MD, USA). The quantity of EPCs was evaluated.
Immunohistochemical detection of CXCR4,
SDF-1α and microvessel density (MVD)
Expression levels of CD105, CXCR4 and SDF-1α were
detected by immunohistochemistry. Sections were dewaxed, endogenous
peroxidase was neutralized with 3% H2O2 and
antigen retrieval was conducted by incubating the sections in 0.01
M citrate buffer (pH 6.0) for 40 min at 92°C, and in normal goat
serum for 20 min at 37°C. Sections were incubated with rabbit
polyclonal CD105 (1:200; Abcam; cat. no. ab107595), rabbit
polyclonal CXCR4 (1:200) and rabbit polyclonal SDF-1α (1:200)
overnight at 4°C. The sections were washed 3 times in PBS and
incubated with polyclonal HRP-conjugated goat anti-rabbit IgG
(1:1,000) secondary antibody at 37°C. Sections were revealed by
3,3′-diaminobenzadine (DAB) (Haoran Biotechnology Co., Ltd.),
counterstained with hematoxylin (Haoran Biotechnology Co., Ltd.),
dehydrated and mounted with neutral balsam (Noble Ryder Beijing
Science and Technology Co., Ltd., Beijing, China). The positive
areas were quantified as the percentage of the total area using
Image Pro Plus 5.0. A negative control was obtained using PBS
rather than a primary antibody.
MVD
MVD was assessed by immunohistochemistry according
to the method by Weidner et al (35) using CD105-positive cells. Each EPC
or cluster of EPCs that could be separated from surrounding
vessels, tumor cells and other connective tissues were counted as a
single capillary. As long as the microvascular structure was not
continuous, the branch structure was also counted as a vessel.
Capillaries were not assessed according to the presence of
erythrocytes or by the presence of a lumen. Capillaries with a
lumen area >8 erythrocytes in diameter and with a thick muscular
layer were not counted, furthermore, capillaries in fibrotic areas
and in areas with scarce tumor cells were not counted. The five
areas with the highest MVD were photographed under an inverted
optical microscope (magnification, ×400). MVD was evaluated by two
independent observers blinded to the experimental conditions. The
average value of the five areas (mean ± standard deviation) was
taken as the MVD value of the tumor.
Statistical analysis
Continuous data are presented as means ± standard
deviation from three independent experiments. One-way analysis of
variance was conducted with the least significant difference for
post hoc tests. SPSS 15.0 (SPSS, Inc., Chicago, IL, USA) was used
for statistical analysis and P<0.05 was considered to indicate a
statistically significant difference.
Results
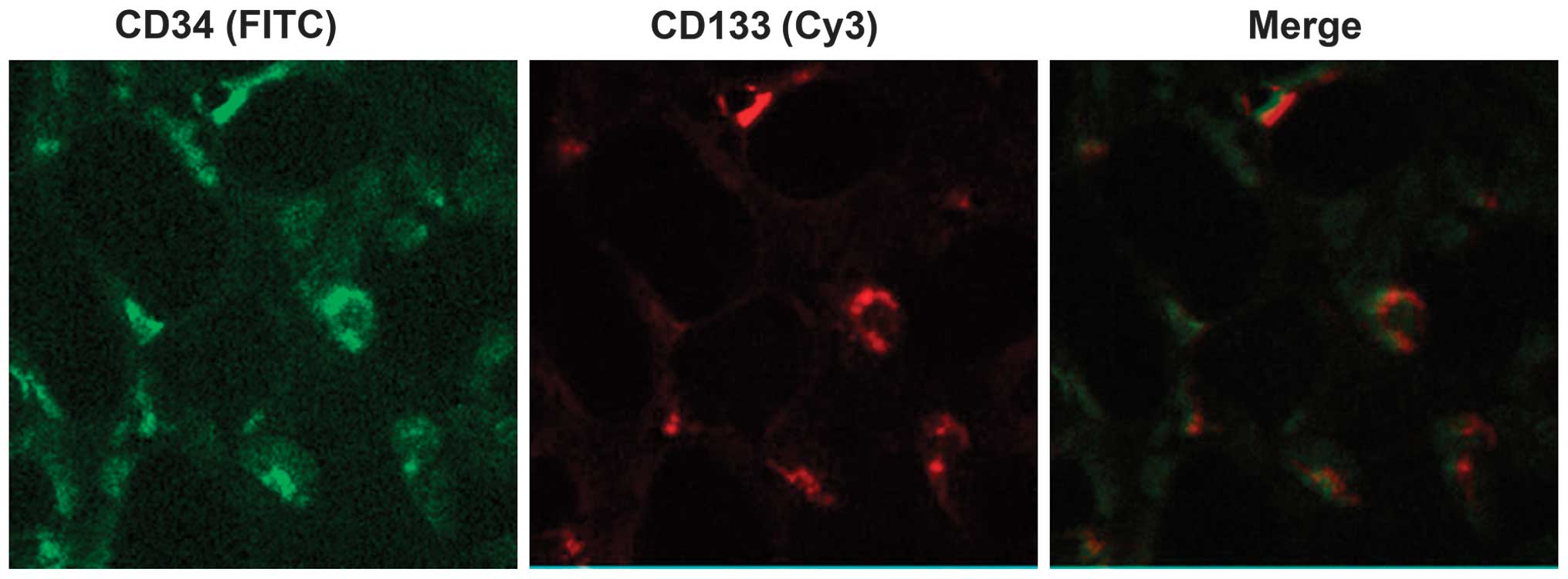
EPCs were successfully isolated
As presented in Fig.
1, cells with a green fluorescence signal are CD34-positive
cells, and cells with a red fluorescence signal are CD133-positive
cells. The merge indicates the CD34/CD133 double-positive cells,
which demonstrate the EPCs (31,32).
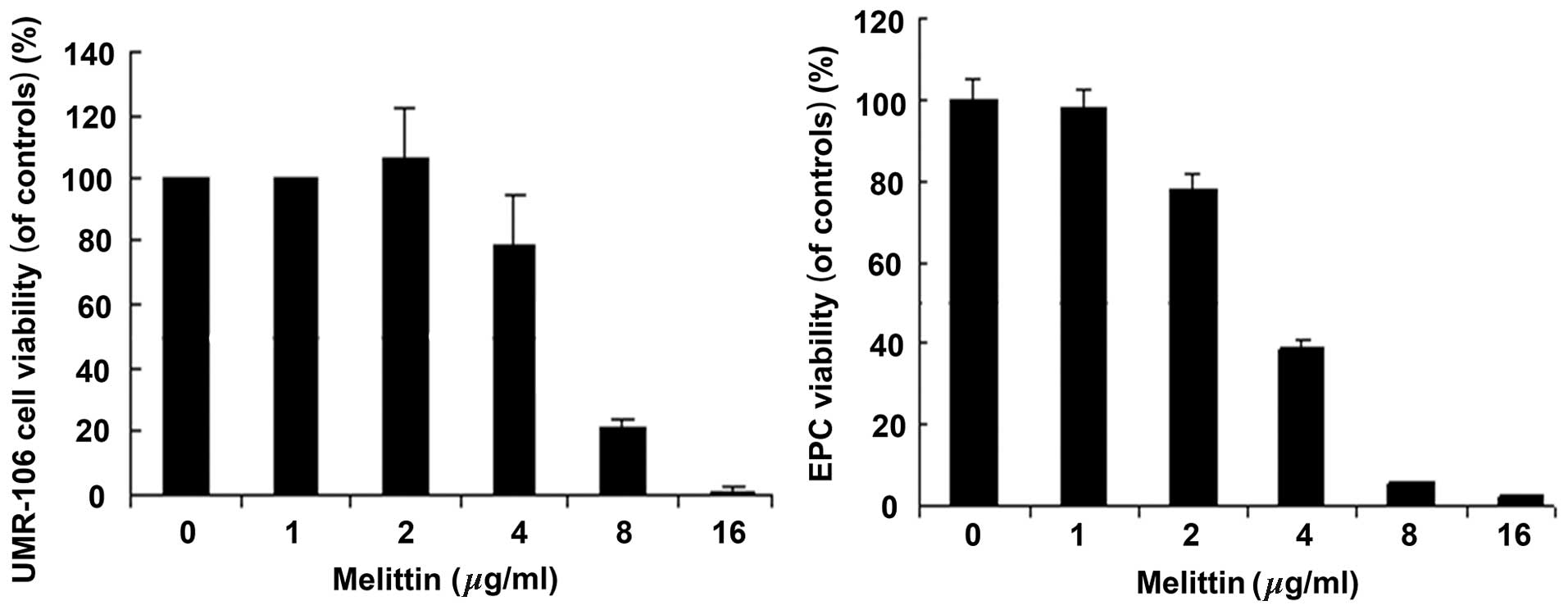
Melittin decreases the viability of
UMR-106 cells and EPCs
Fig. 2 demonstrates
that melittin decreased the viability of UMR-106 cells and EPCs in
a dose-dependent manner. The half maximal inhibitory concentration
(IC50) values for UMR-106 cells and EPCs were 6.33 and
4.51 µg/ml, respectively.
SDF-1α increases EPCs adhesion, which is
decreased by melittin
Fig. 3A indicates
that melittin decreased the adherence of EPCs, which was
facilitated by SDF-1α. The number of adherent cells was 620.8±19.6
in the control, 900.6±16.2 in cells treated with 10 ng/ml SDF-1α
(P<0.01 vs. control), 532.4±5.6 in cells treated with 1
µg/ml melittin + 10 ng/ml SDF-1α (P<0.001 vs. SDF-1α) and
270.2±1.5 in cells treated with 3 µg/ml melittin + 10 ng/ml
SDF-1α (P<0.001 vs. control; P<0.001 vs. SDF-1α).
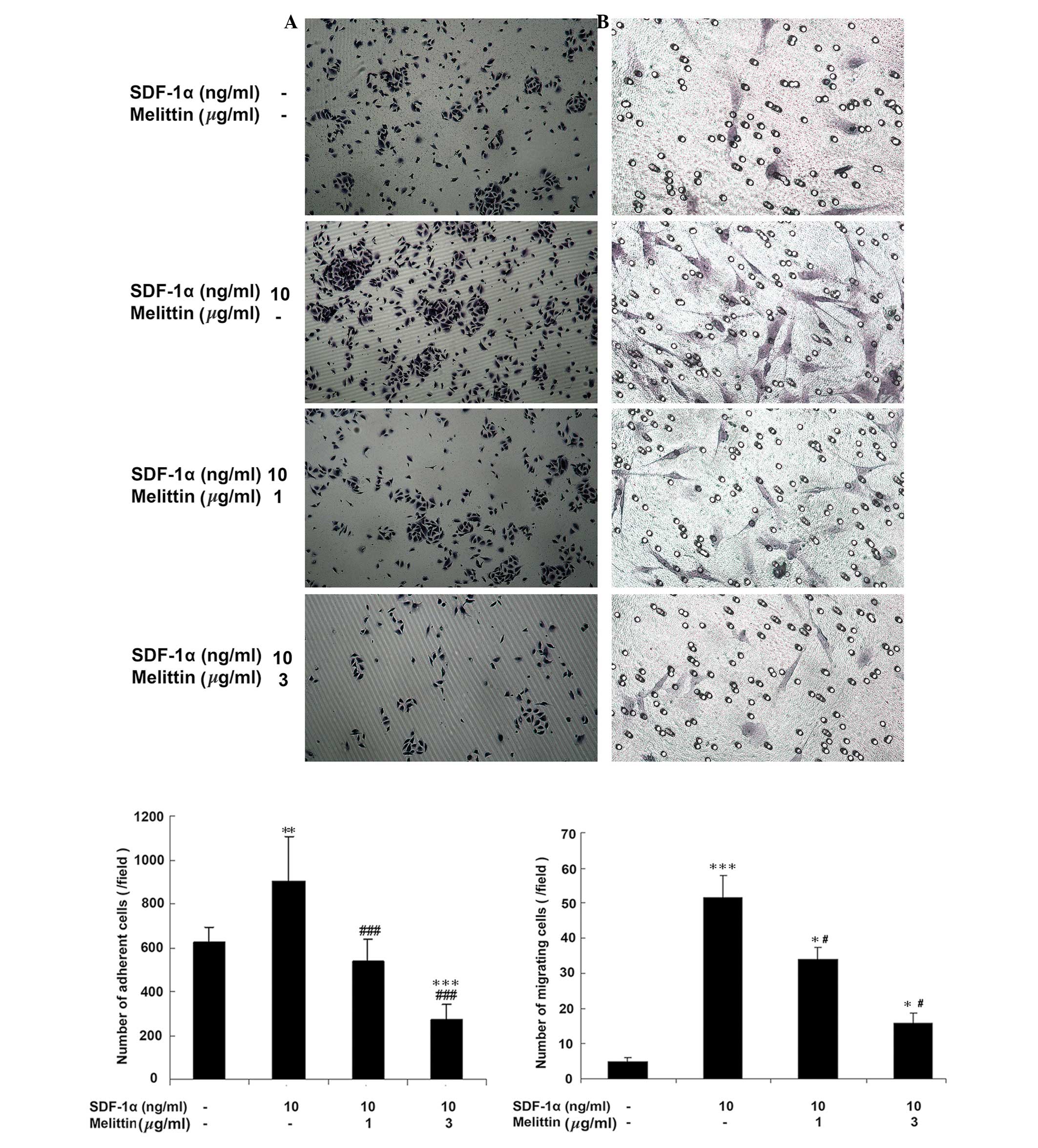 | Figure 3Effects of SDF-1α and melittin on EPC
adhesion and migration. EPCs were divided into four groups: Control
(untreated), 10 ng/ml SDF-1α, 1 µg/ml melittin + 10 ng/ml
SDF-1α, and 3 µg/ml melittin + 10 ng/ml SDF-1α. (A)
Following 2 h of treatment, adhesion of EPCs was determined
(magnification, ×100). (B) Following 6 h of treatment, migration of
EPCs was determined by Transwell migration assay (magnification,
×200). Data are presented as means ± standard deviation.
*P<0.05, **P<0.01,
***P<0.001 vs. controls; #P<0.05,
###P<0.001 vs. SDF-1α. SDF-1α, stromal cell-derived
factor-1α; EPC, endothelial progenitor cell. |
SDF-1α increases EPCs migration, which is
decreased by melittin
Fig. 3B
demonstrates that melittin decreased the number of migrating EPCs,
which was facilitated by SDF-1α. The number of cells that had
migrated was 4.8±6.4 in the control, 51.5±3.5 in cells treated with
10 ng/ml SDF-1α (P<0.001 vs. control), 34.1±3.0 in cells treated
with 1 µg/ml melittin + 10 ng/ml SDF-1α (P<0.05 vs. the
control; P<0.05 vs. SDF-1α) and 15.7±1.8 in cells treated with 3
µg/ml melittin + 10 ng/ml SDF-1α (P<0.05 vs. control;
P<0.05 vs. SDF-1α).
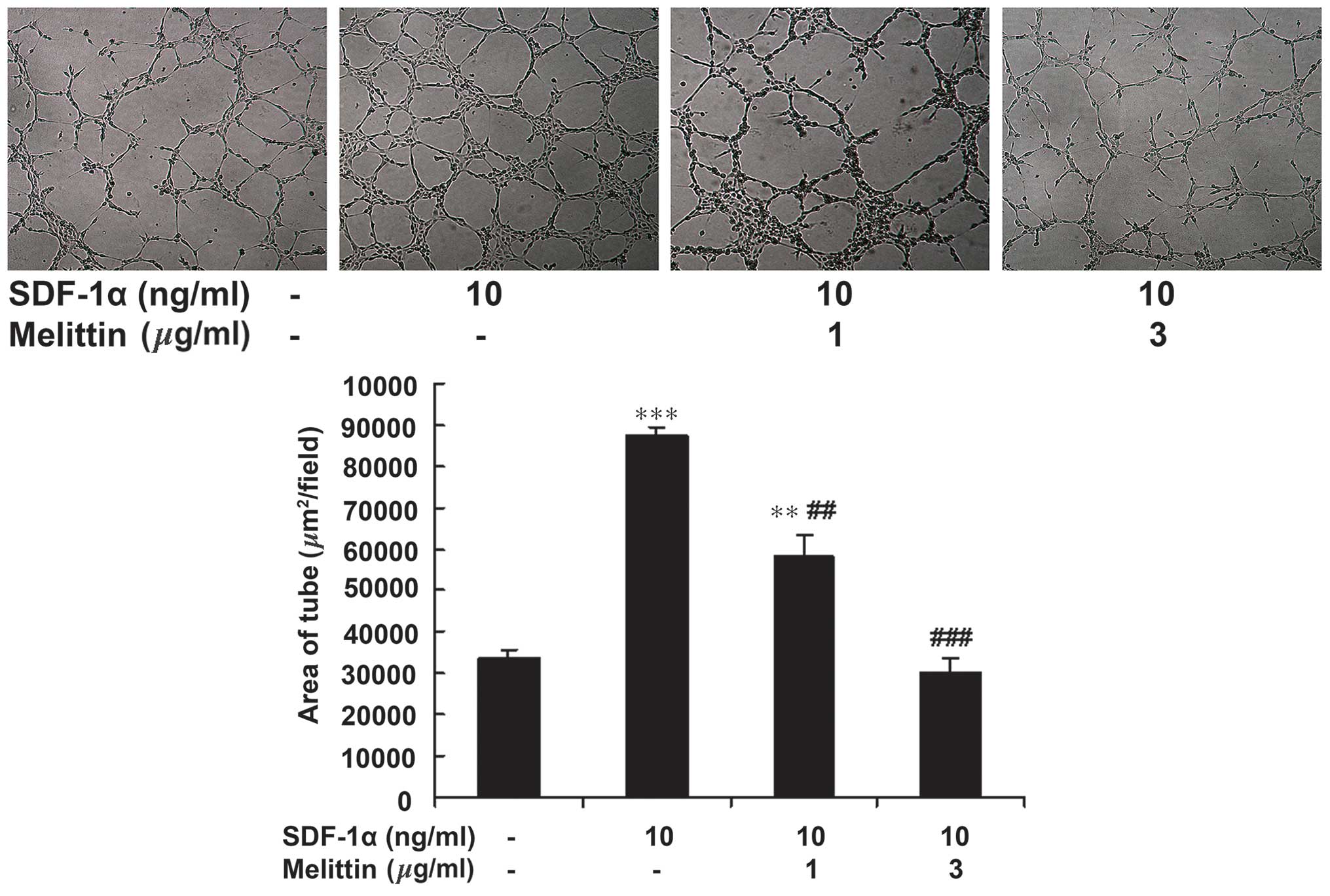
SDF-1α increases EPCs tube formation,
which is decreased by melittin
Fig. 4 indicates
that melittin decreased the tube forming ability of EPCs, which was
facilitated by SDF-1α. The area of the tubes was 33,806±1,703
µm2/field in the control, 87,504±2,102
µm2/field in cells treated with 10 ng/ml SDF-1α
(P<0.001 vs. control), 58,401±4,857 µm2/field
in cells treated with 1 µg/ml melittin + 10 ng/ml SDF-1α
(P<0.01 vs. control; P<0.01 vs. SDF-1α) and 30,165±3,430 in
cells treated with 3 µg/ml melittin + 10 ng/ml SDF-1α
(P<0.001 vs. SDF-1α).
Melittin decreased the protein expression
levels of p-AKT, p-ERK1/2, SDF-1α and CDCR4 in the UMR-106 cells
and EPCs
Fig. 5 presents the
western blots of CXCR4, SDF-1α, p-AKT, AKT, p-ERK1/2, and ERK1/2
expression in UMR-106 cells and EPCs following treatment with
SDF-1α and melittin. Compared with the control, melittin decreased
the expression of p-AKT, p-ERK1/2, SDF-1α and CXCR4 in UMR-106
cells and EPCs.
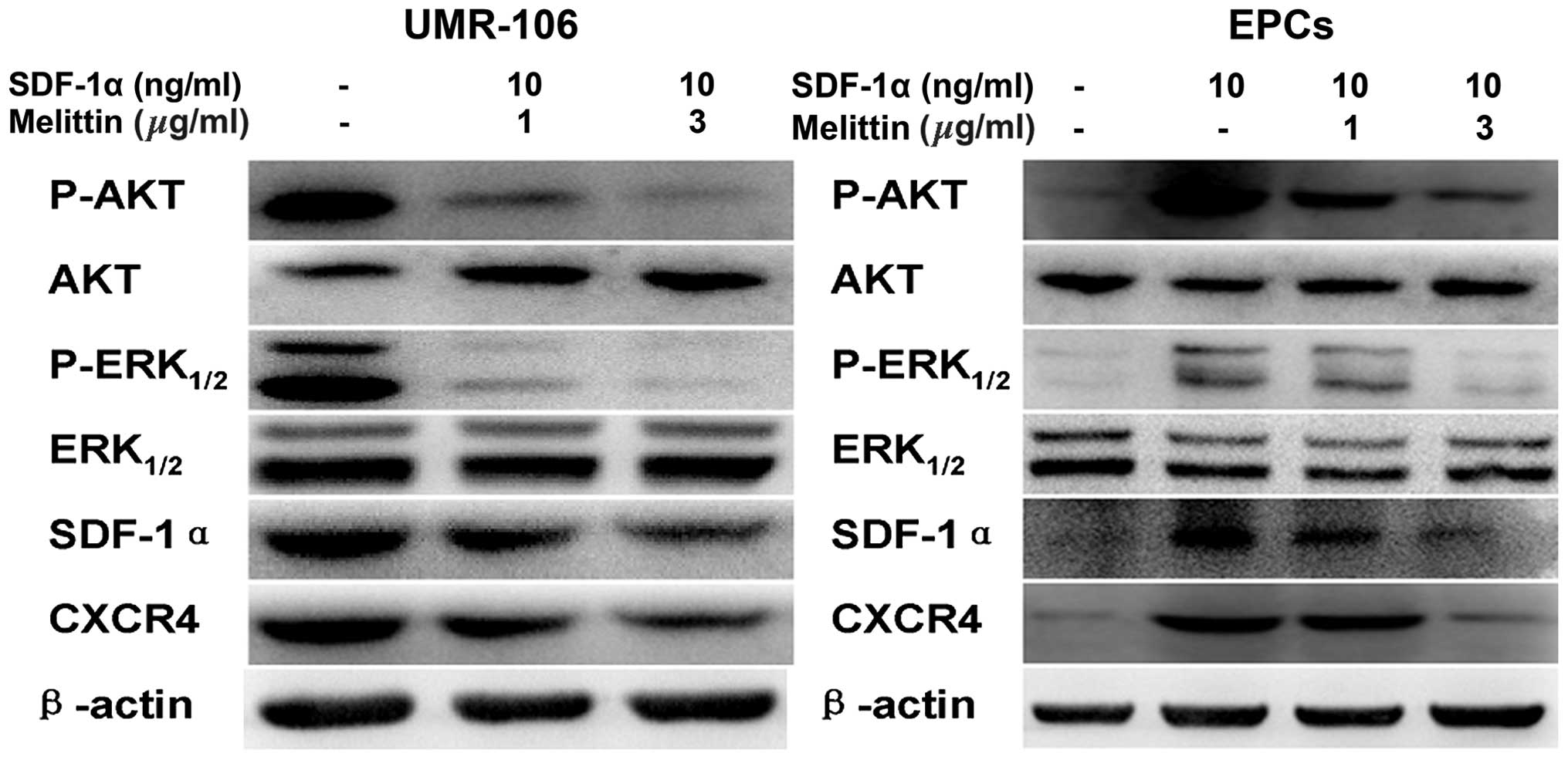 | Figure 5Effects of SDF-1α and melittin on the
expressions of p-AKT, AKT, p-ERK1/2, ERK1/2, SDF-1α, CXCR4 in
UMR-106 cells and EPCs. Protein expression was determined by
western blot analysis and β-actin served as an internal control.
SDF-1α, stromal cell-derived factor-1α; EPC, endothelial progenitor
cell; p, phosphorylated; ERK, extracellular signal-regulated
kinase; CXCR4, C-X-C chemokine receptor type 4. |
Melittin decreases the proportion of
CD34/CD133 cells in an osteosarcoma xenograft mouse model
A mouse model of osteosarcoma was established using
UMR-106 cells and tumors were injected with various doses of
melittin. Intra-tumor EPCs were detected by immunofluorescence
(Fig. 6). The proportion of
CD34/CD133 double-positive cells was 16.4±10.4% in the control,
7.0±4.4% in tumors treated with 160 µg/kg melittin per day
(P<0.01 vs. control), 2.9±1.2% in tumors treated with 320
µg/kg melittin per day (P<0.001 vs. control and P<0.05
vs. 160 µg/kg melittin), and 1.3±0.3% in tumors treated with
640 µg/kg melittin per day (P<0.001 vs. control,
P<0.01 vs. 160 µg/kg melittin, and P<0.05 vs. 320
µg/kg melittin).
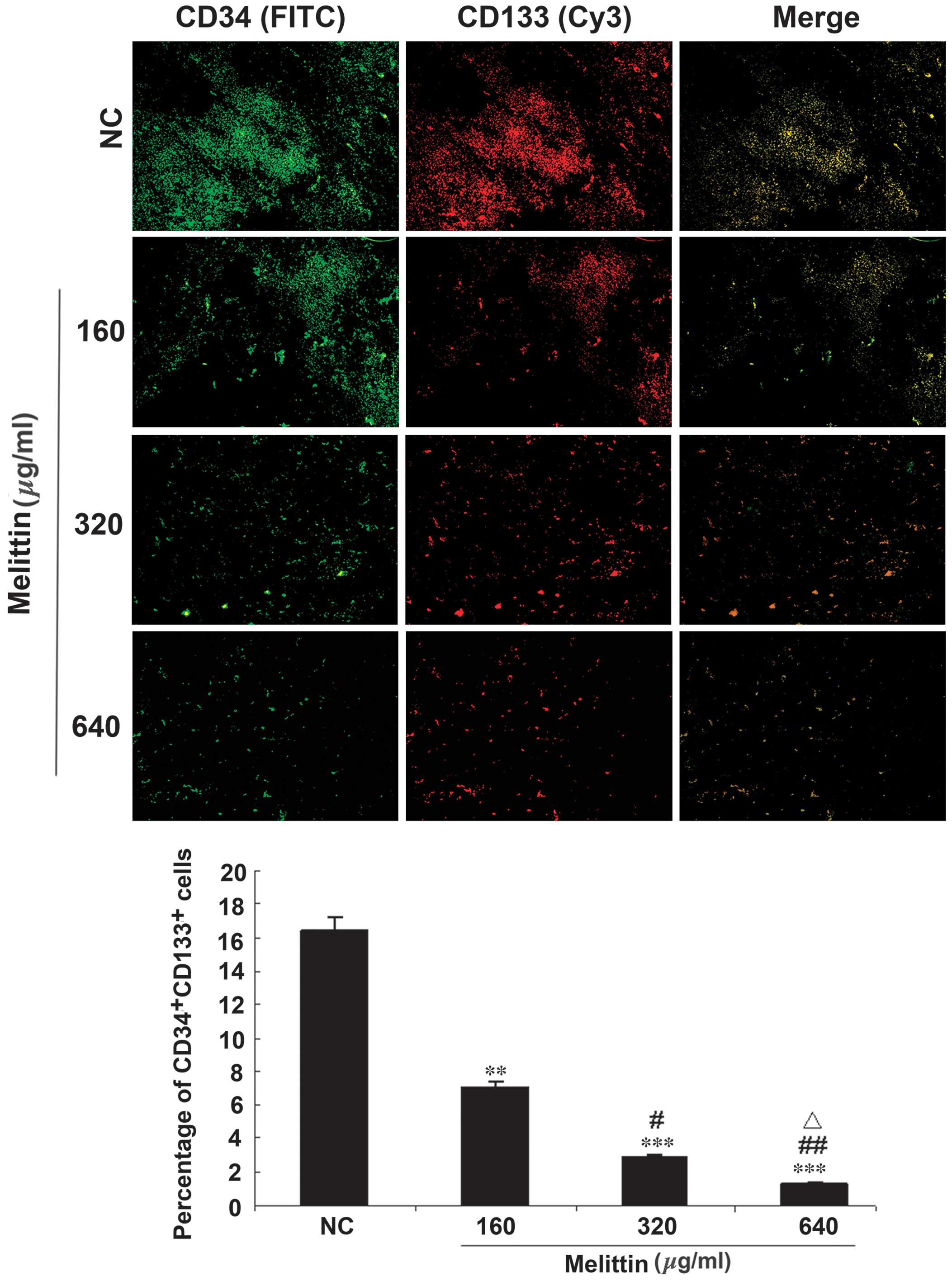 | Figure 6Effects of melittin on the proportion
of CD34/CD133 double-positive cells in the mouse model of UMR 106
osteosarcoma. Intra-tumor multipoint local injections were
administered when the tumors grew to ~0.5×0.5 cm. For the NC group,
tumors were treated with a local injection of normal saline (total
volume, 200 µl) once a day. Tumors were treated with local
injections of 160, 320 or 640 µg/kg melittin, once a day,
for a treatment period of 5 days. Mice were treated for two 5-day
periods, with one day between the two periods. CD34 (green, FITC)
and CD133 (red, Cy3) expression in the tumor tissue samples was
detected using fluorescence microscopy (magnification, ×100). The
proportions of CD34/CD133 double-positive cells are presented as
means ± standard deviation (n=6/group). **P<0.01,
***P<0.001 vs. NC; #P<0.05,
##P<0.01 vs. 160 µg/kg melittin;
△P<0.05 vs. 320 µg/kg melittin. CD, cluster of
differentiation; NC, negative control; FITC, fluorescein
isothiocyanate; Cy3, cyanine 3. |
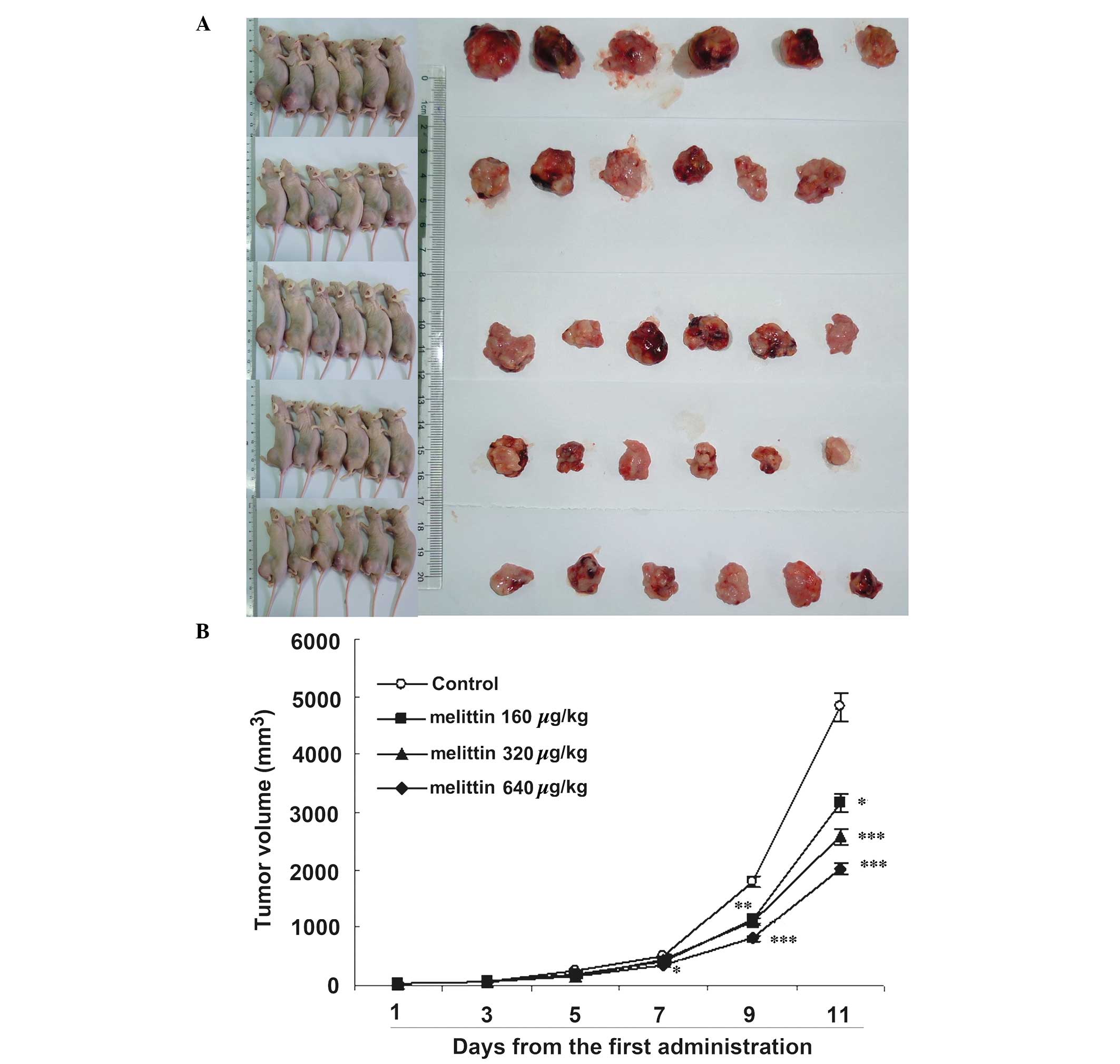
Melittin decreases tumor size in
vivo
Fig. 7 demonstrates
that melittin decreased osteosarcoma growth. Curves began to
separate at 7 days following onset of treatment. At 11 days, the
three doses of melittin resulted in smaller tumors compared with
the control (control, 4.8±1.3 cm3; 160 µg/kg
melittin, 3.2±0.6 cm3; 320 µg/kg melittin,
2.6±0.5 cm3; and 640 µg/kg melittin, 2.0±0.2
cm3; all P<0.05 vs. the control).
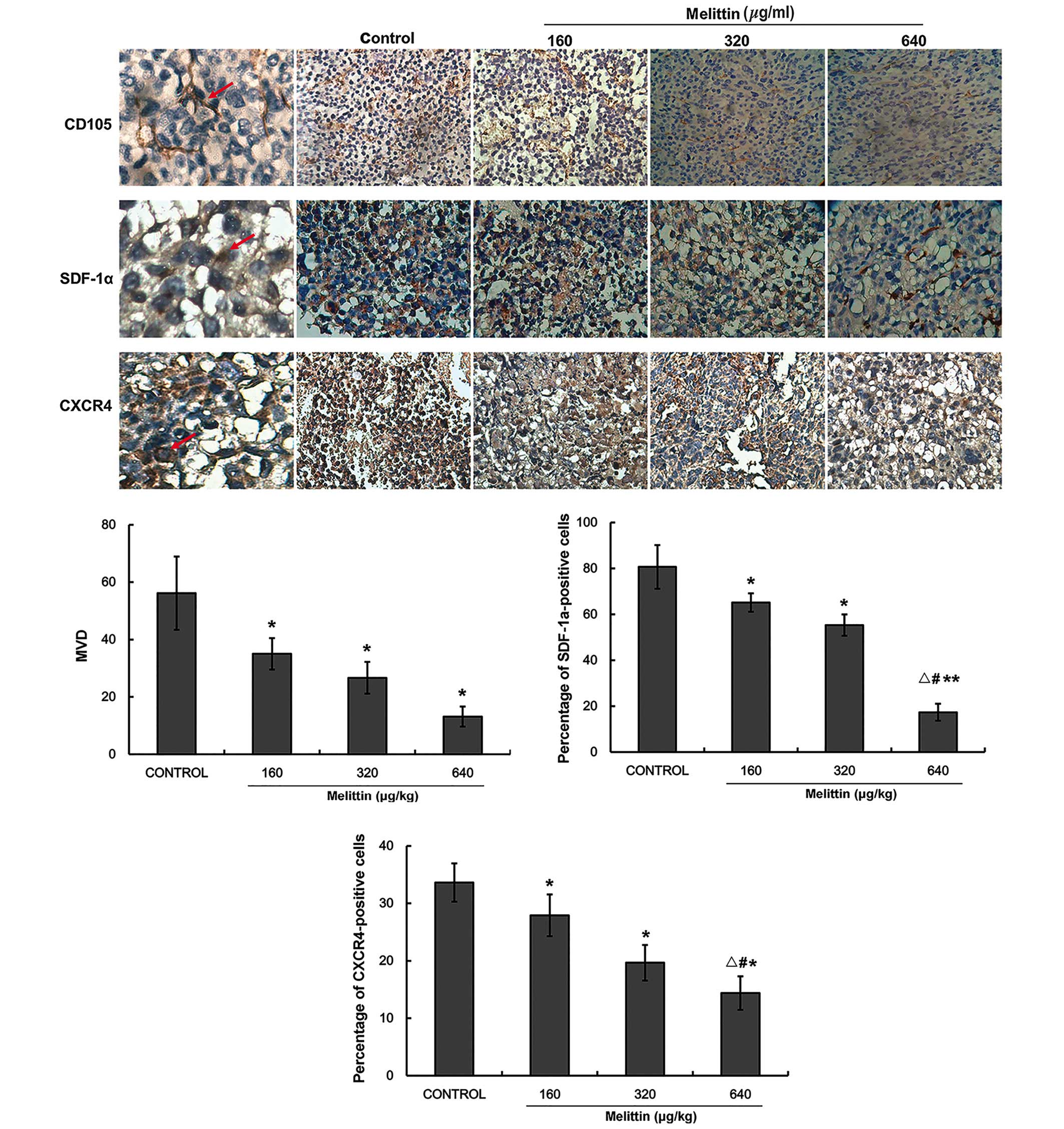
Melittin decreases MDV, CXCR4 and SDF-1α
protein expression levels
CD105 immunohistochemistry was performed to
determine MVD. Fig. 8 indicates
that MVD decreased with melittin administration in a dose-dependent
manner (control, 56.2±12.8; 160 µg/kg melittin, 35.0±5.5;
320 µg/kg melittin, 26.7±5.5; and 640 µg/kg melittin,
13.2±3.5 cm3; all P<0.05 vs. control).
Fig. 8 indicates
that SDF-1α expression decreased with melittin in a dose-dependent
manner (control, 80.7±9.5; 160 µg/kg melittin, 65.2±4.0; 320
µg/kg melittin, 55.3±4.6; and 640 µg/kg melittin,
17.3±3.7; all P<0.05 vs. control).
CXCR4 expression levels decreased with melittin in a
dose-dependent manner (control, 33.6±3.3; 160 µg/kg
melittin, 27.9±3.6; 320 µg/kg melittin, 19.7±3.1; and 640
µg/kg melittin, 14.4±2.9; all P<0.05 vs. control;
Fig. 8).
Discussion
EPCs are important in tumor angiogenesis, SDF-1α and
its receptor, CXCR4, are key in stem cell homing, and melittin (a
component of bee venom) exerts antitumor activity, although its
underlying mechanisms remain unknown. Furthermore, osteosarcomas
maintain a tumor microenvironment, which stimulates angiogenesis
(12). Therefore, the aim of the
present study was to examine the effects of melittin on EPCs and
angiogenesis, and investigate the underlying mechanisms of these
effects.
Melittin exposure decreased the viability of UMR-106
cells and EPCs, with IC50 values of 6.33 and 4.51
µg/ml, respectively. Furthermore, melittin decreased EPC
adhesion, migration and tube formation when compared with the
control and SDF-1α-treated cells. Compared with the control. In
addition, the expression levels of p-AKT, p-ERK1/2, SDF-1α and
CXCR4 in UMR-106 cells and EPCs were decreased by melittin
administration. The proportions of CD34/CD133 double-positive cells
were 16.4±10.4% in the control cells, and 7.0±4.4, 2.9±1.2 and
1.3±0.3% in tumors treated with 160, 320 and 640 µg/kg
melittin per day, respectively (all P<0.05). At 11 days,
melittin reduced the tumor size compared with the control (control,
4.8±1.3 cm3; melittin, 3.2±0.6, 2.6±0.5, and 2.0±0.2
cm3 for 160, 320 and 640 µg/kg, respectively; all
P<0.05). Furthermore, melittin decreased MVD, and SDF-1α and
CXCR4 protein expression levels in tumor tissue samples.
EPCs migrate to tumor tissues and are involved in
tumor angiogenesis; thus they present as a potential target in
treatment strategies against tumor angiogenesis. Numerous stages of
the process could be targeted, such as EPC mobilization,
recruitment and/or differentiation. As osteosarcomas are tumors
that actively promote angiogenesis (12), EPC targeting may provide a novel
approach for osteosarcoma therapy (36).
EPCs are pluripotent stem cells, which have the
potential to differentiate into mature endothelial cells (37). However, the exact role of the
SDF-1α/CXCR4 signaling pathway in EPC-mediated angiogenesis in
osteosarcomas remains unclear. In the present study, melittin
significantly inhibited the angiogenesis ability of EPCs, which is
induced by SDF-1α. The present study demonstrates that this
inhibition is mediated by the melittin-induced inhibition of AKT
and ERK1/2 phosphorylation. Therefore, the results suggest that
melittin inhibits angiogenesis in EPCs via inhibiting the
SDF-1α/CXCR4 signaling pathway. Results from the in vivo
experiment further suggest that melittin may be administered to
control the growth of osteosarcomas. The local injection approach
decreased the likelihood of inducing a systemic reaction. In
addition, previous studies have indicated that melittin exerts
certain beneficial effects against cancer, such as decreased tumor
invasion and decreased expression of proteins involved in tumor
progression and invasion (38,39).
As with white blood cell homing to inflammatory
tissues, EPC homing to tumors is dependent on SDF-1α and its
receptor, CXCR4. The activation of the SDF-1α/CXCR4 signaling
pathway is important in EPC migration and angiogenesis in tumors
(19). The activation of CXCR4 by
SDF-1α stimulates various downstream signaling pathways, such as
phosphoinositide 3-kinase (PI3K)/AKT and mitogen-activated protein
kinase/ERK1/2 (40–43). EPC homing and migration are
improved by the upregulation of CXCR4 (44), as well as by the improvement of
signaling pathways that are mediated by CXCR4 (45). Furthermore, tumor and stromal cells
are the predominant source of SDF-1α in the tumor microenvironment
(46).
CXCR4 is expressed on EPCs (47,48)
and directs the cells toward tumors to induce angiogenesis
(49). In addition, CXCR4 has been
involved in osteosarcoma growth and metastases (50,51).
The present study indicated that melittin may decrease the levels
of p-AKT and ERK1/2 via reduced expression of CXCR4 on EPCs. A
previous study demonstrated that melittin inhibits the
PI3K/AKT/mechanistic target of rapamycin signaling pathway in
breast cancer cells (52).
However, further studies are required to elucidate the precise
mechanisms involved.
In the present study, in vivo experiments
demonstrated that melittin may reduce the number of CD34/CD133
double-positive cells in a mouse tibial tumor in situ.
CD34/CD133 double-positive cells are considered to be EPCs
(53). The melittin-treated tumors
were also observed to have a lower MVD when compared with that of
the controls. MVD is assessed using CD105, which is a vascular
endothelial cell proliferation marker (54). CD105 is more effective than other
endothelial cell markers (such as CD31 CD34, factor VIII-related
antigen) as it is expressed only in the vascular endothelial cells
of tumor tissues and not in the vessels of healthy tissues
(55). This characteristic allows
only the angiogenesis process in tumors to be assessed. The present
study demonstrated that melittin may effectively reduce
osteosarcoma-induced angiogenesis.
There were certain limitations of the present study.
It was performed in animals and further studies in animals are
required prior to eventual clinical trials. In addition, the
present study was not designed to determine the exact mechanisms
underlying EPC-modulated angiogenesis in osteosarcoma. However, the
current study may aid in the design of novel studies to address
this issue.
In conclusion, the present study demonstrates that
the SDF-1α/CXCR4 signaling pathway is important in EPC-modulated
tumor angiogenesis. Melittin decreased EPC viability, migration and
tube formation, and decreased the expression levels of p-AKT and
ERK1/2. Further studies are required to assess the effects of
melittin on angiogenesis modulated by EPCs in osteosarcoma.
Acknowledgments
The present study was supported by the National
Natural Science Foundation of China (grant no. 30873275).
References
|
1
|
Picci P: Osteosarcoma (osteogenic
sarcoma). Orphanet J Rare Dis. 2:62007. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Eilber F, Giuliano A, Eckardt J, Patterson
K, Moseley S and Goodnight J: Adjuvant chemotherapy for
osteosarcoma: A randomized prospective trial. J Clin Oncol.
5:21–26. 1987.PubMed/NCBI
|
|
3
|
Bernthal NM, Federman N, Eilber FR, Nelson
SD, Eckardt JJ, Eilber FC and Tap WD: Long-term results (>25
years) of a randomized, prospective clinical trial evaluating
chemotherapy in patients with high-grade, operable osteosarcoma.
Cancer. 118:5888–5893. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Whelan JS, Jinks RC, McTiernan A, Sydes
MR, Hook JM, Trani L, Uscinska B, Bramwell V, Lewis IJ, Nooji MA,
et al: Survival from high-grade localised extremity osteosarcoma:
Combined results and prognostic factors from three European
Osteosarcoma Intergroup randomised controlled trials. Ann Oncol.
23:1607–1616. 2012. View Article : Google Scholar :
|
|
5
|
Pluda JM: Tumor-associated angiogenesis:
Mechanisms, clinical implications, and therapeutic strategies.
Semin Oncol. 24:203–218. 1997.PubMed/NCBI
|
|
6
|
de Bont ES, Guikema JE, Scherpen F,
Meeuwsen T, Kamps WA, Vellenga E and Bos NA: Mobilized human CD34+
hematopoietic stem cells enhance tumor growth in a nonobese
diabetic/severe combined immunodeficient mouse model of human
non-Hodgkin's lymphoma. Cancer Res. 61:7654–7659. 2001.PubMed/NCBI
|
|
7
|
Duan HX, Cheng LM, Wang J, Hu LS and Lu
GX: Angiogenic potential difference between two types of
endothelial progenitor cells from human umbilical cord blood. Cell
Biol Int. 30:1018–1027. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Asahara T, Masuda H, Takahashi T, Kalka C,
Pastore C, Silver M, Kearne M, Magner M and Isner JM: Bone marrow
origin of endothelial progenitor cells responsible for postnatal
vasculogenesis in physiological and pathological
neovascularization. Circ Res. 85:221–228. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Kryczek I, Wei S, Keller E, Liu R and Zou
W: Stroma-derived factor (SDF-1/CXCL12) and human tumor
pathogenesis. Am J Physiol Cell Physiol. 292:C987–C995. 2007.
View Article : Google Scholar
|
|
10
|
Vajkoczy P, Blum S, Lamparter M,
Mailhammer R, Erber R, Engelhardt B, Vestweber D and Hatzopoulos
AK: Multistep nature of microvascular recruitment of ex
vivo-expanded embryonic endothelial progenitor cells during tumor
angiogenesis. J Exp Med. 197:1755–1765. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Mukai N, Akahori T, Komaki M, Li Q,
Kanayasu-Toyoda T, Ishii-Watabe A, Kobayashi A, Yamaguchi T, Abe M,
Amagasa T and Morita I: A comparison of the tube forming potentials
of early and late endothelial progenitor cells. Exp Cell Res.
314:430–440. 2008. View Article : Google Scholar
|
|
12
|
Uehara F, Tome Y, Miwa S, Hiroshima Y,
Yano S, Yamamoto M, Mii S, Maehara H, Bouvet M, Kanaya F and
Hoffman RM: Osteosarcoma cells enhance angiogenesis visualized by
color-coded imaging in the in vivo Gelfoam® assay. J
Cell Biochem. 115:1490–1494. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Mueller SM, Mizuno S, Gerstenfeld LC and
Glowacki J: Medium perfusion enhances osteogenesis by murine
osteosarcoma cells in three-dimensional collagen sponges. J Bone
Miner Res. 14:2118–2126. 1999. View Article : Google Scholar
|
|
14
|
Kumarasuriyar A, Murali S, Nurcombe V and
Cool SM: Glycosaminoglycan composition changes with MG-63
osteosarcoma osteogenesis in vitro and induces human mesenchymal
stem cell aggregation. J Cell Physiol. 218:501–511. 2009.
View Article : Google Scholar
|
|
15
|
Bonig H, Priestley GV, Oehler V and
Papayannopoulou T: Hematopoietic progenitor cells (HPC) from
mobilized peripheral blood display enhanced migration and marrow
homing compared to steady-state bone marrow HPC. Exp Hematol.
35:326–334. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Müller A, Homey B, Soto H, Ge N, Catron D,
Buchanan ME, McClanahan T, Murphy E, Yuan W, Wagner SN, et al:
Involvement of chemokine receptors in breast cancer metastasis.
Nature. 410:50–56. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Kim J, Mori T, Chen SL, Amersi FF,
Martinez SR, Kuo C, Turner RR, Ye X, Bilchik AJ, Morton DL and Hoon
DS: Chemokine receptor CXCR4 expression in patients with melanoma
and colorectal cancer liver metastases and the association with
disease outcome. Ann Surg. 244:113–120. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Sun YX, Schneider A, Jung Y, Wang J, Dai
J, Wang J, Cook K, Osman NI, Koh-Paige AJ, Shim H, et al: Skeletal
localization and neutralization of the SDF-1(CXCL12)/CXCR4 axis
blocks prostate cancer metastasis and growth in osseous sites in
vivo. J Bone Miner Res. 20:318–329. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Petit I, Jin D and Rafii S: The
SDF-1-CXCR4 signaling pathway: A molecular hub modulating
neo-angiogenesis. Trends Immunol. 28:299–307. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Oren Z and Shai Y: Selective lysis of
bacteria but not mammalian cells by diastereomers of melittin:
Structure-function study. Biochemistry. 36:1826–1835. 1997.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Kubo H, Loegering DA, Adolphson CR and
Gleich GJ: Cytotoxic properties of eosinophil granule major basic
protein for tumor cells. Int Arch Allergy Immunol. 118:426–428.
1999. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Lazarev VN, Parfenova TM, Gularyan SK,
Misyurina OY, Akopian TA and Govorun VM: Induced expression of
melittin, an antimicrobial peptide, inhibits infection by Chlamydia
trachomatis and Mycoplasma hominis in a HeLa cell line. Int J
Antimicrob Agents. 19:133–137. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Li B, Gu W, Zhang C, Huang XQ, Han KQ and
Ling CQ: Growth arrest and apoptosis of the human hepatocellular
carcinoma cell line BEL-7402 induced by melittin. Onkologie.
29:367–371. 2006.PubMed/NCBI
|
|
24
|
Yeo SW, Seo JC and Choi YH: Induction of
the growth inhibition and apoptosis by bee venom in human breast
carcinoma MCF-7 cells. J Kor Acup Mox Soc. 20:45–62. 2003.
|
|
25
|
Jang MH, Shin MC, Lim S, Han SM, Park HJ,
Shin I, Lee JS, Kim KA, Kim EH and Kim CJ: Bee venom induces
apoptosis and inhibits expression of cyclooxygenase-2 mRNA in human
lung cancer cell line NCI-H1299. J Pharmacol Sci. 91:95–104. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Huh JE, Baek YH, Lee MH, Choi DY, Park DS
and Lee JD: Bee venom inhibits tumor angiogenesis and metastasis by
inhibiting tyrosine phosphorylation of VEGFR-2 in LLC-tumor-bearing
mice. Cancer Lett. 292:98–110. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Cho HJ, Jeong YJ, Park KK, Park YY, Chung
IK, Lee KG, Yeo JH, Han SM, Bae YS and Chang YC: Bee venom
suppresses PMA-mediated MMP-9 gene activation via JNK/p38 and
NF-kappaB-dependent mechanisms. J Ethnopharmacol. 127:662–668.
2010. View Article : Google Scholar
|
|
28
|
Liu S, Yu M, He Y, Xiao L, Wang F, Song C,
Sun S, Ling C and Xu Z: Melittin prevents liver cancer cell
metastasis through inhibition of the Rac1-dependent pathway.
Hepatology. 47:1964–1973. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Guo W, Feng JM, Yao L, Sun L and Zhu GQ:
Transplantation of endothelial progenitor cells in treating rats
with IgA nephropathy. BMC Nephrol. 15:1102014. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Quaranta P, Antonini S, Spiga S, Mazzanti
B, Curcio M, Mulas G, Diana M, Marzola P, Mosca F and Longoni B:
Co-transplantation of endothelial progenitor cells and pancreatic
islets to induce long-lasting normoglycemia in
streptozotocin-treated diabetic rats. PLoS One. 9:e947832014.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Massa M, Rosti V, Ramajoli I, Campanelli
R, Pecci A, Viarengo G, Meli V, Marchetti M, Hoffman R and Barosi
G: Circulating CD34+, CD133+, and vascular endothelial growth
factor receptor 2-positive endothelial progenitor cells in
myelofibrosis with myeloid metaplasia. J Clin Oncol. 23:5688–5695.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Fritzenwanger M, Lorenz F, Jung C, Fabris
M, Thude H, Barz D and Figulla HR: Differential number of CD34+,
CD133+ and CD34+/CD133+ cells in peripheral blood of patients with
congestive heart failure. Eur J Med Res. 14:113–117.
2009.PubMed/NCBI
|
|
33
|
Fisher JL, Mackie PS, Howard ML, Zhou H
and Choong PF: The expression of the urokinase plasminogen
activator system in metastatic murine osteosarcoma: An in vivo
mouse model. Clin Cancer Res. 7:1654–1660. 2001.PubMed/NCBI
|
|
34
|
Cai KX, Tse LY, Leung C, Tam PK, Xu R and
Sham MH: Suppression of lung tumor growth and metastasis in mice by
adeno-associated virus-mediated expression of vasostatin. Clin
Cancer Res. 14:939–949. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Weidner N, Semple JP, Welch WR and Folkman
J: Tumor angiogenesis and metastasis--correlation in invasive
breast carcinoma. N Engl J Med. 324:1–8. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Ta HT, Dass CR, Choong PF and Dunstan DE:
Osteosarcoma treatment: State of the art. Cancer Metastasis Rev.
28:247–263. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Xu S, Wen H and Jiang H: Urotensin II
promotes the proliferation of endothelial progenitor cells through
p38 and p44/42 MAPK activation. Mol Med Rep. 6:197–200.
2012.PubMed/NCBI
|
|
38
|
Park JH, Jeong YJ, Park KK, Cho HJ, Chung
IK, Min KS, Kim M, Lee KG, Yeo JH, Park KK and Chan YC: Melittin
suppresses PMA-induced tumor cell invasion by inhibiting NF-kappaB
and AP-1-dependent MMP-9 expression. Mol Cells. 29:209–215. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Fan Q, Hu Y, Pang H, Sun J, Wang Z and Li
J: Melittin protein inhibits the proliferation of MG63 cells by
activating inositol-requiring protein-1α and X-box binding protein
1-mediated apoptosis. Mol Med Rep. 9:1365–1370. 2014.PubMed/NCBI
|
|
40
|
Gao H, Priebe W, Glod J and Banerjee D:
Activation of signal transducers and activators of transcription 3
and focal adhesion kinase by stromal cell-derived factor 1 is
required for migration of human mesenchymal stem cells in response
to tumor cell-conditioned medium. Stem Cells. 27:857–865. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Huang CY, Lee CY, Chen MY, Yang WH, Chen
YH, Chang CH, Hsu HC, Fong YC and Tang CH: Stromal cell-derived
factor-1/CXCR4 enhanced motility of human osteosarcoma cells
involves MEK1/2, ERK and NF-kappaB-dependent pathways. J Cell
Physiol. 221:204–212. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Petit I, Goichberg P, Spiegel A, Peled A,
Brodie C, Seger R, Nagler A, Alon R and Lapidot T: Atypical
PKC-zeta regulates SDF-1-mediated migration and development of
human CD34+ progenitor cells. J Clin Invest. 115:168–176. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Lu DY, Tang CH, Yeh WL, Wong KL, Lin CP,
Chen YH, Lai CH, Chen YF, Leung YM and Fu WM: SDF-1alpha
up-regulates interleukin-6 through CXCR4, PI3K/Akt, ERK, and
NF-kappaB-dependent pathway in microglia. Eur J Pharmacol.
613:146–154. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Smadja DM, Bièche I, Uzan G, Bompais H,
Muller L, Boisson-Vidal C, Vidaud M, Aiach M and Gaussem P: PAR-1
activation on human late endothelial progenitor cells enhances
angiogenesis in vitro with upregulation of the SDF-1/CXCR4 system.
Arterioscler Thromb Vasc Biol. 25:2321–2327. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Walter DH, Haendeler J, Reinhold J,
Rochwalsky U, Seeger F, Honold J, Hoffman J, Urbich C, Lehmann R,
Arenza-Seisdesdos F, et al: Impaired CXCR4 signaling contributes to
the reduced neovascularization capacity of endothelial progenitor
cells from patients with coronary artery disease. Circ Res.
97:1142–1151. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Maksym RB, Tarnowski M, Grymula K,
Tarnowksa J, Wysoczynski M, Liu R, Czerny B, Ratajczak J, Kucia M
and Ratajczak MZ: The role of stromal-derived factor-1--CXCR7 axis
in development and cancer. Eur J Pharmacol. 625:31–40. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Yamaguchi J, Kusano KF, Masuo O, Kawamoto
A, Silver M, Murasawa S, Bosch-Marce M, Masuda H, Losordo DW, Isner
JM and Asahara T: Stromal cell-derived factor-1 effects on ex vivo
expanded endothelial progenitor cell recruitment for ischemic
neovascularization. Circulation. 107:1322–1328. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Loetscher M, Geiser T, O'Reilly T, Zwahlen
R, Baggiolini M and Moser B: Cloning of a human seven-transmembrane
domain receptor, LESTR, that is highly expressed in leukocytes. J
Biol Chem. 269:232–237. 1994.PubMed/NCBI
|
|
49
|
Folkman J: Tumor angiogenesis. Cancer
Medicine. Holland JF, Bast RC and Morton DL: 4th edition. Williams
& Wilkins; Baltimore: pp. 181–204. 1997
|
|
50
|
Lin F, Zheng SE, Shen Z, Tang LN, Chen P,
Sun YJ, Zhao H and Yao Y: Relationships between levels of CXCR4 and
VEGF and blood-borne metastasis and survival in patients with
osteosarcoma. Med Oncol. 28:649–653. 2011. View Article : Google Scholar
|
|
51
|
Zhang P, Dong L, Yan K, Long H, Yang TT,
Dong MQ, Zhou Y, Fan QY and Ma BA: CXCR4-mediated osteosarcoma
growth and pulmonary metastasis is promoted by mesenchymal stem
cells through VEGF. Oncol Rep. 30:1753–1761. 2013.PubMed/NCBI
|
|
52
|
Jeong YJ, Choi Y, Shin JM, Cho HJ, Kang
JH, Park KK, Choe JY, Bae YS, Han SM, Kim CH, Chang HW and Chang
YC: Melittin suppresses EGF-induced cell motility and invasion by
inhibiting PI3K/Akt/mTOR signaling pathway in breast cancer cells.
Food Chem Toxicol. 68:218–225. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Zhu G, Song M, Wang H, Zhao G, Yu Z, Yin
Y, Zhao X and Huang L: Young environment reverses the declined
activity of aged rat-derived endothelial progenitor cells:
involvement of the phosphatidylinositol 3-kinase/Akt signaling
pathway. Ann Vasc Surg. 23:519–534. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Brewer CA, Setterdahl JJ, Li MJ, Johnston
JM, Mann JL and McAsey ME: Endoglin expression as a measure of
microvessel density in cervical cancer. Obstet Gynecol. 96:224–228.
2000.PubMed/NCBI
|
|
55
|
Wikström P, Lissbrant IF, Stattin P,
Egevad L and Bergh A: Endoglin (CD105) is expressed on immature
blood vessels and is a marker for survival in prostate cancer.
Prostate. 51:268–275. 2002. View Article : Google Scholar : PubMed/NCBI
|