Introduction
Tendinopathy is a strain-induced chronic disease
whose causes remain to be elucidated. The frequency of cases of
tendinopathy is increasing, and it currently accounts for ~50% of
all sports injuries (1). As
conservative treatments are limited, they are unsuccessful in
24–45.5% of patients, leading to refractory tendinopathy (2). Therefore, an enhanced understanding
of the pathogenesis of tendinopathy at the cellular and molecular
level is required to develop novel and more effective treatment
strategies.
Tendon stem cells (TSCs) are a type of mesenchymal
stem cells derived from the paratenon of the tendon. Zhang and Wang
(3) indicated that TSCs have a
higher potential for cell proliferation and collagen synthesis as
compared with that of mature tendon cells. Under certain
conditions, TSCs can be differentiated into mature tendon cells
with an enhanced secretion of extracellular matrix to effectively
repair tendon injury (3). However,
the underlying mechanisms of TSC differentiation have remained
elusive. TSCs are pluripotent and thus capable of differentiating
into adipocytes, chondrocytes and osteoblasts (4–8).
Their multi-directional differentiation potential has an important
role in the process of tendon repair. Insight into the regulation
of TSC differentiation may provide novel strategies for the
treatment of tendinopathy, as these cells may be utilized for
tendon injury repair.
Sirt1 is a nicotinamide adenine dinucleotide
(NAD+)-dependent histone deacetylase, which has an
important role in energy metabolism and cell differentiation
(9). A study has shown that Sirt1
can regulate osteogenic differentiation by self-deacetylation in
mesenchymal stem cells (10).
Resveratrol, an activator of Sirt1, can decrease the number of
adipocytes and increase the expression of osteoblast markers
(11). Inhibition of Sirt1 can
increase the number of adipocytes and lipoblast markers and reduce
the expression of osteoblast markers. Thus, the activation of Sirt1
can inhibit adipogenic differentiation and promote osteogenic
differentiation (11). However,
the expression status of Sirt1 in TSCs and the underlying
mechanisms of its possible regulation of osteogenic and adipogenic
differentiation in TSCs have remained elusive.
Using reverse-transcription quantitative polymerase
chain reaction (RT-qPCR) and western blot analyses, the present
study assessed the gene and protein expression of Sirt1 in TSCs
following treatment with a Sirt1 activator or inhibitor for various
time periods. In addition, the present study assessed the resulting
differentiation into osteoblasts and lipoblasts and elucidated the
molecular mechanism of Sirt1-mediated regulation of osteogenic and
adipogenic differentiation of TSCs. The present study provided
insight into the molecular mechanisms of Sirt1-induced TSC
differentiation, which may be harnessed for the development of
novel and effective treatments for tendon injury.
Materials and methods
Isolation and culture of rat TSCs
Three eight-week-old Sprague-Dawley male rats
weighing 250–300 g were purchased from the Shanghai Institute of
Materia Medica, Chinese Academy of Science (Shanghai, China) and
used in the present study. All experiments were approved by the
Animal Research Ethics Committee of the Third Military Medical
University (Chongqing, China). The isolation and culture of rat
TSCs were performed as described previously (12–14).
Culture medium with or without Sirt1 activator SRT1720 (0.04 or
0.02 µM; Sigma-Aldrich, St. Louis, MO, USA) or Sirt1
inhibitor EX527 (0.02 µM; Santa Cruz Biotechnology, Inc.,
Dallas, TX, USA) was changed every three days throughout the
experiments. Analyses were performed on days 1, 3, 7, 10 and 14 of
incubation.
RT-Qpcr
RNA was extracted with TRIzol reagent (Invitrogen;
Thermo Fisher Scientific Inc., Waltham, MA, USA; 1 ml per
1×107 cells) and cDNA was synthesized using the RNA
reverse transcription kit (cat no. DRR047A; Takara, Dalian, China)
according to the manufacturer's instructions. The sequences of the
Sirt1 gene primers were 5′-CCAGATCCTCAAGCCATG T-3′ for the forward
primer and 5′-TTGGATTCCTGCAACCTG-3′ for the reverse primer. The
sequences of the Runx2 gene primers were
5′-TGATGACACTGCCACCTCTGACTT-3′ for the forward primer and
5′-TGGATAGTGCATTCGTGGGTTGGA-3′ for the reverse primer. The
sequences of the PPARγ gene primers were 5′-ATGACCACTCCCATTCCTTT-3′
for the forward primer and 5′-TGATCGCACTTTGGTATTCTT-3′ for the
reverse primer. The sequences of the GAPDH primers were
5′-ATGGAAATCCCATCACCATCTT-3′ for the forward primer and
5′-CGCCCCACTTGATTTTGG-3′ for the reverse primer. The primers were
obtained from Sangon Biotech Co. Ltd. (Shanghai, China). A total of
5 µl cDNA from each sample was obtained and PCR was
performed in 25 µl reactions containing Platinum SYBR Green
qPCR SuperMix-UDG and appropriate primers in the ABI StepOne Plus
system from Applied Biosystems (Foster City, CA, USA). Cycling
parameters were as follows: Denaturation, 95°C for 10 min followed
by 45 cycles consisting of denaturation at 95°C for 20 sec,
annealing at 60°C for 25 sec, extension at 72°C for 30 sec, and a
controlled increase to 95°C with a heating rate of 0.1°C/sec.
Target gene expression was normalized to that of GAPDH. Data were
analyzed using the comparative threshold cycle (Cq) method. The
melting curves were analyzed after amplification. PCR reactions of
each sample were performed in triplicate. Data were analyzed using
the comparative 2−ΔΔCq method.
Protein extraction and western blot
analysis
Cells were scraped and homogenized in lysis buffer
(Beyotime Institute of Biotechnology, Haimen, China) containing a
mixture of proteinase inhibitors (Thermo Fisher Scientific Inc.).
Protein concentrations were measured using a bicinchoninic acid
protein assay kit (Beyotime Institute of Biotechnology). Protein
samples (40 µg/lane) were resolved by 10% SDS-PAGE and
transferred onto polyvinylidene difluoride membranes. After
blocking with 0.1% Tris-buffered saline containing Tween 20
(Beyotime Institute of Biotechnology) and 5% non-fat milk for 1 h
at 20°C, membranes were incubated at 4°C for 1 h sequentially with
primary and secondary antibodies. The following primary antibodies
purchased from Abcam (Cambridge MA, USA) were used: Anti-Sirt1
(monoclonal mouse IgG; 1:1,000 dilution; cat. no. ab50517),
anti-β-catenin (monoclonal mouse IgG; 1:800 dilution; cat. no.
ab22656), anti-Runt-related transcription factor 2 (Runx2;
monoclonal mouse IgG; 1:1,000 dilution; cat. no. ab115899),
anti-CCAAT/enhancer binding protein (CEBP)α (monoclonal mouse IgG;
1:1,000 dilution; cat. no. ab128482) and anti-peroxisome
proliferator-activated receptor (PPAR)γ (monoclonal mouse IgG;
1:1200 dilution; cat. no. ab138004), anti-PI3K (monoclonal mouse;
1:1,000 dilution; cat. no. ab86714), anti-AKT (monoclonal rabbit
IgG; 1:1,500 dilution; cat. no. ab32505), anti-BMP2 (monoclonal
rabbit IgG; 1:1,000 dilution; cat. no. ab183729). Subsequently, the
membranes were incubated at room temperature for 2 h with
peroxidase-labeled anti-mouse IgG antibody (1:1,000 dilution; cat.
no. A0216; Beyotime Institute of Biotechnology) or anti-rabbit IgG
antibody (1:1,000 dilution; cat. no. A0208; Beyotime Institute of
Biotechnology). Proteins were visualized using an enhanced
chemiluminescence (ECL) reagent (Amersham ECL Prime detection; GE
Healthcare, Little Chalfont, UK). Experiments were performed in
triplicate.
Alizarin red staining
Cells were fixed in 70% ethanol for 1 h and stained
with 2% alizarin red solution (Beyotime Institute of Biotechnology;
pH 4.1–4.5) for 30 min at 20°C. The stained cells were examined
under an inverted microscope (ix71; Olympus Corporation, Tokyo,
Japan). Photomicrographs were obtained using a charge-coupled
device camera.
Oil Red O staining
Adipocytes were identified by staining with Oil Red
O (Sigma-Aldrich) as described previously (15). The area of the cells stained with
Oil Red O was measured using ImagePro Plus 6.0 software (Media
Cybernetics, Rockville, MD, USA).
Statistical analysis
SPSS version 17.0 (SPSS, Inc, Chicago, IL, USA)
software was used for statistical analysis. Values are expressed as
the mean + standard deviation. Multiple comparisons were made using
one-way analysis of variance followed by Fisher's exact test.
P<0.05 was considered to indicate a statistically significant
difference between values.
Results
Sirt1 is expressed in TSCs and drives
osteogenic differentiation
In order to investigate the association between
Sirt1 and the osteogenic and adipogenic differentiation of TSCs,
the present study first detected Sirt1 mRNA and protein expression
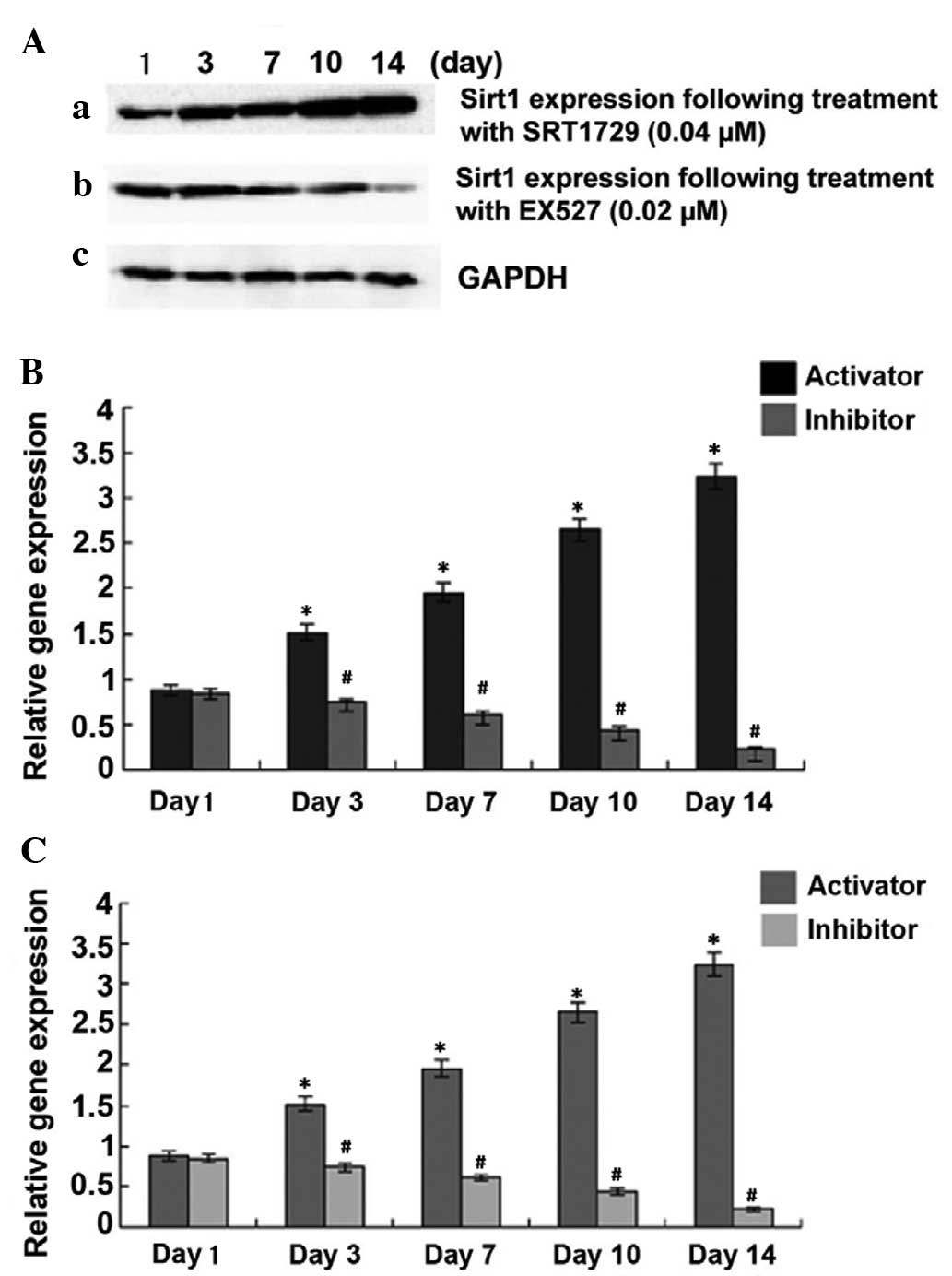
in TSCs. As shown in Fig. 1, TSCs
expressed Sirt1, which was further enhanced by Sirt1 activator
SRT1720 in a concentration- and time-dependent manner. Conversely,
Sirt1 expression decreased in the presence of Sirt1 inhibitor EX527
in a time-dependent manner (P<0.05). Next, the present study
evaluated the expression of adipogenic and osteogenic
differentiation markers in TSCs on days 3, 7, 10 and 14 of
treatment with Sirt1 activator and inhibitor. The expression of
BMP2 and Runx2 appeared to be positively correlated with Sirt1
expression, which indicated that Sirt1 increases the capacity of
TSCs for osteogenic differentiation (Fig. 2). Peak values of BMP2 and Runx2
expression were observed on day 10, followed by a marked decrease
in osteogenic differentiation capacity of TSCs during prolonged
treatment with Sirt1 activator. However, the adipogenic
differentiation capacity of TSCs was gradually reduced
(P<0.05).
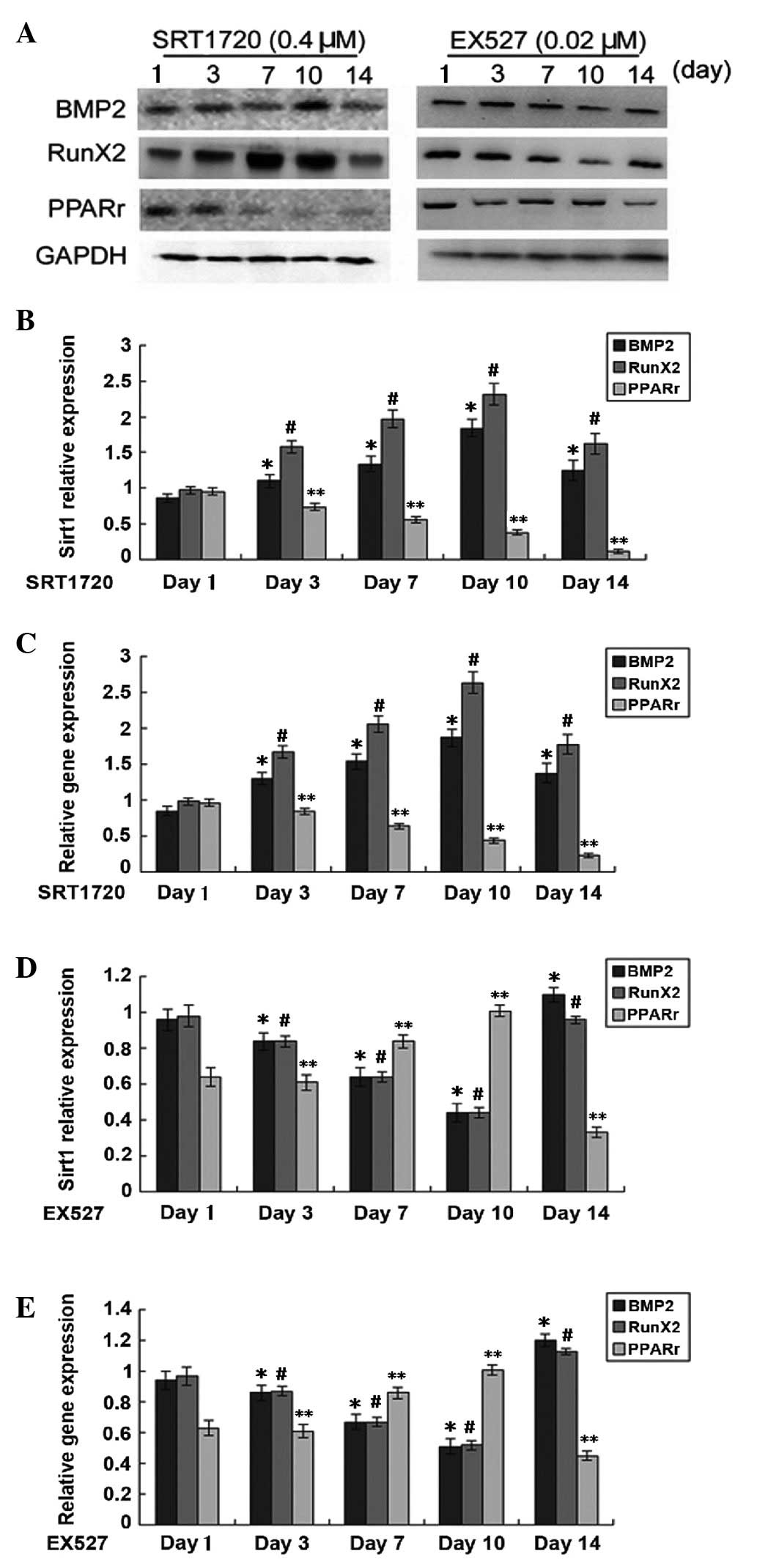 | Figure 2Expression of marker proteins of
osteogenic and adipogenic differentiation on days 3, 7, 10 and 14
after treatment with sirtuin 1 activator SRT1720 (0.4 µM)
and inhibitor EX527 (0.02 µM). (A) Representative western
blots of BMP2, RunX2 and PPARγ. (B) Protein levels following
SRT1720 treatment were quantified by grey value analysis of A with
normalization to GAPDH. (C) Reverse-transcription quantitative
polymerase chain reaction analysis of mRNA levels of BMP2, RunX2
and PPARγ following SRT1720 treatment. (D) Protein levels following
EX527 treatment were quantified by grey value analysis of A with
normalization to GAPDH. (E) Reverse-transcription quantitative
polymerase chain reaction analysis of mRNA levels of BMP2, RunX2
and PPARγ following EX527 treatment. Values are expressed as the
mean ± standard deviation. *P<0.05, compared with the
control group (BMP2); #P<0.05, compared with the
control group (RunX2); **P<0.05, compared with the
control group (PPARγ). BMP, bone morphogenetic protein; RunX2,
Runt-related transcription factor 2; PPAR, peroxisome
proliferator-activated receptor. |
Sirt1 enhances osteogenic and decreases
adipogenic differentiation of tendon stem cells
Alizarin red staining was used to assess the
osteogenic differentiation and oil red O staining was used to
detect the adipogenic differentiation of tendon stem cells after
treatment with SRT1720 (0.4 µM) (Fig. 3). In accordance with the effects of
Sirt1 on the expression of differentiation markers (Fig. 2), staining showed that the capacity
for osteogenic differentiation increased, while the capacity for
adipogenic differentiation decreased following treatment with Sirt1
activator (0.4 µM). Sirt1 treatment was observed to exert
the highest effect on TSC differentiation on day 10.
Sirt1 drives osteogenic differentiation
of TSCs via upregulating β-catenin and Runx2 and represses
adipogenic differentiation of TSCs through downregulating CEBPα and
PPARγ and the PI3K/AKT pathway
The present study showed that increases in Sirt1
expression in TSCs enhanced β-catenin and Runx2 expression, while
the expression of PI3K, AKT, CEBPα and PPARγ was decreased
(Fig. 4). Furthermore, treatment
with Sirt1 inhibitor reduced not only Sirt1 expression but also
β-catenin and Runx2 expression, while enhancing the expression of
PI3K, AKT, CEBPα and PPARγ. These results indicated that Sirt1
promotes osteogenic differentiation of TSCs through upregulating
β-catenin and Runx2 expression, while it inhibits the expression of
CEBPα and PPARγ as well as the PI3K/AKT pathway, thereby inhibiting
the adipogenic differentiation of TSCs (P<0.05).
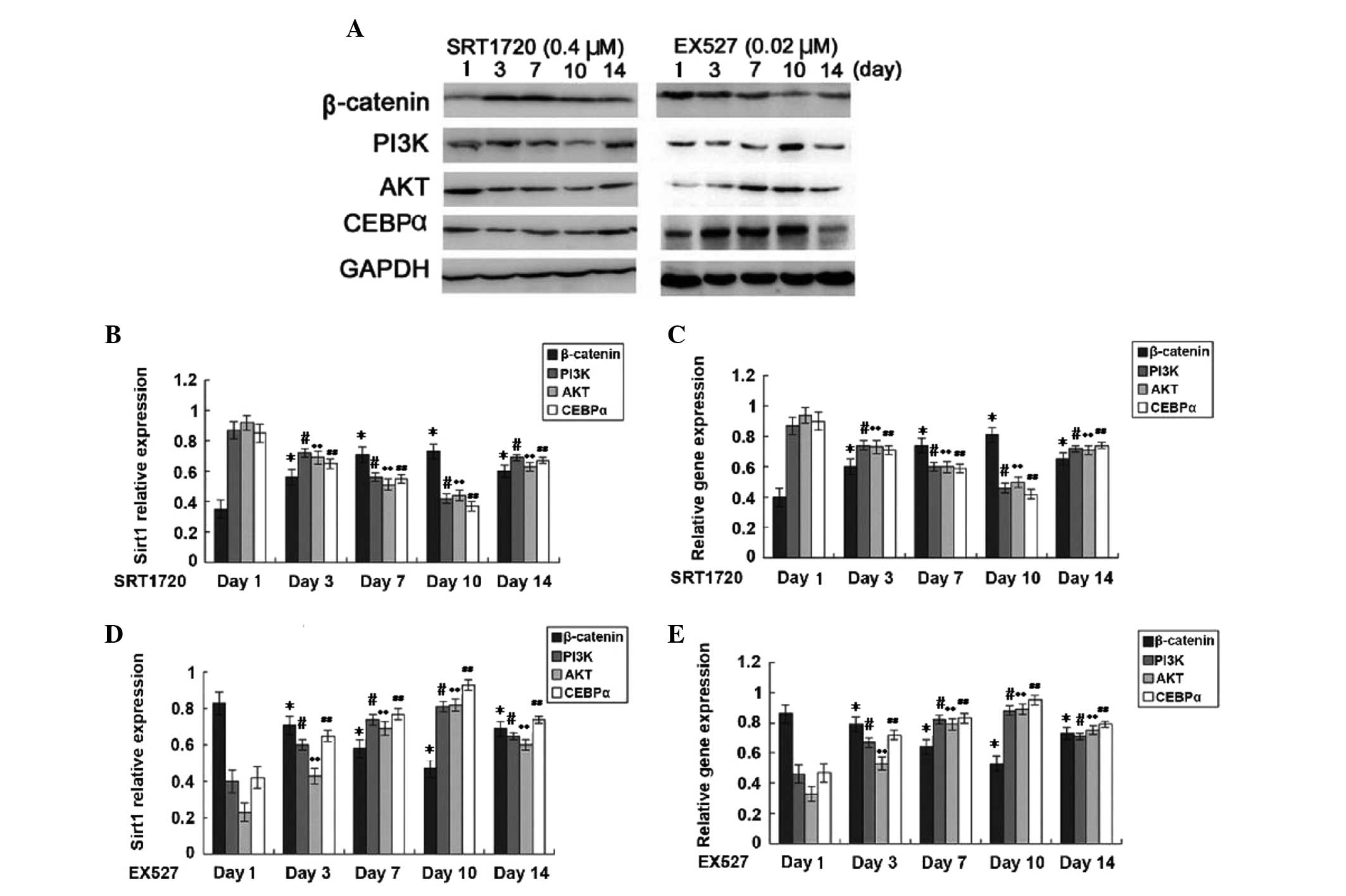 | Figure 4Expression of proteins and genes
involved in signaling pathways on days 3, 7, 10 and 14 of treatment
with sirtuin 1 activator SRT1720 (0.4 µM) or inhibitor EX527
(0.02 µM). (A) Representative western blots of β-catenin,
PI3K, AKT and CEBPα. (B) Protein levels following SRT1720 treatment
were quantified by grey value analysis of A with normalization to
GAPDH. (C) Reverse-transcription quantitative polymerase chain
reaction analysis of mRNA levels of β-catenin, PI3K, AKT and CEBPα
following SRT1720 treatment. (D) Protein levels following EX527
treatment were quantified by grey value analysis of A with
normalization to GAPDH. (E) Reverse-transcription quantitative
polymerase chain reaction analysis of mRNA levels of β-catenin,
PI3K, AKT and CEBPα following EX527 treatment. Values are expressed
as the mean ± standard deviation. *P<0.05, compared
with the control group (β-catenin); #P<0.05, compared
with the control group (PI3K); **P<0.05, compared
with the control group (AKT); ##P<0.05, compared with
the control group (CEBPα). PI3K, phosphoinositide-3 kinase; CEBPα,
CCAAT/enhancer binding protein α. |
Discussion
Sirt1 is an NAD-dependent deacetylase which is
important in numerous biological processes, including cell cycle
regulation, cell differentiation, cell survival and apoptosis.
Sirt1 regulates osteogenic differentiation of mesenchymal stem
cells by deacetylating H3K9 in the promoter sequence of the Sost
gene, which encodes sclerostin (10).
The present study confirmed that Sirt1 is expressed
in TSCs and revealed that it enhances their osteogenic
differentiation capacity in a time-dependent manner. Furthermore,
the gene and protein levels of osteogenesis marker BMP2, were
elevated by Sirt1 in a time-dependent manner. Peak values of BMP2
expression were observed on day 10 followed of treatment with Sirt1
activator. Conversely, the osteogenic differentiation capacity of
TSCs was decreased after treatment with Sirt1 inhibitor. By
contrast, treatment with Sirt1 activator reduced the adipogenic
capacity of TSCs, while it was increased by the Sirt1
inhibitor.
Recently, Simic et al (16) showed that Sirt1 deacetylates Runx2
and β-catenin to regulate the differentiation of mesenchymal stem
cells. Sirt1 activated by resveratrol was reported to promote
osteogenic differentiation of mesenchymal stem cells by increasing
Runx2 expression, increasing its combination with PPARγ and
inhibiting the activity of PPARγ through its cofactor nuclear
receptor co-repressor 1 (NCoR1) (14). PPARγ has an important role in the
balance of adipogenic and osteogenic differentiation of mesenchymal
stem cells (15). Overexpressed
Sirt1 can be anchored to NCoR1 and -2, the inhibitory co-factors of
PPARγ, which in turn suppresses the expression of PPARγ and C/EBPα
and further inhibits the cytopoiesis of adipocytes (i.e. it reduces
the transformation of pre-adipocytes into adipocytes) (14). Furthermore, Sirt1 can inhibit the
transcriptional activity of AKT2 and interact with AKT2 and CEBPα
(16). Silencing of AKT2 and/or
Sirt1 was shown to affect pre-adipogenesis through the PI3K/AKT
pathway, which exhibits a synergy with the CEBPα pathway (16). The present study explored whether
Sirt1 regulates the differentiation of TSCs in a similar manner to
that of mesenchymal stem cells.
In the present study, it was revealed that increased
Sirt1 expression enhanced β-catenin and Runx2 expression in TSCs
and reduced the expression of PI3K, AKT, CEBPα and PPARγ, while
treatment with Sirt1 inhibitor had the opposite effect. It is
therefore concluded that Sirt1 promotes osteogenic differentiation
of TSCs through upregulation of β-catenin and Runx2, while
inhibiting adipogenic differentiation of TSCs via inhibiting the
PI3K/AKT pathway as well as the expression of CEBPα and PPARγ.
In conclusion, to the best of our knowledge, the
present study was the first to report that Sirt1 is endogenously
expressed in TSCs. Sirt1 was demonstrated to time-dependently
promote the osteogenic differentiation of TSCs through upregulating
β-catenin and Runx2 and to inhibit the adipogenic differentiation
of TSCs through inhibiting the PI3K/AKT pathway with downregulation
of CEBPα and PPARγ. During the process of tendon injury repair,
TSCs favor differentiation into cartilage and bone, while the
accumulation of adipose tissue is not conducive to the recovery
from tendinopathy. Therefore, by targeting Sirt1, it may be
possible to regulate osteogenic differentiation of TSCs as well as
adipose accumulation in injured tendons. Upregulation of Sirt1 may
be an efficient strategy for the treatment of refractory
tendinopathy using TSCs. This approach may represent a breakthrough
leading to the development of novel clinical techniques with the
potential to fundamentally improve the treatment of sports
injuries.
Acknowledgments
The present study was supported by the National
Natural Science Foundation of China (grant nos. 81230040 and
81401831).
References
|
1
|
Mahieu NN, Witvrouw E, Stevens V, Van
Tiggelen D and Roget P: Intrinsic risk factors for the development
of achilles tendon overuse injury: A prospective study. Am J Sports
Med. 34:226–235. 2006. View Article : Google Scholar
|
|
2
|
Maffulli N: Achilles tendon pathology,
present and future. Br J Sports Med. 45:e2 View Article : Google Scholar : 2011.
|
|
3
|
Zhang J and Wang JH: Characterization of
differential properties of rabbit tendon stem cells and tenocytes.
BMC Musculoskelet Disord. 11:102010. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Bi Y, Ehirchiou D, Kilts TM, Inkson CA,
Embree MC, Sonoyama W, Li L, Leet AI, Seo BM, Zhang L, et al:
Identification of tendon stem/progenitor cells and the role of the
extracellular matrix in their niche. Nat Med. 13:1219–1227. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Rui YF, Lui PP, Li G, Fu SC, Lee YW and
Chan KM: Isolation and characterization of multipotent rat
tendon-derived stem cells. Tissue Eng Part A. 16:1549–1558. 2010.
View Article : Google Scholar
|
|
6
|
Zhang J and Wang JH: Production of PGE(2)
increases in tendons subjected to repetitive mechanical loading and
induces differentiation of tendon stem cells into non-tenocytes. J
Orthop Res. 28:198–203. 2010.
|
|
7
|
de Mos M, Koevoet WJ, Jahr H, Verstegen
MM, Heijboer MP, Kops N, van Leeuwen JP, Weinans H, Verhaar JA and
van Osch GJ: Intrinsic differentiation potential of adolescent
human tendon tissue: An in-vitro cell differentiation study. BMC
Musculoskelet Disord. 8:162007. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Salingcarnboriboon R, Yoshitake H, Tsuji
K, Obinata M, Amagasa T, Nifuji A and Noda M: Establishment of
tendon-derived cell lines exhibiting pluripotent mesenchymal stem
cell-like property. Exp Cell Res. 287:289–300. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Blander G and Guarente L: The Sir2 family
of protein deacetylases. Annu Rev Biochem. 73:417–435. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Cohen-Kfir E, Artsi H, Levin A, Abramowitz
E, Bajayo A, Gurt I, Zhong L, D'Urso A, Toiber D, Mostoslavsky R
and Dresner-Pollak R: Sirt1 is a regulator of bone mass and a
repressor of Sost encoding for sclerostin, a bone formation
inhibitor. Endocrinology. 152:4514–4524. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Backesjo CM, Li Y, Lindgren U and Haldosén
LA: Activation of Sirt1 decreases adipocyte formation during
osteoblast differentiation of mesenchymal stem cells. Cells Tissues
Organs. 189:93–97. 2009. View Article : Google Scholar
|
|
12
|
Liu J, Chen L, Tao X and Tang K:
Phosphoinositide 3-kinase/Akt signaling is essential for
prostaglandin E2-induced osteogenic differentiation of rat tendon
stem cells. Biochem Biophys Res Commun. 435:514–519. 2013.
View Article : Google Scholar
|
|
13
|
Liu J, Chen L, Zhou Y, Liu X and Tang K:
Insulin-like growth factor-1 and bone morphogenetic protein-2
jointly mediate prostaglandin e2-induced adipogenic differentiation
of rat tendon stem cells. PloS one. 9:e854692014. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Hata K, Nishimura R, Ikeda F, Yamashita K,
Matsubara T, Nokubi T and Yoneda T: Differential roles of Smad1 and
p38 kinase in regulation of peroxisome proliferator-activating
receptor gamma during bone morphogenetic protein 2-induced
adipogenesis. Mol Biol Cell. 14:545–555. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Ji C, Chang W, Centrella M and McCarthy
TL: Activation domains of CCAAT enhancer binding protein delta:
Regions required for native activity and prostaglandin E2-dependent
transactivation of insulin-like growth factor I gene expression in
rat osteoblasts. Mol Endocrinol. 17:1834–1843. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Simic P, Zainabadi K, Bell E, Sykes DB,
Saez B, Lotinun S, Baron R, Scadden D, Schipani E and Guarente L:
SIRT1 regulates differentiation of mesenchymal stem cells by
deacetylating β-catenin. EMBO Mol Med. 5:430–440. 2013. View Article : Google Scholar : PubMed/NCBI
|