Introduction
Glioma is a common type of brain cancer originating
from various types of glial cells residing in the brain, including
ependymal cells, astrocytes and oligodendrocytes. Glioblastoma
multiforme, derived from astrocytes and one of the three types of
glioma, is the most common form of malignant brain tumor (1–4). Due
to the fast-growing nature of astrocytes and abundant vascular
network within glioblastoma, this type of tumor is develops rapidly
and aggressively, and accounts for 15.4% of all primary brain
tumors and 60–75% of astrocytomas (1,2).
Currently, patients with glioblastoma are estimated to have a
median survival time of 14.6 months and a 30% two-year survival
rate. The primary therapies for glioblastoma are conventional
surgery and radiotherapy. However, glioblastoma cells remain
active, therefore tumors may recur and develop resistance to radio-
and chemotherapy. The median time for disease recurrence following
standard therapy is 6.9 months (2,5).
Thus, this type of tumor remains one of the most life-threatening;
identifying novel therapeutic targets and molecular markers is
essential.
Conventional tumor therapies have focused on
targeting the mass of homogenously mutated cells using chemo- and
radiotherapy. However, multiple combinations of these therapies
have yielded limited success; this has led to the development of
the cancer stem cell theory, which outlines a hierarchy in tumor
cells (6–9). Emerging evidence has indicated the
presence of cancer stem cell niches within the tumor, which
initiate tumor growth and metastasis, and survive chemo- or
radiotherapy, leading to disease recurrence. The first evidence for
glioblastoma stem cells (GSCs) was presented in 2003, when Singh
et al (6,7) isolated cluster of differentiation
(CD) 133(+) brain tumor-initiating cells from patients, and
verified the self-renewal and differentiation potential of this
subpopulation. Subsequently, further studies demonstrated that GSCs
possess markedly greater levels of chemoresistant genes, including
O-6-methylguanine-DNA methyltransferase (an enzyme responsible for
DNA repair) (10), breakpoint
cluster region pseudogene 1, and the anti-apoptosis genes B-cell
lymphoma 2, B-cell lymphoma-extra large and the inhibitors of
apoptosis proteins (3,11). Therefore, targeting these cell
types may provide novel strategies for the treatment of
glioblastoma, and the prevention of its recurrence.
Additionally, growing evidence has suggested that
micro (mi)RNAs, a large group of non-coding RNAs, regulate numerous
pathophysiological processes in humans (12–17).
miRNA (miR)-141 has been demonstrated to exhibit antitumor effects
in various types of tumors (12,14,16).
Chen et al (16) reported
that miR-141 may inhibit the proliferation and metastasis of kidney
cancer cells via ephrin type-A receptor 2, and Burk et al
(18) revealed that the
miR-141/200c family may form a feedback loop with zinc finger
E-box-binding homeobox 1, thereby regulating epithelial-mesenchymal
transition (EMT) and metastasis in cancer. However, whether miR-141
possesses antitumor effects in glioblastoma remains unknown. The
present study demonstrated that miR-141 is greatly suppressed in
the CD133(+) glioblastoma cell subpopulation compared with CD133(−)
cells. Thus, miR-141 may serve as an antioncomir in
glioblastoma.
Materials and methods
Ethical approval
The present study was approved by the Ethics
Committee of The First Hospital of Jilin University (Changchun,
China) for both the animal experiments and human tissue sample
processing. Written consent was obtained from patients donating
glioblastoma tissue. This investigation complied with the
principles that govern the use of human tissues outlined in the
Declaration of Helsinki (19).
Isolation of CD133(+) GSCs and CD133(−)
non-glioblastoma stem cells (NSCs). Glioblastoma tissue specimens
were obtained from patients from the Department of Neurosurgery of
The First Hospital of Jilin University.
Glioblastoma tissue was sectioned into pieces and
digested with 0.2% collagenase II (Sigma-Aldrich; Merck Millipore,
Darmstadt, Germany) in Dulbecco's modified Eagle's medium
(DMEM)-high glucose (Corning Life Sciences, Tewksbury, MA, USA) for
2 h at 37°C. The cell suspension was subsequently rinsed with
Dulbecco's PBS three times and incubated with Red Blood Cell Lysis
buffer (Sigma-Aldrich; Merck Millipore) to eliminate erythrocytes.
The remaining cells were incubated with an anti-human-CD133
conjugated with phycoerythrin antibody (cat. no. 130–098-826;
dilution, 1:11; Miltenyi Biotec GmbH, Bergisch Gladbach, Germany)
or a negative control antibody (cat. no. 130–098-845; dilution,
1:11; Miltenyi Biotec GmbH, Bergisch Gladbach, Germany) at 4°C for
30 min, rinsed with PBS three times and resuspended in buffer.
Following this, CD133(+) and CD133(−) cells were separately
isolated via Fluorescence Activated Cell Sorting (FACS). CD133(+)
GSCs were maintained in minimal glucose lacking histidine medium,
which was composed of: DMEM/nutrient mixture F12 (HyClone; GE
Healthcare Life Sciences, Logan, UT, USA), 1X N2 supplement (Thermo
Fisher Scientific, Inc., Waltham, MA, USA), 50 µg/ml bovine serum
albumin (cat. no. 9048-46-8; Beijing Solarbio Science &
Technology Co., Ltd., Beijing, China), 25 µg/ml gentamycin (cat.
no. G8170; Beijing Solarbio Science & Technology Co., Ltd.),
100 U/ml penicillin/streptomycin (Wuhan Boster Biological
Technology, Wuhan, China), 20 ng/ml epidermal growth factor (EGF;
PeproTech, Inc., Rocky Hill, NJ, USA) and 20 ng/ml basic fibroblast
growth factor (bFGF; PeproTech, Inc.). DMEM/F-12 medium (HyClone;
GE Healthcare Life Sciences) supplemented with 20 ng/ml EGF and 20
ng/ml bFGF was prepared for the sphere formation assay.
Sphere formation assay
To evaluate the capability of cancer stem cells to
form spheres, cells were trypsinized, pipetted to a single cell
suspension and adjusted to a concentration of 500 cells/ml. The
single cell suspension (2 µl/well) was seeded into ultra-low
attachment 96-well plates (Corning Incorporated, Corning, NY, USA)
supplemented with 150 µl/well sphere formation medium, of which an
extra 20 µl was added every three days. After two weeks, the total
number of spheres formed by each cell type was counted by
microscope (1X71 Research Inverted System microscope; Olympus
Corporation, Tokyo, Japan) and evaluated as the number of
spheres/96 cells.
Subcutaneous tumor xenograft
assay
A total of 12 male BALB/c nude mice (5-weeks-old)
were obtained from Beijing HFK Bioscience Co., Ltd. (Beijing,
China) and kept in a specific pathogen-free vivarium with
temperature between 26–34°C, humidity at 30–50%, 12/12 h light/dark
cycle and free access to food and water (n=6/group). For the in
vivo tumorigenicity assessment of the GSC-vector and
GSC-miR-141 cell lines, ~1.0×106 GSCs were cultured,
rinsed with precooled PBS three times, and resuspended with
precooled Matrigel (BD Biosciences, San Jose, CA, USA) diluted with
PBS at a ratio of 1:3. This was subsequently injected into either
the right or left axillary fossa of each mouse. From day 7, mouse
tumor volumes were measured with calipers by two experienced lab
members at different time points throughout the day, every 3 days.
The formula to calculate tumor volume was volume=0.5ab2,
where ‘a’ is the long axis and ‘b’ is the short axis of the tumor.
All animals were sacrificed using general anesthesia by 1.5%
isoflurane (Sigma-Aldrich; Merck Millipore) inhalation and tumors
(maximum size, 4.7 mm3) were weighed.
Antibodies and reagents
A Jagged1 rabbit anti-human polyclonal antibody was
purchased from Abcam (cat. no. ab7771; Cambridge, MA, USA) and a
GAPDH rabbit anti-human polyclonal antibody (cat. no. A00227) was
purchased from Wuhan Boster Biological Technology. Horseradish
peroxidase (HRP)-labeled goat anti-mouse IgG (cat. no. G1210-2-A)
and goat anti-rabbit secondary antibodies (cat. no. G1210-2-B) were
obtained from Guge Biotechnology Co., Ltd. (Wuhan, China), and
enhanced chemiluminescence (ECL) reagents were purchased from EMD
Millipore (Billerica, MA, USA). A quantitative polymerase chain
reaction (qPCR) kit was purchased from Takara Bio, Inc. (Otsu,
Japan).
Western blot analysis
Whole cell proteins were extracted using
radioimmunoprecipitation assay buffer supplemented with an
inhibitor cocktail (Roche Diagnostics, Indianapolis, IN, USA), and
subsequently separated by 10% SDS-PAGE with 40 µg loading samples.
Following this, proteins were transferred onto polyvinylidene
difluoride membranes and blocked with 5% skimmed milk in TBS
containing Tween 20 (TBST) for 2 h at room temperature. The
membranes were incubated with anti-Jagged1 (1:1,000) at 4°C
overnight, and rinsed with TBST three times. The membranes were
subsequently incubated with HRP-conjugated secondary antibodies
(1:10,000) for 2 h at 37°C. Following the addition of ECL reagents,
the protein bands were observed and analyzed using the Bio-Rad
imaging system (version, 5.2.1; Bio-Rad Laboratories, Inc.,
Hercules, CA, USA). GAPDH (1:5,000) served as an internal
control.
Plasmids and primers
The hsa-miR-141-3p and Jagged1 overexpression
plasmids were modified from a psicoR vector and a vector containing
a puromycin resistance gene, respectively (Addgene, Inc.,
Cambridge, MA, USA). Dual luciferase plasmids of Jagged1 and its
corresponding mutant were modified from a psiCHECK2 vector (Promega
Corporation, Madison, WI, USA) and dual luciferase assay was
undertaken as instructed by the protocol provided by Promega
Corporation (20). All primers
used are listed in Table I. The
primers for qPCR analysis of miR-141 were purchased from
GeneCopoeia, Inc. (Rockville, MD, USA). Reverse transcription
reagents were purchased and undertaken as the protocol from
GeneCopoeia, Inc. (cat. no. QP018), while PCR reagents were also
purchased from GeneCopoeia Inc. (cat. no. QP015).
 | Table I.Quantitative polymerase chain reaction
primers. |
Table I.
Quantitative polymerase chain reaction
primers.
| Gene | Sequence (5′-3′) |
|---|
| CD133 | F:
GGCCCAGTACAACACTACCAA |
|
| R:
ATTCCGCCTCCTAGCACTGAA |
| Oct-4 | F:
CTGGGTTGATCCTCGGACCT |
|
| R:
CCATCGGAGTTGCTCTCCA |
| Nestin | F:
CAACAGCGACGGAGGTCTC |
|
| R:
GCCTCTACGCTCTCTTCTTTGA |
| C-Myc | F:
TCCCTCCACTCGGAAGGAC |
|
| R:
CTGGTGCATTTTCGGTTGTTG |
| Nanog | F:
AAGGTCCCGGTCAAGAAACAG |
|
| R:
CTTCTGCGTCACACCATTGC |
| Jagged1 | F:
GCCGAGGTCCTATACGTTGC |
|
| R:
CCGAGTGAGAAGCCTTTTCAA |
| TWIST-1 | F:
GTCCGCAGTCTTACGAGGAG |
|
| R:
GCTTGAGGGTCTGAATCTTGCT |
| Vimentin | F:
AGTCCACTGAGTACCGGAGAC |
|
| R:
CATTTCACGCATCTGGCGTTC |
| ZEB-1 | F:
TTACACCTTTGCATACAGAACCC |
|
| R:
TTTACGATTACACCCAGACTGC |
| ZEB-2 | F:
GGAGACGAGTCCAGCTAGTGT |
|
| R:
CCACTCCACCCTCCCTTATTTC |
| N-cadherin | F:
AGCCAACCTTAACTGAGGAGT |
|
| R:
GGCAAGTTGATTGGAGGGATG |
| E-cadherin | F:
ATTTTTCCCTCGACACCCGAT |
|
| R:
TCCCAGGCGTAGACCAAGA |
qPCR
A total of 106 cells of each group were
harvested by TRIzol reagent (cat. no. 15596026, Thermo Fisher
Scientific, Inc.) and total RNA was extracted by traditional
phenol-chloroform method. The reverse transcription was undertaken
according to the protocol form GeneCopoeia, Inc. (cat. no. QP018).
A total of 1 µg RNA for each group was mixed with the following 25
µl reaction mixture: 1 µg total RNA, 1 µl 2.5 U/µl Poly A
Polymerase, 1 µl RTase Mix, 5 µl 5X PAP/RT buffer and
RNase/DNase-free water to make the final 25 µl in total; cat. no.
QP018, GeneCopoeia, Inc.) and was incubated as 37°C of 60 min
followed with enzyme inactivation by inbucation at 85°C for 5 min.
Then, the reverse transcribed cDNA was mixed with other qPCR
reagents in a kit (cat. no. QP015; GeneCopoeia, Inc.) and was
analyzed using the Mx3000P qPCR system (cat. no. 401512, Agilent
Technologies, Santa Clara, USA) with the following thermocycling
conditions: 95°C for 10 min (1 cycle), 95°C for 10 sec followed
with 60°C for 20 sec and 72°C for 10 sec (40 cycles). Data
processing and normalization was undertaken as reported by Livak
and Schmittgen (21).
Transfection
The lentivirus of has-miR-141-3p and Jagged1 from
respective plasmids or empty plasmids were packaged by Hanbio Co.,
Ltd. (Shanghai, China). GSCs were transfected with the virus and
selected with puromycin at 1 µg/ml for 1 week for subsequent in
vitro and in vivo studies.
Statistical analysis
Statistical analyses were performed using SPSS
software version 20.0 (IBM SPSS, Armonk, NY, USA). A one-way
analysis of variance and Fisher's exact test were used to determine
significant differences between groups. Data are presented as the
mean ± standard error unless otherwise indicated. P<0.05 was
considered to indicate a statistically significant difference.
Results
CD133(+) GSCs are self-renewing
In order to verify the FACS sorting result of
CD133(+) GSCs and CD133(−) NSCs, qPCR (Fig. 1A; P<0.01) and western blot
(Fig. 1B; P<0.01) analyses were
performed to analyze the comparative expression levels of CD133 in
these two cell subpopulations. The sorted CD133(+) cells
demonstrated markedly increased expression levels of CD133,
compared with CD133(−) cells. In addition, qPCR was performed on
five stemness-associated [octamer-binding transcription factor 4
(Oct-4), nestin, c-Myc, homeobox protein Nanog (Nanog) and Jagged1]
and six EMT-associated [twist family BHLH transcription factor 1
(TWIST-1), vimentin, zinc finger E-box binding homeobox (ZEB)-1,
ZEB-2, neural (N)-cadherin and epithelial (E)-cadherin] genes to
determine differences in mRNA expression levels between the two
cell subopulations. As presented in Fig. 1C, the CD133(+) GSC subpopulation
exhibited upregulated expression levels of all five stemness genes
(Oct-4, nestin, c-Myc, Nanog and Jagged; all P<0.01) and five
mesenchymal-associated genes (TWIST-1, P<0.01; vimentin,
P<0.01; ZEB-1, P<0.01; ZEB-2, P<0.05; and N-cadherin,
P<0.01) compared with NSCs, which had upregulated E-cadherin
expression levels compared with GSCs. A sphere formation assay was
performed to assess the self-renewal of cells. It revealed that
CD133(+) GSCs exhibited an increased potential to duplicate
themselves compared with CD133(−) NSCs (Fig. 1D; P<0.01).
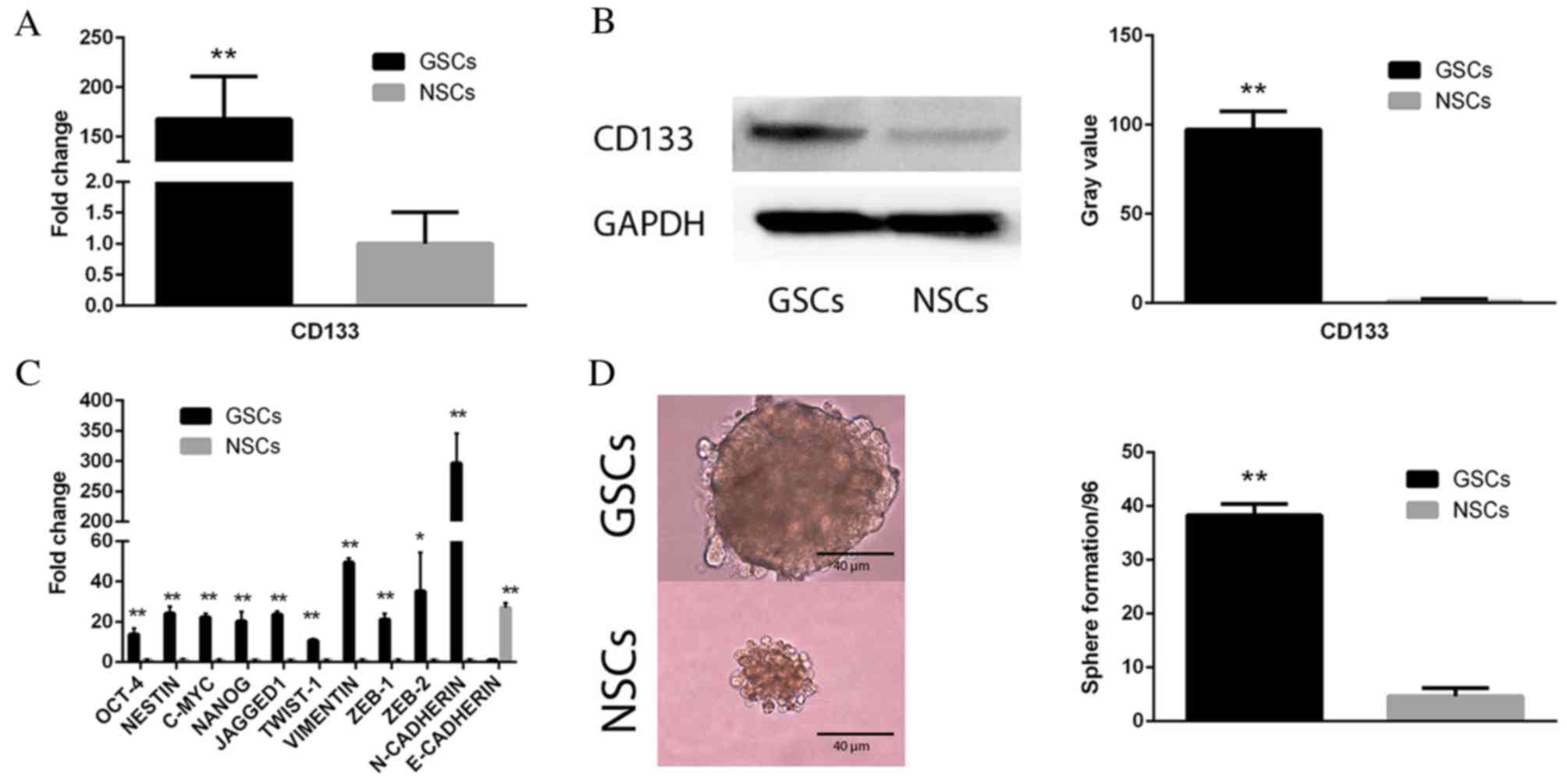 | Figure 1.CD133 expression and self-renewal
following Fluorescence Activated Cell Sorting of GSCs. (A) qPCR and
(B) western blot analysis demonstrating markedly increased
expression levels of CD133 in CD133(+) GSCs. Gray value indicates
protein optical density as analyzed by ImageJ software. GAPDH
served as an internal control. (C) qPCR analysis revealed an array
of stemness- and epithelial-mesenchymal transition-associated genes
were upregulated in CD133(+) GSCs compared with CD133(−) NSCs. (D)
A sphere formation assay demonstrated increased sphere formation in
CD133(+) GSCs compared with CD133(−) NSCs. Data are presented as
the mean ± standard error. *P<0.05 and **P<0.01 vs. NSCs.
GSCs, glioblastoma stem cells; NSCs, non-glioblastoma stem cells;
Oct-4, octamer-binding transcription factor 4; Nanog, homeobox
protein Nanog; TWIST-1, twist family BHLH transcription factor 1;
ZEB, zinc finger E-box binding homeobox; N, neural; E, epithelial;
qPCR, quantitative polymerase chain reaction; CD133, cluster of
differentiation 133. |
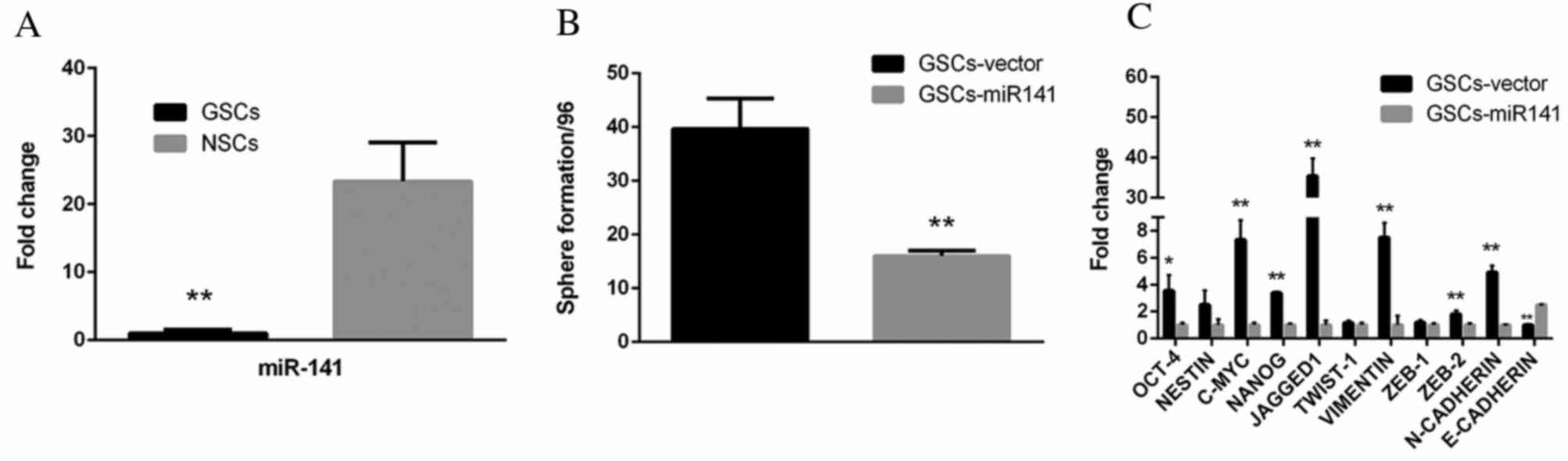
miR-141 expression levels are
suppressed in GSCs and inhibit the self-renewal of GSCs in
vitro
To investigate the role of miR-141 in GSCs, qPCR
analysis was performed. Compared with NSCs, GSCs exhibited markedly
reduced expression levels of miR-141 (Fig. 2A; P<0.01). GSCs transfected with
miR-141 exhibited significantly reduced sphere formation ability,
compared with vector-transfected cells (Fig. 2B; P<0.01). In addition, a set of
stemness (Oct-4, P<0.05; nestin, P>0.05; c-Myc, P<0.01;
Nanog, P<0.01; Jagged, P<0.01; Twist-1, P>0.05) and
EMT-associated genes (vimentin, P<0.01; Zeb-1, P>0.05; Zeb-2,
P<0.01; N-cadherin, P<0.01; E-cadherin, P<0.01) were
downregulated in GSCs-miR-141 compared with GSCs-vector, whereas
E-cadherin was upregulated (Fig.
2C).
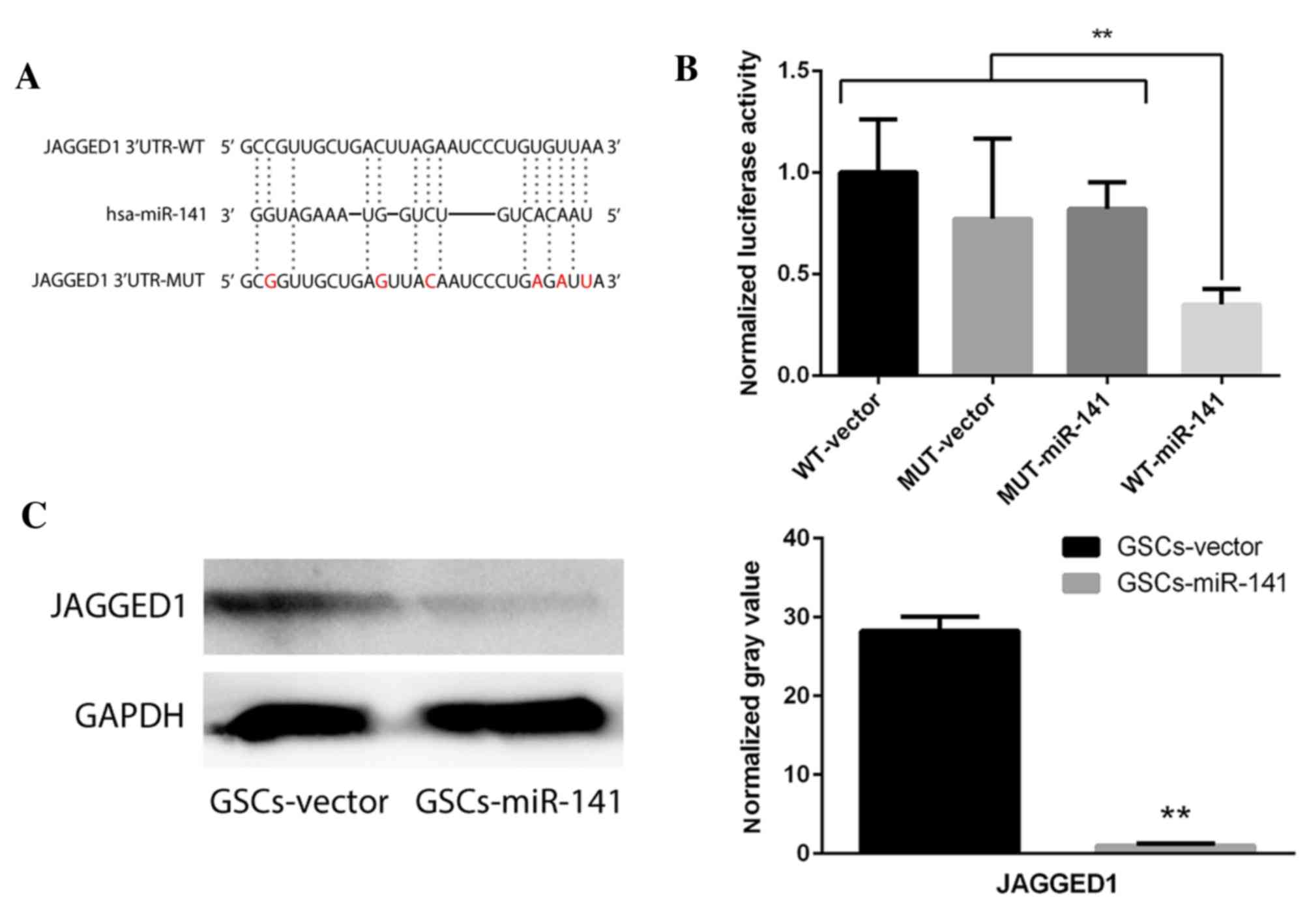
Jagged1 may be inhibited via direct
binding with miR-141
To investigate the potential targets of miR-141, two
dual luciferase constructs were made containing a Jagged1
3′-untranslated region and a mutated version of this region
(Fig. 3A). Following transfection
with miR-141, luminescence significantly decreased in the wild-type
compared with the mutant group (Fig.
3B; P<0.01). Western blot analysis revealed that Jagged1
expression levels were significantly decreased following
overexpression of miR-141 in GSCs (Fig. 3C; P<0.01). To verify the effect
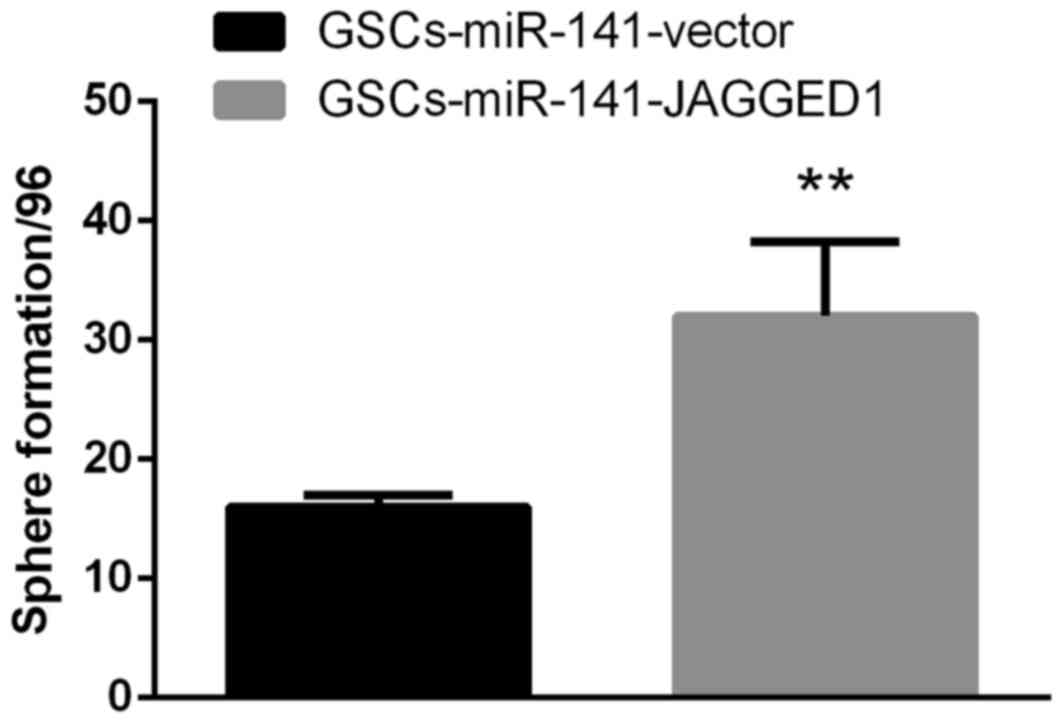
of Jagged1 on self-renewal, Jagged1 overexpression constructs were
cloned to upregulate its expression levels in GSCs-miR-141. The
results demonstrated that the reduced sphere formation ability in
GSCs-miR-141 was recovered (Fig.
4; P<0.01). These results were compared to a
GSC-vector-vector group, and this demonstrated a similar sphere
formation ability, whereby it was only partially recovered
following Jagged1 overexpression (data not shown).
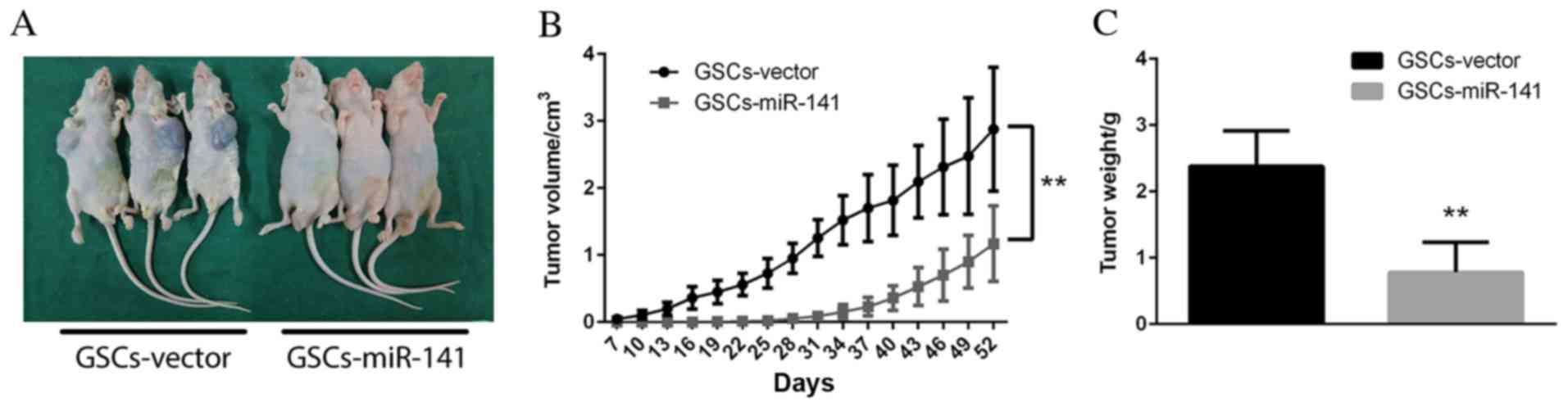
miR-141 may inhibit the tumorigenecity
of GSCs in vivo
To investigate the role of miR-141 in GSCs in
vivo, two groups of mice were transfected with GSCs-vector or
GSCs-miR-141. As presented in Fig.
5A, the tumors produced by GSCs-miR-141 were markedly reduced
in size compared with GSCs-vector. Furthermore, significantly
reduced tumor volumes (Fig. 5B;
P<0.01) and weight (Fig. 5C)
were observed in mice implanted with miR-141 overexpressing GSCs,
compared with those implanted with vector-transfected cells,
suggesting that miR-141 expression may inhibit tumor growth in
vivo (Fig. 5B; P<0.01).
Discussion
The present study demonstrated a direct interaction
between Jagged1 and miR-141 in GSCs. Clement et al (20) reported in 2011 that miR-141 may
inhibit Jagged1, thereby inhibiting the metastasis of prostate
cancer cells; however, the effect of miR-141 on the self-renewal of
cancer stem cells, particularly in glioblastoma, remains
unclear.
Jagged1 is a protein mutated in 60–75% of patients
with Alagille Syndrome (22). It
is encoded by the JAG1 gene, and the translation product is the
ligand for the receptor Notch-1/3 (22). A previous study by Chen et
al (23) revealed that Jagged1
promoted the pathogenesis of ovarian cancer by activating Notch-3.
Furthermore, the canonical stemness-associated Wnt/β-catenin
signaling pathway may upregulate the expression levels of Jagged1,
thus facilitating the development of ovarian cancer. Notably,
Jagged1 expression has been identified to be negatively correlated
with outcome in glioblastoma patients (24). Qiu et al (24) reported that in 82 glioblastoma
patients, reduced expression levels of Jagged1 were consistent with
slower tumor progression and increased overall survival.
Multivariate survival analysis additionally revealed that increased
Jagged1 expression levels in tumor or endothelial cells are an
independent prediction factor for poor prognosis. Consistent with
these findings, the present study demonstrated that miR-141
overexpression results in decreased Jagged1 expression levels and
reduced self-renewal in GSCs. These results may facilitate the
development of therapeutic targets to prolong the overall survival
rate of patients with glioblastoma.
Furthermore, Jagged1 is closely associated with the
Notch signaling pathway. The Notch signaling pathway is involved in
numerous physiological and pathophysiological processes, including
differentiation and stemness maintenance. There are primarily four
types of Notch family receptors: Notch-1, -2, -3 and -4 (12,23,25–27),
which may promote proliferative signaling during neuron
development. The Notch receptor ligands in mammals primarily
include delta-like proteins and jagged proteins (23). There has been increasing evidence
to suggest Notch's involvement in cancer cell growth; Farnie et
al (26) demonstrated that a
Notch-4-neutralizing antibody may markedly reduce the self-renewal
of breast cancer stem cells. Furthermore, Fan et al
(27) identified a feedback loop
between human epidermal growth factor receptor 2/proto-oncogene Neu
and Notch in breast cancer stem cells. In glioblastoma, Fan et
al (27) demonstrated that the
Notch signaling pathway blockade by gamma-secretase inhibitors may
deplete CD133(+) glioblastoma cells. The results of the present
study suggested that the signal attenuation of the Notch signaling
pathway via inhibition of Jagged1 by miR-141 may serve as a
potential strategy to eradicate glioblastoma cells within
tumors.
EMT has been associated with cancer stem cell
maintenance and development (28).
Previous studies have demonstrated that the activation of EMT
confers stemness traits to healthy and neoplastic cells, thus EMT-
and stemness-associated genes may be upregulated in parallel or
sequentially (12,26,28).
The present study revealed that the EMT-associated genes TWIST-1,
vimentin, ZEB-1, ZEB-2 and N-cadherin, were co-upregulated with the
stemness genes Oct-4, nestin, C-Myc, Nanog and Jagged1, in GSCs
compared with NSCs. miR-141-overexpressing GSCs exhibited
downregulated EMT- and stemness-associated gene expression, and
upregulation of E-cadherin. Therefore, miR-141 may additionally
serve a role in EMT in glioblastoma and reduce tumor aggression,
leading to tumor demarcation that may facilitate surgical
resection.
In conclusion, the present study demonstrated that
miR-141 may serve as an antioncomir in GSCs and markedly inhibit
their self-renewal via downregulating Jagged1 expression levels
in vitro and in vivo. Similar to commercially
available small interfering RNA treatment agents, it is
hypothesized that miR-141 may be utilized for glioblastoma
treatment. Targeting GSCs or the source of glioblastoma remains a
promising strategy to impede metastasis and resistance to
chemotherapy or radiotherapy. The potential of miR-141 to inhibit
cell metastasis requires further investigation, to facilitate the
development of novel treatment strategies for glioblastoma.
Acknowledgements
The authors thank all the patients who participated
in this program and all coworkers who contributed to this
study.
References
|
1
|
Luo JW, Wang X, Yang Y and Mao Q: Role of
micro-RNA (miRNA) in pathogenesis of glioblastoma. Eur Rev Med
Pharmacol Sci. 19:1630–1639. 2015.PubMed/NCBI
|
|
2
|
Rachet B, Mitry E, Quinn MJ, Cooper N and
Coleman MP: Survival from brain tumours in England and Wales up to
2001. Br J Cancer. 99:(Suppl 1). S98–S101. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Cho DY, Lin SZ, Yang WK, Lee HC, Hsu DM,
Lin HL, Chen CC, Liu CL, Lee WY and Ho LH: Targeting cancer stem
cells for treatment of glioblastoma multiforme. Cell Transplant.
22:731–739. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Bao S, Wu Q, Sathornsumetee S, Hao Y, Li
Z, Hjelmeland AB, Shi Q, McLendon RE, Bigner DD and Rich JN: Stem
cell-like glioma cells promote tumor angiogenesis through vascular
endothelial growth factor. Cancer Res. 66:7843–7848. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Stupp R, Mason WP, van den Bent MJ, Weller
M, Fisher B, Taphoorn MJ, Belanger K, Brandes AA, Marosi C, Bogdahn
U, et al: Radiotherapy plus concomitant and adjuvant temozolomide
for glioblastoma. N Engl J Med. 352:987–996. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Singh SK, Clarke ID, Terasaki M, Bonn VE,
Hawkins C, Squire J and Dirks PB: Identification of a cancer stem
cell in human brain tumors. Cancer Res. 63:5821–5828.
2003.PubMed/NCBI
|
|
7
|
Singh SK, Hawkins C, Clarke ID, Squire JA,
Bayani J, Hide T, Henkelman RM, Cusimano MD and Dirks PB:
Identification of human brain tumour initiating cells. Nature.
432:396–401. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Furnari FB, Fenton T, Bachoo RM, Mukasa A,
Stommel JM, Stegh A, Hahn WC, Ligon KL, Louis DN, Brennan C, et al:
Malignant astrocytic glioma: Genetics, biology, and paths to
treatment. Genes Dev. 21:2683–2710. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Chen J, Li Y, Yu TS, McKay RM, Burns DK,
Kernie SG and Parada LF: A restricted cell population propagates
glioblastoma growth after chemotherapy. Nature. 488:522–526. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Ji J, Black KL and Yu JS: Glioma stem cell
research for the development of immunotherapy. Neurosurg Clin N Am.
21:159–166. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Liu G, Yuan X, Zeng Z, Tunici P, Ng H,
Abdulkadir IR, Lu L, Irvin D, Black KL and Yu JS: Analysis of gene
expression and chemoresistance of CD133+ cancer stem
cells in glioblastoma. Mol Cancer. 5:672006. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Wang Z, Li Y, Kong D, Ahmad A, Banerjee S
and Sarkar FH: Cross-talk between miRNA and Notch signaling
pathways in tumor development and progression. Cancer Lett.
292:141–148. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Bier A, Giladi N, Kronfeld N, Lee HK,
Cazacu S, Finniss S, Xiang C, Poisson L, deCarvalho AC, Slavin S,
et al: MicroRNA-137 is downregulated in glioblastoma and inhibits
the stemness of glioma stem cells by targeting RTVP-1. Oncotarget.
4:665–676. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Wong JW: MicroRNA-induced silencing of
glioma progression. J Neurosci. 30:3868–3869. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Wu N, Xiao L, Zhao X, Zhao J, Wang J, Wang
F, Cao S and Lin X: miR-125b regulates the proliferation of
glioblastoma stem cells by targeting E2F2. FEBS Lett.
586:3831–3839. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Chen X, Wang X, Ruan A, Han W, Zhao Y, Lu
X, Xiao P, Shi H, Wang R, Chen L, et al: miR-141 is A key regulator
of renal cell carcinoma proliferation and metastasis by controlling
EphA2 expression. Clin Cancer Res. 20:2617–2630. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Guessous F, Alvarado-Velez M,
Marcinkiewicz L, Zhang Y, Kim J, Heister S, Kefas B, Godlewski J,
Schiff D, Purow B and Abounader R: Oncogenic effects of miR-10b in
glioblastoma stem cells. J Neurooncol. 112:153–163. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Burk U, Schubert J, Wellner U, et al: A
reciprocal repression between ZEB1 and members of the miR-200
family promotes EMT and invasion in cancer cells. EMBO reports.
9:582–589. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
World Medical Association Declaration of
Helsinki: Ethical principles for medical research involving human
subjects. JAMA. 310:2191–2194. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Clement T, Salone V and Rederstorff M:
Dual luciferase gene reporter assays to study miRNA function.
Methods Mol Biol. 1296:187–198. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta Delta C(T)) Method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Spinner NB, Colliton RP, Crosnier C,
Krantz ID, Hadchouel M and Meunier-Rotival M: Jagged1 mutations in
alagille syndrome. Hum Mut. 17:18–33. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Chen X, Stoeck A, Lee SJ, Shih Ie M, Wang
MM and Wang TL: Jagged1 expression regulated by Notch3 and
Wnt/b-catenin signaling pathways in ovarian cancer. Oncotarget.
1:210–218. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Qiu XX, Wang CH, You N, Chen BJ, Wang XF,
Chen YP and Lin ZX: High Jagged1 expression is associated with poor
outcome in primary glioblastoma. Med Oncol. 32:3412015. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Vallejo DM, Caparros E and Dominguez M:
Targeting Notch signalling by the conserved miR-8/200 microRNA
family in development and cancer cells. EMBO J. 30:756–769. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Farnie G, Clarke RB, Spence K, Pinnock N,
Brennan K, Anderson NG and Bundred NJ: Novel cell culture technique
for primary ductal carcinoma in situ: Role of Notch and epidermal
growth factor receptor signaling pathways. J Natl Cancer Inst.
99:616–627. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Fan X, Khaki L, Zhu TS, Soules ME, Talsma
CE, Gul N, Koh C, Zhang J, Li YM, Maciaczyk J, et al: NOTCH pathway
blockade depletes CD133-positive glioblastoma cells and inhibits
growth of tumor neurospheres and xenografts. Stem Cells. 28:5–16.
2010.PubMed/NCBI
|
|
28
|
Scheel C and Weinberg RA: Cancer stem
cells and epithelial-mesenchymal transition: Concepts and molecular
links. Semin Cancer Biol. 22:396–403. 2012. View Article : Google Scholar : PubMed/NCBI
|